Introduction
Head and neck cancer (HNC) frequently infiltrates
and metastasizes bones (1),
resulting in HNC-associated bone pain (HNC-BP) (2). Cancer pain is a prognostic factor of
poor clinical outcomes (3). HNC-BP
often reduces the ability to eat and swallow, thus posing a
significant challenge to the quality of life of patients presenting
with advanced HNC. The pathophysiology of bone pain associated with
HNC is poorly understood (4), and
HNC-BP is frequently inadequately treated.
Dead cells, dendritic cells, and cancer cells
releases damage-associated molecular patterns (DAMPs), which
initiate noninfectious inflammatory responses (5). Among the various DAMPs, high mobility
group box 1 (HMGB1) was first discovered as a conserved non-histone
DNA-binding protein and has been the most intensively studied
(6). A 2015 study revealed that
HMGB1 mediates inflammatory and immune reactions in the nervous
system (7), and emerging evidence
indicates that HMGB1 plays an essential role in neuropathic pain
(8). Extracellular HMGB1 activates
the receptor for advanced glycation end products (RAGE) and
Toll-like receptor 4 (TLR4), promoting pain signals (9,10). It
was revealed that HMGB1 is overexpressed in the serum and tissues
in several types of cancer, including breast cancer, malignant
mesothelioma, and other malignancies (11,12).
However, the role of cancer-derived HMGB1 in HNC-BP remains
unknown.
Cancer cells in bone metastasis and at bone invasion
sites secrete significant amounts of growth factors, promoting
osteoclastogenesis (13,14). The best-known molecular pain inducer
in HNC-BP is osteoclast-derived proton (H+) (15). In the cancer bone microenvironment,
cancer cells and osteoclasts secrete protons through the proton
pump, creating an acidic microenvironment in the bone marrow
(16). This environment activates
pH-sensitive sensory neurons via acid-sensing nociceptors such as
transient receptor potential vanilloid 1 (TRPV1), eliciting bone
pain (17). The findings of the
present study provide initial evidence that HNC-derived HMGB1
directly activates sensory neurons and increases the expression of
TRPV1, thus indirectly promoting acid-induced HNC-BP.
Materials and methods
Reagents
Puromycin dihydrochloride (#P9620) was purchased
from Sigma-Aldrich; Merck KGaA. TLR4 antagonist TAK-242 (#13871)
and RAGE antagonist FPS-ZM1 (#11909) were purchased from Cayman
Chemical Co. Chicken anti-HMGB1 polyclonal antibody (#326052233)
was purchased from Shino-Test. Control shRNA plasmid-A
(#sc-108060), HMG-1 shRNA plasmid (#sc-37982-SH), and anti-RAGE
antibody (anti-mouse, monoclonal, sc-80652) were purchased from
Santa Cruz Biotechnology, Inc. HMGB1 antibody (anti-mouse,
monoclonal, GTX628834) was purchased from GeneTex, Inc.
Anti-phospho-p44/42 MAPK antibody (p-ERK; anti-rabbit, monoclonal,
#4370), anti-p44/42 MAPK antibody (ERK; anti-rabbit, monoclonal,
#4695), horseradish peroxidase (HRP)-conjugated IgG antibody (goat
anti-rabbit, monoclonal, #7074), HRP-conjugated IgG antibody (goat
anti-mouse, monoclonal, #7076), and Alexa Fluor 488-conjugated IgG
(H+L) F(ab′)2 fragment (goat anti-rabbit, monoclonal, #4412) were
purchased from Cell Signaling Technology, Inc. Anti-TLR4 antibody
(anti-rabbit, polyclonal, #ab13556), anti-β-actin antibody
(anti-mouse, monoclonal, #ab49900), anti-CGRP antibody (anti-goat,
polyclonal, #ab36001), and Alexa Fluor 647-conjugated IgG H&L
antibody (donkey anti-goat, monoclonal, #ab150135) were purchased
from Abcam.
Cell lines and culture conditions
The human HNC cell lines SAS (#JCRB0260), HSC-2
(#JCRB0622), HSC-3 (#JCRB0623), HSC-4 (#JCRB0624), OSC-19
(#JCRB0198), and mouse fibroblast cell line 3T6 (#JCRB9059) were
obtained from the Human Science Resources Bank (Osaka, Japan). All
of these cell lines were cultured in Dulbecco's modified Eagle's
medium (DMEM) (Thermo Fisher Scientific, Inc.) supplemented with
10% heat-inactivated fetal bovine serum (FBS) and 1%
penicillin-streptomycin. The rat DRG neuronal cell line 50B11 was
kindly provided by Dr A. Hoke (Department of Neurology, School of
Medicine, Johns Hopkins University, Baltimore, MD, USA) and
cultured in neurobasal medium. MLO-A5 cells provided by Dr T.
Bellido (Department of Anatomy and Cell Biology, School of
Medicine, Indiana University, Indianapolis, IN, USA) and primary
mouse osteoblast cells (OBC12) (Cosmo Bio, Tokyo) were cultured in
α-MEM (minimal essential medium) containing 5% FBS. Cells of the
mouse macrophage cell line RAW264.7 were cultured in α-MEM
containing 10% FBS.
Rat DRG cells were obtained from Lonza Japan and
cultured according to the supplier's instructions. All cell lines
were cultured in an atmosphere of 5% CO2 at 37°C.
Immunohistochemical analysis
We analyzed the expression of HMGB1 in HNC tissue
and a normal tissue microarray (#OR601d; US Biomax). The antigen
was activated by cooking in a citric acid solution. For the
immunohistochemical analysis, the specimens were incubated with
anti-HMGB1 antibody (dilution 1:200) overnight at 4°C. The slides
were then treated with a streptavidin-biotin complex (EnVision
System labeled polymer, HRP; Dako; Agilent Technologies, Inc.) for
60 min at a dilution of 1:100. The immunoreaction was visualized
with the use of a DAB substrate-chromogen solution (Dako Cytomation
Liquid DAB Substrate Chromogen System, Agilent Technologies, Inc.).
The cells were counted using a light microscope and evaluated.
Analysis of HMGB1 expression in SAS
cells
SAS cells were transfected with 5.0 µg control short
hairpin (sh)RNA or HMGB1 shRNA with the use of 4D-Nucleofector™
(Lonza Group, Ltd.). Two days later, the cells were cultured in
DMEM + 10% FBS for 5 days in the presence of 1.6 µg/ml puromycin
dihydrochloride for the selection of cells that stably expressed
the shRNAs.
The hairpin sequence was as follows:
GATCCAAGCACCCAGATGCTTCAGTTTCAAGAGAACTGAAGCATCTGGGTGCTTTTTTT. The
corresponding siRNA sequences were sense, AAGCACCCAGAUGCUUCAGUtt
and antisense, ACUGAAGCAUCUGGGUGCUUtt. All sequences are provided
in 5′→3′ orientation.
Cell proliferation assay
SAS cells were plated in 24-well plates at
1.5×104 cells per well. Their number was counted 72 h
later with a TC20 automated cell counter according to the
manufacturer's instructions (Bio-Rad Laboratories, Inc.).
Western blot analysis
The cell lysates were mixed with 4X Laemmli sample
buffer (Bio-Rad Laboratories, Inc.) and heated at 95°C for 5 min.
The samples were electrophoresed on 4–12% SDS-PAGE gels, and the
proteins were transferred onto PVDF membranes (Bio-Rad
Laboratories, Inc.). The membranes were incubated with primary and
secondary antibodies according to the ECL chemiluminescence
protocol (RPN2109; Amersham Biosciences) to detect secondary
antibody binding. Antibodies against HMGB1 (1:1,000), TLR-4
(1:500), RAGE (1:1,000), p-ERK (1:1,000), ERK (1:1,000), TRPV1
(1:1,000), and β-actin (1:10,000) were used as the primary
antibodies. HRP-conjugated anti-rabbit antibody (1:2,000) and
HRP-conjugated anti-mouse antibody (1:2,000) were used as the
secondary antibodies.
We evaluated the effects of three redox forms of
HMGB1 on sensory neuronal excitation. Cells of the DRG neuronal
cell line 50B11 were cultured on 6-well plates
(5×104/well) in DMEM and incubated for 24 h after the
addition of each isoform of HMGB1 (HMGB1 Isoform Kit, #HM-030,
HMGBiotech). The protein was then collected. A ChemiDoc MP system
(Bio-Rad Laboratories, Inc.) was used for the analysis of the
western blots.
Sensory neuron fiber sprouting
assay
DRG cells were plated on 48-well plates
(1×104/well) in neuron growth medium for 24 h. The
conditioned medium of SAS, short hairpin (sh)-control SAS, and
sh-HMGB1 SAS cells was added to each well at 30% of the total
medium volume. The conditioned medium had been collected after the
incubation of 5.0×106 of each cell in DMEM plus 2% FBS
for 24 h.
We also evaluated an HMGB1 neutralizing antibody
which was applied to the wells with conditioned medium of SAS at
the following concentrations: Anti-HMGB1 polyclonal antibody (100
ng/ml). DRG fibers were visualized by calcein acetoxymethyl (AM)
staining on Day 5. DRG neuronal fibers were observed with a Keyence
microscope (magnification, ×200) (BZ-8100; Osaka, Japan).
Animal experiments
We established a mouse model of bone invasion by
human HNC in 7-week-old male BALB/c nude mice (each group, n=8;
total, n=56; mean body weight, 24.3 g; Charles River Laboratories,
Yokohama, Japan) by the inoculation of 1×105 SAS cells
(parental, sh-control, sh-HMGB1) with a 29-gauge needle into the
bone marrow space of the right tibial edge of the mouse under
anesthesia with 0.4 mg/kg of medetomidine, 4.0 mg/kg of midazolam,
and 5.0 mg/kg of butorphanol. The sham procedure was only a
puncture with a 29-gauage needle into the right tibial cavity. Body
condition scoring was applied and body weight was monitored daily.
Mice were maintained in housing conditions at a temperature of
23±1°C, humidity of 40–80% and lighting time of 12 h from 8 am to 8
pm. Food and water were taken freely.
Mechanical hyperalgesia was evaluated every other
day on postoperative day (POD)1-9 in each group of mice. We also
evaluated the effects of the HMGB1 neutralizing antibody and
receptor antagonist. SAS-inoculated mice were treated with
intraperitoneal injections of anti-HMGB1 polyclonal antibody, RAGE
antagonist FPS-ZM1, and TLR4 antagonist TAK-242 on POD10, and then
we performed the assay every 6 h over a 24-h period. The mice were
individually placed in a cage with a mesh floor, over a moveable
pressure-stimulating filament. The filament was positioned under
the center of the hind paw, and the time (sec) from stimulus onset
to withdrawal was recorded. Previously we reported that SAS tumor
progression in the tibia promotes bone destruction and fracture
around POD20 (18). In this
research we evaluated the direct cancer bone pain before fracture.
On POD10, the mice underwent cardiac blood collection under
anesthesia with 0.4 mg/kg of medetomidine, 4.0 mg/kg of midazolam
and 5.0 mg/kg of butorphanol (i.p), followed by cervical
dislocation. Ipsilateral L4-L5 DRGs and the right tibia were then
harvested. The criteria of humane endpoints for euthanasia was loss
of >20 percent of body weight compared to the age-matched
controls. Death of the animal was verified by cessation of
cardiovascular and respiratory movements. All of the animal
experimental protocols were approved by the Ethics Review Committee
for Animal Experimentation of the Okayama University Graduate
School of Medicine and Dentistry (OKU-2018701, 20/Nov/2018).
In vivo analysis of HNC-BP
We evaluated the mechanical hyperalgesia in the mice
with the use of a Dynamic Plantar Aesthesiometer (#37450; Ugo
Basile), which measures an animal's withdrawal latency to
non-painful pressure at the proximal half of the plantar surface of
the ipsilateral hind paw, and we applied an increasing force (2.75
g/sec). Prior to this examination, the mice were allowed to
acclimate to the testing environment for 30 min. The testing
environment consisted of translucent plastic-walled individual
chambers with a metal mesh bottom. When the mouse withdrew its hind
paw, the mechanical stimulus stopped automatically and the latency
was recorded by the device precisely in 0.1-sec increments. The
test was performed every other day on POD1-9 in the sham, parental,
sh-control and sh-HMGB1 groups. For the evaluations of the effects
of the HMGB1 neutralizing antibody and receptor antagonist,
intraperitoneal injections of anti-HMGB1 antibody (2 mg/kg),
FPS-ZM1 (10 mg/kg), and TAK-242 (10 mg/kg) were respectively
conducted for SAS-inoculated mice, and the assays were performed
every 6 h over the 24-h period on POD10. The concentration of
agents used in vivo are referenced from previous studies
(19–21).
In vivo radiography and the
measurement of osteolytic lesion areas
Osteolytic bone destruction was assessed on
radiographs. The bones were placed against films (22×27 cm; Fuji
Industrial Film FR; Fuji Photo Film) and exposed to soft X-rays at
35 kV for 15 sec with the use of a Sofron apparatus (Sofron).
HMGB1 concentration measurement
The HMGB1 concentrations in the mouse tibia, whole
blood serum, and conditioned medium were evaluated by enzyme-linked
immunosorbent assay (ELISA). Both ends of the tibias were cut, and
the bone marrow serum was extracted by centrifugation. The culture
supernatant had been collected after the incubation of
5.0×106 parental, sh-control and sh-HMGB1 SAS cells in
DMEM plus 2% FBS for 24 h. Each concentration of HMGB1 was measured
by the HMGB1 ELISA Kit (HMGB1 ELISA Kit II; Shino-Test). The
protocol of the manufacturer was followed.
DRG processing
Corrected DRGs were homogenized in RIPA lysis buffer
with 1 mM PMSF and phosphatase inhibitor
(Na3VO4 and NaF) added. The lysate was
centrifuged at 15,000 × g for 5 min at 4°C, and the supernatant was
collected as total protein. Some of the collected DRGs were fixed
in 10% neutral-buffered formalin and then embedded in paraffin.
Western blotting and immunofluorescence were performed using these
DRGs.
Immunofluorescence analysis
We conducted an immunofluorescence analysis to
determine the expressions of p-ERK in DRGs from each group of mice.
The specimens were incubated with 3% bovine serum albumin-phosphate
buffered saline (BSA-PBS) blocking solution, and then with p-ERK
antibody (dilution 1:200) and anti-CGRP antibody (dilution 1:200)
overnight at 4°C as primary antibodies, followed by Alexa Fluor 488
anti-rabbit IgG (dilution 1:1,000) or and Alexa Fluor 647 anti-goat
IgG (1:1,000) as secondary antibodies. Nuclei were counterstained
with Fluoroshield mounting medium with DAPI (#ab104139; Abcam).
Statistical analyses
The data were analyzed using an unpaired Student's
t-test for comparisons of two groups and by performing a one-way
analysis of variance (ANOVA) and a post hoc Tukey's test for the
analysis of multiple group comparisons with Graph Pad Prism, ver.
7.0 (GraphPad Software, Inc.). The results are expressed as the
mean ± standard deviation (SD). Probability (P)-values <0.05
were considered significant.
Results
HMGB1 expression in the human HNC
samples
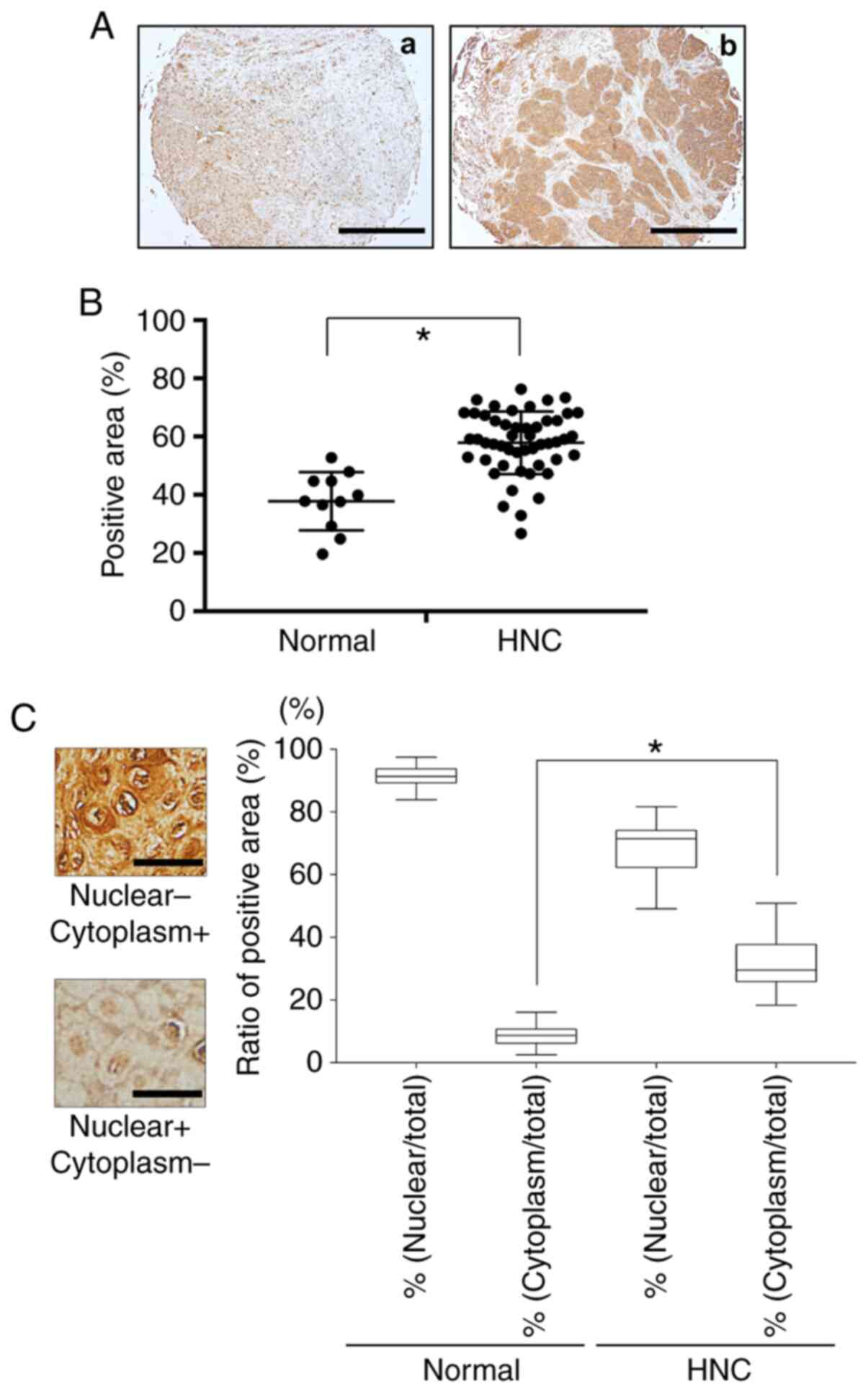
Fig. 1 provides a
representative histologic pattern of normal oral tissue and HNC
tissue. HMGB1 was highly expressed in the HNC patient samples
compared to the normal head and neck samples (Fig. 1A and B). The ratios of
HMGB1-positive nuclei in each HNC sample and the normal oral
tissues were the same, but the ratio of cytoplasm HMGB1-positive
cells was much higher in the HNC tissue compared to the normal oral
tissue (Fig. 1C).
Effect of HNC-derived HMGB1 on neurite
sprouting
To determine whether HNC cells express HMGB1 in
vitro, we performed a western blot analysis of HMGB1 expression
in the HNC cell lines. As shown in Fig.
2A, the results of the western blot analysis revealed high
expression of HMGB1 in the SAS cells. To examine the role of
cancer-derived HMGB1 in neurite sprouting in vitro, we first
evaluated the expression of HMGB1 in sensory neurons. Fig. 2B demonstrates that the bone and
neuronal cell lines expressed both RAGE and TLR4.
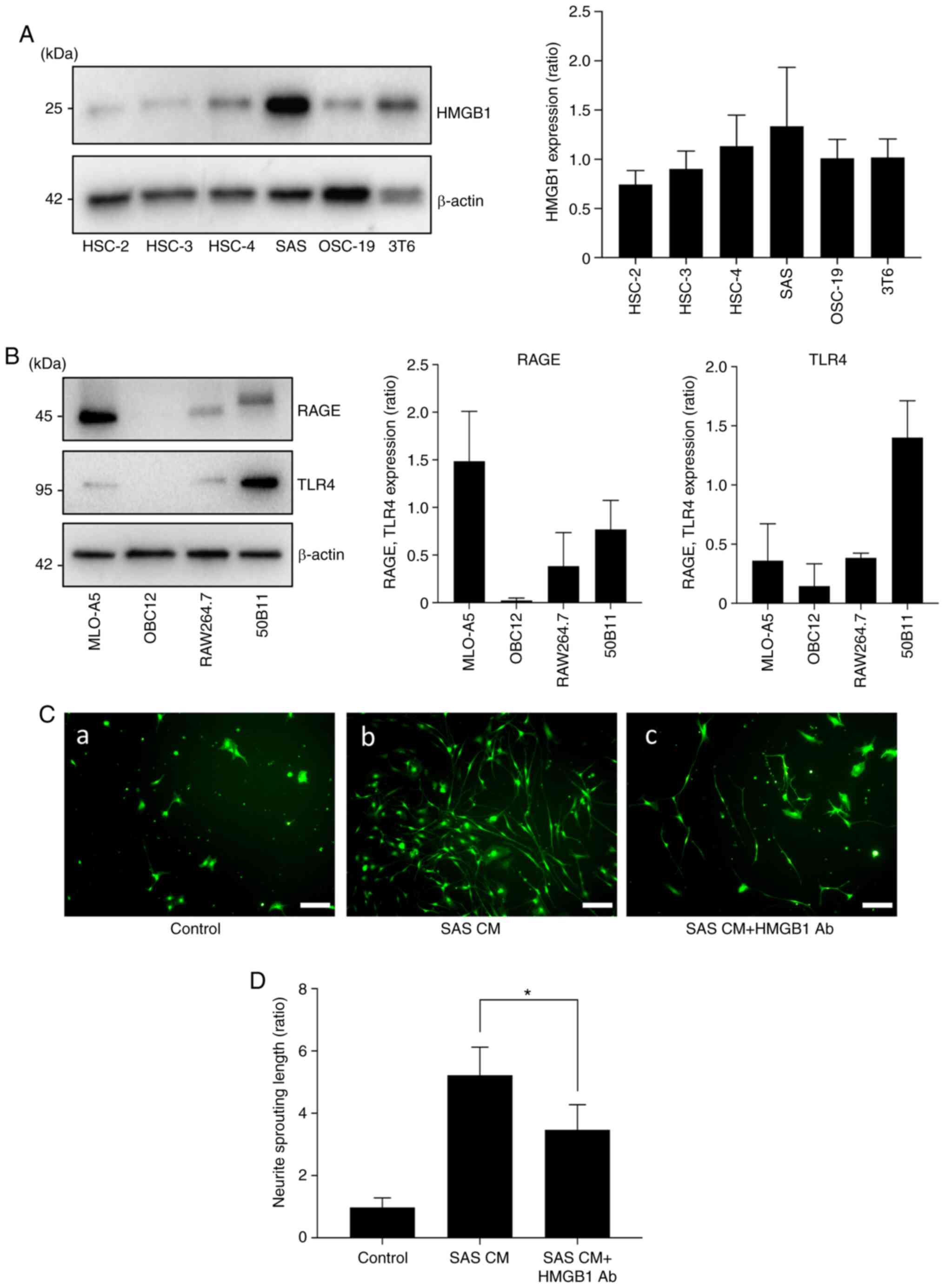 | Figure 2.Effects of HMGB1 on neurite induction
and outgrowth. (A) Expression of HMGB1 in HNC cell lines (HSC-2,
HSC-3, HSC-4, SAS, OSC-19) and mouse fibroblast cell line (3T6) by
western blot analysis. The right panel shows the quantification
(n=3). (B) Expressions of RAGE and TLR4 in each cell line:
Osteocytes (MLO-A5), osteoblasts (OBC12), macrophages prior to
osteoclast differentiation (RAW264.7), and neurons (50B11). The
right panel shows the quantification (n=3). (C) Neurite outgrowth
from primary sensory neuron cells in neuron growth medium
containing conditioned medium of SAS (CM) with/without HMGB1
neutralizing antibody. (a) Control, (b) SAS CM, and (c) SAS CM with
HMGB1 antibody (Ab) for 5 days, labeled with calcein AM. Scale bar,
100 µm. (D) Quantitative data of neurite length shown in C. Error
bars: Mean ± SD; *P<0.01 vs. SAS CM. HMGB1, high mobility group
box 1; HNC, head and neck cancer; RAGE, advanced glycation end
products; TLR4, Toll-like receptor 4. |
We next cultured the DRG sensory neuron cells in
neuron growth medium containing conditioned medium of SAS (CM) with
or without HMGB1 neutralizing antibody (Ab) to evaluate sensory
neuron axis sprouting. Fig. 2C and
D shows that the cultures with SAS CM exhibited increased
neuron lengths, and the HMGB1 neutralizing antibody suppressed that
effect.
HMGB1 neutralizing antibody suppresses
HNC-BP and sensory nerve excitation in vivo
The mouse tibias injected with SAS cells developed
aggressive proliferation of SAS cells in the bone marrow and bone
destruction (Fig. 3A). To evaluate
the role of HMGB1 in SAS-induced HNC-BP, we investigated SAS
injection-induced mechanical hyperalgesia in the mouse bone marrow
after the injection of SAS cells. At POD10, we observed that the
treatment with the HMGB1 neutralizing antibody significantly
reduced hyperalgesia compared to the parental SAS cell-injected
mice (Fig. 3B).
In parallel with the HNC-BP results, the DRGs from
the SAS-injected mice demonstrated an increased expression of
phosphorylated (p)-ERK1/2, which is a molecular indicator of neuron
excitation. In contrast, the DRGs from the sham-operated mice and
the HMGB1 neutralizing antibody-treated mice showed decreased
p-ERK1/2 expression in the western blot analysis (Fig. 3C).
RAGE and the TLR4 antagonist suppress
the sensory nerve excitation and HNC-BP in vivo
To evaluate the role of the HMGB1 receptor in
SAS-induced HNC-BP, we investigated SAS injection-induced
mechanical hyperalgesia. Treatment with the RAGE antagonist FPS-ZM1
or TLR4 antagonist TAK-242 both significantly reduced hyperalgesia
compared to the SAS cell-injected mice (Fig. 4A). As expected, the DRGs from the
mice administered the RAGE antagonist or TLR4 antagonist showed
significantly decreased p-ERK1/2 expression in the western blot and
immunofluorescence analyses (Fig. 4B
and C). Calcitonin gene-related peptides (CGRP) are
neuropeptides expressed in sensory nerve, and are a positive
control of sensory neurons.
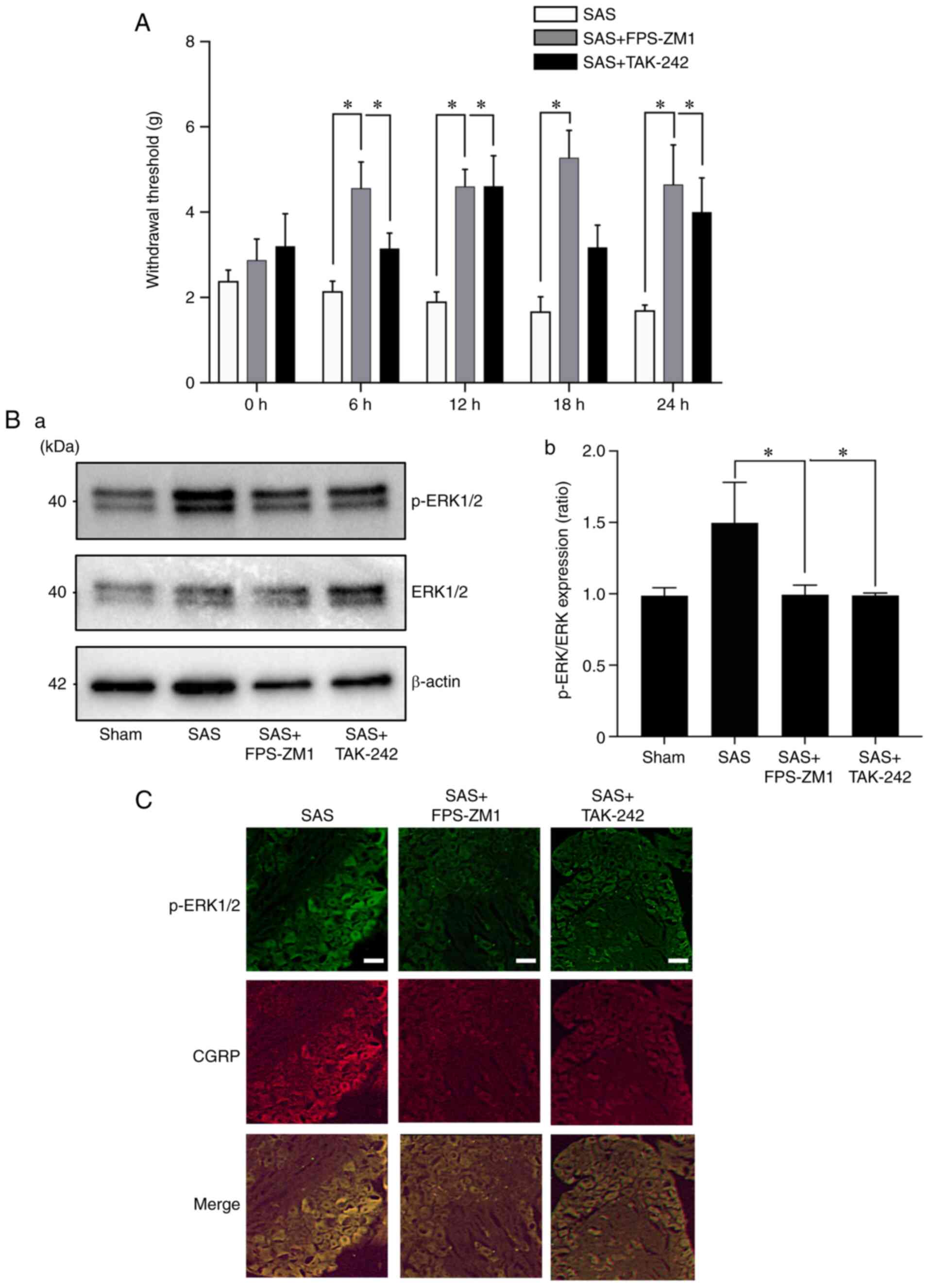 | Figure 4.Effects of the HMGB1 receptor
antagonists in HNC-BP in vivo. (A) The 24-h time course
analysis of mechanical hyperalgesia following treatment with
vehicle (0.1% DMSO in PBS), RAGE antagonist FPS-ZM1, or TLR4
antagonist TAK-242 (both 10 mg/kg) in SAS-injected mice on POD10
(n=8). Error bars: Mean ± SD. *P<0.05 vs. the SAS group. (B)
Expression of phosphorylated (p-)ERK1/2 in the dorsal root ganglia
(DRG) of the mice shown in panel A by western blot analysis. (a)
Representative blot and (b) quantification with densitometry of
p-ERK/ERK (n=3). Error bars: Mean ± SD. *P<0.05 vs. the SAS
group. (C) Immunofluorescence analysis of DRGs. Upper panels,
p-ERK1/2 (green); middle panels, CGRP (red); lower panels, merged.
Scale bar, 50 µm. HMGB1, high mobility group box 1; HNC, head and
neck cancer; HNC-BP, HNC-associated bone pain; CGRP, calcitonin
gene-related peptide. |
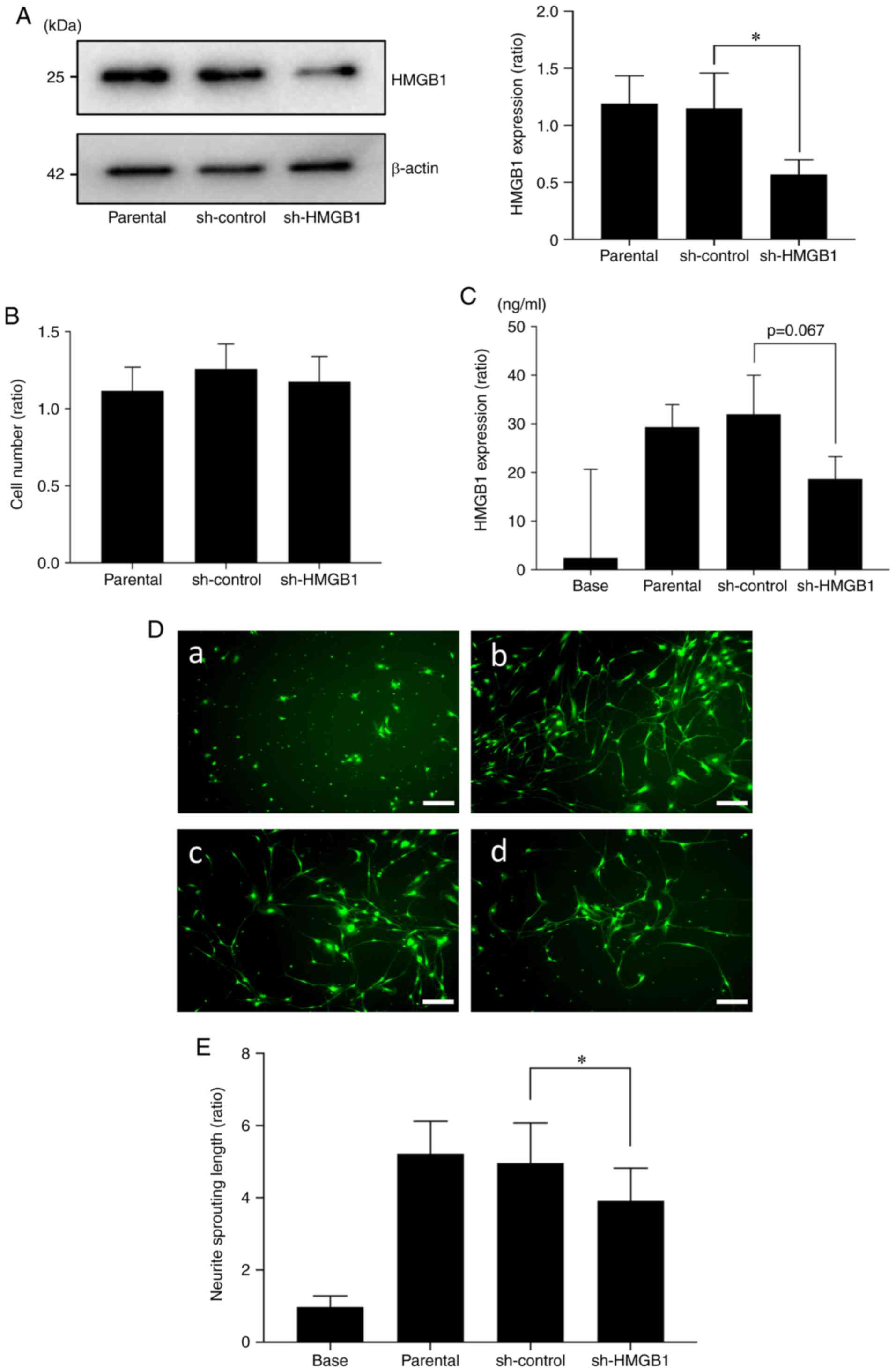
Reduction in HMGB1 in SAS cells
decreases sensory nerve sprouting
Next, to investigate the role of cancer-derived
HMGB1 in HNC-BP, we introduced an shRNA plasmid targeting HMGB1
into SAS cells by an electroporation system. As shown in Fig. 5A, a 60% suppression of the
expression of HMGB1 protein in SAS was observed in the
shHMGB1-transfected group (sh-HMGB1) compared to the parental SAS
cells and control shRNA (sh-control) plasmid-introduced group. The
proliferation ability of the HMGB1-knockdown SAS cells did not
differ significantly from that of the sh-control SAS cells
(Fig. 5B). HMGB1 in conditioned
medium tended to be decreased in the shHMGB1-transfected SAS cells
(Fig. 5C). This conditioned medium
promoted significantly less sensory neuron sprouting in the in
vitro cultures (Fig. 5D and
E).
HMGB1 derived from HNC promotes HNC-BP
and the excitation of sensory nerves in vivo
To evaluate cancer-derived HMGB1, we injected the
parental, sh-control, and sh-HMGB1 SAS cells into mouse tibial bone
marrow and determined the systemic and local serum HMGB1
concentrations on POD10. Tumor area of the SAS parental (Fig. 6A-a), sh-control (Fig. 6A-b) and sh-HMGB1 (Fig. 6A-c) cell groups are shown. At a
point on POD10, there were no significant differences in regards to
tumor burden in the tibia and body weight between the
HMGB1-knockdown SAS vs. parental and sh-control SAS cell-injected
mice (Fig. 6A-d and -e). There were
no significant between-group differences in the HMGB1
concentrations in the cardiac tissues (Fig. 6B-a). Compared to the parental SAS
and sh-control SAS cell-injected mice, the injection of sh-HMGB1
SAS cells significantly decreased the HMGB1 levels in the bone
marrow (Fig. 6B-b). In parallel
with these results, HMGB1 knockdown significantly reduced
hyperalgesia compared to the parental SAS- and sh-control SAS
cell-injected mice on POD5, 7 and 9 (Fig. 6C). Similar to the HNC-BP results,
the DRGs from the parental SAS or sh-control SAS cell-injected mice
demonstrated an increased expression of p-ERK1/2, which is a
molecular indicator of neuron excitation. In contrast, the DRGs
from the HMGB1-knockdown SAS cell-injected mice showed decreased
p-ERK1/2 expression in the western blot and immunofluorescence
analyses (Fig. 6D and E).
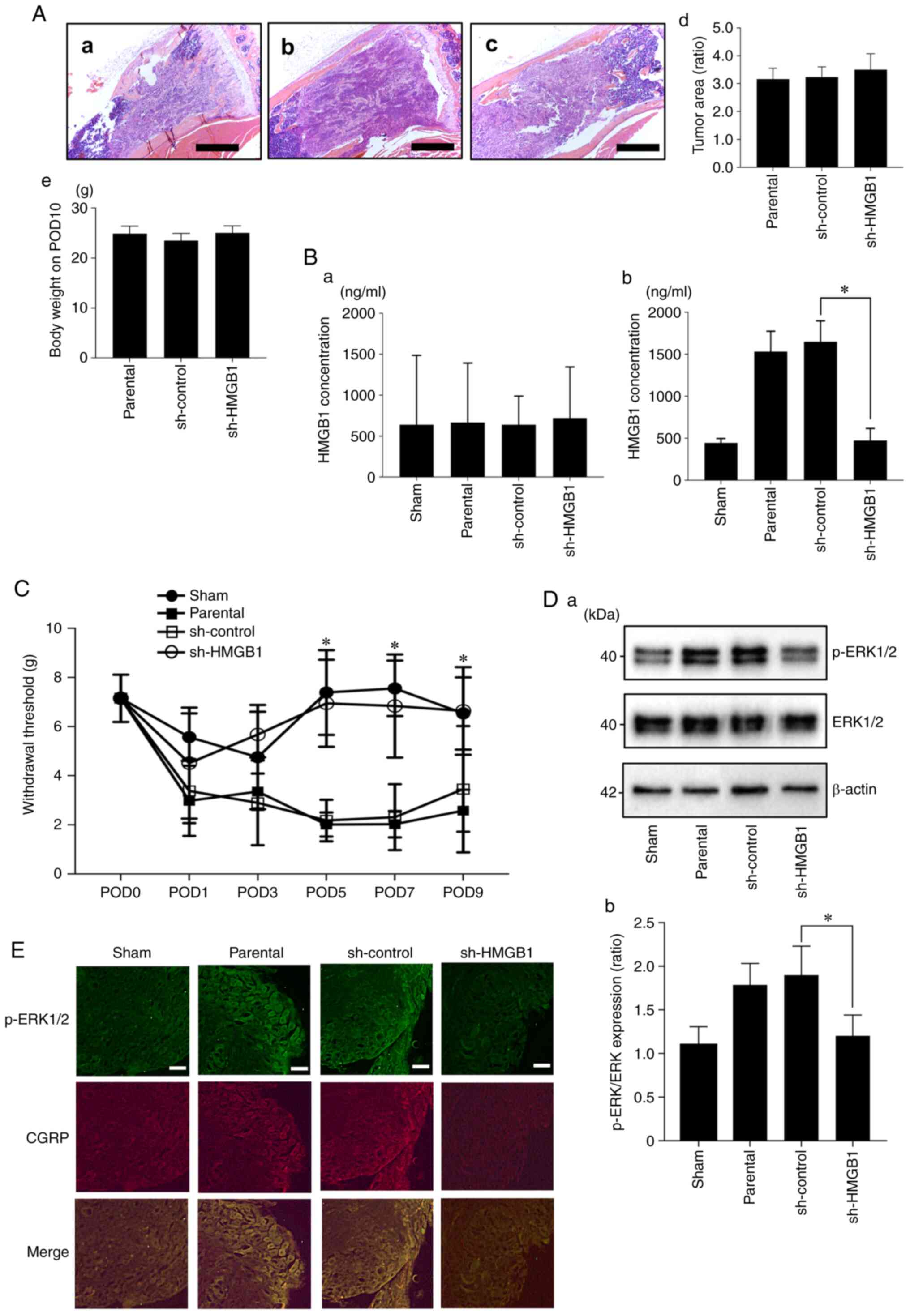 | Figure 6.Effect of the reduction of HMGB1 on
HNC-BP and sensory neuron excitation in vivo. (A) Tumor area
of SAS parental, sh-control and sh-HMGB1 groups in tibia bone
marrow (hematoxylin and eosin) at ×40 magnification. Scale bar, 1
mm. (a) Parental, (b) sh-control and (c) sh-HMGB1 groups. (d)
Quantitative data (n=4). There was no significant difference
between each of them. (e) Body weight of each group on POD10 (n=8).
(B) HMGB1 concentrations in cardiac blood samples and tibia bone
marrow. (a) HMGB1 concentration in the cardiac blood (n=3). (b)
HMGB1 concentration in the tibia bone marrow (n=4). Error bars:
Mean ± SD. *P<0.01 vs. sh-control. (C) Mechanical hyperalgesia
results of each group. The test was performed every other day from
POD1 to POD9 (n=8). Error bars: Mean ± SD. *P<0.05 vs.
sh-control. (D) Excitation of sensory neurons determined by
phosphorylated (p-)ERK1/2 expression in the DRGs of the sham,
parental, sh-control, sh-HMGB1 SAS-injected mice by western
blotting analysis. (a) Representative blot and (b) quantification
with densitometry of p-ERK/ERK (n=3). Error bars: Mean ± SD.
*P<0.05 vs. sh-control. (E) Immunofluorescence analysis of DRGs.
Upper panels, p-ERK1/2 (green); middle panels, CGRP (red); lower
image, merged. Scale bar, 50 µm. HMGB1, high mobility group box 1;
HNC, head and neck cancer; HNC-BP, HNC-associated bone pain; POD,
postoperative day; CGRP, calcitonin gene-related peptide. |
The three redox forms of HMGB1 have
different roles in sensory neuron excitation
HMGB1 contains three conserved cysteine residues:
Cys23, Cys45 and Cys106. The functions of HMGB1 depend on
redox-sensitive cysteine residues (Cys23, Cys45 and Cys106) of the
protein. HMGB1 has at least three redox forms with different
biological functions: i) All-thiol HMGB1 (all three cysteine
residues in the thiol state), ii) ‘disulfide HMGB1’ (an
intramolecular disulfide bridge between Cys23 and Cys45; Cys106 in
the thiol state), and ii) ‘Terminal oxidized HMGB1’ (all three
cysteines in the hyperoxidized sulfonic acid state). The western
blot results indicated that all-thiol HMGB1 and disulfide HMGB1
directly stimulated DRG sensory neuron cells as determined by a
significant increase in p-ERK. Furthermore, they both sensitized
the acid-induced stimulation by significantly increasing the TRPV1
expression, and also HMGB1 neutralizing antibody addition
suppressed TRPV1 expression (Fig. 7A
and B).
Discussion
Bone-invasive cancer including head and neck cancer
(HNC), breast cancer, and multiple myeloma frequently invade and
metastasize to bone, promoting bone pain. Cancer cells in bone
marrow release various cytokines and growth factors (13), and these cytokines contribute to the
activation of sensory neurons, leading to cancer-induced bone pain
(22). High mobility group box 1
(HMGB1) is a 25-kDa non-histone DNA binding protein that is
generally distributed in the cell nucleus. The best-studied roles
of HMGB1 are those in the immune system. Nuclear HMGB1 acts as a
DNA chaperone which regulates DNA repair and transcription
(23). HMGB1 is also a
damage-associated molecular pattern (DAMP), that is released from
dead cells and dendritic cells (24,25).
HMGB1 is considered an inflammatory cytokine released from
activated monocytes and macrophages (26,27).
It has been reported that HMGB1 secreted from
various cancer cells and the blood serum HMGB1 concentration are
both correlated with poor patient prognosis (28,29).
Another study indicated that HMGB1 promotes sensory neuron
sprouting and neuropathic pain in diseases such as pancreatitis,
bladder pain, and arthritis (30).
However, the role of HMGB1 in HNC bone pain (HNC-BP) remains
unclear. Our present findings demonstrated that HNC SAS cells
express HMGB1 in the cytoplasm and subsequently release HMGB1 to
the extracellular space. In addition, our in vivo data
revealed that HNC cells actively effuse HMGB1 to the extracellular
space in bone marrow.
HMGB1 was found to evoke the influx of Ca in the
neuron axis (31). Our present
findings showed that SAS cells promote sensory neuron sprouting and
elongation of the axis, and HMGB1 neutralizing antibody suppressed
SAS conditioned medium-induced sensory neuron sprouting. These
results indicate cancer-derived HMGB1 sensitization via increasing
the length of sensory neuron axis.
Cytokines released from cancer promote
osteoclastogenesis, and osteoclasts subsequently resorb the bone
matrix by protons (32). The
extracellular environment that is created by releasing protons from
osteoclasts and by cancer contributes to the activation of sensory
neurons and bone pain (15,33). Osteoclasts release protons via
a3V-ATPase, which is a V-type proton pump (34). Cancer cells released protons as
lactic acid in a metabolic process (35).
The phosphorylation of ERK and CREB is a marker of
sensory neuron excitation. Our present results demonstrated that
SAS cell-derived HMGB1 increased the acid sensitivity of sensory
neuron cells via the HMGB1 axis. Calcium influx is a direct
indicator of cell excitation. Herein, the HMGB1 neutralizing
antibody and RAGE antagonist similarly suppressed the cell
excitation by proton stimulation followed by SAS conditioned medium
co-incubation. We therefore speculate that HMGB1 from cancer cells
enhances HNC-BP via acid sensitization.
As expected, the HMGB1 neutralizing antibody
suppressed the cancer bone pain that was caused by SAS cell
injection into mouse tibial bone. HMGB1 is released from various
cells such as dendritic cells and necrotic cells. To investigate
the importance of cancer-derived HMGB1 for HNC-BP but not other
microenvironmental cells, we established HMGB1-knockdown SAS cells.
The results showed that the HMGB1 level in the bone marrow was
correlated with the HMGB1 expression in SAS cells. In addition, the
HMGB1 knockdown in SAS cells decreased the sensory neuron
excitation in vitro, and it decreased the HNC-BP in mice
in vivo. This result indicated that the cancer-derived HMGB1
promoted HNC-BP.
HMGB1 has two receptors, i.e., TLR4 and RAGE
(20). The question of which
receptor(s) play a critical role in HNC-BP remains unanswered. It
was reported that different redox states of the three cysteines of
HMGB1 endow it with mutually exclusive activities (36). Fully reduced HMGB1 binds mainly to
RAGE, and disulfide HMGB1 binds to TLR4 (37). RAGE signaling is suspected to
promote the phosphorylation of ERK, which is a marker of sensory
neuron activation (38). The
results of the present investigation demonstrated that compared to
the partly oxidized form (disulfide HMGB1), the fully reduced form
(All-thiol HMGB1) evoked sensory neuron excitation in vitro.
In the in vivo experiment, the RAGE antagonist strongly
suppressed HNC-BP compared to the TLR4 antagonist. Our results
demonstrated that RAGE (and not TLR4) is critical for DRG
excitation and HNC-BP.
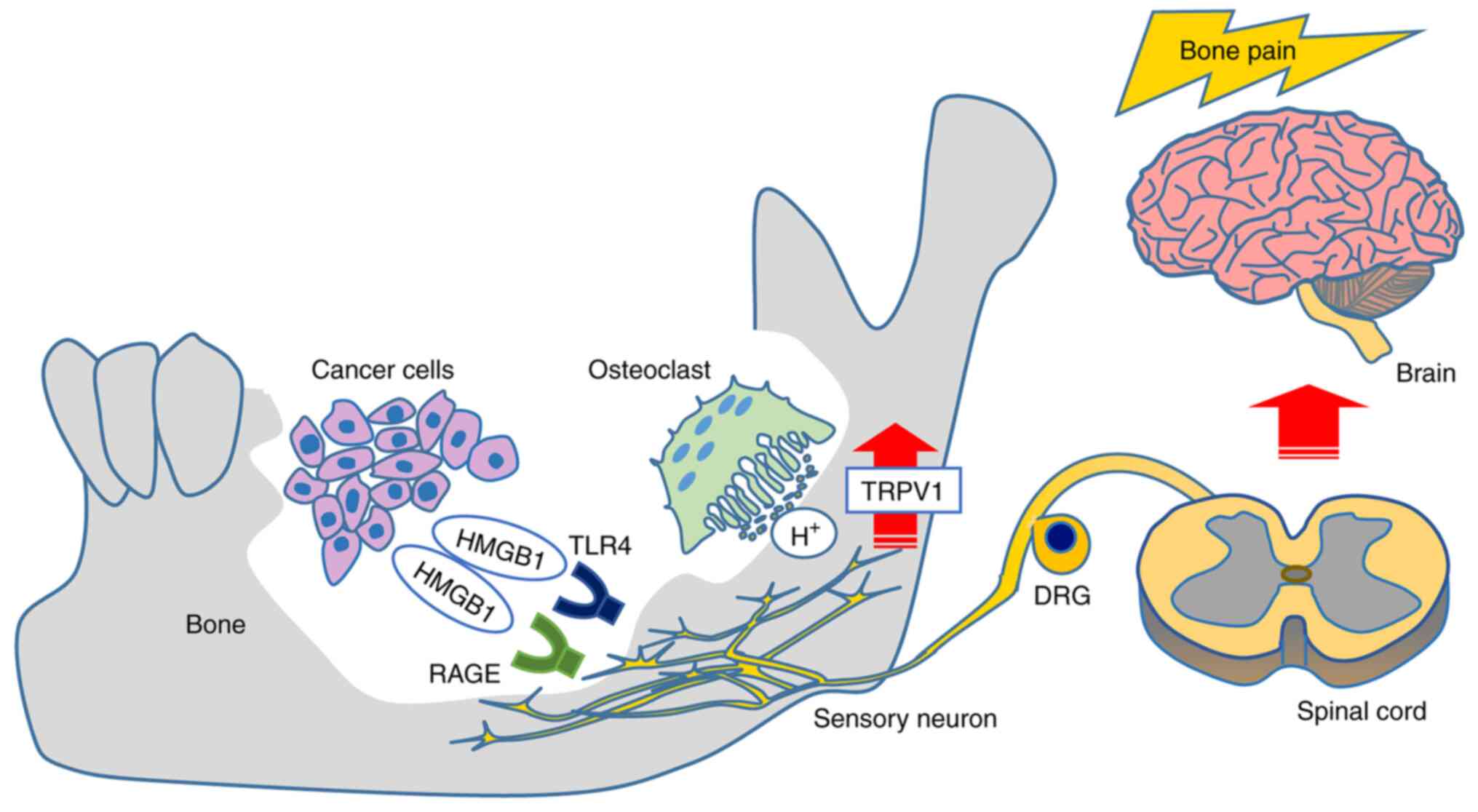
In conclusion, our present findings demonstrated
that the HMGB1 released from HNC cells promotes HNC bone pain via
sensory neurons innervating bone marrow, acid sensing receptor
TRPV1 activation, and a direct excitation of sensory neurons by
RAGE signaling rather than by a TLR4 pathway (Fig. 8). Targeting these pathways may
provide effective mechanism-based therapies for the control of
HNC-BP, which is currently undertreated. Furthermore, TLR4 and RAGE
antagonist are expected as newly therapeutic agents against
diabetes mellitus, sepsis and rheumatoid arthritis that are also
painful diseases (39). Future
research for HMGB1 will improve the treatment methods of these
diseases.
Acknowledgements
Not applicable.
Funding
This study was supported by a Grant-in-Aid for Young
Scientists (#18K17225 to TO) and a Grant-in-Aid for Scientific
Research (B) (#20H03889 to AS) from the Ministry of Education,
Culture, Sports, Science, and Technology of Japan.
Availability data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TO conceived and designed the experiments. TN, TO
and KH performed the experiments. TO and TN analyzed and
interpreted the data. TN, SR, TO, KH, YK, SI, AS, KOno, KOba, and
MM performed the data acquisition. TO and TN wrote the manuscript.
TO, TS, SI and AS conducted the manuscript revision/review. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All of the animal experimental protocols were
approved by the Ethics Review Committee for Animal Experimentation
of the Okayama University Graduate School of Medicine and Dentistry
(OKU-2018701, 20/Nov/2018).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zheng Y, Zhou H, Dunstan CR, Sutherland RL
and Seibel MJ: The role of the bone microenvironment in skeletal
metastasis. J Bone Oncol. 2:47–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schmidt BL: The neurobiology of cancer
pain. J Oral Maxillofac Surg. 73 (Suppl 12):S132–S135. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Montazeri A: Quality of life data as
prognostic indicators of survival in cancer patients: An overview
of the literature from 1982 to 2008. Health Qual Life Outcomes.
7:1022009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Viet CT and Schmidt BL: Biologic
mechanisms of oral cancer pain and implications for clinical
therapy. J Dent Res. 91:447–453. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Garg AD and Agostinis P: Cell death and
immunity in cancer: From danger signals to mimicry of pathogen
defense responses. Immunol Rev. 280:126–148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carballo M, Puigdomenech P and Palau J:
DNA and histone H1 interact with different domains of HMG 1 and 2
proteins. EMBO J. 2:1759–1764. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Man LL, Liu F and Wang YJ, Song HH, Xu HB,
Zhu ZW, Zhang Q and Wang YJ: The HMGB1 signaling pathway activates
the inflammatory response in Schwann cells. Neural Regen Res.
10:1706–1712. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Das N, Dewan V, Grace PM, Gunn RJ, Tamura
R, Tzarum N, Watkins LR, Wilson IA and Yin H: HMGB1 activates
proinflammatory signaling via TLR5 leading to allodynia. Cell Rep.
17:1128–1140. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sims GP, Rowe DC, Rietdijk ST, Herbst R
and Coyle AJ: HMGB1 and RAGE in inflammation and cancer. Annu Rev
Immunol. 28:367–388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shao Y, Sha M, Chen L, Li D, Lu J and Xia
S: HMGB1/TLR4 signaling induces an inflammatory response following
high-pressure renal pelvic perfusion in a porcine model. Am J
Physiol Renal Physiol. 311:F915–F925. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Jiang Z, Yan J and Ying S: HMGB1
as a potential biomarker and therapeutic target for malignant
mesothelioma. Dis Markers. 2019:41831572019.PubMed/NCBI
|
|
12
|
Sun S, Zhang W, Cui Z, Chen Q, Xie P, Zhou
C, Liu B, Peng X and Zhang Y: High mobility group box-1 and its
clinical value in breast cancer. Onco Targets Ther. 8:413–419.
2015.PubMed/NCBI
|
|
13
|
Yi B, Williams PJ, Niewolna M, Wang Y and
Yoneda T: Tumor-derived platelet-derived growth factor-BB plays a
critical role in osteosclerotic bone metastasis in an animal model
of human breast cancer. Cancer Res. 62:917–923. 2002.PubMed/NCBI
|
|
14
|
Guise TA, Yin JJ, Taylor SD, Kumagai Y,
Dallas M, Boyce BF, Yoneda T and Mundy GR: Evidence for a causal
role of parathyroid hormone-related protein in the pathogenesis of
human breast cancer-mediated osteolysis. J Clin Invest.
98:1544–1549. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hiasa M, Okui T, Allette YM, Ripsch MS,
Sun-Wada GH, Wakabayashi H, Roodman GD, White FA and Yoneda T: Bone
pain induced by multiple myeloma is reduced by targeting V-ATPase
and ASIC3. Cancer Res. 77:1283–1295. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoneda T, Hiasa M, Nagata Y, Okui T and
White F: Contribution of acidic extracellular microenvironment of
cancer-colonized bone to bone pain. Biochim Biophys Acta.
1848:2677–2684. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wakabayashi H, Wakisaka S, Hiraga T, Hata
K, Nishimura R, Tominaga M and Yoneda T: Decreased sensory nerve
excitation and bone pain associated with mouse Lewis lung cancer in
TRPV1-deficient mice. J Bone Miner Metab. 36:274–285. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ryumon S, Okui T, Kunisada Y, Kishimoto K,
Shimo T, Hasegawa K, Ibaragi S, Akiyama K, Thu Ha NT, Monsur Hassan
NM and Sasaki A: Ammonium tetrathiomolybdate enhances the antitumor
effect of cisplatin via the suppression of ATPase copper
transporting beta in head and neck squamous cell carcinoma. Oncol
Rep. 42:2611–2621. 2019.PubMed/NCBI
|
|
19
|
Ueno H, Matsuda T, Hashimoto S, Amaya F,
Kitamura Y, Tanaka M, Kobayashi A, Maruyama I, Yamada S, Hasegawa
N, et al: Contributions of high mobility group box protein in
experimental and clinical acute lung injury. Am J Respir Crit Care
Med. 170:1310–1316. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu C, Ding X, Zhou C, Ye P, Sun Y, Wu J,
Zhang A, Huang X, Ren L, Wang K, et al: Inhibition of intimal
hyperplasia in murine aortic allografts by administration of a
small-molecule TLR4 inhibitor TAK-242. Sci Rep. 7:157992017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang F, Wang Z, Zhang JH, Tang J, Liu X,
Tan L, Huang QY and Feng H: Receptor for advanced glycation
end-product antagonist reduces blood-brain barrier damage after
intracerebral hemorrhage. Stroke. 46:1328–1336. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu XM, Yang W, Du LX, Cui WQ, Mi WL,
Mao-Ying QL, Chu YX and Wang YQ: Vascular endothelial growth factor
a signaling promotes spinal central sensitization and pain-related
behaviors in female rats with bone cancer. Anesthesiology.
131:1125–1147. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Javaherian K, Liu JF and Wang JC:
Nonhistone proteins HMG1 and HMG2 change the DNA helical structure.
Science. 199:1345–1346. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Antoine DJ, Jenkins RE, Dear JW, Williams
DP, McGill MR, Sharpe MR, Craig DG, Simpson KJ, Jaeschke H and Park
BK: Molecular forms of HMGB1 and keratin-18 as mechanistic
biomarkers for mode of cell death and prognosis during clinical
acetaminophen hepatotoxicity. J Hepatol. 56:1070–1079. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dumitriu IE, Baruah P, Valentinis B, Voll
RE, Herrmann M, Nawroth PP, Arnold B, Bianchi ME, Manfredi AA and
Rovere-Querini P: Release of high mobility group box 1 by dendritic
cells controls T cell activation via the receptor for advanced
glycation end products. J Immunol. 174:7506–7515. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lotze MT and Tracey KJ: High-mobility
group box 1 protein (HMGB1): Nuclear weapon in the immune arsenal.
Nat Rev Immunol. 5:331–342. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang W, Li J, Gallowitsch-Puerta M,
Tracey KJ and Pisetsky DS: The effects of CpG DNA on HMGB1 release
by murine macrophage cell lines. J Leukoc Biol. 78:930–936. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fang J, Ge X, Xu W, Xie J, Qin Z, Shi L,
Yin W, Bian M and Wang H: Bioinformatics analysis of the prognosis
and biological significance of HMGB1, HMGB2, and HMGB3 in gastric
cancer. J Cell Physiol. 235:3438–3446. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu T, Zhang W, Yang G, Li H, Chen Q, Song
R and Zhao L: HMGB1 overexpression as a prognostic factor for
survival in cancer: A meta-analysis and systematic review.
Oncotarget. 7:50417–50427. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saleh A, Smith DR, Tessler L, Mateo AR,
Martens C, Schartner E, Van der Ploeg R, Toth C, Zochodne DW and
Fernyhough P: Receptor for advanced glycation end-products (RAGE)
activates divergent signaling pathways to augment neurite outgrowth
of adult sensory neurons. Exp Neurol. 249:149–159. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Allette YM, Due MR, Wilson SM, Feldman P,
Ripsch MS, Khanna R and White FA: Identification of a functional
interaction of HMGB1 with receptor for advanced glycation
end-products in a model of neuropathic pain. Brain Behav Immun.
42:169–177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Teitelbaum SL: Bone resorption by
osteoclasts. Science. 289:1504–1508. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Caterina MJ, Schumacher MA, Tominaga M,
Rosen TA, Levine JD and Julius D: The capsaicin receptor: A
heat-activated ion channel in the pain pathway. Nature.
389:816–824. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu H, Xu G and Li YP: Atp6v0d2 is an
essential component of the osteoclast-specific proton pump that
mediates extracellular acidification in bone resorption. J Bone
Miner Res. 24:871–885. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hasegawa K, Okui T, Shimo T, Ibaragi S,
Kawai H, Ryumon S, Kishimoto K, Okusha Y, Monsur Hassan NM and
Sasaki A: Lactate transporter monocarboxylate transporter 4 induces
bone pain in head and neck squamous cell carcinoma. Int J Mol Sci.
19:33172018. View Article : Google Scholar
|
|
36
|
Janko C, Filipovic M, Munoz LE, Schorn C,
Schett G, Ivanovic-Burmazovic I and Herrmann M: Redox modulation of
HMGB1-related signaling. Antioxid Redox Signal. 20:1075–1085. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yamasoba D, Tsubota M, Domoto R, Sekiguchi
F, Nishikawa H, Liu K, Nishibori M, Ishikura H, Yamamoto T, Taga A
and Kawabata A: Peripheral HMGB1-induced hyperalgesia in mice:
Redox state-dependent distinct roles of RAGE and TLR4. J Pharmacol
Sci. 130:139–142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Balosso S, Liu J, Bianchi ME and Vezzani
A: Disulfide-containing high mobility group box-1 promotes
N-methyl-D-aspartate receptor function and excitotoxicity by
activating Toll-like receptor 4-dependent signaling in hippocampal
neurons. Antioxid Redox Signal. 21:1726–1740. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
VanPatten S and Al-Abed Y: High mobility
group Box-1 (HMGb1): Current wisdom and advancement as a potential
drug target. J Med Chem. 61:5093–5107. 2018. View Article : Google Scholar : PubMed/NCBI
|