Introduction
Colon cancer is a lethal malignancy in the
gastrointestinal tract with high incidence and recurrence, leading
to approximately 1,096,601 new cases and 551,269 deaths in 2018
(1–3). Currently, the methods of colon cancer
therapy are surgery combined with chemotherapy, radiotherapy and
targeted therapy (4). Although
treatment methods have advanced, the incidence of colon cancer
continues to increase and the prognosis of patients remains dismal
(5). Therefore, it is necessary to
elucidate the pathogenic mechanism of colon cancer.
Recently, numerous studies have verified that the
use of anesthetics during surgery may affect the progression of
cancers (6). Sevoflurane is a
commonly used inhaled anesthetic in clinical practice, which plays
a vital role in the progression of various tumor cells. For
example, Liang et al demonstrated that sevoflurane could
suppress the metastasis of lung cancer cells (7). Gao et al revealed that
sevoflurane suppressed the proliferation and metastasis of glioma
cells (8). In the present study,
the functions and mechanisms of sevoflurane in colon cancer were
investigated.
Exosomes are discoid vesicles with a diameter of
50–140 nm (9). Exosomes secreted by
tumor cells can transfer some tumor-specific biological information
to neighboring cells or even distant cells and then promote the
occurrence and development of tumors via delivering proteins,
mRNAs, circular RNAs (circRNAs), microRNAs (miRNAs) and other
bioactive substances (10,11). CircRNAs are a special class of
non-coding RNAs (ncRNAs), which are characterized by closed ring
structures (12). CircRNAs have
emerged as crucial regulators in different types of cancers,
including colon cancer. For example, Zhang et al revealed
that circ-PIP5K1A was abnormally increased and could promote the
progression of colon cancer by inducing cell viability and
metastasis (13). Xu et al
reported that circ_000984 served as an oncogene in colon cancer and
circ_000984 knockdown hampered cell growth, metastasis and tumor
formation (14). It has been
reported that circRNA 3-hydroxy-3-methylglutaryl-CoA synthase 1
(circ-HMGCS1) is associated with the progression of hepatoblastoma
(HB) and colorectal cancer (CRC) (15, 16). However, the studies on
circ-HMGCS1 in colon cancer remain limited.
miRNAs, a series of ncRNAs with approximately 22
nucleotides, mainly alter gene expression by recognizing the
3′-untranslated region (3′UTR) of target mRNAs (17). Multiple miRNAs have been confirmed
to participate in the development of colon cancer via binding to
target genes. For example, miR-28a-5p exerted its tumor-suppressive
role in colon cancer by targeting CAMTA2 (18). miR-223-3p facilitated colon cancer
cell growth and metastasis by binding to PRDM1 (19). miR-204-3p targeted HMGA2 to suppress
cell viability and metastasis and facilitated cell apoptosis in
colon cancer (20). Previous
reports revealed that miR-34a-5p was reduced in CRC and the
increase of miR-34a-5p suppressed tumor metastasis (21,22).
Sphingosine-1-phosphate phosphatase 1 (SGPP1) has been demonstrated
to promote cell growth and migration and hinder cell apoptosis in
CRC (23). However, whether
miR-34a-5p can target SGPP1 to take part in the regulation of colon
cancer remains unclear.
The purpose of this research was to explore the
functions of sevoflurane in colon cancer cell viability, apoptosis
and invasion. In addition, the roles and potential mechanisms of
exosomal circ-HMGCS1, miR-34a-5p and SGPP1 in colon cancer
progression were investigated.
Materials and methods
Human serum collection
The serum samples were collected from 30 colon
cancer patients (19 males and 11 females; age, 50–70 years) and 30
healthy volunteers (17 males and 13 females; age, 48–65 years) at
the First Affiliated Hospital of Zhengzhou University from March
2015 to October 2017. The experiment was conducted following the
approval that was obtained from the Ethics Committee of the First
Affiliated Hospital of Zhengzhou University and written informed
consents were signed by all participants. The collected samples
were stored at −80°C until use.
Cell culture
Two colon cancer cell lines (ATCC®
CCL-228™, SW480; and ATCC® CCL-229, LOVO) were purchased
from the American Type Culture Collection and a normal human colon
mucosal epithelial cell line (C0972; NCM460) was obtained from
Guandao Biological Company (https://www.biomart.cn/infosupply/37016225.htm). All
cells were grown in Roswell Park Memorial Institute (RPMI)-1640
medium (cat. no. A1049101; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (cat. no. 16140063; FBS;
Gibco; Thermo Fisher Scientific, Inc.) at an atmosphere of 5%
CO2 and 37°C.
Sevoflurane treatment
SW480 and LOVO cells (2×103) at the
exponential growth phase were seeded into plates and incubated
overnight. Next, the plates were placed in an airtight glass
chamber. Sevoflurane (product code YZ-1612540; Beijing Solarbio
Science & Technology Co., Ltd.) was added into the chamber
through an anesthetic vaporizer (BS-S6100 Plus; Guangzhou Bisen
Medical Co., Ltd.). A gas monitor (PM8060; Drager) was employed to
monitor the concentrations of sevoflurane. Cells were treated with
various doses (1.7, 3.4 and 5.1%) of sevoflurane for 6 h, and then
maintained in normal conditions for 24 h for further study. Cells
without treatment were used as the control.
Cell transfection
The overexpression plasmid of circ-HMGCS1
(circ-HMGCS1) and its control (pcDNA), small interfering RNA
targeting circ-HMGCS1 (si-circ-HMGCS1; 5′-TGGAAGCCUUGGGGCUUCGU-3′)
and its control (si-NC; 5′-UUCUCCGAACGUGUCACGUTT−3′), miR-34a-5p
mimic (miR-34a-5p; 5′-GAUGGACGUGCUUGUCGUGAAAC-3′) and its control
(miR-NC; 5′-UUCUCCGAACGUGUCACGUTT−3′), miR-34a-5p inhibitor
(anti-miR-34a-5p; 5′-CUACCUGCACCAACAGCACUU−3′) and its control
(anti-miR-NC; 5′-CAGUACUUUUGUGUAGUACAA-3′), lentivirus-mediated
short hairpin against circ-HMGCS1 (sh-circ-HMGCS1;
5′-TTTGGGGCTTCGTGGGACACA−3′) and its control (sh-NC;
5′-TTCTCCGAACGTGTCACGT-3′) were synthesized by Shanghai GenePharma
Co., Ltd. Cell transfection was carried out with the Lipofectamine
2000 reagent (cat. no. 11668019; Invitrogen; Thermo Fisher
Scientific, Inc.).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
The viability of SW480 and LOVO cells was assessed
by MTT assay after relevant treatment. In brief, cells were seeded
into 96-well plates at a density of 1×103 cells/well and
incubated overnight. Then 20 µl MTT (5 mg/ml; item no. IM0280;
Beijing Solarbio Science & Technology Co., Ltd.) was added into
each well after incubation for 24, 48 and 72 h followed by
incubation for another 4 h at 37°C. Next, dimethyl sulfoxide (DMSO;
D8371; Beijing Solarbio Science & Technology Co., Ltd.) was
added to dissolve the formazan crystals. The absorbance at 490 nm
was determined using a microplate reader (Elx808™; BioTek
Instruments, Inc.).
Flow cytometric assay
The Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) Apoptosis Detection Kit (C1062M;
Beyotime Institute of Biotechnology) was employed for the detection
of cell apoptosis according to the instructions of manufacturers.
In brief, SW480 and LOVO cells (1×104) were harvested
and washed with phosphate-buffered saline (PBS; P1022; Beijing
Solarbio Science & Technology Co., Ltd.) after relevant
sevoflurane treatment and transfection. Then 5 µl Annexin V-FITC
and 5 µl PI were added to stain cells for 15 min at room
temperature in the dark. Finally, the rate of apoptotic cells was
examined via a FACScan® flow cytometer (BD Biosciences)
within 1 h and analyzed with software FlowJo (7.6.1; FlowJo LLC).
The apoptotic rate was calculated as the sum of the early apoptosis
rate and the late apoptosis rate.
Transwell assay
The invasion of SW480 and LOVO cells was tested
using a Transwell insert (3379; 8 µm pore size; Corning
Incorporated) which was pre-coated with Matrigel (product no.
356234; Beijing Solarbio Science & Technology Co., Ltd.). Cells
(1×104) in serum-free RPMI-1640 medium were added into
the upper chamber. RPMI-1640 medium supplemented with 10% FBS was
added into the bottom chamber. After 48 h, cells remaining on the
upper chamber were removed and the cells that invaded to the lower
surface were fixed with 70% methanol for 30 min at room temperature
and stained with 0.1% crystal violet (IC0600; Beijing Solarbio
Science & Technology Co., Ltd.) for 15 min at room temperature.
The number of invaded cells was analyzed under a light microscope
(Olympus Corporation) at a magnification of ×100.
Western blot analysis
After being extracted from serums, cells and
exosomes using RIPA buffer (P0013C; Beyotime Institute of
Biotechnology), total protein (20 µg) was quantified by a BCA
Protein Quantification Kit (E112-01/02; Vazyme) and then separated
through 10% sodium dodecyl sulfonate-polyacrylamide gel (SDS-PAGE;
P1200; Beijing Solarbio Science & Technology Co., Ltd.). Then
the proteins were transferred onto polyvinylidene difluoride
membranes (PVDF; 3010040001; EMD Millipore) and blocked in 5% skim
milk for 2 h at room temperature. Next, the membranes were
incubated with primary antibodies against cyclin D1 (product code
ab16663; 1:200), p21 (product code ab109199; 1:1,000), matrix
metallopeptidase 9 (MMP9; product code ab38898; 1:1,000), CD9
(product code ab92726; 1:2,000), CD63 (product code ab68418;
1:2,000), SGPP1 (product code ab108435; 1:2,000), GAPDH (product
code ab181602; 1:5,000) or β-actin (product code ab8227; 1:5,000;
all from Abcam) overnight at 4°C and secondary antibody goat
anti-rabbit IgG H&L (product code ab150077; 1:5,000; Abcam) for
2 h at room temperature. Finally, the protein bands were visualized
by an enhanced chemiluminescence reagent (E411-03/04/05; Vazyme)
and analyzed by ImageJ software (v1.8.0; National Institutes of
Health).
Exosome isolation
The serum-exosomes were isolated with ExoQuick
precipitation kit (EXOQ5A-1; System Biosciences) according to the
manufacturer's instructions. Briefly, ExoQuick solution was added
into the serum samples and incubated for 30 min at 4°C. Then the
mix was centrifuged for 30 min at 1,500 × g at room temperature.
Subsequently, the supernatant was carefully removed and then
centrifuged for 5 min at 1,500 × g to remove the extra liquid.
Exosome pellets were resuspended in PBS (P1022; Beijing Solarbio
Science & Technology Co., Ltd.) and preserved at −80°C.
Exosomes were isolated from cultured cells through differential
ultracentrifugation as previously described (24).
Transmission electron microscopy
(TEM)
Exosome pellets were suspended in PBS (P1022;
Beijing Solarbio Science & Technology Co., Ltd.) and then fixed
with 4% paraformaldehyde (E672002; Shanghai Sangon Biotech Co.,
Ltd.) overnight at 4°C and 4% glutaraldehyde (A600875; Shanghai
Sangon Biotech Co., Ltd.) for 30 min at 4°C in phosphate buffer (pH
7.4), and maintained overnight at 4°C until the TEM assay. The
exosomes were placed on a 400-mesh carbon-coated copper grid and
stained with 2% phosphotungstic acid solution (pH 7.0; G1599;
Solarbio) for 2 min at room temperature. The morphologies of the
samples were examined with a JEM-1200EX transmission electron
microscope (JEOL, Ltd.) at the magnification of ×50,000.
Nanoparticle tracking analysis
(NTA)
The size of the exosomes was examined using the
Nanosight NS 300 system (NanoSight Technology) according to the
manufacturer's instructions.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from serums, cells and
exosomes with TRIzol reagent (15596018; Invitrogen; Thermo Fisher
Scientific, Inc.). Then reverse transcription was conducted using
High Capacity cDNA Reverse Transcription Kit (4368814; Applied
Biosystems; Thermo Fisher Scientific, Inc.) or miRNA 1st Strand
cDNA Synthesis Kit (MR101-01/02; Vazyme) according to the
manufacturer's instructions. Subsequently, RT-qPCR was conducted
using AceQ Universal SYBR qPCR Master Mix (Q511-02; Vazyme) on an
ABI 7500 PCR system (4351104; Applied Biosystems; Thermo Fisher
Scientific, Inc.) under the thermocycling conditions: i) Initial
denaturation at 95°C for 5 min; ii) 40 cycles of 95°C for 10 sec
and 60°C for 30 sec; iii) 95°C for 15 sec, 60°C for 60 sec and 95°C
for 15 sec. The expression was analyzed with the 2−ΔΔCq
method (25). Glyceraldehyde
3-phosphate dehydrogenase (GAPDH) or U6 was used as an internal
control. The primer sequences were listed as follows: circ-HMGCS1
forward, 5′-TCTAGCTCGGATGTTGCTGA-3′ and reverse,
5′-TCAGGCTTGTAAAAATCATAGGC-3′; miR-34a-5p forward,
5′-CTGGGAGGTGGCAGTGTCTTAGC-3′ and reverse,
5′-TCAACTGGTGTCGTGGAGTCGG-3′; SGPP1 forward,
5′-TGGTCCTCCTCACCTATGGC-3′ and reverse, 5′-CTAGAGAACACCAGCAGGGA-3′;
GAPDH forward, 5′-CCGGGAAACTGTGGCGTGATGG-3′ and reverse,
5′-AGGTGGAGGAGTGGGTGTCGCTGTT-3′; U6 forward,
5′-TGCGGGTGCTCGCTTCGGCAGC-3′ and reverse,
5′-CCAGTGCAGGGTCCGAGGT-3′.
Dual-luciferase reporter assay
The potential binding sites between miR-34a-5p and
circ-HMGCS1 or SGPP1 were predicted by starBase v2.0 (http://starbase.sysu.edu.cn/index.php)
and verified by dual-luciferase reporter assay. The fragments of
circ-HMGCS1 or 3′UTR of SGPP1 containing the predicted wild-type or
mutant miR-34a-5p binding sequences were cloned into the pmirGLO
vector (E1330; Promega Corporation) to construct luciferase
reporter plasmids WT-circ-HMGCS1, MUT-circ-HMGCS1, SGPP1 3′UTR-WT
and SGPP1 3′UTR-MUT, respectively. Then miR-34a-5p or miR-NC was
transfected into SW480 or LOVO cells along with relevant plasmid.
Dual-Luciferase Reporter Assay Kit (E1910; Promega Corporation) was
used to detect the luciferase activity according to the
manufacturer's instructions. The Renilla luciferase activity
was normalized to firefly luciferase activity.
RNA immunoprecipitation (RIP)
assay
Magna RIP RNA Binding Protein Immunoprecipitation
Kit (17–700; EMD Millipore) was used to conduct RIP assays.
Briefly, SW480 and LOVO cells were lysed in RIP buffer and then
incubated overnight with magnetic beads coated with antibody
against Argonaute2 (Anti-Ago2; product code ab32381; 1:2,000;
Abcam) or immunoglobulin G (Anti-IgG; product code ab109489;
1:5,000; Abcam) at 4°C. Subsequently, the samples were washed and
incubated with proteinase K (P9460; Beijing Solarbio Science &
Technology Co., Ltd.) at 55°C for 30 min to isolate the RNA-protein
complexes from beads. Finally, RNAs in the magnetic complexes were
extracted and the enrichment of circ-HMGCS1, miR-34a-5p and SGPP1
was analyzed via RT-qPCR.
Murine xenograft model
A total of 24 male nude mice (17–23 g) were
purchased from Shanghai SLAC Laboratory Animals Co., Ltd and
divided into 3 groups (n=8/group): sh-NC, sh-NC+SEV and
sh-circ-HMGCS1+SEV. All the mice were housed in a pathogen-free
condition at 28°C and 45% humidity with a 12-h light/dark cycle and
fed sterile fodder and drinking water. After being transfected with
sh-circ-HMGCS1 or sh-NC for 48 h and treated with 5.1% sevoflurane
for 6 h, 4×106 SW480 cells were cultured overnight and
then subcutaneously inoculated into the right forelimb of the nude
mice. After 7 days, the mice were administered with sevoflurane
every 4 days and the tumor weight was monitored concurrently. The
tumor weight was calculated with the formula: (length ×
width2)/2. Subsequently, 27 days later, the mice were
sacrificed by cervical dislocation. The criteria for confirming the
death of the mice were absence of breathing for 2–3 min and absence
of blink reflex. Tumors were collected and weighted. The collected
tumor samples were stored at −80°C. The experiment was approved by
the Ethics Committee of Animal Research of the First Affiliated
Hospital of Zhengzhou University. The mice with tumors were
monitored daily for proper treatment and for signs of convulsions,
self-injury, dyspnea and other phenomena, where corresponding
countermeasures would be taken. The humane endpoints of the animal
studies were as follows: A tumor burden greater than 10% body
weight and a tumor that did not exceed 20 mm in any one dimension.
The maximal tumor volume in our experiments was 960
mm3.
Statistical analysis
The data collected from three independent
experiments were displayed as the mean ± standard deviation (SD),
and processed by using software GraphPad Prism 7 (GraphPad
Software, Inc.). Paired Student's t-test or one-way analysis
of variance (ANOVA) followed by Tukey's test was applied to compare
differences among groups. The correlations between the expression
of miR-34a-5p and circ-HMGCS1, as well as miR-34a-5p and SGPP1 were
analyzed using Spearman's correlation coefficient analysis. A
P-value <0.05 was considered to indicate a statistically
significant difference.
Results
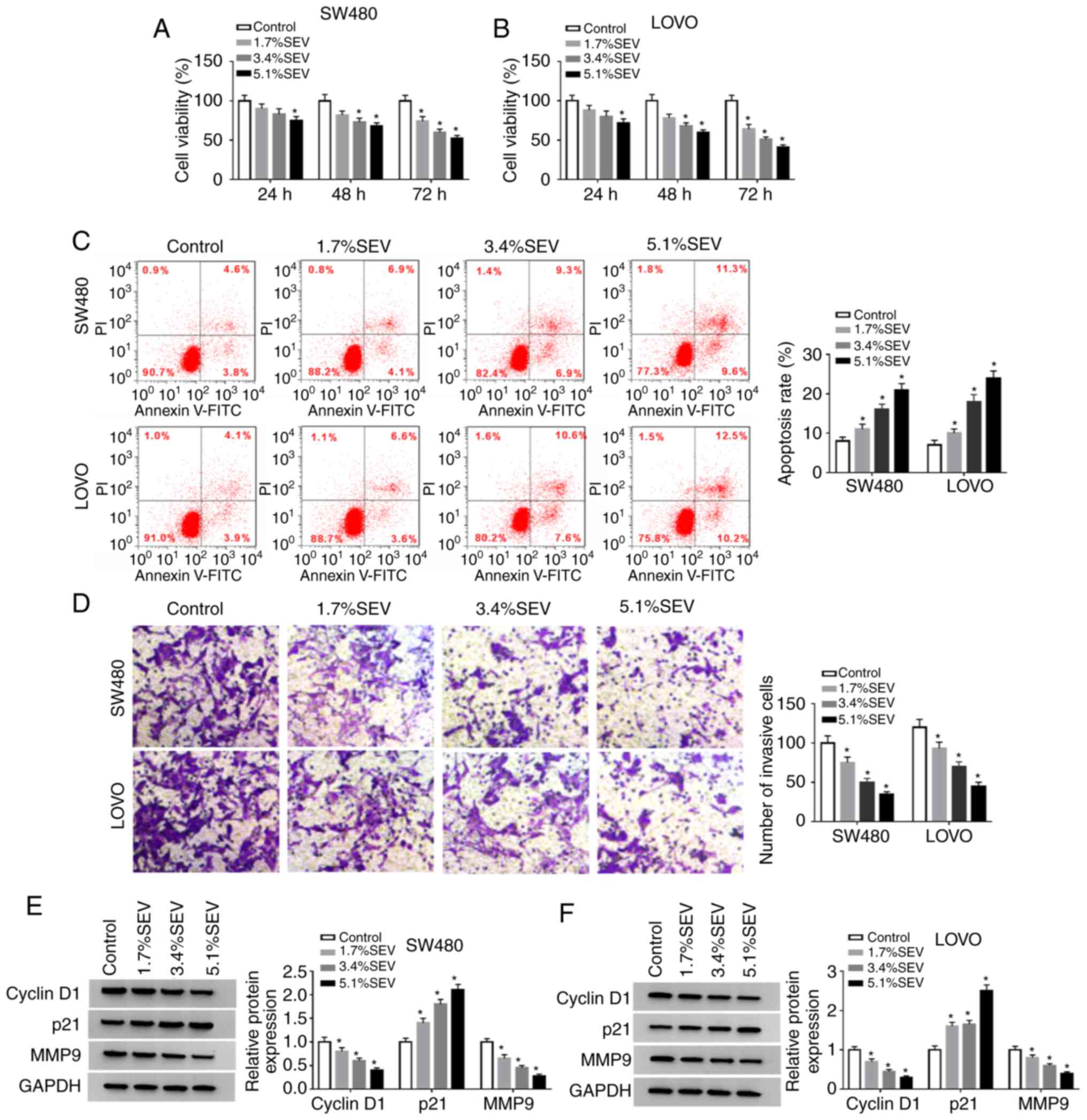
Sevoflurane suppresses cell viability
and invasion and facilitates cell apoptosis in colon cancer
cells
In order to explore the function of sevoflurane in
the progression of colon cancer, SW480 and LOVO cells were treated
with various concentrations (1.7, 3.4 and 5.1%) of sevoflurane and
then cell viability, apoptosis and invasion were investigated. The
data of the MTT assay indicated that compared to control group,
cell viability was significantly suppressed by sevoflurane in a
concentration-dependent manner in both SW480 and LOVO cells
(Fig. 1A and B). Flow cytometric
analysis revealed that the apoptosis of SW480 and LOVO cells was
significantly promoted by the treatment of sevoflurane compared to
control group (Fig. 1C). The number
of invaded cells was significantly reduced in a dose-dependent
manner in SW480 and LOVO cells by sevoflurane treatment compared
with the control group, as determined by Transwell assays (Fig. 1D). In addition, the protein levels
of cyclin D1, p21 and MMP9 in SW480 and LOVO cells treated with
sevoflurane were measured via western blotting. The data revealed
that cyclin D1 and MMP9 were decreased while p21 was increased in a
concentration-dependent manner in SW480 and LOVO cells after
sevoflurane treatment (Fig. 1E and
F). Collectively, the data indicated that sevoflurane could
inhibit the progression of colon cancer cells.
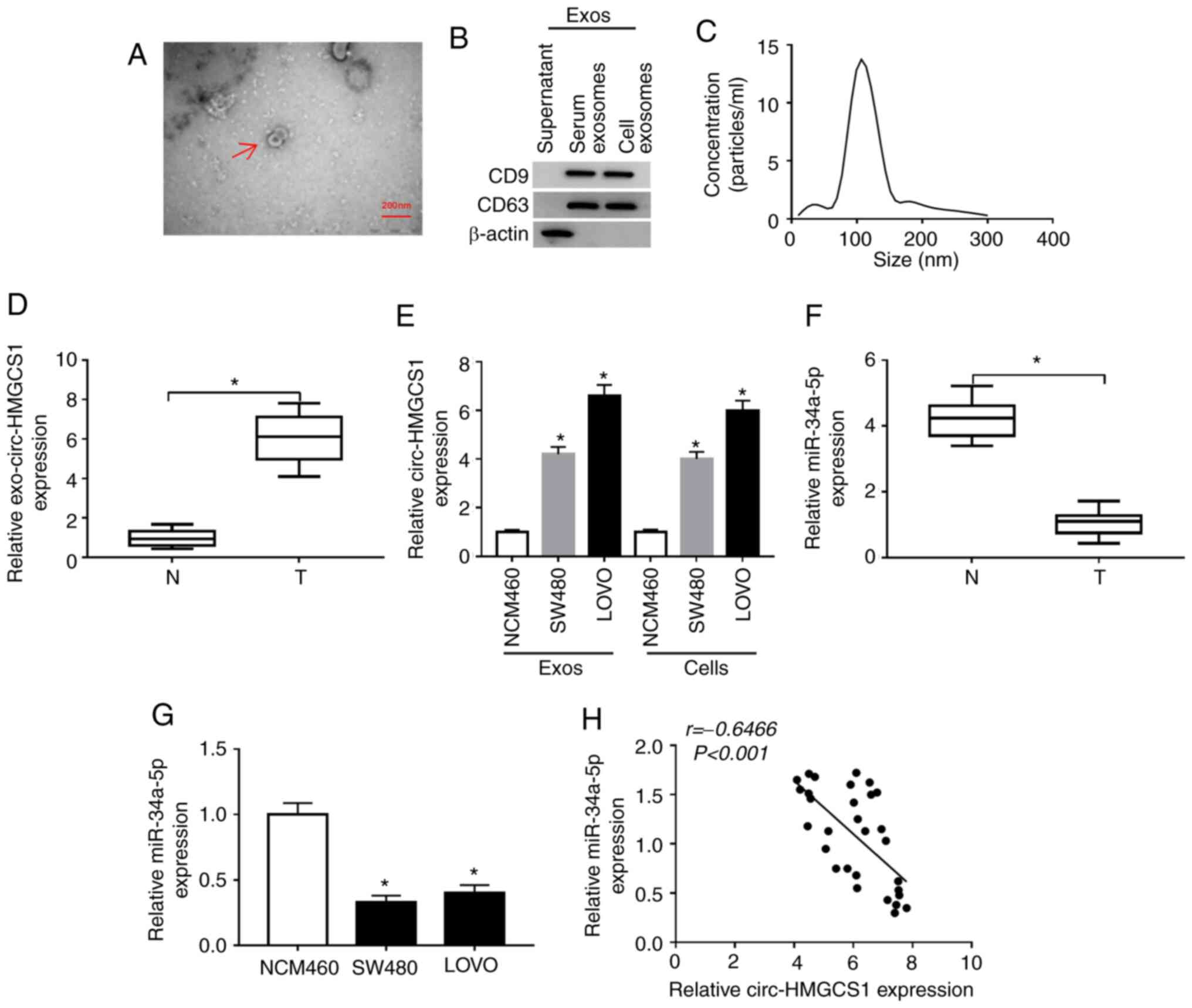
Exosomal circ-HMGCS1 is increased and
cytoplasm miR-34a-5p is decreased in the serums and cells of colon
cancer patients
To analyze the function of circ-HMGCS1 packaged in
exosomes, serum derived from colon cancer patients and colon cancer
cell-derived exosomes were first isolated. Then the morphology of
particles isolated from the serums of the patients was observed by
TEM. It was observed that the particles exhibited typical lipid
bilayer membrane-encapsulated nanoparticles (Fig. 2A). The data of the western blot
assay revealed that the markers of exosomes (CD9 and CD63) could be
detected in the serums of the colon cancer patients and in the
colon cancer cell-derived particles (Fig. 2B). The NTA assay revealed that the
diameters of the particles isolated from the serums of the patients
were 80–130 nm (Fig. 2C). These
data indicated that the isolated particles were exosomes.
Subsequently, the expression of circ-HMGCS1 in the exosomes was
assessed by RT-qPCR. The data revealed that circ-HMGCS1 was
expressed at a higher level in the exosomes from the serums of
colon cancer patients and colon cancer cells (SW480 and LOVO cells)
compared to the exosomes derived from normal serums and NCM460
cells (Fig. 2D and E). In addition,
the expression of miR-34a-5p in the serums of colon cancer patients
and colon cancer cells was examined. As revealed in Fig. 2F and G, miR-34a-5p was downregulated
in the serums of colon cancer patients and colon cancer cells
compared to normal serums and cells. There was an inverse
correlation between circ-HMGCS1 and miR-34a-5p in the serums of
patients with colon cancer, as analyzed by Spearman's correlation
coefficient analysis (Fig. 2H).
Collectively, the aberrant expression of circ-HMGCS1 and miR-34a-5p
may be involved in the progression of colon cancer.
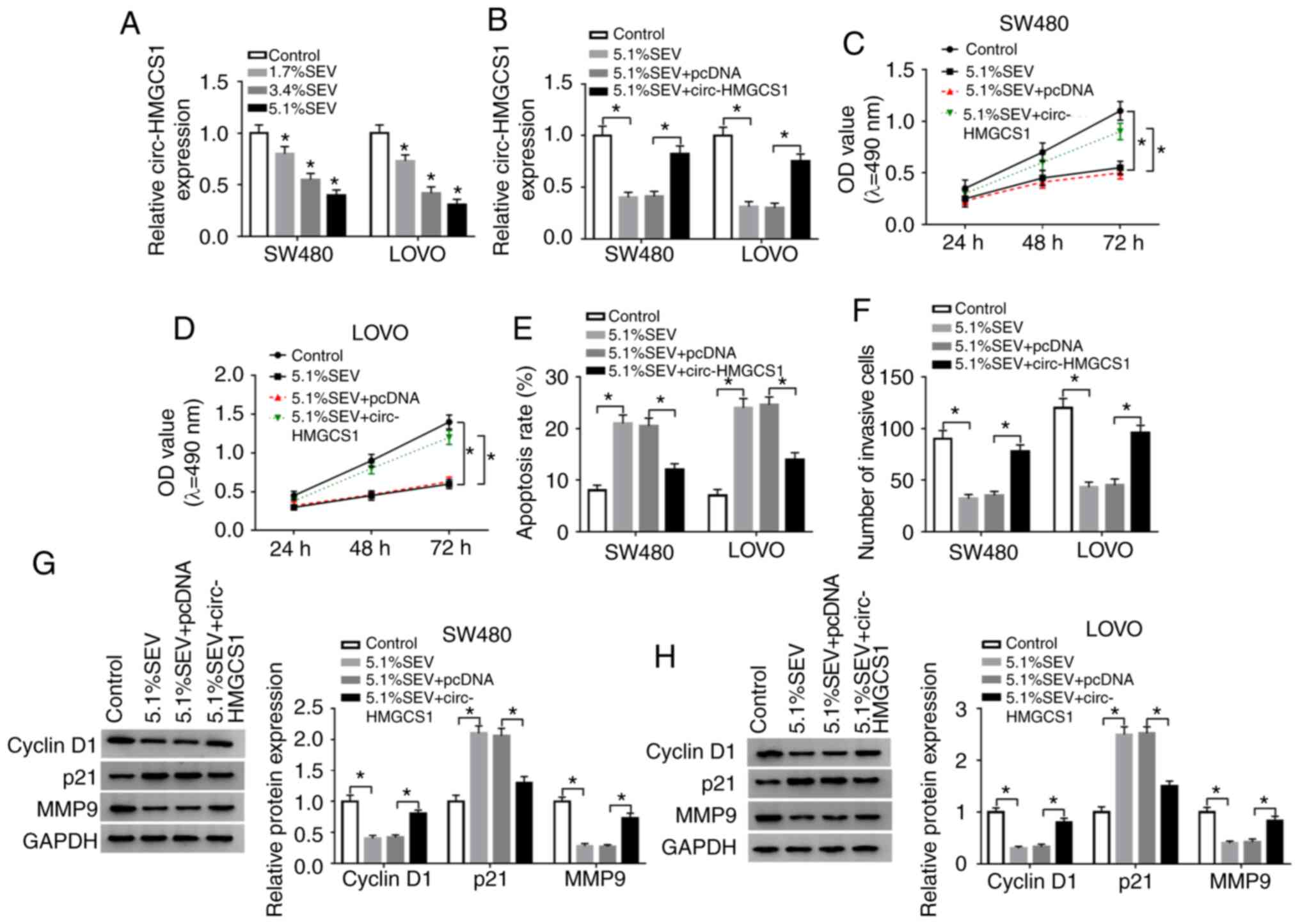
Overexpression of circ-HMGCS1
abrogates the effects of sevoflurane on cell viability, apoptosis
and invasion in colon cancer cells
In order to confirm whether circ-HMGCS1 was involved
in the progression of colon cancer cells, SW480 and LOVO cells were
treated with various concentrations (1.7, 3.4 and 5.1%) of
sevoflurane. Then the effect of sevoflurane on circ-HMGCS1
expression was determined by RT-qPCR. The data revealed that
sevoflurane treatment led to a significant reduction of circ-HMGCS1
in a dose-dependent manner in SW480 and LOVO cells (Fig. 3A). SW480 and LOVO cells exposed to a
concentration of 5.1% SEV were used for the subsequent functional
experiments due to the stronger suppression in circ-HMGCS1
expression. Subsequently, SW480 and LOVO cells were transfected
with pcDNA or circ-HMGCS1 and then exposed to 5.1% SEV for 6 h. As
revealed in Fig. 3B, the
downregulation of circ-HMGCS1 in SW480 and LOVO cells caused by
sevoflurane exposure was partly reversed by the administration of
circ-HMGCS1. An MTT assay revealed that the inhibitory effect on
cell viability mediated by sevoflurane was effectively abolished by
the overexpression of circ-HMGCS1 in both SW480 and LOVO cells
(Fig. 3C and D). As revealed by
flow cytometric analysis, the apoptosis of SW480 and LOVO cells was
facilitated by the treatment of sevoflurane, while circ-HMGCS1
overexpression reversed this effect (Fig. 3E). Transwell assay data indicated
that sevoflurane treatment resulted in a significant suppression in
cell invasion in SW480 and LOVO cells, however, circ-HMGCS1
overexpression abolished this suppression (Fig. 3F). Western blot analysis revealed
that sevoflurane treatment decreased the levels of cyclin D1 and
MMP9 and increased the level of p21 in SW480 and LOVO cells,
whereas the effects were rescued by circ-HMGCS1 (Fig. 3G and H). To sum up, circ-HMGCS1
promoted sevoflurane-mediated proliferation and invasion and
suppressed sevoflurane-mediated apoptosis in colon cancer
cells.
Circ-HMGCS1 silencing suppresses cell
viability and invasion and promotes cell apoptosis by targeting
miR-34a-5p in colon cancer cells
Since circ-HMGCS1 and miR-34a-5p were dysregulated
in colon cancer patients, it was theorized that miR-34a-5p may be a
target of circ-HMGCS1. By searching the online website starBase
v2.0, it was revealed that miR-34a-5p contained the complementary
sequences of circ-HMGCS1 (Fig. 4A).
Next, dual-luciferase reporter and RIP assays were conducted. The
dual-luciferase reporter assay revealed that compared to miR-NC and
WT-circ-HMGCS1 co-transfected groups, the luciferase activity in
SW480 and LOVO cells co-transfected with miR-34a-5p and
WT-circ-HMGCS1 was inhibited, whereas no change was observed in
MUT-circ-HMGCS1 groups (Fig. 4B and
C). The RIP assay revealed that miR-34a-5p and circ-HMGCS1
combined to an Ago2 immunoprecipitation complex were both
significantly enriched compared to the IgG group in SW480 and LOVO
cells (Fig. 4D and E). As revealed
in Fig. 4F and G, si-circ-HMGCS1
transfection led to a significant decrease in circ-HMGCS1
expression and a significant increase in miR-34a-5p expression in
SW480 and LOVO cells, while circ-HMGCS1 transfection exhibited the
opposite results. To further explore the association between
circ-HMGCS1 and miR-34a-5p in the progression of colon cancer
cells, SW480 and LOVO cells were transfected with si-NC,
si-circ-HMGCS1, si-circ-HMGCS1+anti-miR-NC or
si-circ-HMGCS1+anti-miR-34a-5p. It was revealed that the
upregulation of miR-34a-5p caused by circ-HMGCS1 silencing was
reversed following the inhibition of miR-34a-5p (Fig. 4H). Furthermore, circ-HMGCS1
deficiency resulted in a significant suppression in cell viability
and invasion and a significant promotion in cell apoptosis, while
these influences were all weakened by miR-34a-5p inhibition in
SW480 and LOVO cells (Fig. 4I-L).
In addition, the levels of cyclin D1 and MMP9 were reduced and the
level of p21 was increased in SW480 and LOVO cells transfected with
si-circ-HMGCS1; however, inhibitors of miR-34a-5p abolished the
effects (Fig. 4M and N).
Collectively, miR-34a-5p inhibition alleviated the inhibitory
effect of circ-HMGCS1 knockdown on colon cancer cell
progression.
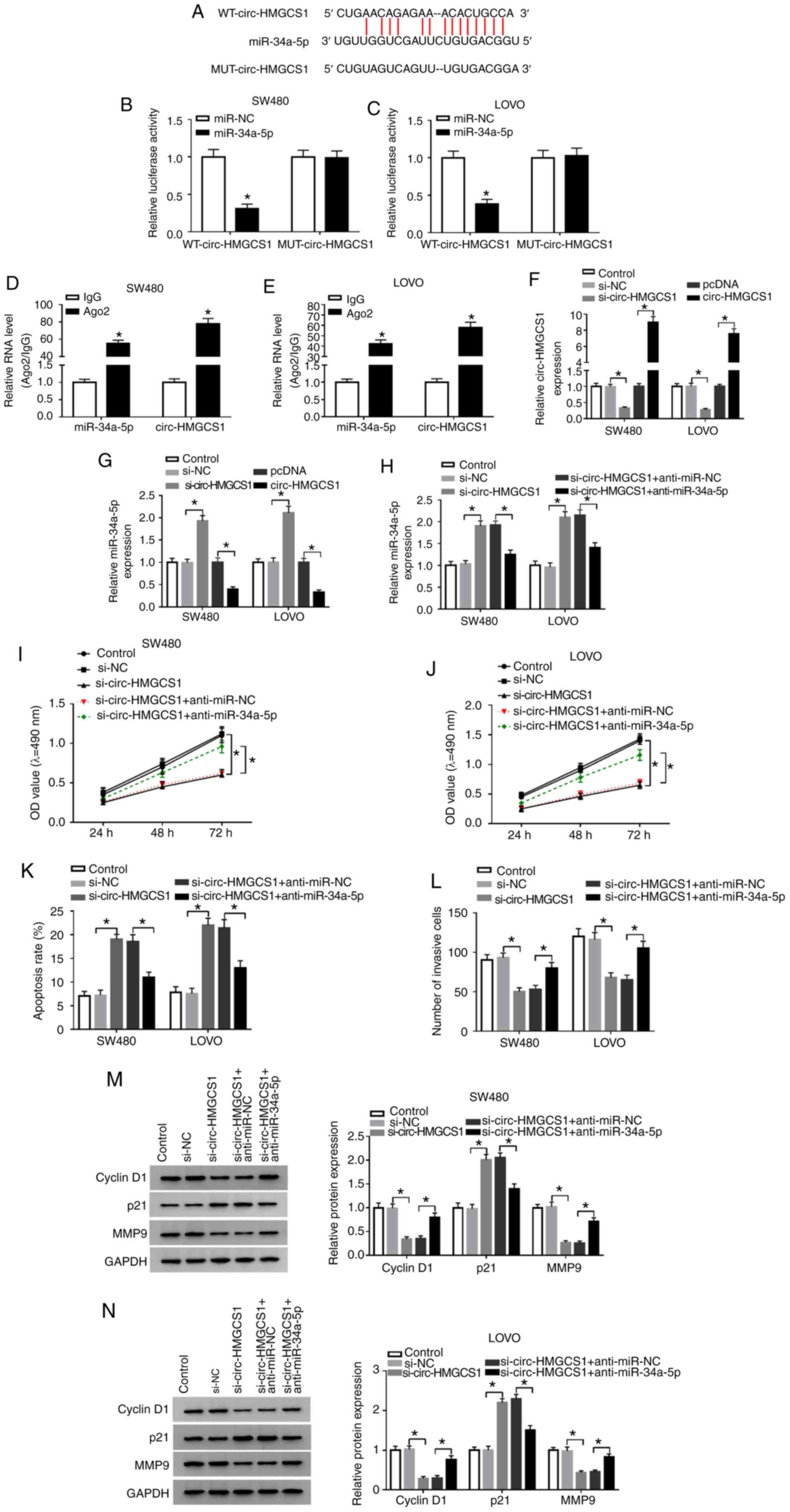 | Figure 4.miR-34a-5p downregulation reverses
the effects of circ-HMGCS1 silencing on cell viability, apoptosis
and invasion in colon cancer cells. (A) The potential binding
sequences between circ-HMGCS1 and miR-34a-5p. (B and C) A
dual-luciferase reporter assay was conducted to verify the
association between circ-HMGCS1 and miR-34a-5p. *P<0.05 vs. the
miR-NC group. (D and E) The enrichment of miR-34a-5p and
circ-HMGCS1 combined to Ago2/IgG precipitation complexes was
determined by RT-qPCR. *P<0.05 vs. the IgG group. (F and G) The
levels of circ-HMGCS1 and miR-34a-5p in SW480 and LOVO cells
transfected with si-NC, si-circ-HMGCS1, pcDNA or circ-HMGCS1 were
measured by RT-qPCR. *P<0.05 vs. the si-NC group or the pcDNA
group. (H) SW480 and LOVO cells were divided into 5 groups:
Control, si-NC, si-circ-HMGCS1, si-circ-HMGCS1+anti-miR-NC and
si-circ-HMGCS1+anti-miR-34a-5p. (H) The expression of miR-34a-5p
was measured by RT-qPCR. *P<0.05 vs. the si-NC group or the
si-circ-HMGCS1+anti-miR-NC group. (I and J) Cell viability, (K)
apoptosis and (L) invasion were evaluated by MTT assay, flow
cytometric analysis and Transwell assay, respectively. (M and N)
The levels of cyclin D1, p21 and MMP9 in SW480 and LOVO cells were
examined using western blotting. *P<0.05 vs. the si-NC group or
the si-circ-HMGCS1+anti-miR-NC group. miR, microRNA; circ-HMGCS1,
circular RNA 3-hydroxy-3-methylglutaryl-CoA synthase 1; Ago2,
Argonaute2; IgG, immunoglobulin G; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; si, small
interfering; NC, negative control; MMP9, matrix metallopeptidase 9;
WT, wild-type; MUT, mutated. |
miR-34a-5p inhibition alleviates
sevoflurane-mediated effects on cell viability, apoptosis and
invasion in colon cancer cells
To further reveal the underlying mechanism of
sevoflurane in the development of colon cancer, the effect of
sevoflurane on miR-34a-5p expression was investigated. The data
revealed that miR-34a-5p was significantly increased following the
treatment of sevoflurane in a dose-dependent manner (Fig. 5A). Then anti-miR-34a-5p or
anti-miR-NC was transfected into SW480 and LOVO cells and then
these cells were exposed to 5.1% SEV. It was observed that
sevoflurane-mediated upregulation in miR-34a-5p was partially
reversed by the transfection of anti-miR-34a-5p (Fig. 5B). Moreover, the suppressive effects
of sevoflurane on cell viability (Fig.
5C and D) and cell invasion (Fig.
5F) and the promoting effect on cell apoptosis (Fig. 5E) were all weakened following
miR-34a-5p inhibition in SW480 and LOVO cells. As indicated by
western blot analysis, miR-34a-5p inhibition effectively rescued
the reduction in cyclin D1 and MMP9 expression and the increase in
p21 expression caused by sevoflurane in SW480 and LOVO cells
(Fig. 5G and H). These data
indicated that the influence of sevoflurane on cell progression was
restored by miR-34a-5p depletion in colon cancer.
 | Figure 5.Effects of sevoflurane on cell
viability, apoptosis and invasion are reversed by miR-34a-5p
inhibition in colon cancer cells. (A) SW480 and LOVO cells were
exposed to various concentrations (1.7, 3.4 and 5.1%) of
sevoflurane and then miR-34a-5p expression was detected by RT-qPCR.
(B-H) SW480 and LOVO cells were divided into 4 groups: Control,
5.1% SEV, 5.1% SEV+anti-miR-NC and 5.1% SEV+anti-miR-34a-5p. (B)
miR-34a-5p expression in SW480 and LOVO cells was measured by
RT-qPCR. (C and D) The viability, (E) apoptosis and (F) invasion of
SW480 and LOVO cells were analyzed via MTT assay, flow cytometric
analysis and Transwell assay, respectively. (G and H) The protein
levels of cyclin D1, p21 and MMP9 were determined using western
blotting. *P<0.05 vs. the control group or the 5.1%
SEV+anti-miR-NC group. miR, microRNA; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; SEV,
sevoflurane; NC, negative control; MMP9, matrix metallopeptidase 9;
OD, optical density. |
SGPP1 is a target gene of
miR-34a-5p
By further searching online software starBase v2.0,
it was revealed that the 3′UTR of SGPP1 contained the putative
binding sites of miR-34a-5p (Fig.
6A). Dual-luciferase reporter assays revealed that
co-transfection of miR-34a-5p and SGPP1 3′UTR-WT led to a
significant suppression in the luciferase activity in SW480 and
LOVO cells compared with miR-NC and SGPP1 3′UTR-WT co-transfected
cells, while the luciferase activity was not affected in the SGPP1
3′UTR-MUT groups (Fig. 6B and C).
RIP assays indicated that miR-34a-5p and SGPP1 levels were
increased in SW480 and LOVO cells after Ago2 RIP compared to the
IgG control group (Fig. 6D and E).
As anticipated, the mRNA and protein levels of SGPP1 in the serums
of colon cancer patients were markedly increased compared to those
in the serums of healthy volunteers (Fig. 6F and G). Similarly, the protein
level of SGPP1 was increased in SW480 and LOVO cells compared to
NCW460 cells (Fig. 6H). Moreover,
SGPP1 expression was negatively correlated with miR-34a-5p
expression in the serums of colon cancer patients (Fig. 6I). As revealed in Fig. 6J-L, miR-34a-5p transfection
significantly enhanced miR-34a-5p expression and decreased SGPP1
protein expression in SW480 and LOVO cells, while anti-miR-34a-5p
transfection exhibited the opposite results. Collectively,
miR-34a-5p negatively modulated SGPP1 expression by directly
targeting SGPP1 in colon cancer cells.
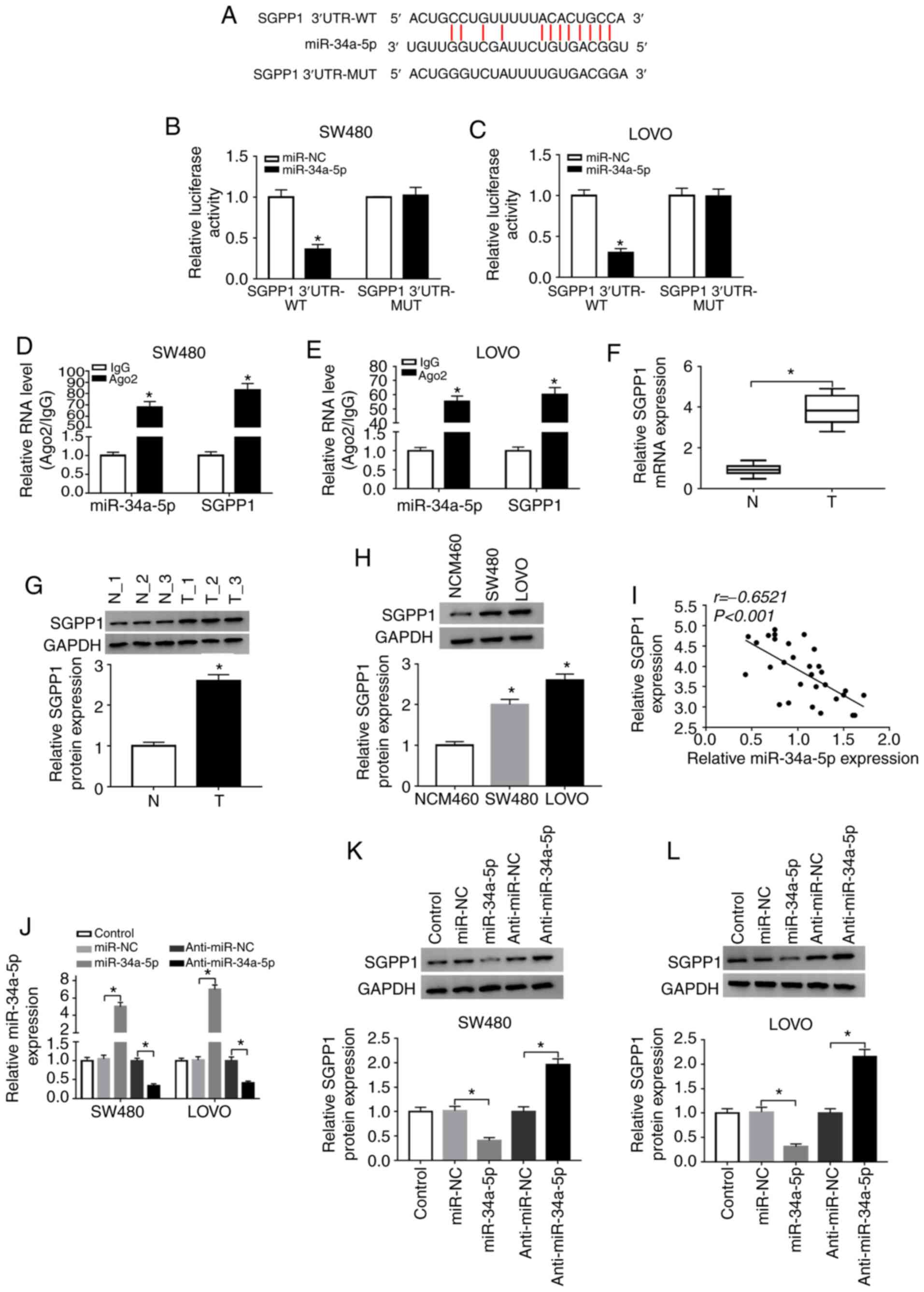 | Figure 6.miR-34a-5p directly interacts with
SGPP1 and negatively regulates SGPP1 expression. (A) The predicted
binding sites between miR-34a-5p and SGPP1. (B and C) The
luciferase activity in miR-NC or miR-34a-5p and SGPP1 3′UTR-WT or
SGPP1 3′UTR-MUT co-transfected SW480 and LOVO cells was measured by
dual-luciferase reporter assay. *P<0.05 vs. the miR-NC group. (D
and E) The levels of miR-34a-5p and SGPP1 in Ago2 or IgG
immunoprecipitation complexes in SW480 and LOVO cells were detected
by RT-qPCR assay. *P<0.05 vs. the IgG group. (F and G) RT-qPCR
assay and western blotting were conducted to examine the mRNA and
protein levels of SGPP1 in the serums collected from colon cancer
patients and healthy participants, respectively. *P<0.05 vs. the
normal group. (H) The protein level of SGPP1 in NCM460, SW480 and
LOVO cells was examined by western blotting. *P<0.05 vs. the
NCM460 group. (I) The correlation between SGPP1 and miR-34a-5p was
analyzed by Spearman's correlation coefficient analysis. (J-L) The
expression of miR-34a-5p and SGPP1 in SW480 and LOVO cells
transfected with miR-NC, miR-34a-5p, anti-miR-NC or anti-miR-34a-5p
was detected by RT-qPCR and western blotting, respectively.
*P<0.05 vs. miR-NC or the anti-miR-NC group. miR, microRNA;
SGPP1, sphingosine-1-phosphate phosphatase 1; NC, negative control;
WT, wild-type; MUT, mutated; Ago2, Argonaute2; IgG, immunoglobulin
G; RT-qPCR, reverse transcription-quantitative polymerase chain
reaction. |
Sevoflurane suppresses SGPP1
expression via the circ-HMGCS1/miR-34a-5p axis in colon cancer
cells
To investigate the relationship among sevoflurane,
circ-HMGCS1, miR-34a-5p and SGPP1 in colon cancer cells, SW480 and
LOVO cells were transfected with si-NC, si-circ-HMGCS1,
si-circ-HMGCS1+anti-miR-NC or si-circ-HMGCS1+anti-miR-34a-5p. The
data revealed that circ-HMGCS1 knockdown decreased the expression
of SGPP1 in both SW480 and LOVO cells, while miR-34a-5p inhibition
partly restored the effect, indicating that circ-HMGCS1 could
suppress SGPP1 expression via sponging miR-34a-5p (Fig. 7A and C). In addition, it was
revealed that sevoflurane treatment caused a significant decrease
in SGPP1 expression in SW480 and LOVO cells, however circ-HMGCS1
overexpression reversed the decrease (Fig. 7B and D). Collectively the results
demonstrated that sevoflurane could modulate SGPP1 expression via
circ-HMGCS1/miR-34a-5p axis.
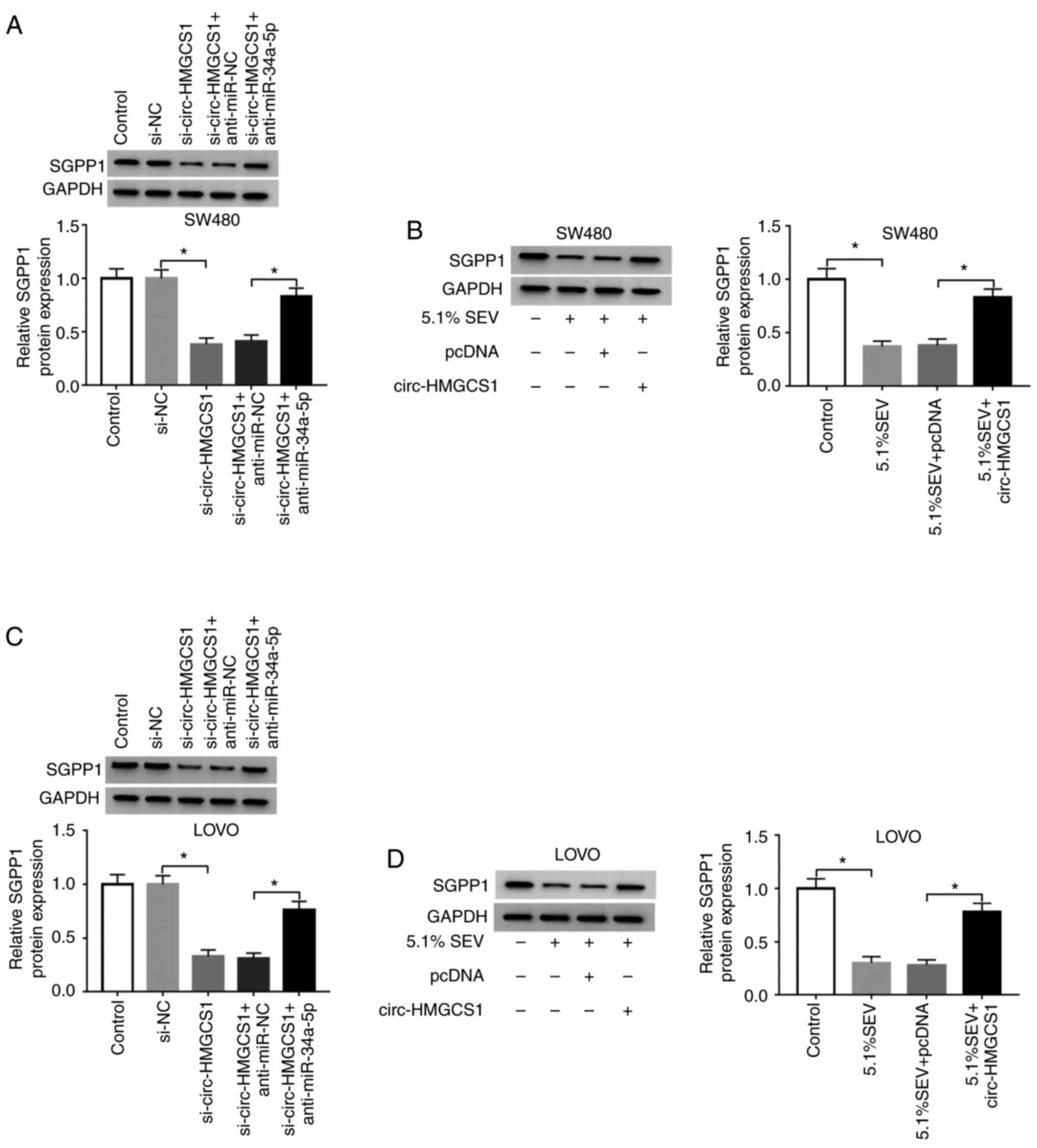 | Figure 7.Sevoflurane decreases SGPP1
expression by regulating the circ-HMGCS1/miR-34a-5p axis in colon
cancer cells. (A and C) SW480 and LOVO cells were assigned to
Control, si-NC, si-circ-HMGCS1, si-circ-HMGCS1+anti-miR-NC and
si-circ-HMGCS1+anti-miR-34a-5p groups. *P<0.05 vs. si-NC or
si-circ-HMGCS1+anti-miR-NC. (B and D) SW480 and LOVO cells were
assigned to Control, 5.1% SEV, 5.1% SEV+pcDNA and 5.1%
SEV+circ-HMGCS1 groups. *P<0.05 vs. the control group or the
5.1% SEV+pcDNA group. (A-D) SGPP1 protein levels in SW480 and LOVO
cells was analyzed using western blotting. circ-HMGCS1, circular
RNA 3-hydroxy-3-methylglutaryl-CoA synthase 1; miR, microRNA; si,
small interfering; NC, negative control; SEV, sevoflurane; SGPP1,
sphingosine-1-phosphate phosphatase 1. |
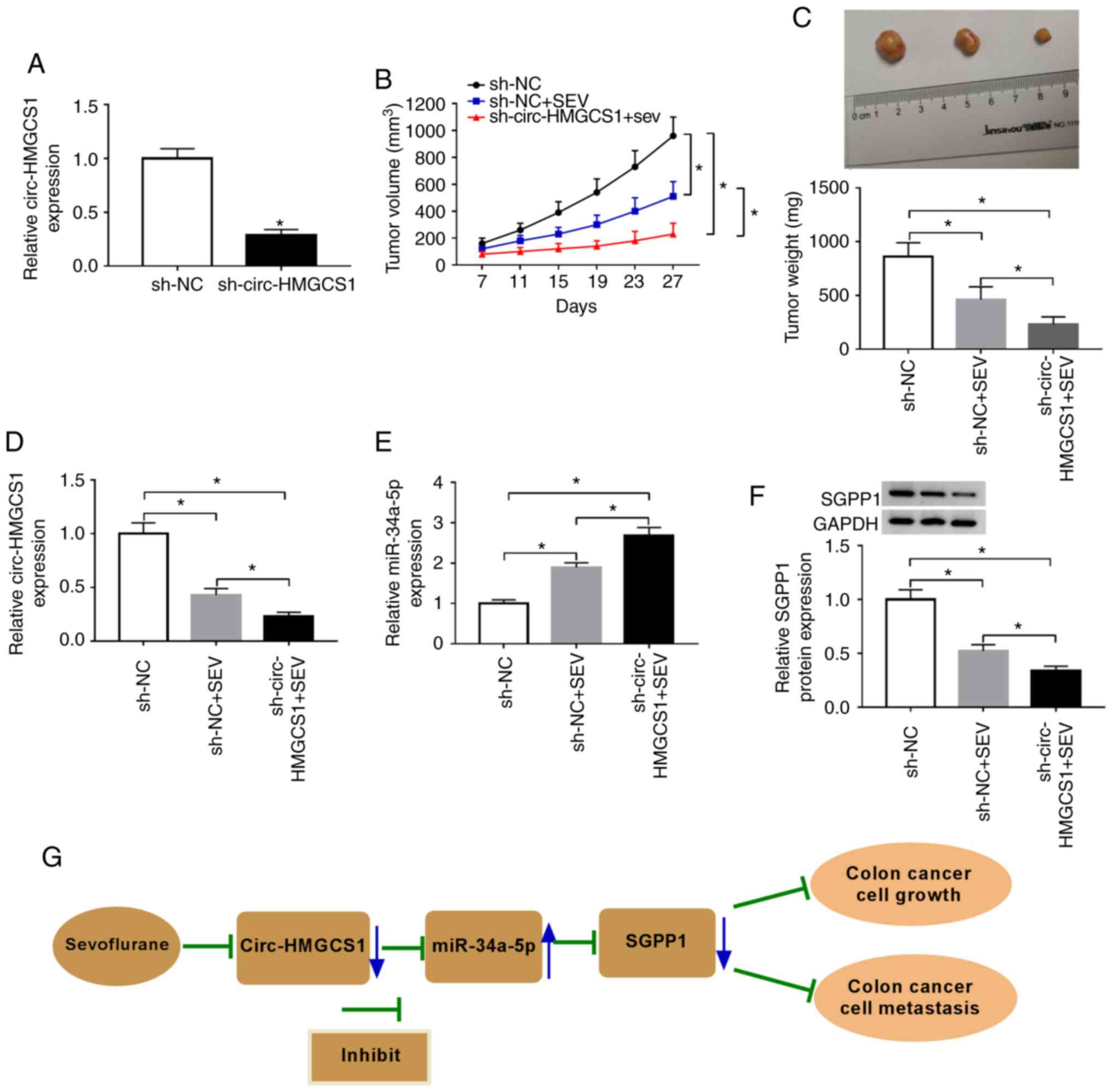
Circ-HMGCS1 knockdown suppresses
tumorigenesis of colon cancer in vivo
To reveal the effect of circ-HMGCS1 in tumor
progression in vivo, SW480 cells were stably transfected
with sh-circ-HMGCS1 or sh-NC and then stimulated with sevoflurane.
As revealed in Fig. 8A,
sh-circ-HMGCS1 transfection significantly reduced the expression
level of circ-HMGCS1 in SW480 cells compared to the sh-NC group,
indicating the successful transfection of sh-circ-HMGCS1. Then,
SW480 cells were injected into the nude mice to establish a murine
xenograft model. It was revealed that the tumor volume and tumor
weight were decreased by circ-HMGCS1 knockdown (Fig. 8B and C). Subsequently, the levels of
circ-HMGCS1, miR-34a-5p and SGPP1 were determined in the collected
tumors. The data revealed that circ-HMGCS1 and SGPP1 levels were
significantly decreased and the level of miR-34a-5p was
significantly increased in the tumors collected from the
sh-circ-HMGCS+SEV group compared to that in the sh-NC+SEV group
(Fig. 8D-F). These results
demonstrated that silencing of circ-HMGCS1 could block tumor growth
of colon cancer in vivo.
Based on all the experimental results, we arrived at
the conclusion that sevoflurane treatment could inhibit the
circHMGCS1/miR-34a-5p/SGPP1 axis, thereby suppressing colon cancer
cell growth and metastasis (Fig.
8G).
Discussion
Emerging evidence has revealed that anesthetic
techniques or drugs can affect the development of human cancers
(26). In the present study, the
function and underlying mechanisms of sevoflurane in colon cancer
progression were explored and it was demonstrated that sevoflurane
suppressed colon cancer cell viability and invasion and facilitated
apoptosis via regulating the circ-HMGCS1/miR-34a-5p/SGPP1 axis.
Sevoflurane exerts a tumor-suppressive role in colon
cancer, as demonstrated by former studies. For example, Yang et
al revealed that sevoflurane suppressed cell viability and
motility and induced cell apoptosis and autophagy in colon cancer
(27). Fan et al
demonstrated that sevoflurane led to an evident suppression of CRC
cell metastasis in a dose-dependent manner (28). Consistently, we observed that
sevoflurane hindered colon cancer cell viability and invasion and
contributed to apoptosis in a concentration-dependent manner.
Subsequently, the potential mechanisms of
sevoflurane in colon cancer were explored. It was determined that
exosomal circ-HMGCS1 was increased in the serums of colon cancer
patients and colon cancer cells. Moreover, cytoplasm circ-HMGCS1
was increased in colon cancer cells. These data indicated that
circ-HMGCS1 may be a diagnostic and prognostic biomarker for
patients with colon cancer. Furthermore, the effect of sevoflurane
on circ-HMGCS1 expression was explored and it was revealed that
circ-HMGCS1 was suppressed by sevoflurane treatment in a
dose-dependent manner. Zhen et al demonstrated that
circ-HMGCS1 was significantly increased in HB and circ-HMGCS1
deficiency hindered cell growth and facilitated cell apoptosis
in vitro and blocked tumorigenesis in vivo (15). Dong et al revealed that the
increase of circ-HMGCS1 predicted a poor prognosis and circ-HMGCS1
silencing inhibited cell growth and promoted apoptosis in CRC
(16). In the present study, the
effects on cell viability, apoptosis and invasion mediated by
sevoflurane were abolished by the overexpression of circ-HMGCS1 in
colon cancer, indicating that circ-HMGCS1 overexpression could
enhance colon cancer cell viability and invasion and suppress
apoptosis. Moreover, circ-HMGCS1 silencing could hinder tumor
growth in vivo. All these data demonstrated that
exosome-transmitted circ-HMGCS1 functioned as an oncogene in colon
cancer.
CircRNAs contain the binding sites of miRNAs and act
as sponges of miRNAs to regulate gene transcription (29,30).
Herein, miR-34a-5p was identified as a direct target of
circ-HMGCS1. Gao et al revealed that miR-34a-5p was
downregulated in CRC, and ectopic expression of miR-34a-5p
suppressed cell growth and metastasis and induced apoptosis in CRC
(21). Sun et al
demonstrated that miR-34a inhibitors restored the inhibitory
effects of sevoflurane on CRC cell viability and metastasis by
binding to ADAM10 (31). In line
with these data, we revealed that the suppressive roles of
circ-HMGCS1 silencing on cell viability and invasion and the
promoting role of circ-HMGCS1 silencing in cell apoptosis in colon
cancer were all reversed by miR-34a-5p inhibition. Moreover,
miR-34a-5p inhibition could reverse the inhibition in the
progression of colon cancer cells caused by sevoflurane treatment.
In addition, it was revealed that SGPP1 was a direct target gene of
miR-34a-5p. Gao et al demonstrated that SGPP1 was increased
in CRC tissues and cells, and miR-27a could target SGPP1 to hinder
cell growth and migration and facilitate cell apoptosis in CRC
(21). In the present study, SGPP1
was increased in colon cancer patients and cells and SGPP1 was
negatively modulated by miR-34a-5p. Moreover, it was revealed that
circ-HMGCS1 could promote SGPP1 expression through sponging
miR-34a-5p.
In summary, sevoflurane hindered cell viability and
invasion and facilitated cell apoptosis in colon cancer by
regulating the exosome-transmitted circ-HMGCS1/miR-34a-5p/SGPP1
axis. These findings facilitated our understanding of sevoflurane
on colon cancer progression and may provide an experimental basis
for selecting more reasonable anesthetics for patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
Conceptualization and methodology was performed by
JH, HZ and XL. DW, YW and YA performed the formal analysis and data
curation. JH and JY conducted the validation and investigation. JH,
HZ, XL and DW drafted the manuscript, wrote, reviewed and edited
the manuscript. All authors approved the final manuscript and
agreed be accountable for all aspects of the work in ensuring that
questions related to the accuracy or integrity of any part of the
work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics Review
Committee of The First Affiliated Hospital of Zhengzhou University.
Written informed consent was obtained from all enrolled patients.
The animal experiments were approved by the Ethics Committee of
Animal Research of the First Affiliated Hospital of Zhengzhou
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ,
Meester RGS, Barzi A and Jemal A: Colorectal cancer statistics,
2017. CA Cancer J Clin. 67:177–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moriarity A, O'Sullivan J, Kennedy J,
Mehigan B and McCormick P: Current targeted therapies in the
treatment of advanced colorectal cancer: A review. Ther Adv Med
Oncol. 8:276–293. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vasile L, Olaru A, Munteanu M, Plesea IE,
Surlin V and Tudorascu C: Prognosis of colorectal cancer: Clinical,
pathological and therapeutic correlation. Rom J Morphol Embryol.
53:383–391. 2012.PubMed/NCBI
|
|
6
|
Green JS and Tsui BC: Impact of anesthesia
for cancer surgery: Continuing professional development. Can J
Anaesth. 60:1248–1269. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liang H, Gu M, Yang C, Wang H, Wen X and
Zhou Q: Sevoflurane inhibits invasion and migration of lung cancer
cells by inactivating the p38 MAPK signaling pathway. J Anesth.
26:381–392. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gao C, Shen J, Meng ZX and He XF:
Sevoflurane Inhibits Glioma Cells Proliferation and Metastasis
through miRNA-124- 3p/ROCK1 Axis. Pathol Oncol Res. 26:947–954.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun Z, Shi K, Yang S, Liu J, Zhou Q, Wang
G, Song J, Li Z, Zhang Z and Yuan W: Effect of exosomal miRNA on
cancer biology and clinical applications. Mol Cancer. 17:1472018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sundararajan V, Sarkar FH and Ramasamy TS:
The multifaceted role of exosomes in cancer progression: Diagnostic
and therapeutic implications [corrected]. Cell Oncol (Dordr).
41:223–252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nedaeinia R, Manian M, Jazayeri MH,
Ranjbar M, Salehi R, Sharifi M, Mohaghegh F, Goli M, Jahednia SH,
Avan A and Ghayour-Mobarhan M: Circulating exosomes and exosomal
microRNAs as biomarkers in gastrointestinal cancer. Cancer Gene
Ther. 24:48–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li X, Yang L and Chen LL: The biogenesis,
functions, and challenges of circular RNAs. Mol Cell. 71:428–442.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Q, Zhang C, Ma JX, Ren H, Sun Y and
Xu JZ: Circular RNA PIP5K1A promotes colon cancer development
through inhibiting miR-1273a. World J Gastroenterol. 25:5300–5309.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu XW, Zheng BA, Hu ZM, Qian ZY, Huang CJ,
Liu XQ and Wu WD: Circular RNA hsa_circ_000984 promotes colon
cancer growth and metastasis by sponging miR-106b. Oncotarget.
8:91674–91683. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhen N, Gu S, Ma J, Zhu J, Yin M, Xu M,
Wang J, Huang N, Cui Z, Bian Z, et al: CircHMGCS1 promotes
hepatoblastoma cell proliferation by regulating the IGF signaling
pathway and glutaminolysis. Theranostics. 9:900–919. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dong J, Li J, Luo J and Wu W: CircHMGCS1
is upregulated in colorectal cancer and promotes proliferation of
colorectal cancer cells by targeting microRNA-503-5p. Eur J Inflam.
17:1–11. 2019. View Article : Google Scholar
|
|
17
|
Reddy KB: MicroRNA (miRNA) in cancer.
Cancer Cell Int. 15:382015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luan XF, Wang L and Gai XF: The
miR-28-5p-CAMTA2 axis regulates colon cancer progression via
Wnt/beta-catenin signaling. J Cell Biochem. Nov 10–2019.(Epub ahead
of print). doi: 10.1002/jcb.29536. View Article : Google Scholar
|
|
19
|
Chai B, Guo Y, Cui X, Liu J, Suo Y, Dou Z
and Li N: MiR-223-3p promotes the proliferation, invasion and
migration of colon cancer cells by negative regulating PRDM1. Am J
Transl Res. 11:4516–4523. 2019.PubMed/NCBI
|
|
20
|
Xi X, Teng M, Zhang L, Xia L, Chen J and
Cui Z: MicroRNA-204-3p represses colon cancer cells proliferation,
migration, and invasion by targeting HMGA2. J Cell Physiol.
235:1330–1338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao J, Li N, Dong Y, Li S, Xu L, Li X, Li
Y, Li Z, Ng SS, Sung JJ, et al: miR-34a-5p suppresses colorectal
cancer metastasis and predicts recurrence in patients with stage
II/III colorectal cancer. Oncogene. 34:4142–4152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kara M, Yumrutas O, Ozcan O, Celik OI,
Bozgeyik E, Bozgeyik I and Tasdemir S: Differential expressions of
cancer-associated genes and their regulatory miRNAs in colorectal
carcinoma. Gene. 567:81–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bao Y, Chen Z, Guo Y, Feng Y, Li Z, Han W,
Wang J, Zhao W, Jiao Y, Li K, et al: Tumor suppressor microRNA-27a
in colorectal carcinogenesis and progression by targeting SGPP1 and
Smad2. PLoS One. 9:e1059912014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lässer C, Eldh M and Lötvall J: Isolation
and characterization of RNA-containing exosomes. J Vis Exp.
e30372012.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Niwa H, Rowbotham DJ, Lambert DG and Buggy
DJ: Can anesthetic techniques or drugs affect cancer recurrence in
patients undergoing cancer surgery? J Anesth. 27:731–741. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang X, Zheng YT and Rong W: Sevoflurane
induces apoptosis and inhibits the growth and motility of colon
cancer in vitro and in vivo via inactivating Ras/Raf/MEK/ERK
signaling. Life Sci. 239:1169162019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fan L, Wu Y, Wang J, He J and Han X:
Sevoflurane inhibits the migration and invasion of colorectal
cancer cells through regulating ERK/MMP-9 pathway by up-regulating
miR-203. Eur J Pharmacol. 850:43–52. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Militello G, Weirick T, John D, Doring C,
Dimmeler S and Uchida S: Screening and validation of lncRNAs and
circRNAs as miRNA sponges. Brief Bioinform. 18:780–788.
2017.PubMed/NCBI
|
|
30
|
Rong D, Sun H, Li Z, Liu S, Dong C, Fu K,
Tang W and Cao H: An emerging function of circRNA-miRNAs-mRNA axis
in human diseases. Oncotarget. 8:73271–73281. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun SQ, Ren LJ, Liu J, Wang P and Shan SM:
Sevoflurane inhibits migration and invasion of colorectal cancer
cells by regulating microRNA-34a/ADAM10 axis. Neoplasma.
66:887–895. 2019. View Article : Google Scholar : PubMed/NCBI
|