Introduction
Circular RNAs (circRNAs) differ from other types of
RNA due to their covalently closed-loop structures, and as such,
have received increasing research interest since their discovery in
1976 (1). As members of the
non-coding RNA family, circRNAs have been considered as secondary
byproducts of linear mRNA splicing events (2). However, recent advancements in
experimental and next-generation RNA sequencing methods have helped
to identity and elucidate the functions of various novel circRNAs
(3). CircRNAs have been revealed to
influence a number of disease processes, including those involved
in cardiovascular diseases, cancer and neurological disorders.
Increasing research has confirmed the aberrant expression of
numerous circRNAs in gynecological tumors, which influence the
occurrence and development of these malignancies through multiple
channels. This provides further evidence for the potential of
circRNAs as biomarkers for the diagnosis, treatment and prognosis
of gynecological cancers. The aim of the present review was to
summarize our knowledge of the biogenesis and function of circRNAs,
and to discuss the advantages and limitations of circRNA as
biomarkers for cervical, ovarian and endometrial cancer, with the
aim to identify novel biomarkers.
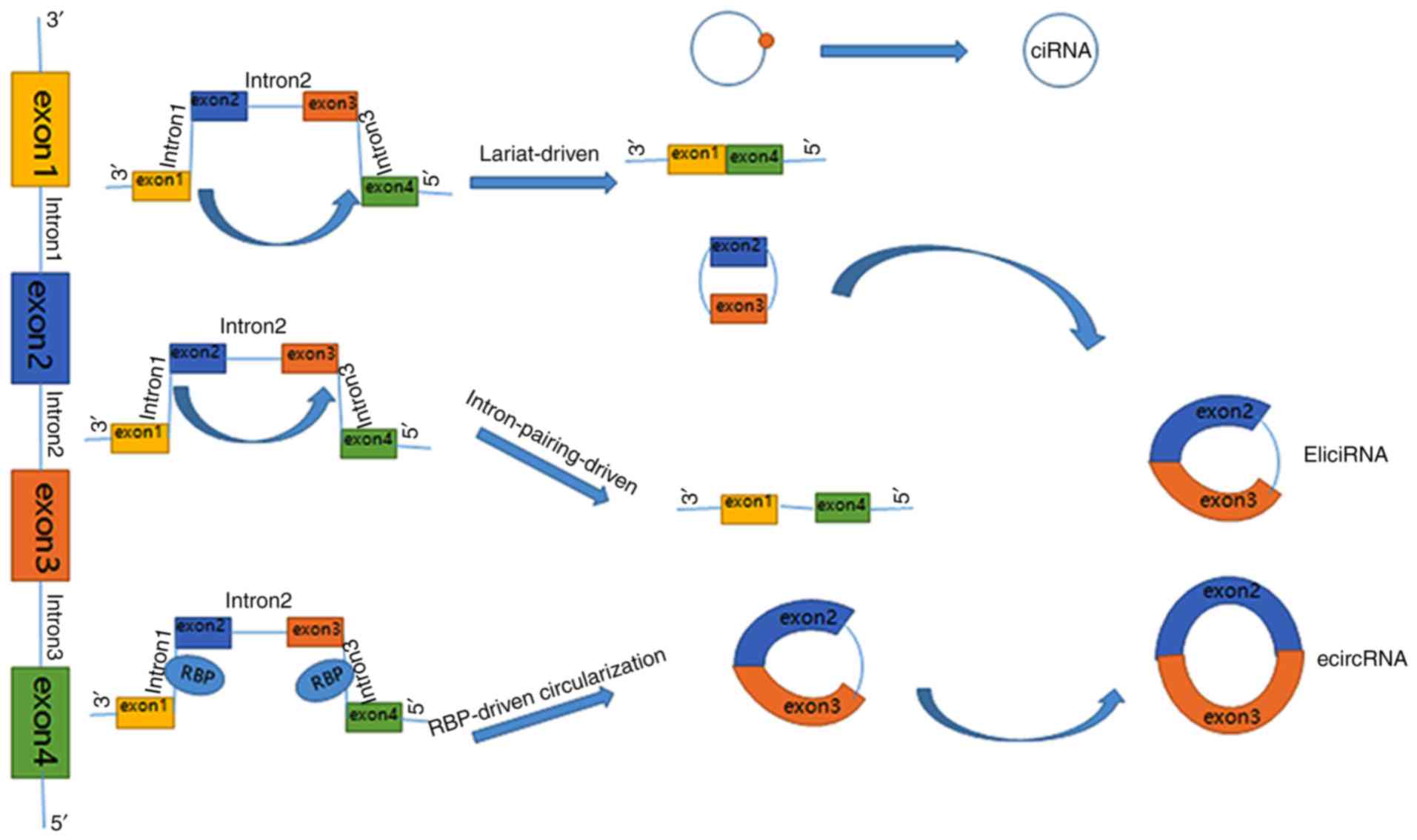
CircRNA biogenesis
As the name implies, circRNAs are closed circular
structures with covalently linked 3′ and 5′ ends, which distinguish
them from other types of RNA (1).
Most circRNAs are generated by pre-mRNA splicing, and their
circular structures make them relatively stable and resistant to
RNase digestion (4,5). Due to their various different sources,
circRNAs can be categorized into three types: i) Exonic circRNAs
(ecircRNAs), which are the product of back-spliced exons; ii)
intronic circRNAs (ciRNAs), the product of intronic sequences; and
iii) exon-intron circRNAs (EliciRNAs), the products of circularized
exons with introns retained between them (3). These circRNAs are distributed
differently within cells; ciRNAs and EliciRNAs are primarily
present in the nucleus, while ecircRNAs are largely located in the
cytoplasm (6). There are also three
primary models for the formation of circRNA loops, namely
lariat-driven, intron-pairing-driven and RNA-binding protein
(RBP)-driven circularization (Fig.
1). These models can be further divided into two mechanisms;
direct back-splicing and exon skipping (1). In the lariat-driven circularization
model (exon skipping), a splice site 30 nucleotides upstream of an
exon is ligated into a site 50 nucleotides downstream, leading to
exon-skipping and the formation of an RNA lariat consisting of
several exons and introns. The introns are then removed to generate
ecircRNAs (4). Additionally, exons
are circularized with introns that are retained between the exons,
resulting in the generation of EliciRNAs (2). For the intron-pairing- and RBP-driven
circularization models (direct back-splicing), reverse
complementary sequences such as Alu repeats (located in the
upstream and downstream introns), and certain trans-acting
activator RNA-binding proteins binding to each of the flanking
introns, bring the splice donor and acceptor sites close enough to
form a loop (7–9). As for circular intron RNA biogenesis,
the conserved motifs at both ends, including a 7-nt GU-rich element
near the 5′ splice site and an 11-nt C-rich element near the
branch-point site, are joined together (10) to prevent introns from forming looped
branches, and instead promote the formation of a loop structure.
The 3′end of the intron is then relocated to the branch point to
produce a stable circular structure.
CircRNAs are widely present in various human cell
types. Their expression is cell type-specific; cells with low
proliferative capacity, such as cardiomyocytes, have higher
expression levels compared with cells that proliferate more
readily, such as those in the liver (7). In addition, the high level of circRNAs
in some tissues is primarily the result of accumulation, which is
likely due to the high stability of these molecules. Due to a lack
of free ends, circRNAs are more stable and resistant to RNase R
compared with linear RNAs (11).
Owing to these aforementioned features, we hypothesized that
circRNAs may serve as novel biomarkers for the diagnosis and
treatment of essential hypertension (12), inflammatory bowel disease (13) and a variety of different cancer
types (14). For example, in
hepatocellular carcinoma, hsa_circ_0001649 is more sensitive and
specific than the known biomarker alpha fetoprotein (15). Another circRNA, hsa_circ_025016, has
proven to be a potential plasma biomarker for the prediction of
postoperative atrial fibrillation (16).
CircRNA functions
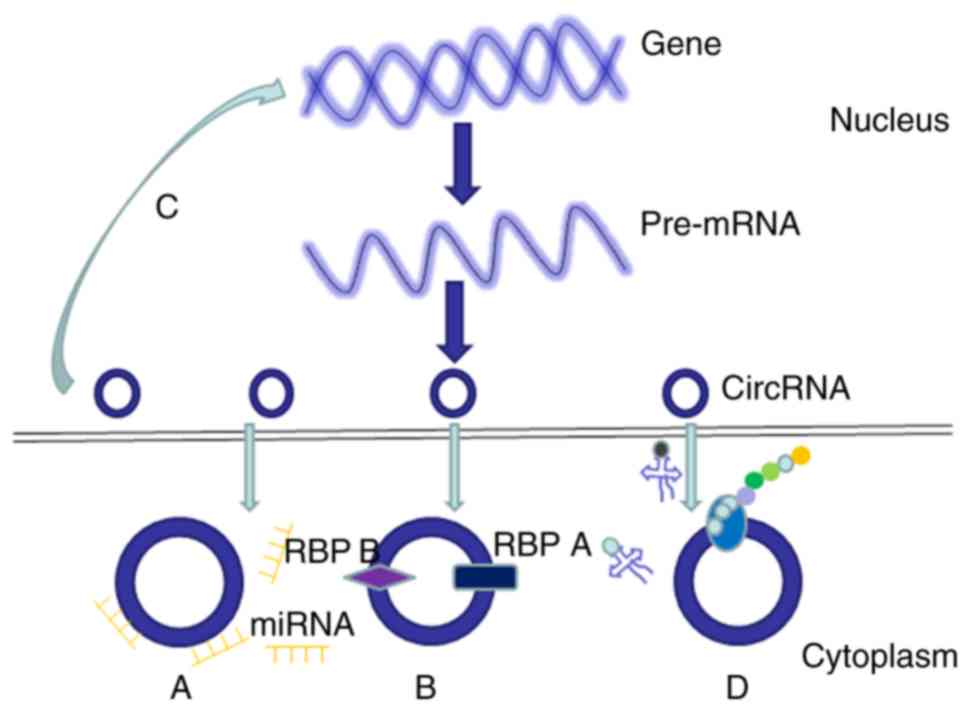
MicroRNA (miRNA/miR) sponging
An increasing number of scholars are focusing on the
roles of circRNAs in cancer, of which miRNA sponging is a major
mechanism (Fig. 2A). CDR1as/ciRS-7
is a well-known circRNA that acts as an miRNA sponge to inhibit
tumor growth in breast, colorectal, gastric and cervical cancer
(17–20). Another highly investigated, typical
competing endogenous RNA is circHIPK3, which is also involved in
antioncogenic processes. By sponging miR-558, circHIPK3 suppresses
the malignant properties of bladder cancer cells, including
migration, invasiveness and angiogenesis (21). Similarly, another study indicated
that circHIPK3 overexpression significantly suppressed the
proliferation, migration and invasiveness of osteosarcoma cells
in vitro (22). Furthermore,
circHIPK3 was found to promote the proliferation and progression of
gallbladder and lung cancer cells, potentially by sponging miR-124
(23,24). These findings suggest that the
properties of circRNAs are a double-edged sword.
Protein interactions
Another regulatory function of circRNAs is achieved
through their ability to interact with RNA-binding proteins
(Fig. 2B), the results of which
depend on the protein (and circRNA) in question. For example, p21
and cyclin-dependent kinase 2 strongly and specifically bind with
circFoxo3 to form a circFoxo3-Cdk2-p21 complex, which inhibits
transition from the G1 to the S phase of the cell cycle
(25). Additionally, Du et
al (26) revealed that
circFoxo3 could bind p53 and mouse double minute 2 homolog to
promote the ubiquitination and further degradation induced by p53.
circMbl is able to bind the mannose-binding lectin (MBL);
Ashwal-Fluss et al (27)
revealed that the biosynthesis of circMbl was affected by the
levels MBL via a feedback loop mechanism, and the process of MBL
pre-mRNA translation into circMbl was also affected by high levels
of MBL protein. Moreover, circMbl was found to bind MBL and
attenuate its bioavailability. Another study identified a number of
other circRNAs that bind multiple different proteins, serving
regulatory roles in cell functioning (28).
Gene expression regulation
The majority of the circRNAs that regulate gene
expression through sponge activity are localized in the cytoplasm
(Fig. 2C). However, recent research
has reported a small number of circRNAs that are localized in the
nucleus, which can regulate gene expression at the transcriptional
level. Li et al (29) found
that circEIF3J and circPAIP2 are localized to the nucleus and
promote the expression of parental genes, which involves the RNA
polymerase II, U1 small nuclear RNA and several promoter regions.
ciRNAs identified by Zhang et al (10) were also revealed to act as positive
regulators of RNA polymerase-II transcription, and to promote
parental gene expression. Similarly, ci-ankrd52 and ci-sirt7 were
revealed to interact with RNA polymerase II to modulate the
transcriptional rate of parental genes by accumulating at the
active transcription site (10).
Moreover, FECR1 circRNA, which binds to the promoter region and
recruits TET1 DNA demethylase, was revealed to regulate the
expression of the FLI1 gene, inducing DNA demethylation (30). These aforementioned circRNAs include
those of intronic, exonic and exon-intron origin.
Protein translation
Although circRNAs are members of the non-coding RNA
family, increasing data has revealed that circRNAs are also
involved in protein translation (Fig.
2D). Unlike linear RNAs, circRNAs lack 7-methylguanosine cap
structures and poly(A) tails, which prevents ribosomal recognition
and translation into protein. Since the discovery of the internal
ribosome entry site, the translation of circRNAs has become evident
(31). A recent study indicated
that circ-ZNF609, which regulates myoblast proliferation, contains
an open reading frame that can be translated into protein with in
cap-independent manner (32).
Circ-FBXW7 is abundantly expressed in the normal human brain, and
encodes a novel protein that regulates the proliferation and
cycling of cancer cells (33).
Furthermore, circ-SHPRH encodes SHPRH-146aa, which regulates the
proliferation and tumorigenicity of cancer cells in vitro
(34). However, to date, few
circRNAs have been found to be translated into protein, thus
further studies in this area are warranted.
CircRNAs as potential biomarkers of
gynecological tumors
A growing number of studies have focused on the
relationship between gynecological tumors and circRNAs. The present
review summarizes the most recent findings surrounding circRNA
regulation in cervical, ovarian and endometrial cancer, with the
aim to identify novel biomarkers and therapeutic approaches.
CircRNAs and cervical cancer (Table I)
Cervical cancer is one of the most common malignant
tumors in women, with an incidence rate second only to that of
breast cancer (35). It has a high
mortality rate and is the second major cause of tumor-related death
worldwide (36). Continuous
infection with high-risk human papillomavirus (HPV) is a major
pathogenic factor for cervical cancer progression (37). Although the administration of HPV
vaccines among patients with cervical cancer is increasing
globally, mortality and morbidity rates remain high in numerous
developed countries (38). In 2018,
~570,000 newly-diagnosed cases of cervical cancer, and ~311,000
deaths, were recorded worldwide (39). Surgery followed by radiotherapy and
chemotherapy is currently the primary method of cervical cancer
treatment. However, the poor prognosis and low survival rate caused
by distant metastasis and high lymphatic metastasis are still
problematic (40). Therefore, the
identification of novel treatments and biomarkers for earlier
diagnosis is necessary for increasing survival rates.
A previous study revealed that circRNAs are stable
and expressed to a high degree in various cancer cell lines
(41). Additionally, circRNA
detection methods are more sensitive and specific than those for
the detection of proteins, and circRNAs are more easily detectable
than miRNAs (42). For these
reasons, there is a high probability that circRNAs will make
suitable novel biomarkers for cancers. Currently, several studies
have investigated the roles of circRNAs in cervical cancer, of
which miRNA sponging may be the most significant mechanism. The
existing mechanisms (to the best of our knowledge) are discussed in
detail below.
Expression of circRNAs in cervical
cancer
Various studies have highlighted the presence of
multiple circRNAs in cervical cancer. Among 526 dysregulated
circRNAs, Zheng et al (43)
found that 352 were upregulated and 174 were downregulated,
indicating that circRNAs may be potential biomarkers for the
diagnosis and treatment of cervical cancer. Gao et al
(44) conducted a high-throughput
microarray study using four pairs of cervical cancer and adjacent
noncancerous tissues, revealing 19 downregulated and 26 upregulated
circRNAs in cervical cancer. Jiao et al (45) studied circRNA expression profiles
using microarray analysis, and found that 178 circRNAs were
differentially expressed between cervical cancer and matched normal
cervical tissues, of which 101 were downregulated and 77 were
upregulated. By classifying the 77 upregulated circRNAs, they also
identified 68 exonic, 3 intronic, 1 antisense and 5 intragenic
circRNAs in cervical cancer tissues, as well as 101 downregulated
circRNAs (including 81 exonic, 13 intronic, 3 antisense and 4
intragenic variants).
Although numerous studies aim to investigate the
expression of ring-shaped RNAs in cervical cancer, there are often
certain limitations. For example, the sample size of these studies
is small, making it especially difficult to understand the
expression profiles of different histological grades and
pathological stages. It is therefore necessary to perform
larger-scale sequencing studies to obtain more detailed data on the
expression of circRNAs in cervical cancer.
i) CircRNAs as biomarkers of cervical
cancer
The early diagnosis and treatment of cervical cancer
can effectively improve the survival rates of patients. With the
development of high-throughput sequencing and biochip technologies,
a growing number of circRNAs have exhibited potential as diagnostic
biomarkers.
ii) CircRNA expression in cervical
cancer tissues samples
Research by Wang et al (46) highlighted that the expression level
of hsa_circ_0001038 was higher in cervical cancer tissues than in
adjacent non-cancerous tissues. Moreover, Chen et al
(47) discovered that
circRNA_0000285 was frequently upregulated in cervical cancer
tissues compared with adjacent normal tissues.
iii) CircRNAs as biomarkers of
cervical cancer metastasis
The expression of circ_0067934 was found to be
increased in cervical cancer tissues and cell lines, compared with
that in adjacent normal tissues (48). High circ_0067934 expression was also
associated with positive lymphatic metastasis in patients with
cervical cancer, indicating its use as a potential biomarker of
cervical cancer metastasis. Ding et al (49) demonstrated that circ-ATP8A2 was
upregulated in cervical cancer tissues and cell lines, and further
confirmed that high levels of circ-ATP8A2 were associated with a
higher FIGO stage, positive lymph node invasion and myometrial
invasion. In addition, has-circ-u0000745 was upregulated in
cervical cancer tissues and cell lines, which was found to be
correlated with various clinicopathological features, such as
poorly differentiated tumors and positive vascular/lymphatic
invasion (45). Another circRNA,
circ_0005576, has been associated with a higher pathological stage
and lymph node metastasis (50).
However, alternative studies have indicated that reduced circRNA
expression also promotes metastasis; Jiao et al (51) revealed that low expression levels of
circRNA_101308 were associated with deep myometrial invasion and
lymph node metastasis. Therefore, selecting the appropriate circRNA
biomarker for a specific pathology requires more thorough
consideration.
CircRNAs as biomarkers of cervical
cancer prognosis
It is important to identify patients with a poor
prognosis in order to implement effective treatment. Therefore,
there is an urgent requirement for the discovery of novel
prognostic biomarkers for cervical cancer. CircRNAs are
differentially expressed in tumors of different histological grades
and pathological stages, and those with lymph node or distant
metastases, suggesting that circRNAs have potential prognostic
value as biomarkers of cervical cancer. For instance, Ji et
al (52) revealed that the
expression level of circSLC26A4 was upregulated in cervical cancer
tissues compared with adjacent normal tissues, and that high
circSLC26A4 expression was associated with a poor patient
prognosis. Circ_0018289 has also been revealed to be markedly
upregulated in cervical cancer tissues compared with adjacent
normal tissues (53), which was
significantly correlated with decreased overall survival,
suggesting that circ_0018289 may be a potential biomarker for
evaluating prognosis. Additionally, hsa_circ_0023404 was found to
be upregulated in cervical cancer tissues and cell lines (54). As a result of Kaplan-Meier analysis,
patients with high hsa_circ_0023404 expression levels exhibited
poorer overall survival rates than those with low hsa_circ_0023404
expression levels. The primary associated mechanism was that
hsa_circ_0023404 sponged miR-136, inducing yes-associated protein
(YAP) pathway activation by promoting α globin transcription factor
CP2 (TFCP2) expression, resulting in the development and
progression of cervical cancer. This study revealed that
hsa_circ_0023404 plays a pivotal role in the regulation of cervical
cancer progression through the hsa_circ_0023404-miR-136-TFCP2-YAP
axis. A number of other circRNAs, such as hsa_circ_0000515
(55), circ_0005576 (50) and circ-0000745 (45) have also been revealed to be highly
expressed in cervical cancer, and associated with poor patient
prognosis. The discovery of these circRNAs provides predictive
possibility for the prognosis of cervical cancer.
CircRNAs suppress tumor
progression
Cervical cancer results from the complex interaction
between various factors and signaling pathways. Currently, the
primary consensus is that this involves persistent infection with
high-risk HPV. Multiple circRNAs have been revealed to play
important roles in the progression of cervical cancer, and may
therefore help to improve our understanding of the mechanisms
underlying cancer progression.
CircSMARCA5 (hsa_circ_0001445), which is located on
chr4:144464661-144465125 and forms the circular exon 15 and 16 of
SMARCA5, is widely expressed in human cells. Previous studies have
found that circSMARCA5 can regulate cellular proliferation by
sponging miRNAs. For example, in prostate cancer, circSMARCA5
promoted tumor progression by sponging miR-432, but suppressed the
progression of multiple myeloma by targeting miR-767-5p (56,57).
In cervical cancer, circSMARCA5 acted as a tumor suppressor with
reduced expression in cancerous tissues, binding and inhibiting the
expression of miR-620. High expression levels of circSMARCA5
suppressed the proliferation, migration and invasiveness of cancer
cells by sponging miR-620 (58).
CircRNA_101308 expression was also revealed to be decreased in
cervical cancer tissues and cell lines, compared with that in
normal tissues. CircRNA_101308 can bind various different miRNAs
and regulate their downstream genes to suppress the proliferation,
invasiveness and metastasis of cervical cancer cells, both in
vitro and in vivo (51).
Furthermore, circ-ITCH has been revealed to be associated with the
regulation of tumor growth, and the proliferation and apoptosis of
cancer cells in bladder (59) and
breast cancer (60). In cervical
cancer, circ-ITCH expression was low and functioned as an miRNA
sponge for miR-93-5p. As the target of miR-93-5p, forkhead box
protein K2 (FOXK2) regulated multiple cancer cell features. In
cervical cancer, the overexpression of circ-ITCH suppressed the
proliferation and metastasis of cervical cancer cells by
interacting with the circ-ITCH/miR-93-5p/FOXK2 axis, and thus
upregulating the expression of FOXK2 (61).
CircRNAs promote tumor
progression
Previous research on circRNAs has improved our
understanding of cervical cancer pathogenesis. Ma et al
(50) recently discovered a novel
circ_0005576-miR-153-3p-kinesin family member 20A (KIF20A) axis
that regulates cervical cancer growth and invasion. They identified
a potentially new form of circ_0005576 that is primarily located in
the cytoplasm, and is significantly upregulated in cervical cancer.
Circ_0005576-knockdown was found to suppress cellular
proliferation, colony formation and metastasis, and circ_0005576
overexpression increased the expression of KIF20A by sponging
miR-153-3p, which may provide a new perspective into the
pathogenesis of cervical cancer. Moreover, hsa_circ_0000263 was
revealed to be upregulated in cervical cancer cells, and cellular
proliferation, migration and the cell cycle were inhibited by
knocking down hsa_circ_0000263. In a murine model, the expression
of murine double minute 4 (MDM4) was regulated by hsa_circ_0000263,
which ultimately affected the expression of the p53 gene.
Therefore, the hsa_circ_0000263/miR-150-5p/MDM4/p53 network has
been suggested to serve a vital role in the regulation of cervical
cancer (62). Furthermore, Hu et
al (48) revealed that
circ_0067934 was upregulated and associated with advanced stage,
lymph node metastasis, and poor prognosis, providing new insights
into the pathogenesis of cervical cancer. Tumor growth and cellular
proliferation were inhibited by knocking down circ_0067934 in
vitro and in vivo, and loss-of-function analysis
revealed that increased miR-545 expression suppressed eukaryotic
translation initiation factor 3 subunit C (EIF3C) expression by
silencing circ_0067934. This study revealed that circ_0067934
influences cellular proliferation, migration and invasiveness in
cervical cancer via the circ_0067934/miR-545/EIF3C axis.
CircRNAs as potential therapeutic
targets
CircRNAs may represent potential targets for the
treatment of cervical cancer. A previous study revealed that
hsa_circ_0000515 was upregulated in cervical cancer, and acted as a
ceRNA of miR-326 to increase the expression of ETS transcription
factor (ELK1). The in vitro results indicated that
proliferation and invasiveness, as well as induced apoptosis and
autophagy, were suppressed by silencing hsa_circ_0000515 or
overexpressing miR-326. ELK1 overexpression also suppressed
apoptosis and autophagy, but enhanced the proliferation and
invasiveness of cervical cancer cells. These findings indicate that
hsa_circ_0000515 may be a tumor promoter and provide evidence for
its therapeutic use in cervical cancer (55). Another circRNA, has-circ-0000745,
was revealed to be upregulated in patients with cervical cancer,
and high expression levels were associated with poorly
differentiated tumors or vascular/lymphatic invasion. Knocking down
hsa_circ_0000745 was found to upregulate E-cadherin expression,
inhibiting the proliferation, migration and invasiveness of
cervical cancer cells, indicating a potential target for cervical
cancer treatment (45). Ding et
al (49) revealed that
circ-ATP8A2, which is highly expressed in cervical cancer specimens
and cell lines, enhanced cellular proliferation, migration,
invasiveness and apoptosis in cervical cancer. The same effects
were observed for ectopically expressed circ-ATP8A2. The
experimental results indicated that circ-ATP8A2 exerted its
inhibitory effects on epidermal growth factor receptor (EGFR)
expression at the post-transcriptional level, by sponging miR-433.
These findings suggest a therapeutic effect of regulating the
circ-ATP8A2/miR-433/EGFR axis to mediate tumor progression.
Furthermore, the expression levels of circRNA_0000285 were higher
in cervical cancer samples than in adjacent, non-cancerous tissues,
and circRNA_0000285-knockdown in vitro significantly
inhibited the proliferation and migration abilities of cervical
cancer cells. circRNA_0000285 expression may also be correlated
with that of the RNA-binding protein FUS, which was found to be
regulated by circRNA_0000285. In a nude mouse model, the formation
and metastasis of cervical cancer were significantly inhibited by
circRNA_0000285-knockdown (47).
Collectively, these data provide insights into the pathogenic
mechanisms and potential treatment targets for cervical cancer.
Perspectives
At present, the association between cervical cancer
and circRNAs is receiving extensive research attention. Researchers
have identified numerous circRNAs that are aberrantly expressed in
cervical cancer, as well as the molecular mechanisms underlying
these phenomena. However, the mechanisms of a large number of
circRNAs are yet to be elucidated. Currently, cervical cancer and
normal adjacent tissues are the preferred specimen types for the
study of circRNAs, and there is a lack of more readily available
clinical specimen types, such as serum and urine, which would be
more convenient for cervical cancer diagnosis. Therefore,
additional research is required to improve our understanding of the
relationship between circRNAs and cervical cancer.
CircRNAs and ovarian cancer (Table II)
Ovarian cancer is a common, but life-threatening
gynecological malignancy that represents 3.6% of neoplasms in women
worldwide (63). Although it ranks
20th among the most common global cancer types, ovarian cancer has
the highest mortality rate of all gynecological malignancies, and
is the eighth leading cause of death among women worldwide
(63,64). In 2018, ovarian cancer resulted in
an estimated 295,414 new cases and 184,799 deaths worldwide
(39). Tumor debulking surgery
followed by platinum and paclitaxel chemotherapy is currently the
preferred clinical treatment (65).
However, the survival rate of patients with advanced ovarian cancer
is ~30%, which is primarily due to late diagnosis and
chemoresistance, the latter of which is influenced by the tumor
microenvironment and the inherent resistance of ovarian cancer
cells to chemotherapy (66). Thus,
improving the response to treatment and developing novel therapies
is crucial for improving survival rates.
Expression of circRNA in ovarian
cancer
Among 4,505 newly identified circRNAs, Teng et
al (67) highlighted 2,431 that
were significantly upregulated and 3,120 that were downregulated.
Furthermore, circHIPK3 was indicated to regulate the proliferation
and apoptosis of ovarian cancer and normal ovarian epithelial cells
via the circHIPK3-miRNA-mRNA axis. Another study revealed that
among 4,388 circRNAs with an expression fold change of ≥2, 2,556
were upregulated and 1,832 were downregulated. Further analysis
indicated that differentially expressed circRNAs may serve a
pathogenetic role in epithelial ovarian cancer, as well as acting
as potential diagnostic and prognostic biomarkers (68). Moreover, of 710 differentially
expressed circRNAs screened via high-throughput sequencing,
circRNA1656 exhibited the highest fold change, with low expression
in ovarian cancer tissues and cell lines, which was correlated with
FIGO stage in high-grade serous ovarian cancer (69).
CircRNAs as biomarkers for the
diagnosis, metastasis and prognosis of ovarian cancer
Early diagnosis prior to cancer metastasis
significantly improves prognosis, and the prognostic evaluation of
ovarian cancer also extends patient survival time (70). The pathogenesis of ovarian cancer is
complex, and current diagnostic methods, including the detection of
tumor markers, ultrasound, CT, MRI and histopathology, are not
sufficient. Therefore, novel indicators for diagnosis, metastasis
and prognosis are urgently required.
Researchers are now paying close attention to the
regulation of circRNAs in ovarian cancer, with the aim to identify
novel diagnosis biomarkers. Pei et al (71) discovered that hsa_circ_0013958 was
highly expressed in ovarian cancer tissues and cell lines, and that
hsa_circ_0013958 upregulation was closely associated with patient
FIGO stage and lymph node metastasis. Further analysis revealed
that hsa_circ_0013958 was a highly sensitive and specific indicator
for the diagnosis of ovarian cancer. Previous studies have revealed
that circLARP4 sponges miR-424 to regulate gastric cancer
progression. In ovarian cancer, the expression of circLARP4 was
found to be significantly downregulated, which was associated with
FIGO stage and lymph node metastasis. Survival analysis revealed
that a low level of circLARP4 was an independent risk factor for
ovarian cancer prognosis, suggesting its potential use as a
prognostic biomarker (72).
Furthermore, circRNA_MYLK was revealed to be highly expressed in
ovarian cancer compared with adjacent normal tissues, and high
circRNA_MYLK expression was positively associated with pathological
stage. Kaplan-Meier survival analysis indicated that high
expression levels of circRNA_MYLK were closely related to a poor
prognosis in patients with ovarian cancer (73). CiRS-7 has also been revealed to be
upregulated in ovarian cancer, which is positively associated with
TNM stage, lymph node metastasis status and poor prognosis.
Moreover, Fan et al (70)
discovered that circMAN1A2 was highly expressed in the serum of
patients with various malignant tumors, including ovarian cancer,
indicating its potential use as a serum biomarker. However, further
clinical investigation is required to support these findings.
CircRNAs in tumor progression
Currently, the most common clinical treatment for
ovarian cancer is tumor debulking surgery followed by platinum and
paclitaxel chemotherapy. However, the poor prognosis of patients
with advanced ovarian cancer remains a problem worldwide, and
exploratory studies of pathogenesis and potential therapeutic
methods are necessary. Zhang et al (74) revealed that hsa_circ_0051240 was
upregulated in ovarian cancer compared with normal tissues, and
that silencing hsa_circ_0051240 inhibited the proliferation,
migration, invasion and tumor formation of ovarian cancer in
vivo. The data also indicated that hsa_circ_0051240 was able to
sponge miR-637, which directly targets KLK4 mRNA in ovarian cancer
cells. It was therefore speculated that the
hsa_circ_0051240-miR-637-KLK4 axis may be a potential treatment
target for regulating the proliferation, migration and invasion of
ovarian cancer.
circ-ABCB10 was also revealed to be upregulated in
ovarian cancer, and was closely associated with large tumor size,
poor differentiation, poor prognosis and an advanced FIGO stage.
Chen et al (75) found that
upregulated circ-ABCB10 expression reduced apoptosis but promoted
cellular proliferation in vitro, and was also associated
with the negative regulation of miR-1271, miR-1252 and miR-203. In
addition, the upregulation of circ-SMAD7 negatively regulated
Krüppel-like factor 6, mediating ovarian cancer progression,
metastasis and cellular proliferation (76). Moreover, the capacity of
VPS13C-has-circ-001567 to regulate the apoptosis and proliferation
of ovarian cancer cells indicates that it may hold potential as a
therapeutic target. VPS13C-has-circ-001567-knockdown inhibited the
proliferation and tumorigenicity, but promoted the apoptosis of
ovarian cancer in vitro; the cell cycle was arrested at the
G1 phase, and the percentage of S1 phase
cells was decreased (77).
Collectively, these findings suggest that circRNAs play
indispensable roles in the regulation of ovarian cancer cell
proliferation, migration and invasiveness, highlighting their
potential use as therapeutic agents.
CircRNAs suppress ovarian cancer
progression
Chen et al (78) observed that circRNA CDR1as acted as
a vital factor in the growth and metastasis of ovarian cancer.
Hypoxia-inducible factor 1α inhibitor (HIF1AN) is an asparagine
hydroxylase that is closely associated with cancer progression. As
an miRNA sponge, CDR1 as inhibited miR-135b-5p to increase the
expression of downstream HIF1AN, thus ultimately contributing to
the suppression of ovarian cancer. hsa_circ_0078607 has also been
found to be significantly downregulated in ovarian cancer (79). Additionally, hsa_circ_0078607
sponging of miR-518a-5p enhanced the expression of tumor necrosis
factor receptor superfamily member 6 (Fas), which interacts with
its receptor Fas ligand to induce the death signal cascade,
resulting in apoptosis. This indicated that hsa_circ_0078607
suppressed ovarian cancer progression via the miR-518a-5p/Fas
signaling pathway. Furthermore, Li et al (80) observed that circ_100395 facilitated
the expression of p53 by modulating miR-1228, further inhibiting
the growth and metastasis of ovarian tumors.
CircRNAs as biomarkers of ovarian
cancer chemoresistance
Chemoresistance is a common phenomenon during cancer
therapy that is caused by drug resistance, and results in treatment
failure and poor patient prognosis. Chemoresistance frequently
occurs in those with ovarian cancer, making it difficult to meet
the chemotherapeutic standards of efficacy. It is therefore
necessary to improve our understanding of the molecular mechanisms
involved in ovarian cancer chemoresistance.
Wang et al (81) determined the levels of serum
circSETDB1 in 60 patients with serous ovarian cancer, in order to
evaluate its association with progression-free survival. They
observed that higher levels of serum circSETDB1 were significantly
associated with lymph node metastasis and advanced clinical stage,
and could act as distinguishing indicators of patients and healthy
volunteers. Notably, the levels of serum circSETDB1 were
significantly increased in patients with primary chemoresistance,
suggesting that serum circSETDB1 may be a predictor of progression,
response to chemotherapy and relapse in ovarian cancer. To
investigate the potential mechanisms involved, Zhao et al
(82) evaluated the expression of
circRNAs in cisplatin-resistant and -sensitive ovarian cancer
tissues. Cisplatin-resistant patient tissues and cells exhibited
lower expression levels of Cdr1as. Also, Cdr1as was determined to
sponge miR-1270 and regulate the suppression of cancer cell
invasiveness. These data indicate that ovarian cancer sensitivity
to platinum may be enhanced by Cdr1as via the miR-1270/SCAI
signaling pathway.
Perspectives
To date, the abnormal expression of numerous
circRNAs has been detected in ovarian cancer, as well as the
underlying mechanisms of some of these circRNAs. However, the
resulting data are still limited by insufficient sample size and
the lack of an expression profile for pathological classification.
Therefore, further studies are required to fully address this
clinical issue.
CircRNAs and endometrial cancer
(Table III)
Endometrial cancer is the fourth most common female
malignant tumor worldwide, with ~300,000 newly diagnosed cases
annually (39). The incidence of
endometrial cancer is region-dependent, but is increasing with the
aging of the population (83).
Obesity, diabetes, polycystic ovary syndrome, infertility, early
menarche and late menopause have all been reported as potential
risk factors (84,85). Although obesity and hyperinsulinemia
in young women have increased significantly, the primary age of
onset for endometrial cancer is in postmenopausal women, and the
average age of cancer diagnosis is 63 years (86,87).
According to histological subtype, endometrial cancer can be
classified into four types, including endometrioid, serous, clear
cell and mixed endometrial cancer. It can also be divided into two
types depending on the tumor response to estrogen. Type I tumors
are estrogen-responsive, account for ~80–90% of endometrial cancer
cases, and are associated with a more favorable prognosis. By
contrast, type II tumors are usually estrogen-independent, and are
associated with a high grade and poor prognosis; despite accounting
for only 10–20% of all cases, type II tumors are responsible for
40% of the total deaths from endometrial cancer (87–89). A
recent study of overall mutational burden introduced novel
classification criteria with different clinical outcomes. The
Cancer Genome Atlas includes four types of endometrial cancer,
including those with p53, polymerase epsilon and phosphatase and
TENsin homolog mutations, microsatellite instability and histology
(90). Early detection tends to
result in improved prognosis; although postmenopausal bleeding is
the most common symptom of endometrial cancer, it is a diagnostic
indicator in only 5–10% of cases. At present, the primary screening
methods for endometrial cancer are transvaginal ultrasound
scanning, outpatient hysteroscopy and endometrial biopsy. These
procedures are expensive, and due to interference, can be difficult
to perform effectively. Surgery combined with chemotherapy is
currently the most commonly used therapeutic method for endometrial
cancer, and a combination of carboplatin and paclitaxel is the
first-line treatment of choice (91). However, poor prognosis with a rapid
onset of resistance is still a significant issue. In summary, it is
necessary to identity novel diagnostic and therapeutic methods to
increase the survival rate of patients with endometrial cancer.
Expression and biomarker potential of
circRNAs in endometrial cancer
A number of potential cancer biomarkers have been
identified using high-throughput technologies. These include
circRNAs, which serve regulatory roles in various physiological and
pathological processes. In endometrial cancer, the roles of
circRNAs in cells and tissues are gradually being revealed. Recent
studies have shown that 75,928 unique circRNAs are abnormally
expressed in endometrial cancer, and that 62,167 of these are
significantly up- or downregulated in endometrial cancer tissues
compared with adjacent normal tissues (92)C. The top 10 up- and downregulated
circRNAs were selected for further investigation. Among them, the
expression levels of hsa_circ_0001610 and hsa_circ_0039569 were
lower in grade 1–2, compared with grade 3 endometrial cancer
tissues, and age, tumor size, lymph node metastasis, myometrial
invasion and FIGO stage were not associated with hsa_circ_0039569
expression level. However, hsa_circ_0001610 and hsa_circ_0039569
were significantly associated with tumor differentiation. The
correlation between these circRNAs and miRNAs was also analyzed,
indicating that that the
hsa_circ_0039569-has-miR-542-3p/has-let-7c-5p axis serves a
regulatory role in grade 3 endometrial cancer. In summary, this
study indicated that circRNAs may play a vital role in the
diagnosis and treatment of grade 3 endometrial cancer, for which
hsa_circ_0039569 may be a predictive indicator. Another study
identified 209 upregulated and 66 downregulated circRNAs in
extracellular vesicles, that were separated from the serum of
patients with endometrial cancer. Of these 275 differentially
expressed circRNAs, only hsa_circ_0109046 and hsa_circ_0002577
reached a fold-change >2, thus the potential interactions
between these two circRNAs and miRNAs were predicted.
circRNAs have also been revealed to be potential
biomarkers for the diagnosis of endometrial cancer (93). Chen et al (94) highlighted that the overall abundance
of circRNAs is higher in the normal endometrium than in endometrial
cancer tissues. Further research revealed 120 circRNAs that are
differentially expressed between normal endometrial and endometrial
cancer tissues, the majority of which are derived from exons. The
transcriptional products of these circRNAs predispose endometrial
tissues to malignancy. In summary, the results of the
aforementioned studies indicate that circRNAs may serve as
potential biomarkers for the diagnosis and progression of
endometrial cancer.
The tumor suppressor gene PTEN plays an
indispensable role in a number of biological processes, including
cellular survival and apoptosis, proliferation and the maintenance
of genomic stability. Circ-ITCH can combine with miRNA-17 and
miRNA-224 to induce the differential expression of p21 and PTEN. In
animal models, PTEN-knockdown promoted the development of
endometrial cancer and its precursors (95). Also, women carrying germline PTEN
mutations are at a higher risk of developing endometrial cancer
(96), suggesting that different
molecular mechanisms are involved in the regulation of circRNAs in
this malignancy. Another study revealed that two circRNAs, HSPG2
and RP11255H23.4, are expressed in normal endometrial tissues, but
not in endometrial cancer tissues. In normal tissues, the
expression levels of the corresponding miRNAs were also increased,
indicating that circRNAs can competitively bind to related miRNAs,
promoting the development of endometrial cancer. The interaction
between HSPG2 and various growth factors was also found to regulate
endothelial growth and regeneration. Collectively, these findings
suggest that circRNAs may act as diagnostic biomarkers and
therapeutic targets for endometrial cancer.
CircRNAs in endometrial cancer
progression
At present, few studies have investigated the roles
of circRNAs in endometrial cancer progression. Liu et al
(97) discovered that circWHSC1,
which is highly expressed in cancer tissues compared with normal
tissues, could regulate nucleophosmin 1 (NPM1), a downstream target
of miR-646. This was subsequently found to be associated with the
clinical stage and histological grade of endometrial cancer.
CircWHSC1 may therefore promote the development of endometrial
cancer by sponging miR-646 and targeting NPM1.
Conclusions
Bioinformatics and sequencing techniques have been
used to identity numerous novel circRNAs involved in the
development of cancer. Current studies indicate that circRNAs play
an important role in the regulation of cellular proliferation,
migration and invasiveness in cervical, ovarian and endometrial
cancer, primarily via the miRNA sponging mechanism. Due to the
stability of circRNAs, they possess great potential in tumor
diagnosis and treatment. Furthermore, circRNAs can regulate
gynecological tumors through a variety of molecular mechanisms, and
blocking these pathways may represent novel therapeutic
methodologies.
Although sequencing techniques have identified a
growing number of circRNAs, their functions in gynecological cancer
are still largely unclear. Moreover, the sample populations of
recent studies have been relatively small and from a single study
center, thus the reliability of the results cannot be fully
guaranteed. Therefore, the introduction of more clinically
available sample types, such as serum and urine, would improve
study convenience. In addition, current research is primarily
focused on the sponging function of circRNAs, with limited research
into other functions. Moreover, due to the complexity of tumor
pathogenesis, the mechanisms of circRNAs in tumors are not
particularly clear, and further research is required to investigate
these mechanisms in more depth.
Based on current research, the present review
summarizes the potential of circRNAs as novel biomarkers for the
diagnosis and treatment of cervical, ovarian cancer and endometrial
cancer. However, further research is required to support these
findings.
Acknowledgements
Not applicable.
Funding
This project was supported by the National Natural
Science Foundation of China (grant no. 81802586).
Availability of data and materials
Not applicable.
Authors' contributions
YS and RH contributed to the conception of the
manuscript and wrote the draft. YY collected and prepared the
literature. YH and KS revised the manuscript. LZ and BW provided
funding and proofread the manuscript. All authors read and reviewed
the final manuscript version.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:3852–3856. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen LL: The biogenesis and emerging roles
of circular RNAs. Nat Rev Mol Cell Biol. 17:205–211. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Han B, Chao J and Yao H: Circular RNA and
its mechanisms in disease: From the bench to the clinic. Pharmacol
Ther. 187:31–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Enuka Y, Lauriola M, Feldman ME, Sas-Chen
A, Ulitsky I and Yarden Y: Circular RNAs are long-lived and display
only minimal early alterations in response to a growth factor.
Nucleic Acids Res. 44:1370–1383. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bachmayr-Heyda A, Reiner AT, Auer K,
Sukhbaatar N, Aust S, Bachleitner-Hofmann T, Mesteri I, Grunt TW,
Zeillinger R and Pils D: Correlation of circular RNA abundance with
proliferation-exemplified with colorectal and ovarian cancer,
idiopathic lung fibrosis, and normal human tissues. Sci Rep.
5:80572015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kramer MC, Liang D, Tatomer DC, Gold B,
March ZM, Cherry S and Wilusz JE: Combinatorial control of
drosophila circular RNA expression by intronic repeats, hnRNPs, and
SR proteins. Genes Dev. 29:2168–2182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Suzuki H, Zuo Y, Wang J, Zhang MQ,
Malhotra A and Mayeda A: Characterization of RNase R-digested
cellular RNA source that consists of lariat and circular RNAs from
pre-mRNA splicing. Nucleic Acids Res. 34:e632006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zheng S, Gu T, Bao X, Sun J, Zhao J, Zhang
T and Zhang L: Circular RNA hsa_circ_0014243 may serve as a
diagnostic biomarker for essential hypertension. Exp Ther Med.
17:1728–1736. 2019.PubMed/NCBI
|
|
13
|
Ye YL, Yin J, Hu T, Zhang LP, Wu LY and
Pang Z: Increased circulating circular RNA_103516 is a novel
biomarker for inflammatory bowel disease in adult patients. World J
Gastroenterol. 25:6273–6288. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lei B, Tian Z, Fan W and Ni B: Circular
RNA: A novel biomarker and therapeutic target for human cancers.
Int J Med Sci. 16:292–301. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P
and Wu M: CircRNA: Functions and properties of a novel potential
biomarker for cancer. Mol Cancer. 16:942017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Aufiero S, Reckman YJ, Pinto YM and
Creemers EE: Circular RNAs open a new chapter in cardiovascular
biology. Nat Rev Cardiol. 16:503–514. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kong D, Piao YS, Yamashita S, Oshima H,
Oguma K, Fushida S, Fujimura T, Minamoto T, Seno H, Yamada Y, et
al: Inflammation-induced repression of tumor suppressor miR-7 in
gastric tumor cells. Oncogene. 31:3949–3960. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu S, Zhang P, Chen Z, Liu M, Li X and
Tang H: MicroRNA-7 downregulates XIAP expression to suppress cell
growth and promote apoptosis in cervical cancer cells. FEBS Lett.
587:2247–2253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang H, Cai K, Wang J, Wang X, Cheng K,
Shi F, Jiang L, Zhang Y and Dou J: MiR-7, inhibited indirectly by
lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of
breast cancer stem cells by downregulating the STAT3 pathway. Stem
Cells. 32:2858–2868. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang N, Li X, Wu CW, Dong Y, Cai M, Mok
MT, Wang H, Chen J, Ng SS, Chen M, et al: microRNA-7 is a novel
inhibitor of YY1 contributing to colorectal tumorigenesis.
Oncogene. 32:5078–5088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Zheng F, Xiao X, Xie F, Tao D, Huang
C, Liu D, Wang M, Wang L, Zeng F and Jiang G: CircHIPK3 sponges
miR-558 to suppress heparanase expression in bladder cancer cells.
EMBO Rep. 18:1646–1659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xiao-Long M, Kun-Peng Z and Chun-Lin Z:
Circular RNA circ_HIPK3 is down-regulated and suppresses cell
proliferation, migration and invasion in osteosarcoma. J Cancer.
9:1856–1862. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu H, Chen Y and Jiang P: Circular RNA
HIPK3 exerts oncogenic properties through suppression of miR-124 in
lung cancer. Biochem Biophys Res Commun. 506:455–462. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kai D, Yannian L, Yitian C, Dinghao G, Xin
Z and Wu J: Circular RNA HIPK3 promotes gallbladder cancer cell
growth by sponging microRNA-124. Biochem Biophys Res Commun.
503:863–869. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Du WW, Yang W, Liu E, Yang Z, Dhaliwal P
and Yang BB: Foxo3 circular RNA retards cell cycle progression via
forming ternary complexes with p21 and CDK2. Nucleic Acids Res.
44:2846–2858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang
Z and Yang BB: Induction of tumor apoptosis through a circular RNA
enhancing Foxo3 activity. Cell Death Differ. 24:357–370. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing.
Molecular Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Q, Du WW, Wu N, Yang W, Awan FM, Fang
L, Ma J, Li X, Zeng Y, Yang Z, et al: A circular RNA promotes
tumorigenesis by inducing c-myc nuclear translocation. Cell Death
Differ. 24:1609–1620. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen N, Zhao G, Yan X, Lv Z, Yin H, Zhang
S, Song W, Li X, Li L, Du Z, et al: A novel FLI1 exonic circular
RNA promotes metastasis in breast cancer by coordinately regulating
TET1 and DNMT1. Genome Biol. 19:2182018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen CY and Sarnow P: Initiation of
protein synthesis by the eukaryotic translational apparatus on
circular RNAs. Science. 268:415–417. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Legnini I, Di Timoteo G, Rossi F, Morlando
M, Briganti F, Sthandier O, Fatica A, Santini T, Andronache A, Wade
M, et al: Circ-ZNF609 is a circular RNA that can be translated and
functions in myogenesis. Molecular Cell. 66:22–37.e9. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao
F, Huang N, Yang X, Zhao K, Zhou H, et al: Novel role of FBXW7
circular RNA in repressing glioma tumorigenesis. J Natl Cancer
Inst. 110:304–315. 2018. View Article : Google Scholar
|
|
34
|
Zhang M, Huang N, Yang X, Luo J, Yan S,
Xiao F, Chen W, Gao X, Zhao K, Zhou H, et al: A novel protein
encoded by the circular form of the SHPRH gene suppresses glioma
tumorigenesis. Oncogene. 37:1805–1814. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Torre LA, Siegel RL, Ward EM and Jemal A:
Global cancer incidence and mortality rates and trends-an update.
Cancer Epidemiol Biomarkers Prev. 25:16–27. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang S and Batur P: Human papillomavirus
in 2019: An update on cervical cancer prevention and screening
guidelines. Cleve Clin J Med. 86:173–178. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yee GP, de Souza P and Khachigian LM:
Current and potential treatments for cervical cancer. Curr Cancer
Drug Targets. 13:205–220. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mezei AK, Armstrong HL, Pedersen HN,
Campos NG, Mitchell SM, Sekikubo M, Byamugisha JK, Kim JJ, Bryan S
and Ogilvie GS: Cost-effectiveness of cervical cancer screening
methods in low- and middle-income countries: A systematic review.
Int J Cancer. 141:437–446. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9:e10037772013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Han YN, Xia SQ, Zhang YY, Zheng JH and Li
W: Circular RNAs: A novel type of biomarker and genetic tools in
cancer. Oncotarget. 8:64551–64563. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zheng SR, Zhang HR, Zhang ZF, Lai SY,
Huang LJ, Liu J, Bai X, Ding K and Zhou JY: Human papillomavirus 16
E7 oncoprotein alters the expression profiles of circular RNAs in
Caski cells. J Cancer. 9:3755–3764. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gao YL, Zhang MY, Xu B, Han LJ, Lan SF,
Chen J, Dong YJ and Cao LL: Circular RNA expression profiles reveal
that hsa_circ_0018289 is up-regulated in cervical cancer and
promotes the tumorigenesis. Oncotarget. 8:86625–86633. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiao J, Zhang T, Jiao X, Huang T, Zhao L,
Ma D and Cui B: hsa_circ_0000745 promotes cervical cancer by
increasing cell proliferation, migration, and invasion. J Cell
Physiol. 235:1287–1295. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang Y, Wang L, Wang W and Guo X:
Overexpression of circular RNA hsa_circ_0001038 promotes cervical
cancer cell progression by acting as a ceRNA for miR-337-3p to
regulate cyclin-M3 and metastasis-associated in colon cancer 1
expression. Gene. 733:1442732019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen RX, Liu HL, Yang LL, Kang FH, Xin LP,
Huang LR, Guo QF and Wang YL: Circular RNA circRNA_0000285 promotes
cervical cancer development by regulating FUS. Eur Rev Med
Pharmacol Sci. 23:8771–8778. 2019.PubMed/NCBI
|
|
48
|
Hu C, Wang Y, Li A, Zhang J, Xue F and Zhu
L: Overexpressed circ_0067934 acts as an oncogene to facilitate
cervical cancer progression via the miR-545/EIF3C axis. J Cell
Physiol. 234:9225–9232. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ding L and Zhang H: Circ-ATP8A2 promotes
cell proliferation and invasion as a ceRNA to target EGFR by
sponging miR-433 in cervical cancer. Gene. 705:103–108. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ma H, Tian T, Liu X, Xia M, Chen C, Mai L,
Xie S and Yu L: Upregulated circ_0005576 facilitates cervical
cancer progression via the miR-153/KIF20A axis. Biomed
Pharmacother. 118:1093112019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiao J, Jiao X, Liu Q, Qu W, Ma D, Zhang Y
and Cui B: The Regulatory Role of circRNA_101308 in cervical cancer
and the prediction of its mechanism. Cancer Manag Res.
12:4807–4815. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ji F, Du R, Chen T, Zhang M, Zhu Y, Luo X
and Ding Y: Circular RNA circSLC26A4 accelerates cervical cancer
progression via miR-1287-5p/HOXA7 axis. Mol Ther Nucleic Acids.
19:413–420. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
He J, Lv X and Zeng Z: A potential disease
monitoring and prognostic biomarker in cervical cancer patients:
The clinical application of circular RNA_0018289. J Clin Lab Anal.
34:e233402020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang J, Zhao X, Zhang J, Zheng X and Li
F: Circular RNA hsa_circ_0023404 exerts an oncogenic role in
cervical cancer through regulating miR-136/TFCP2/YAP pathway.
Biochem Biophys Res Commun. 501:428–433. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tang Q, Chen Z, Zhao L and Xu H: Circular
RNA hsa_circ_0000515 acts as a miR-326 sponge to promote cervical
cancer progression through up-regulation of ELK1. Aging (Albany
NY). 11:9982–9999. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu H, Wu Y, Wang S, Jiang J, Zhang C,
Jiang Y, Wang X, Hong L and Huang H: Circ-SMARCA5 suppresses
progression of multiple myeloma by targeting miR-767-5p. BMC
Cancer. 19:9372019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dong C, Fan B, Ren Z, Liu B and Wang Y:
CircSMARCA5 facilitates the progression of prostate cancer through
miR-432/PDCD10 axis. Cancer Biother Radiopharm. 2020.(Online ahead
of print). View Article : Google Scholar
|
|
58
|
Tian JDC and Liang L: Involvement of
circular RNA SMARCA5/microRNA-620 axis in the regulation of
cervical cancer cell proliferation, invasion and migration. Eur Rev
Med Pharmacol Sci. 22:8589–8598. 2018.PubMed/NCBI
|
|
59
|
Yang C, Yuan W, Yang X, Li P, Wang J, Han
J, Tao J, Li P, Yang H, Lv Q and Zhang W: Circular RNA circ-ITCH
inhibits bladder cancer progression by sponging miR-17/miR-224 and
regulating p21, PTEN expression. Mol Cancer. 17:192018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wang ST, Liu LB, Li XM, Wang YF, Xie PJ,
Li Q, Wang R, Wei Q, Kang YH, Meng R and Feng XH: Circ-ITCH
regulates triple-negative breast cancer progression through the
Wnt/β-catenin pathway. Neoplasma. 66:232–239. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Li J, Guo R, Liu Q, Sun J and Wang H:
Circular RNA Circ-ITCH inhibits the malignant behaviors of cervical
cancer by microRNA-93-5p/FOXK2 Axis. Reprod Sci. 27:860–868. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Cai H, Zhang P, Xu M, Yan L, Liu N and Wu
X: Circular RNA hsa_circ_0000263 participates in cervical cancer
development by regulating target gene of miR-150-5p. J Cell
Physiol. 234:11391–11400. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics, 2018. CA Cancer J Clin. 68:284–296.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hackshaw A, Gershenson D and Ledermann J:
Mucinous Ovarian Carcinoma. N Engl J Med. 381:e32019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang L, Wang Q, Xu Y and Han L: Advances
in the treatment of ovarian cancer using PARP inhibitors and the
underlying mechanism of resistance. Current Drug Targets.
21:167–178. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hansen JM, Coleman RL and Sood AK:
Targeting the tumour microenvironment in ovarian cancer. Eur J
Cancer. 56:131–143. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Teng F, Xu J, Zhang M, Liu S, Gu Y, Zhang
M, Wang X, Ni J, Qian B, Shen R and Jia X: Comprehensive circular
RNA expression profiles and the tumor-suppressive function of
circHIPK3 in ovarian cancer. Int J Biochem Cell Biol. 112:8–17.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ning L, Long B, Zhang W, Yu M, Wang S, Cao
D, Yang J, Shen K, Huang Y and Lang J: Circular RNA profiling
reveals circEXOC6B and circN4BP2L2 as novel prognostic biomarkers
in epithelial ovarian cancer. Int J Oncol. 53:2637–2646.
2018.PubMed/NCBI
|
|
69
|
Gao Y, Zhang C, Liu Y and Wang M: Circular
RNA profiling reveals circRNA1656 as a novel biomarker in high
grade serous ovarian cancer. Biosci Trends. 13:204–211. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fan CM, Wang JP, Tang YY, Zhao J, He SY,
Xiong F, Guo C, Xiang B, Zhou M, Li XL, et al: circMAN1A2 could
serve as a novel serum biomarker for malignant tumors. Cancer Sci.
110:2180–2188. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Pei C, Wang H, Shi C, Zhang C and Wang M:
CircRNA hsa_circ_0013958 may contribute to the development of
ovarian cancer by affecting epithelial-mesenchymal transition and
apoptotic signaling pathways. J Clin Lab Anal. 34:e232922020.
View Article : Google Scholar
|
|
72
|
Zou T, Wang PL, Gao Y and Liang WT:
Circular RNA_LARP4 is lower expressed and serves as a potential
biomarker of ovarian cancer prognosis. Eur Rev Med Pharmacol Sci.
22:7178–7182. 2018.PubMed/NCBI
|
|
73
|
Zhao Y, Hu Y, Shen Q, Chen Q, Zhu XJ,
Jiang SS and Zhang Q: CircRNA_MYLK promotes malignant progression
of ovarian cancer through regulating microRNA-652. Eur Rev Med
Pharmacol Sci. 24:5281–5291. 2020.PubMed/NCBI
|
|
74
|
Zhang M, Xia B, Xu Y, Zhang Y, Xu J and
Lou G: Circular RNA (hsa_circ_0051240) promotes cell proliferation,
migration and invasion in ovarian cancer through miR-637/KLK4 axis.
Artif Cells Nanomed Biotechnol. 47:1224–1233. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Chen Y, Ye X, Xia X and Lin X: Circular
RNA ABCB10 correlates with advanced clinicopathological features
and unfavorable survival, and promotes cell proliferation while
reduces cell apoptosis in epithelial ovarian cancer. J Nurs
Scholarsh. 26:151–161. 2019.
|
|
76
|
Zhao Y, Qin XP, Lang YP, Kou D and Shao
ZW: Circular RNA circ-SMAD7 promoted ovarian cancer cell
proliferation and metastasis by suppressing KLF6. Eur Rev Med
Pharmacol Sci. 23:5603–5610. 2019.PubMed/NCBI
|
|
77
|
Bao L, Zhong J and Pang L: Upregulation of
Circular RNA VPS13C-has-circ-001567 promotes ovarian cancer cell
proliferation and invasion. Cancer Biother Radiopharm. 34:110–118.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chen H, Mao M, Jiang J, Zhu D and Li P:
Circular RNA CDR1as acts as a sponge of miR-135b-5p to suppress
ovarian cancer progression. Onco Targets Ther. 12:3869–3879. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhang N, Jin Y, Hu Q, Cheng S, Wang C,
Yang Z and Wang Y: Circular RNA hsa_circ_0078607 suppresses ovarian
cancer progression by regulating miR-518a-5p/Fas signaling pathway.
J Ovarian Res. 13:642020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Li X, Lin S, Mo Z, Jiang J, Tang H, Wu C
and Song J: CircRNA_100395 inhibits cell proliferation and
metastasis in ovarian cancer via regulating
miR-1228/p53/epithelial-mesenchymal transition (EMT) axis. J
Cancer. 11:599–609. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang W, Wang J, Zhang X and Liu G: Serum
circSETDB1 is a promising biomarker for predicting response to
platinum-taxane-combined chemotherapy and relapse in high-grade
serous ovarian cancer. Onco Targets Ther. 12:7451–7457. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhao Z, Ji M, Wang Q, He N and Li Y:
Circular RNA Cdr1as upregulates SCAI to suppress cisplatin
resistance in ovarian cancer via miR-1270 suppression. Mol Ther
Nucleic Acids. 18:24–33. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Lortet-Tieulent J, Ferlay J, Bray F and
Jemal A: International patterns and trends in endometrial cancer
incidence, 1978–2013. J Natl Cancer Inst. 110:354–361. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
McAlpine JN, Temkin SM and Mackay HJ:
Endometrial cancer: Not your grandmother's cancer. Cancer.
122:2787–2798. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Braun MM, Overbeek-Wager EA and Grumbo RJ:
Diagnosis and management of endometrial cancer. Am Fam Physician.
93:468–474. 2016.PubMed/NCBI
|
|
86
|
Moore K and Brewer MA: Endometrial cancer:
Is this a new disease? Am Soc Clin Oncol Educ Book. 37:435–442.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Urick ME and Bell DW: Clinical
actionability of molecular targets in endometrial cancer. Nat Rev
Cancer. 19:510–521. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Bokhman JV: Two pathogenetic types of
endometrial carcinoma. Gynecol Oncol. 15:10–17. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Suarez AA, Felix AS and Cohn DE: Bokhman
Redux: Endometrial cancer ‘types’ in the 21st century. Gynecol
Oncol. 144:243–249. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Cancer Genome Atlas Research Network, .
Kandoth C, Schultz N, Cherniack AD, Akbani R, Liu Y, Shen H,
Robertson AG, Pashtan I, Shen R, et al: Integrated genomic
characterization of endometrial carcinoma. Nature. 497:67–73. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Colombo N, Creutzberg C, Amant F, Bosse T,
González-Martín A, Ledermann J, Marth C, Nout R, Querleu D, Mirza
MR, et al: ESMO-ESGO-ESTRO Consensus Conference on Endometrial
Cancer: diagnosis, treatment and follow-up. Ann Oncol. 27:16–41.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ye F, Tang QL, Ma F, Cai L, Chen M, Ran
XX, Wang XY and Jiang XF: Analysis of the circular RNA
transcriptome in the grade 3 endometrial cancer. Cancer Manag Res.
11:6215–6227. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Xu H, Gong Z, Shen Y, Fang Y and Zhong S:
Circular RNA expression in extracellular vesicles isolated from
serum of patients with endometrial cancer. Epigenomics. 10:187–197.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Chen BJ, Byrne FL, Takenaka K, Modesitt
SC, Olzomer EM, Mills JD, Farrell R, Hoehn KL and Janitz M:
Analysis of the circular RNA transcriptome in endometrial cancer.
Oncotarget. 9:5786–5796. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wang H, Douglas W, Lia M, Edelmann W,
Kucherlapati R, Podsypanina K, Parsons R and Ellenson LH: DNA
mismatch repair deficiency accelerates endometrial tumorigenesis in
Pten heterozygous mice. Am J Pathol. 160:1481–1486. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Stambolic V, Tsao MS, Macpherson D, Suzuki
A, Chapman WB and Mak TW: High incidence of breast and endometrial
neoplasia resembling human Cowden syndrome in pten+/- mice. Cancer
Res. 60:3605–3611. 2000.PubMed/NCBI
|
|
97
|
Liu Y, Chen S, Zong ZH, Guan X and Zhao Y:
CircRNA WHSC1 targets the miR-646/NPM1 pathway to promote the
development of endometrial cancer. J Cell Mol Med. 24:6898–6907.
2020. View Article : Google Scholar : PubMed/NCBI
|