Introduction
Voltage-dependent anion channel (VDAC)1, as a
mitochondrial porin, controls the entry and exit of metabolites and
energy between the mitochondria and the cytosol. Therefore, it is a
target convergence for both cell death and mitochondria-mediated
apoptosis (1). In 2008, the 3D
structure of VDAC1 was determined by NMR spectroscopy and X-ray
crystallography (2–4). VDAC1 comprises a barrel formed by 19
β-strands and an N-terminal α-helix folded into the barrel
interior. The N-terminus is composed of hydrophilic amino acid
residues and has a higher positive charge density than negative
charge density (4). The N-terminus
stabilizes the barrel, preventing it from adopting a partially
dilapidated, low-conductance closed state. In addition, the
N-terminus reportedly contributes to VDAC1 oligomerization and
cellular apoptosis through interactions with hexokinase and
adenosine triphosphate (ATP) (5).
In the mitochondrial outer membrane (MOM), VDAC1, adenine
nucleotide translocase (ANT) and cyclophilin D, constitute the
permeability transition pore (PTP), which is involved in the
release of pro-apoptotic factors from the intra-mitochondrial space
(6).
VDAC1 channel permeability, which is an important
feature of the PTP, is mediated by ions, small molecules, and
protein kinases (7,8). Studies have demonstrated that VDAC1
interacts with hexose kinase (5,9–11), the
pro-apoptotic proteins Bax and Bak, and the anti-apoptotic
proteins, Bcl-2 and Bcl-xL (12,13).
Previous mechanistic studies have revealed that PKA-dependent VDAC1
phosphorylation decreases VDAC1 conductance, glycogen synthase
kinase 3β (GSK3β)-mediated VDAC1 phosphorylation appears to promote
channel opening and chemotherapy-induced cytotoxicity, and the
AMPK/mTOR signalling axis also controls mitochondrial metabolism
(14,15). These studies highlight the
importance of the VDAC1 protein for cancer cell survival and
mitochondrial apoptosis.
The mitochondria prefer to be localized within the
tubulin-microtubule network and to move along microtubules, which
are cylindrical polymers composed of α and β tubulin. Microtubules
are the primary components of the cytoskeleton and mediate crucial
cellular functions. Tubulin proteins exhibit a high affinity for
VDAC1 by binding to the mitochondria. A previous study confirmed
that tubulins are inherent components of the mitochondrial membrane
and may be involved in modulating mitochondrial permeability and
subsequently controlling respiration (16). The same previous study also
demonstrated that dimeric tubulin at nanomolar levels induces
reversible closure of VDAC in a voltage-sensitive manner. The
tubulin protein has an extended C-terminal tail with a negative
charge. These anionic C-terminal tails are essential for the
interaction of tubulin with the cationic N-terminus of VDAC in the
barrel at the mitochondria-cytosol interface. In the tubulin-VDAC
interaction model, the anionic C-terminus of tubulin penetrates
into the VDAC cationic channel lumen and specifically interacts
with VDAC, thus blocking channel conductance (17). This indicates that the level of
dimeric tubulin or free α/β tubulin may regulate the permeability
of VDAC in the MOM and then influence mitochondria-mediated
cellular apoptosis. Paclitaxel and other microtubule-targeting
anti-tumour drugs reportedly modify the interactions of
microtubules and/or tubulin with VDAC and induce overexpression of
VDAC1 (18). The high levels of
VDAC1 may shift VDAC1 from a monomeric to an oligomeric assembly,
which promotes CytC release from the mitochondria, leading to
apoptosis (18). However, the
critical factor is whether these drugs change the levels of free
tubulins that interact with VDAC in the MOM and subsequently affect
VDAC permeability. This question is of special interest and is
worth exploring.
Geldanamycin and its analogue, 17-AAG, inhibitors of
heat shock protein (Hsp)90, have been shown to interact with the
mitochondria, particularly with VDAC, through hydrophobic
interactions independent of Hsp90, increasing the intracellular
Ca(2+) concentrations and decreasing the plasma membrane cationic
current (19). In previous studies,
it was found that geldanamycin and its analogue, 17-AAG, both
induced lung cancer cell apoptosis effectively by inhibiting Hsp90
(20). Therefore, the present study
aimed to investigate whether tanespimycin (17-AAG) associates with
VDAC by interacting with it in the MOM during apoptosis induction.
Herein, the experiments revealed that tanespimycin induced
α-tubulin acetylation and increased the expression of VDAC and Bax.
In particular, docetaxel, a microtubule stabilizer that promotes
the hyperpolarization of microtubules, enhanced the levels of
acetylated α-tubulin, VDAC and Bax when combined with tanespimycin.
As a result, cellular apoptosis was induced. This implies that the
two drugs have a similar mechanism of apoptosis induction. However,
at present, the molecular mechanisms underlying the association
between α-tubulin acetylation and VDAC in apoptosis upon treatment
with tanespimycin remain unclear. Therefore, the present study
aimed to explore the mechanisms between α-tubulin acetylation and
VDAC upregulation in apoptosis in lung cancer cells.
Materials and methods
Reagents
Tanespimycin was provided by LC Laboratories.
Docetaxel was purchased from American Radiolabeled Chemicals, Inc.
Rapamycin was purchased from Cell Signaling Technology, Inc. Each
of these compounds was dissolved in dimethyl sulfoxide (DMSO) at a
given concentration, and aliquots were stored at −20°C. The stock
solutions were diluted to the final appropriate concentration
immediately prior to use.
Antibodies
Mouse monoclonal anti-caspase-3 antibody (cat. no.
31A1067) and anti-caspase-8 antibody (cat. no. 9746) were purchased
from Imegenex and Cell Signaling Technology, Inc., respectively.
Rabbit anti-VDAC1 antibody (cat. no. 12454), mouse anti-caspase-9
antibody (cat. no. 9508) and anti-poly (ADP-ribose) polymerase
(PARP) antibody (cat. no. 9542) were obtained from Cell Signaling
Technology, Inc. A mouse anti-Hsp90α antibody (cat. no. ab128483)
and rabbit anti-Hsp90β antibody (cat. no. ab2927) were purchased
from Abcam. A goat anti-α-tubulin acetyltransferase 1 (αTAT1)
antibody (cat. no. sc-101911) and mouse polyclonal anti-β-actin
antibody (cat. no. sc-47778) were purchased from Santa Cruz
Biotechnology, Inc. Mouse polyclonal anti-acetylated α-tubulin
(Ac-α-tubulin) antibody (cat. no. 2152) and α-tubulin antibody
(cat. no. 3873) were obtained from Cell Signaling Technology, Inc.
Rabbit anti-p-Akt antibody (cat. no. 4060), anti-Akt antibody (cat.
no. 9272) and anti-GSK3β antibody (cat. no. 5676) were purchased
from Cell Signaling Technology, Inc. Protein A-Agarose (cat. no.
05015979001) and Protein G-Agarose (cat. no. 05015952001) were
obtained from Roche Diagnostics.
Cell line and cell culture
The human non-small cell lung cancer cell line,
Calu-1, was provided by the American Type Culture Collection
(ATCC). The cells were grown in RPMI-1640 (Gibco; Thermo Fisher
Scientific, Inc.) with 5% fetal bovine serum in a humidified
atmosphere of 5% CO2 and 95% air at 37°C.
Cell survival assay
First, Calu-1 cells were seeded in 96-well plates at
an appropriate density of 8×103 cells per 100 µl of
culture medium per well. On the second day, Calu-1 cells were
treated with tanespimycin (0, 0.5 and 1.0 µM) or combination with
docetaxel (0.5 nM) and the total volume per well was 200 µl. Calu-1
cells were treated for 48 h, and cell survival was estimated by a
sulforhodamine B (SRB) assay as previously described (20). First, the culture supernatants were
discarded, and 100 µl of 10% TCA (trichloroacetic acid,) were then
added per well. After washing with water thrice, 50 µl SRB were
then added per well by agitating for 5 min. After recycling the SRB
solution and washing with 1% acetic acid 5 times, 100 µl tris-base
(10 mM, pH 10.5) were added per well and followed by agitation for
5 min. The absorbance value was measured by a microplate reader at
540 nm. The relative fold-change of cell survival was determined by
comparing with the control. At least 4 independent experiments were
performed.
Western blot analysis
Whole-cell protein lysate preparation and western
blot analysis were performed as previously described in the
literature (21), in which various
apoptotic proteins were induced and examined by western blot
analysis. Cells were lysed in RIPA buffer and total lysate proteins
were assessed by Biorad Bradford assay (Bio-Rad Laboratoreis,
Inc.). SDS-PAGE (10%) and immunoblotting were performed with 40 µg
of proteins from each sample and proteins were transferred to a
polyvinylidene difluoride membrane (PVDF). The dilution of the
primary antibodies (Cell Signaling Technology) used for probing was
usually 1:1,000. The secondary antibodies were usually diluted at
1:5,000. In the blocking protocol, 5% skim milk was used at room
temperature for 1 h. The primary antibody was incubated at 4°C
overnight and the secondary antibody was incubated at room
temperature for 1 h. Images were developed using ECL reagent (cat.
no. 32109; Thermo Fisher Scientific, Inc.) with an Amersham Imager
600 (GE Healthcare). All immunoblots were repeated thrice, and the
band densities were quantified using ImageJ 1.53a software. β-actin
was used as a loading control.
Silencing of VDAC1, Hsp90α/β and αTAT1
with siRNA oligos
siRNAs were synthesized by GenePharma. The Hsp90α/β
siRNA and VDAC1 siRNA target sequences were synthesized as
previously described (22,23): siRNA transfection (1.5 µl) was
conducted as previously described (24) and the sequences were as follows.
HiPerFect Transfection Reagent was used and at 6 h following
transfection (siRNA, 25 pmol), the cells were cultured in new
medium for 18–24 h and then treated with the given drugs for 48 h.
The sequences were as follows: Control siRNA oligos,
5′-UUCUCCGAACGUGUCACGUTT-3′; VDAC1 siRNA oligos,
5′-GCTTGGTCTAGGACTGGAA-3′; Hsp90α siRNA oligos,
5′-GTTTGAGAACCTCTGCAAA-3′; Hsp90β siRNA oligos,
5′-CGACAAGAAUGAUAAGGCA-3′; and αTAT1 siRNA oligos,
5′-GGGAAACUCACCAGAACGA-3′.
Immunoprecipitation (IP)
Calu-1 cells were lysed with RIPA lysis buffer, and
the supernatant was treated with Protein A-Agarose and Protein
G-Agarose (1:1) and with 10 µg of VDAC1 antibody. The mixture was
rotated at 4°C overnight, and the pelleted beads were washed
according to the manufacturers instructions. The precipitated
proteins were dissolved in 2X SDS sample buffer and boiled for 5
min. The pulled down proteins were then examined by western blot
analysis.
Microscopy
First, Calu-1 cells were seeded in 6-cm culture
plates at an appropriate density (approximately 5.3×105)
and on the second day, various concentrations (0, 0.5 and 1.0 µM)
of tanespimycin were added and the Calu-1 cells were cultured for
the indicated time period (48 h). The Calu-1 cells were then
observed under inverted phase contrast microscope (Nikon TS100) at
×200 magnification.
Statistical analysis
Data from the siRNA experiments of Hsp90, VDAC1 and
αTAT1 were compared with a multiple t-test (Holm-Sidak correction
was applied to the P-values) and the comparison of interest is the
relative protein in Cki (−) treatment. Comparisons between groups
in IP were carried out with an unpaired Student's t-test. The data
are expressed as the means ± standard deviation (SD) and error bars
in the figures represent the SD values. Data from cell survival
assay were analyzed by an unpaired Student's t-test and the
analysis results were the means of 4 independent experiments. Data
from Calu-1 apoptotic cells were calculated using ImageJ software
and comparisons between groups were carried out with repeated
measures (RM) one-way ANOVA and a Geisser-Greenhouse correction was
applied to the P-values obtained from RM one-way ANOVA to control
for comparisons. The analysis results are the means of 4 replicate
determinations and bars standard deviation (SD). Statistical
analyses were performed using GraphPad Prism 8 software (GraphPad
Software, Inc.). P-values of <0.05 were considered to indicate
statistically significant differences.
Results
The molecular chaperone, Hsp90,
mediates the expression of VDAC1
Tanespimycin, an inhibitor of Hsp90, may disrupt the
molecular chaperone activity of Hsp90 and influence cell survival.
It has been found that geldanamycin and its derivative, 17-AAG
(also known as tanespimycin), associate with the mitochondria,
specifically VDAC1, via a hydrophobic interaction (19). The present study revealed that
tanespimycin induced VDAC1 upregulation. The present study wished
to determine whether an association exists the expression of Hsp90
and that of VDAC1, and whether Hsp90 plays a role in modulating
VDAC expression and function to induce cell apoptosis. For this
purpose, Hsp90α/β was silenced by siRNA to decrease the expression
level of Hsp90. Hsp90α/β was knocked down in Calu-1 cells, and the
cells were treated with tanespimycin for 48 h (Fig. 1A). The results of western blot
analysis and GraphPad software statistical analyses revealed that
the expression level of VDAC1 was upregulated in both the untreated
cells in which Hsp90α/β was knocked down and in the cells in which
Hsp90α/β was knocked down and treated with tanespimycin compared
with the control cells (Fig. 1),
suggesting that a reduction in Hsp90 expression or the loss of its
activity may lead to an increase in VDAC1 expression. Thus, Hsp90
may mediate the expression level of VDAC or Hsp90 and VDAC in the
mitochondrial outer membrane and this may be regulated in parallel
or in a coordinated manner.
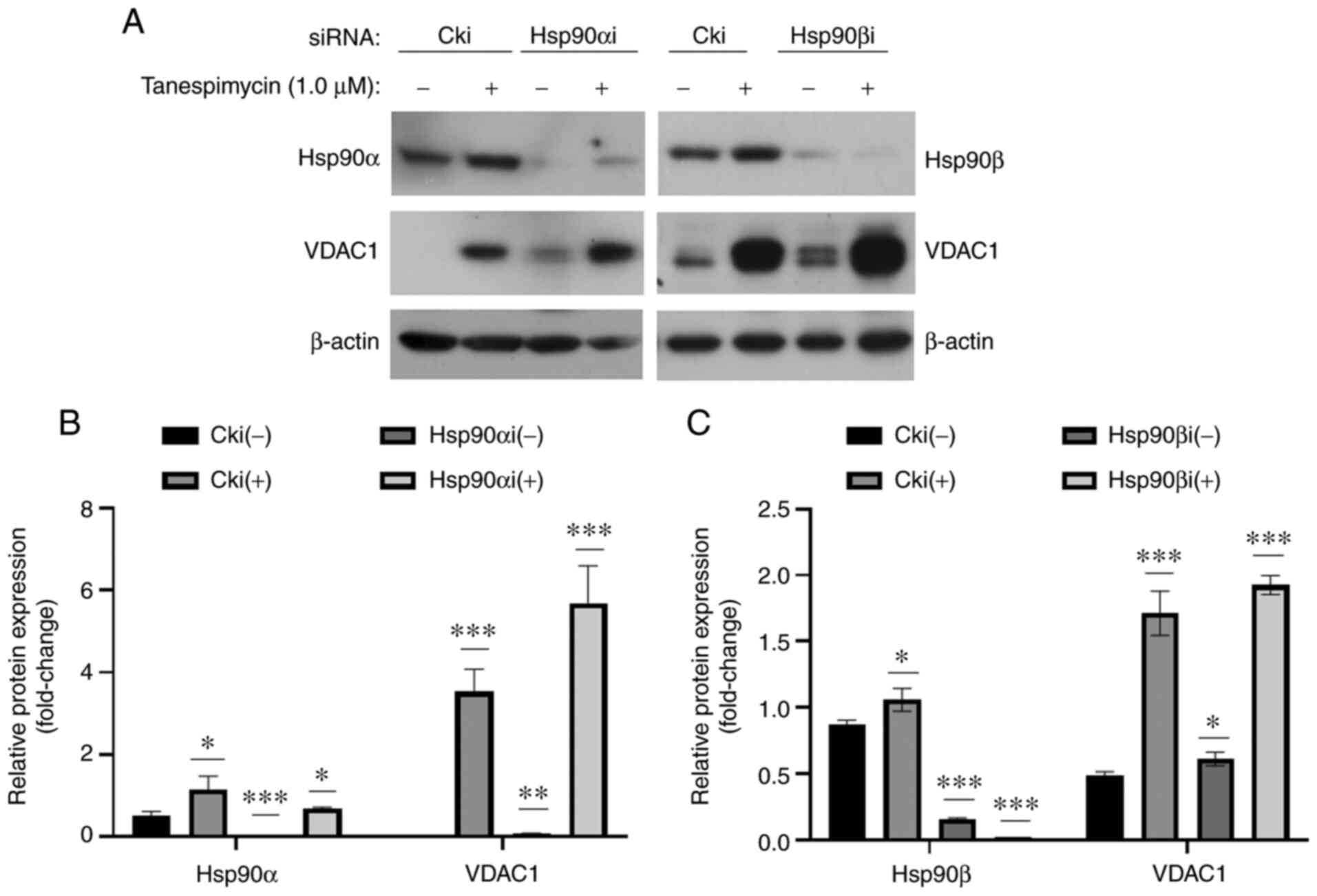 | Figure 1.Hsp90α/β knockdown mediates VDAC1
expression. (A) Calu-1 cells were cultured in 6-well plates and on
the 2nd day they were transfected with control siRNA (Cki) or
Hsp90α/β siRNA (Hsp90αi/βi). At 24 h following transfection, cells
were reseeded in a 6-well plate and treated with 1.0 µM
tanespimycin for 48 h and the cells were then harvested for the
preparation of whole-cell protein lysates for western blot
analysis, and the indicated proteins Hsp90α/β, VDAC1 and β-actin
were analysed. β-actin was used as the loading control (Hsp90α, 90
kDa; Hsp90β, 90 kDa; VDAC1, 32 kDa; β-actin, 45 kDa). (B and C) The
above western blot analysis results were analyzed by ImageJ
software and GraphPad Prism 8 software, and the analysis results
are the means of 3 independent experiments. The western blot shown
is representative of 3 independent experiments. Comparisons between
groups were carried out with the Multiple t-test (Holm-Sidak
correction was applied to the P-values of multiple comparison) and
the comparison of interest is the relative protein in Cki(−)
treatment. Values of P<0.05 were considered to indicate a
statistically significant difference; *P<0.05, **P<0.005 and
***P<0.001 compared with the untreated cells. Error bars in the
figures represent the SD values. Hsp, heat shock protein; VDAC1,
voltage-dependent anion channel 1. |
Upregulation of VDAC1 is involved in
mitochondria-dependent apoptosis
To further elucidate the roles of VDAC1 in lung
cancer cell apoptosis, VDAC1 was silenced by siRNA in Calu-1 cells,
and the cells were then treated with 1.0 µM tanespimycin for 48 h.
The results of western blot analysis revealed that when VDAC1
protein was knocked down, the activation of caspase-8, particularly
a distinct anti-caspase-8-p18 protein band, was clearly inhibited,
and PARP expression was decreased (Fig.
2A and B). VDAC1 is reportedly required for intrinsic apoptosis
through its ability to process procaspase-8 into its active p18
form (25). In the present study,
in the inverted phase contrast microscopy images, Calu-1 cell
apoptosis was evident (P<0.001) and this was also confirmed by
statistical analyses (Fig. 2C and
D). Therefore, the present study demonstrated that VDAC1
upregulation was involved in mitochondria-dependent apoptosis
induced by tanespimycin in lung cancer cells.
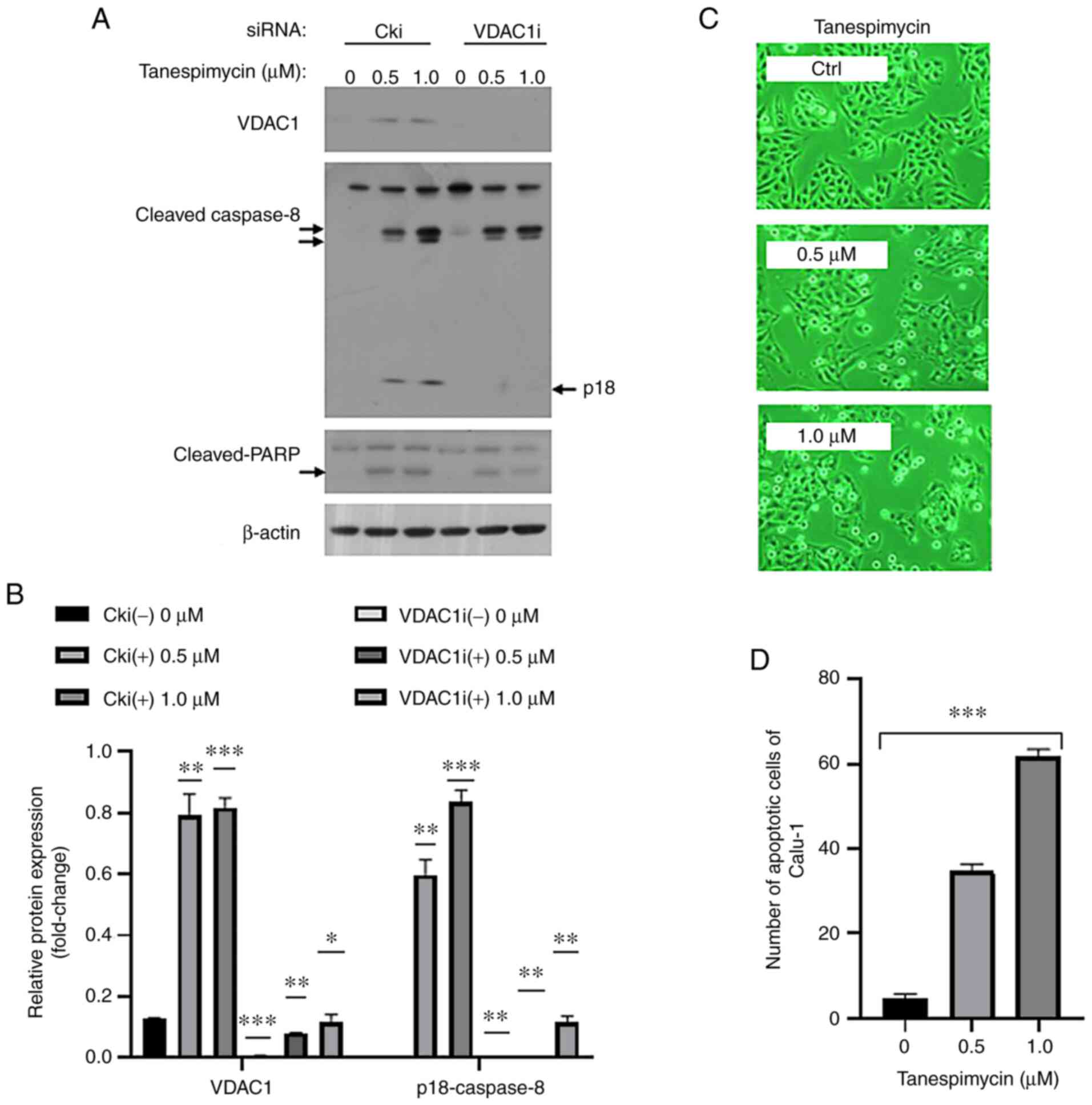 | Figure 2.VDAC1 upregulation induced by
tanespimycin is involved in cell apoptosis. (A) Calu-1 cells were
cultured in 6-well plate and on the 2nd day they were transfected
with control siRNA (Cki) or VDAC1 siRNA (VDAC1i). At 24 h following
transfection, cells were reseeded in 6-well plate and treated with
tanespimycin (0, 0.5 and 1.0 µM) for 48 h. The cells were then
harvested for the preparation of whole-cell protein lysates for
following western blot analysis to detect VDAC1, Caspase-8, PARP
and β-actin levels. β-actin was used as the loading control (VDAC1,
32 kDa; caspase-8, 18, 43, 57 kDa; PARP, 89, 116 kDa; β-actin, 45
kDa). (B) The above western blot analysis results of VDAC1 and
p18-caspase8 were analyzed by ImageJ software and GraphPad Prism 8
software and the analysis results were the mean of 3 independent
experiments. The western blot shown is representative of 3
independent experiments. Comparison betweens groups were carried
out with the Multiple t-test (Holm-Sidak correction was applied to
the P-values of multiple comparison). The comparison of interest is
the relative protein in Cki (−) treatment; *P<0.05, **P<0.005
and ***P<0.001. Error bars in the figures represent the SD
values. (C) Calu-1 cells were treated with tanespimycin (0, 0.5 and
1.0 µM), and then the cells were subjected to take photos by
inverted phase contrast microscope (×200 magnification) to observe
the cell apoptotic state. (D) The number of Calu-1 apoptotic cells
were calculated by ImageJ software. Comparison between groups was
carried out with the RM one-way ANOVA method (a Geisser-Greenhouse
correction was applied to the P-values). ***P<0.001. The
analysis results are the means of 4 replicate determinations; bars
represent SD. P<0.001. VDAC1, voltage-dependent anion channel
1. |
Expression of Ac-α-tubulin induced by
tanespimycin is elevated in apoptosis
Microtubules consist of α/β-tubulin dimers and exert
significant effects on cell growth state and signal transduction.
When the levels of acetylated α/β-tubulin dimers or acetylated
microtubules are increased, their cellular activities are altered
(26). The authors and other
researchers have reported that tanespimycin (17-AAG) induces cell
death via the downregulation of c-FLIPL in lung cancer
cells (20,27). In the present study, it was
simultaneously found that when the Calu-1 cells were exposed to
tanespimycin (1.0 µM for 48 h), the expression of Ac-α-tubulin
induced by tanespimycin evidently increased in a dose-dependent
manner, as detected by western blot analysis (Fig. 3A), and that the cell survival rate
was reduced to approximately 30% of the control level, as
determined by SRB assay (Fig. 3B).
To further determine the functions of tanespimycin-induced
Ac-α-tubulin in apoptosis, Calu-1 cells were treated for 48 h with
a combination of tanespimycin (0, 0.5 and 1.0 µM) and docetaxel
(0.5 nM). As a microtubule stabilizer, docetaxel potentiated the
expression of Ac-α-tubulin and cellular apoptosis induced by
tanespimycin monotherapy, as determined by western blot analysis
and SRB assay, in which the cell survival rate was reduced to 23%
of the control level (Fig. 3A and
B). According to these combination experiments, cellular
apoptosis was induced, as evidenced by a higher degree of caspase-9
and PARP activation, and the expression of Ac-α-tubulin, VDAC and
Bax was elevated (Fig. 3A).
Furthermore, the expression of Ac-α-tubulin was positively
associated with apoptosis, suggesting that the acetylation of
α-tubulin plays important roles in the induction of the apoptosis
of non-small cell lung cancer (NSCLC) cells.
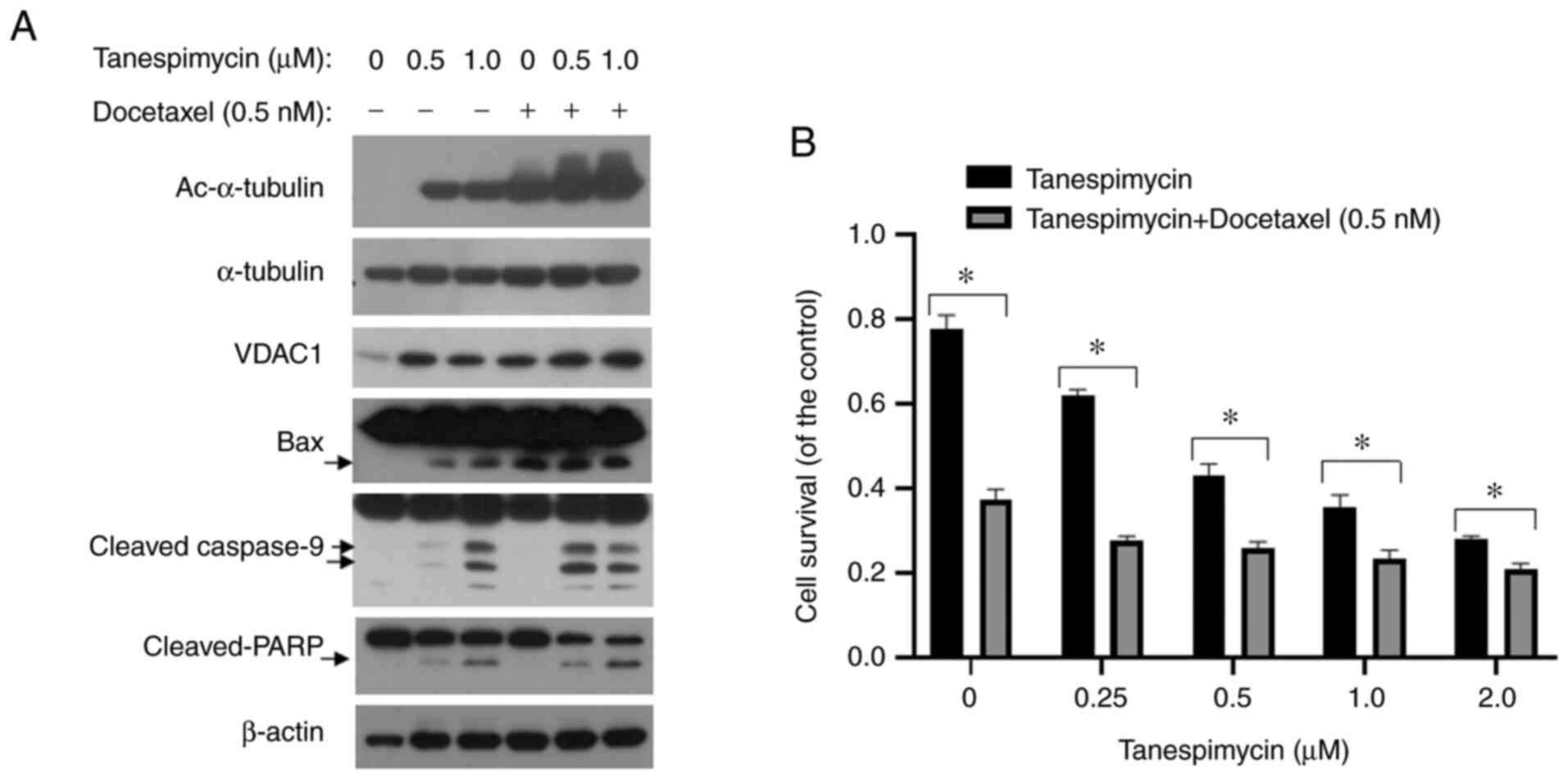 | Figure 3.Docetaxel enhances the upregulation
of VDAC1, Ac-α-tubulin and cellular apoptosis induced by
tanespimycin. (A) Calu-1 cells were treated with the given
concentrations of tanespimycin (0, 0.5 and 1.0 µM) or in
combination with docetaxel (0.5 nM) for 48 h, and the the cells
were then subjected to the preparation of whole-cell protein
lysates and the given proteins Ac-α-tubulin, α-tubulin, VDAC1, Bax,
caspase-9, PARP and β-actin were detected by western blot analysis.
β-actin was used as the loading control (Ac-α-tubulin, 52 kDa;
α-tubulin, 52 kDa; VDAC1, 32 kDa; Bax, 20 kDa; caspase-3, 17, 19,
35 kDa; PARP, 89, 116 kDa; β-actin, 45 kDa). (B) Calu-1 cells were
seeded in 96-well plate and on the 2nd day treated with the given
concentrations of tanespimycin (0, 0.25, 0.5, 1.0 and 2.0 µM) and
docetaxel (0.5 nM) for 48 h. Cell number was estimated by SRB assay
for calculation of cell survival. Comparison between groups was
carried out with an unpaired Student's t-test. The analysis results
were the mean of 4 replicate determinations; bars, SD. *P<0.05.
VDAC1, voltage-dependent anion channel 1; Ac-α-tubulin, acetylated
α-tubulin. |
Acetylation level of α-tubulin is
modulated by the acetyltransferase, αTAT1
The acetyltransferase, αTAT1, has been reported to
catalyse α-tubulin acetylation at Lys40 inside the microtubule
lumen to alter microtubule-based processes (28). The acetylation level of microtubules
(MTs) influences their stability and can even render MTs
inflexible, brittle, and more inclined to break under stress
(29). Thus, the present study
wished to determine whether Ac-α-tubulin induced by tanespimycin
resulted from the breakage of acetylated MTs or the acetylation of
free α/β-tubulin and to identify the association between α-tubulin
acetylation and the acetyltransferase, αTAT1. To determine whether
the acetyltransferase αTAT1 is the enzyme that catalyses the
tanespimycin-induced acetylation of α-tubulin in Calu-1 lung cancer
cells, a time-course experiment and a siRNA experiment were
performed. In the time-course experiment, the expression levels of
αTAT1 were consistent with the increasing levels of Ac-α-tubulin
induced by tanespimycin (Fig. 4A).
When αTAT1 was knocked down, the expression level of Ac-α-tubulin
decreased simultaneously compared with the control treated with
tanespimycin, as shown by western blot analysis and GraphPad
software analyses (Fig. 4B and C).
These results indicate that αTAT1 is the acetyltransferase of
α-tubulin in lung cancer cells and that it may regulate the levels
of acetylated α-tubulin.
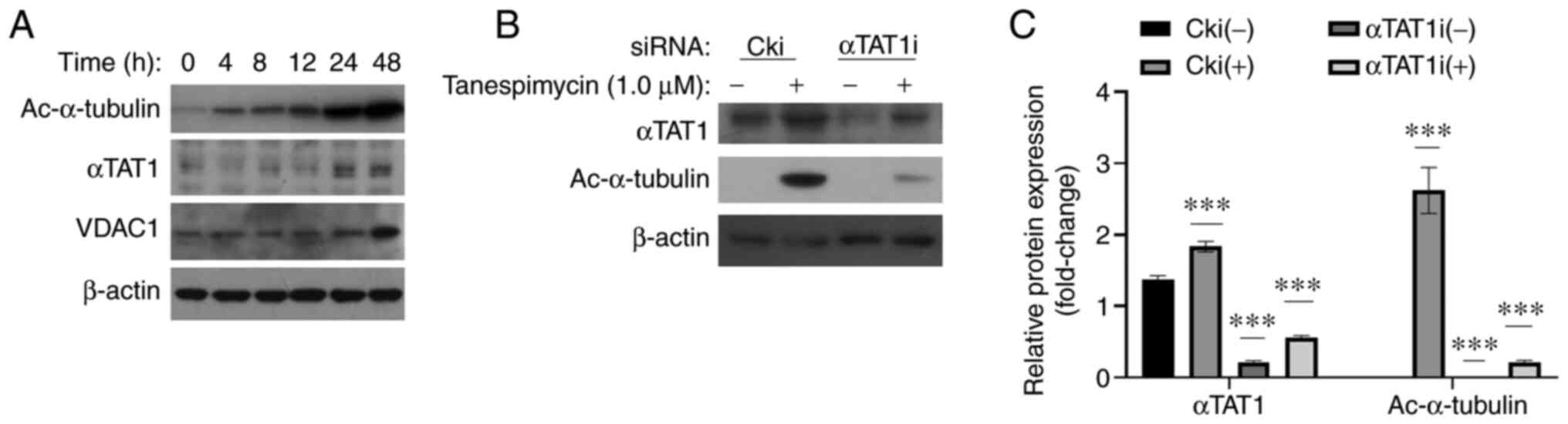 | Figure 4.Expression level of Ac-α-tubulin is
modulated by the acetyltransferase, αTAT1. (A) Calu-1 cells were
treated with 1.0 µM tanespimycin for the indicated periods of time
(0, 4, 8, 12, 24 and 48 h); the cells were then subjected to the
preparation of whole-cell protein lysates and the given proteins
Ac-α-tubulin, αTAT1, VDAC1 and β-actin were detected by western
blot analysis. (B) Calu-1 cells were cultured in 6-well plate and
on the 2nd day they were transfected with control siRNA (Cki) or
αTAT1 siRNA (αTAT1i). At 24 h following transfection, cells were
treated with 1.0 µM tanespimycin for 48 h. The cells were then
harvested for preparation of whole-cell protein lysates for
following western blot analysis to detect the expression levels of
the given proteins Ac-α-tubulin, αTAT1 and β-actin. β-actin was
used as the loading control (αTAT1, 43 kDa; Ac-α-tubulin, 52 kDa;
VDAC1, 32 kDa; β-actin, 45 kDa). (C) The above western blot results
of αTAT1 and Ac-α-tubulin were analyzed by ImageJ software and
GraphPad Prism 8 software and the analysis results are the means of
3 independent experiments. The western blot shown is representative
of 3 independent experiments. Comparisons between groups were
carried out with Multiple t-test (Holm-Sidak correction was applied
to the P-values of multiple comparison) and the comparison of
interest is the relative protein in Cki (−) treatment.
***P<0.001 compared with the untreated cells. Error bars in the
figures represent the SD values. VDAC1, voltage-dependent anion
channel 1; Ac-α-tubulin, acetylated α-tubulin; αTAT1, α-tubulin
acetyltransferase 1. |
Acetylation of α-tubulin promotes
VDAC1 permeability in mitochondria-mediated apoptosis
In the above studies, we also observed that VDAC1
expression was elevated in both the time-course experiment and in
the drug combination experiments and was accompanied by an increase
in Ac-α-tubulin (Figs. 3A and
4A). The present study then wished
to determine the functions of both proteins in apoptosis. In the
tubulin-VDAC1 interaction model, the anionic C-terminal tail of
tubulin penetrates into the VDAC1 cationic channel lumen and
interacts with VDAC1, thus blocking channel conductance (17). In the present study, the acetylation
of α-tubulin was markedly induced, and this change was accompanied
by lung cancer cell apoptosis. Therefore, it was hypothesized that
the acetylation of α-tubulin in microtubules, particularly the
acetylation of α/β dimers, would decrease the effective level of
free tubulin in the MOM and that the density of the C-terminal tail
of α-tubulin, which interacts with VDAC1, would be reduced in the
interior β-barrel of VDAC1. To examine this hypothesis, Calu-1
cells were treated with 1.0 µM tanespimycin for 32 h (not for 48 h
as treatment for this duration induces apoptosis that is overly
severe), and an IP experiment was then conducted. It was observed
that α-tubulin and Ac-α-tubulin were co-precipitated in the IP
complexes, as detected by western blot analysis (Fig. 5A). However, in the IP complexes, 2
distinct (136 and 185 kDa) protein bands were clearly detected by
an anti-VDAC1 antibody and were found to correspond to a tetramer
and hexamer of VDAC1. However, there were no bands that represented
VDAC1 monomers (32 kDa). The level of VDAC1 was not apparently
increased in either the lysate or the IP complex between the
tanespimycin-treated cells and the control cells (not treated with
tanespimycin) (Fig. 5A), while the
level of α-tubulin was slightly decreased in either the lysate or
the IP complex in the treated cells compared to the untreated
cells. Based on the results of IP analyzed by GraphPad software
(Fig. 5B), following treatment with
tanespimycin for 32 h, the level of VDAC1 in the IP complex was not
increased. This may have been because 32 h pf tanespimycin
treatment was not sufficient to induce VDAC1 upregulation, although
it was sufficient to cause a shift from the monomeric form to the
oligomeric form and to induce VDAC1 permeability and cell apoptosis
(Fig. 5A and B).
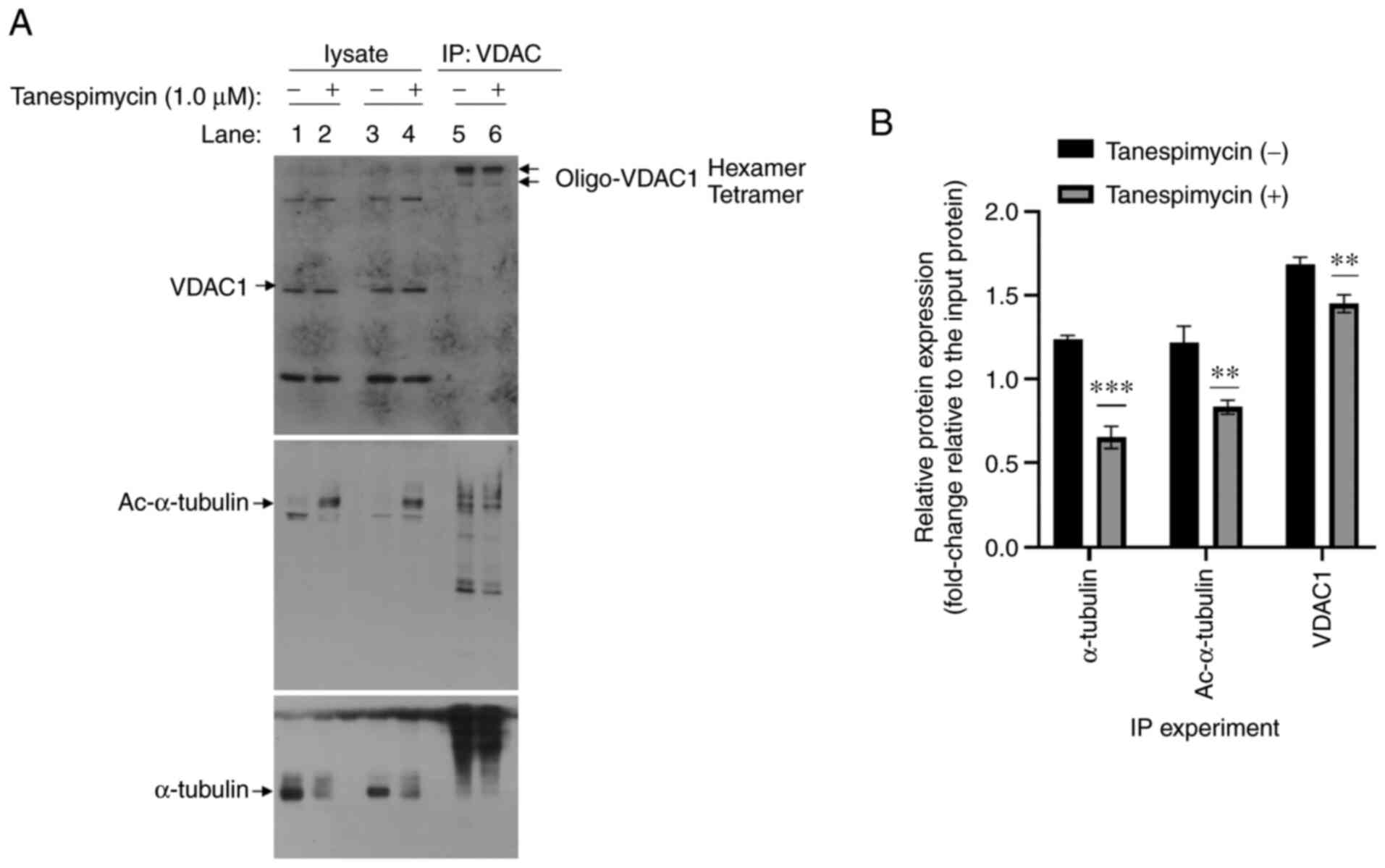 | Figure 5.α-tubulin interacts with VDAC1 in
lung cancer apoptosis induced by tanespimycin. (A) Calu-1 cells
were treated with 1.0 µM tanespimycin for 32 h and cell lysates
were then subjected to immunoprecipitation with anti-VDAC1
antibodies and Protein-A/G Agarose (1:1) and the complexes were
incubated at 4°C overnight. The precipitated proteins and the
whole-cell protein lysates were isolated by SDS-PAGE and detected
by western blot analysis with anti-α-tubulin, anti-Ac-α-tubulin and
anti-VDAC1 antibodies (lanes 1, 2, 3, 4 represent the whole cell
lysate samples; lane 5 and 6 represent the IP samples; lanes 2, 4
and 6 represent samples treated with by 1.0 µM tanespimycin for 32
h; Ac-α-tubulin, 52 kDa; α-tubulin, 52 kDa; VDAC1, 32 kDa). (B) The
above western blot analysis results of α-tubulin, Ac-α-tubulin and
VDAC1 in the IP experiments were analyzed respectively using ImageJ
software and GraphPad Prism 8 software and the analysis results are
the means of 3 independent experiments. Comparisons between groups
was carried out with an unpaired Student's t-test. **P<0.005 and
***P<0.001 compared with untreated cells. Error bars in the
figures represent the SD values. VDAC1, voltage-dependent anion
channel 1; Ac-α-tubulin, acetylated α-tubulin. |
It has been shown that GSK3β can phosphorylate VDAC
and induce VDAC opening. GSK3β and VDAC are two direct substrates
of the kinase, AKT. Phosphorylated AKT (p-AKT) can promote GSK3β
and VDAC phosphorylation; however, phosphorylated GSK3β does not
catalyse VDAC phosphorylation, resulting in mitochondrial
permeability blockade (14). To
further examine the state of VDAC1 permeability, the present study
performed dose-dependent experiments and it was found that the
increase in GSK3β (total) levels paralleled the upregulation of
acetylated α-tubulin and VDAC1. At the same time, it was observed
that the p-AKT levels were decreased in Calu-1 cells (Fig. 6A).
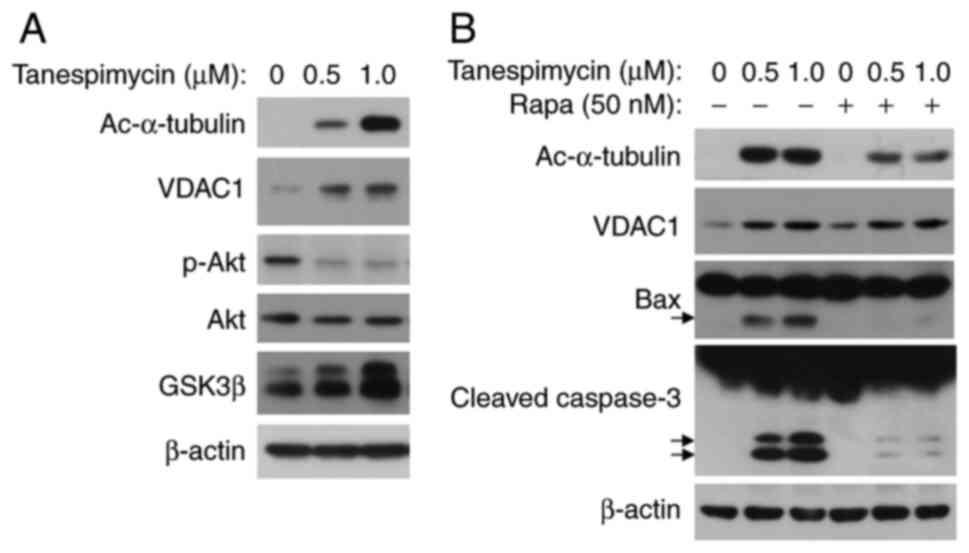 | Figure 6.p-Akt/GSK3β signaling pathway
modulates the phosphorylation of VDAC1. (A) Calu-1 cells were
treated with the given concentrations of tanespimycin (0, 0.5 and
1.0 µM) for 48 h, and the cells were then harvested for the
preparation of whole-cell protein lysates for following western
blot analysis to detect the expression levels of the given
proteins, Ac-α-tubulin, VDAC1, p-Akt, Akt, GSK3β and β-actin. (B)
Calu-1 cell lines were treated with the given concentrations of
tanespimycin (0, 0.5 and 1.0 µM) or in combination with rapamycin
(50 nM) for 48 h and the cells were then subjected to the
preparation of whole-cell protein lysates, and the given proteins
Ac-α-tubulin, VDAC, Bax, caspase 3 and β-actin were detected by
western blot analysis. β-actin was used as the loading control
(Ac-α-tubulin, 52 kDa; VDAC1, 32 kDa; p-Akt, 60 kDa; Akt, 60 kDa;
GSK3β, 46 kDa; Bax, 20 kDa; caspase-9, 35, 37, 47 kDa; PARP, 89,
116 kDa; β-actin, 45 kDa). VDAC1, voltage-dependent anion channel
1; Ac-α-tubulin, acetylated α-tubulin; GSK3β, glycogen synthase
kinase 3β. |
To further elucidate the mechanisms underlying the
role of VDAC1 in apoptosis, tanespimycin (0, 0.5 and 1.0 µM) was
used in combination with rapamycin (50 nM) for 48 h, an mTOR
inhibitor that has been reported to promote cell survival (30). The results revealed that the
combination of tanespimycin and rapamycin markedly reduced the
levels of Ac-α-tubulin and Bax, and partially inhibited caspase-3
activation (Fig. 6B). In the
combination experiment, VDAC1 expression was not reduced, which
suggests that VDAC expression is not affected by rapamycin
treatment and that rapamycin has a greater influence on α-tubulin
acetylation and cell death. That is, acetylation of α-tubulin is
related to the mTOR signalling pathway. According to the
above-mentioned results, an increase in Ac-α-tubulin expression
induced by tanespimycin weakens the interaction between VDAC1 and
tubulin, and results in an increase in MOM permeability and
cellular apoptosis through the intrinsic apoptotic pathway.
Discussion
The present study demonstrated that tanespimycin
induced α-tubulin acetylation, VDAC1 upregulation and cell death,
during which the conformational structure of VDAC1 may shift from
the monomeric form to the oligomeric form. The quantity of
acetylation of α-tubulin may decrease the level of free α-tubulin
that interacts with VDAC1 and induce VDAC1 permeability to release
proapoptotic materials from the mitochondria. It is well known that
the mitochondria are cell organelles in which ATP and intermediates
are produced for energy, and mitochondria play a crucial role in
deciding cell fate for survival or apoptosis (1,31). The
permeability of VDAC1, as a major protein in the mitochondrial
outer membrane, is modulated by a variety of factors, and the
switch of the VDAC1 channel is also influenced by its
conformational structure (32–34).
VDAC1 has been reported to be presented as a monomer or oligomer
during apoptosis induction (6).
This finding is consistent with the results of the present study.
The overexpression of VDAC1 induced by tanespimycin may alter the
balance between its monomeric and oligomeric forms, and is
conducive to the formation of oligomers, as previously reported
(35).
VDAC1 can also form hetero-oligomers with Bax or
Bak, which are two essential mediators of apoptosis (36,37).
In the present study, it was also found that Bax expression was
elevated along with cellular apoptosis. However, it is not known
whether VDAC1 forms hetero-oligomers with Bax, and this warrants
further investigation. Studies have confirmed that VDAC1
overexpression triggers the oligomerization of VDAC1 with itself or
with other proteins, resulting in cellular apoptosis (38). When encountering apoptosis signals,
Bax translocates to the mitochondria to increase the VDAC1 pore
size, which leads to the permeabilization of the MOM (39,40).
In the present study, tanespimycin not only inhibited Hsp90
biological activity, but also induced an increase in VDAC1
expression. Indeed, it was found that the upregulation of VDAC1 and
Bax was associated with an increase in tanespimycin concentration.
VDAC1, Bax and Bak are all localized in the mitochondrial outer
membrane. Another study demonstrated that VDAC1 knockdown inhibited
Bax activation, but not Bak activation in CDDP-induced cellular
apoptosis (12). The present study
also revealed that Bax (not Bak) was activated following elevation
of VDAC1 expression (Fig. 3A).
Therefore, it was hypothesized that VDAC1 and Bax may form
homo-oligomers or hetero-oligomers to generate channels that are
large enough to allow CytC release from the mitochondria, inducing
cellular apoptosis.
The acetylation of α-tubulin during apoptosis is
controversial at present as it is unclear whether α-tubulin
acetylation is the cause or an effect of apoptosis. In the present
study, the combination of tanespimycin and docetaxel further
promoted α-tubulin acetylation accompanied by VDAC1 and Bax
elevation, implying that apoptosis induced by tanespimycin is
associated with both microtubules and mitochondria. Docetaxel is an
anti-tumour drug that inhibits microtubule depolymerization by
binding to tubulin, which leads to an apparent decrease in free
tubulin and induces cancer cell apoptosis. To further evaluate the
association between apoptosis and increases in α-tubulin
acetylation and VDAC1, tanespimycin was used in combination with
rapamycin. The results revealed that apoptosis was alleviated, as
indicated by a marked reduction in cleaved-caspase-3 expression,
and that α-tubulin acetylation was simultaneously decreased,
although that the change in VDAC1 expression was minimal. These
results demonstrate that there is a positive association between
cellular apoptosis and α-tubulin acetylation induced by
tanespimycin.
Protein acetylation is a reversible
post-translational modification that is modulated by
acetyltransferase and deacetylase. The present study demonstrated
that the acetylation levels of α-tubulin are consistent with the
expression of the acetyltransferase, αTAT1. The depletion of the
tubulin acetyltransferase, αTAT1, has been reported to lead to an
apparent increase in the frequency of microtubule disruption, which
indicates that acetylation is necessary to maintain the persistence
of long-lived microtubules (41,42).
Surprisingly, in the present study, Hsp90 inhibition by
tanespimycin induced the acetylation of α-tubulin acetylation
(43), and also simultaneously
caused cellular apoptosis and the enhanced expression of VDAC1 and
Bax. However, the present study wished to determine the origin of
this Ac-α-tubulin, namely to determine whether it is the result of
acetylated microtubule breakage or the acetylation of free
α-tubulin. It also needs to be determined whether the level of free
α-tubulin that interacts with VDAC1 was influenced. Based on the
results of the present study, it was concluded that Hsp90 may
mediate the expression level of VDAC1 and that VDAC1 plays its
normal physiological roles in the MOM under normal conditions.
However, when Hsp90 activity is disrupted by tanespimycin or when
Hsp90 levels are reduced, higher VDAC1 expression is induced, which
may result in VDAC permeability through the formation of
homo-oligomers or hetero-oligomers by VDAC and Bax. Based on the
tubulin-VDAC interaction model and the current IP experiments, it
was elucidated that abundant acetylation of α-tubulin may reduce
the level of free α-tubulin or α/β tubulin dimers in the MOM and
subsequently affect VDAC permeability. The level of free α-tubulin
or α/β tubulin dimers (although the level of β tubulin was not
determined) in the MOM affected VDAC1 permeability, as evidenced by
a decrease in p-AKT levels, which indicates that this interaction
may be mediated by the AKT/GSK3β signalling pathway.
Hexose kinase (HK) is an important protein that
interacts with VDAC1, and HK-I and HK-II, which are overexpressed
in many types of cancers, control the rates of tumour growth and
migration. We suspect that tanespimycin can compete with HK to bind
to VDAC1. This may be an interesting area of future study for
identifying the association between mitochondria-mediated apoptosis
and the Warburg effect.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from
doctoral scientific research funds (318051315), the Science and
Technology Planning Project for Colleges and Universities in
Shandong Province (J17KA233), and the National Natural Science
Foundation of China (30971479 and 31071215).
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
QW and XL designed the study. QW conducted the
experiments, analysed the data and drafted the manuscript. XL was
involved in the interpretation of the data. Both authors
contributed to, and read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Ac-α-tubulin
|
acetylated α-tubulin
|
|
ANT
|
adenine nucleotide translocase
|
|
GSK3β
|
glycogen synthase kinase
3β;
|
|
HDAC6
|
histone deacetylase 6
|
|
HK
|
hexose kinase
|
|
Hsp27
|
heat shock protein 27
|
|
Hsp90
|
heat shock protein 90
|
|
IP
|
immunoprecipitation
|
|
LC3B-II
|
microtubule-associated protein 1 light
chain 3B-II
|
|
MOM
|
mitochondrial outer membrane
|
|
PARP
|
poly ADP-ribose polymerase
|
|
PKA
|
protein kinase A
|
|
PTP
|
permeability transition pore
|
|
SRB
|
sulforhodamine B
|
|
VDAC1
|
voltage-dependent anion channel 1
|
|
αTAT1
|
α-tubulin acetyltransferase 1
|
References
|
1
|
Shoshan-Barmatz V, Krelin Y and Chen Q:
VDAC1 as a player in mitochondria-mediated apoptosis and target for
modulating apoptosis. Curr Med Chem. 24:4435–4446. 2017.PubMed/NCBI
|
|
2
|
Bayrhuber M, Meins T, Habeck M, Becker S,
Giller K, Villinger S, Vonrhein C, Griesinger C, Zweckstetter M and
Zeth K: Structure of the human voltage-dependent anion channel.
Proc Natl Acad Sci USA. 105:15370–15375. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hiller S, Garces RG, Malia TJ, Orekhov VY,
Colombini M and Wagner G: Solution structure of the integral human
membrane protein VDAC-1 in detergent micelles. Science.
321:1206–1210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maldonado EN, Sheldon KL, DeHart DN,
Patnaik J, Manevich Y, Townsend DM, Bezrukov SM, Rostovtseva TK and
Lemasters JJ: Voltage-dependent anion channels modulate
mitochondrial metabolism in cancer cells: Regulation by free
tubulin and erastin. J Biol Chem. 288:11920–11929. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shuvo SR, Ferens FG and Court DA: The
N-terminus of VDAC: Structure, mutational analysis, and a potential
role in regulating barrel shape. Biochim Biophys Acta.
1858:1350–1361. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Keinan N, Tyomkin D and Shoshan-Barmatz V:
Oligomerization of the mitochondrial protein voltage-dependent
anion channel is coupled to the induction of apoptosis. Mol Cell
Biol. 30:5698–5709. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tian M, Xie Y, Meng Y, Ma W, Tong Z, Yang
X, Lai S, Zhou Y, He M and Liao Z: Resveratrol protects
cardiomyocytes against anoxia/reoxygenation via dephosphorylation
of VDAC1 by Akt-GSK3 β pathway. Eur J Pharmacol. 843:80–87. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heslop KA, Rovini A, Hunt EG, Fang D,
Morris ME, Christie CF, Gooz MB, DeHart DN, Dang Y, Lemasters JJ
and Maldonado EN: JNK activation and translocation to mitochondria
mediates mitochondrial dysfunction and cell death induced by VDAC
opening and sorafenib in hepatocarcinoma cells. Biochem Pharmacol.
171:1137282020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Abu-Hamad S, Zaid H, Israelson A, Nahon E
and Shoshan-Barmatz V: Hexokinase-I protection against apoptotic
cell death is mediated via interaction with the voltage-dependent
anion channel-1: Mapping the site of binding. J Biol Chem.
283:13482–13490. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shoshan-Barmatz V, Ben-Hail D, Admoni L,
Krelin Y and Tripathi SS: The mitochondrial voltage-dependent anion
channel 1 in tumor cells. Biochim Biophys Acta. 1848:2547–2575.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mazure NM: VDAC in cancer. Biochim Biophys
Acta Bioenerg. 1858:665–673. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tajeddine N, Galluzzi L, Kepp O, Hangen E,
Morselli E, Senovilla L, Araujo N, Pinna G, Larochette N, Zamzami
N, et al: Hierarchical involvement of Bak, VDAC1 and Bax in
cisplatin-induced cell death. Oncogene. 27:4221–4232. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Matsubara H, Tanaka R, Tateishi T, Yoshida
H, Yamaguchi M and Kataoka T: The human Bcl-2 family member
Bcl-rambo and voltage-dependent anion channels manifest a genetic
interaction in Drosophila and cooperatively promote the activation
of effector caspases in human cultured cells. Exp Cell Res.
381:223–234. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pastorino JG, Hoek JB and Shulga N:
Activation of glycogen synthase kinase 3beta disrupts the binding
of hexokinase II to mitochondria by phosphorylating
voltage-dependent anion channel and potentiates
chemotherapy-induced cytotoxicity. Cancer Res. 65:10545–10554.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Head SA, Shi W, Zhao L, Gorshkov K,
Pasunooti K, Chen Y, Deng Z, Li RJ, Shim JS, Tan W, et al:
Antifungal drug itraconazole targets VDAC1 to modulate the
AMPK/mTOR signaling axis in endothelial cells. Proc Natl Acad Sci
USA. 112:E7276–E7285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rostovtseva TK, Sheldon KL, Hassanzadeh E,
Monge C, Saks V, Bezrukov SM and Sackett DL: Tubulin binding blocks
mitochondrial voltage-dependent anion channel and regulates
respiration. Proc Natl Acad Sci USA. 105:18746–18751. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rovini A: Tubulin-VDAC interaction:
Molecular basis for mitochondrial dysfunction in
chemotherapy-induced peripheral neuropathy. Front Physiol.
10:6712019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shoshan-Barmatz V, Mizrachi D and Keinan
N: Oligomerization of the mitochondrial protein VDAC1: From
structure to function and cancer therapy. Prog Mol Biol Transl Sci.
117:303–334. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xie Q, Wondergem R, Shen Y, Cavey G, Ke J,
Thompson R, Bradley R, Daugherty-Holtrop J, Xu Y, Chen E, et al:
Benzoquinone ansamycin 17AAG binds to mitochondrial
voltage-dependent anion channel and inhibits cell invasion. Proc
Natl Acad Sci USA. 108:4105–4110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Q, Sun W, Hao X, Li T, Su L and Liu
X: Down-regulation of cellular FLICE-inhibitory protein (Long Form)
contributes to apoptosis induced by Hsp90 inhibition in human lung
cancer cells. Cancer Cell Int. 12:542012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu X, Yue P, Zhou Z, Khuri FR and Sun SY:
Death receptor regulation and celecoxib-induced apoptosis in human
lung cancer cells. J Natl Cancer Inst. 96:1769–1780. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chatterjee M, Jain S, Stühmer T, Andrulis
M, Ungethüm U, Kuban RJ, Lorentz H, Bommert K, Topp M, Krämer D, et
al: STAT3 and MAPK signaling maintain overexpression of heat shock
proteins 90alpha and beta in multiple myeloma cells, which
critically contribute to tumor-cell survival. Blood. 109:720–728.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abu-Hamad S, Sivan S and Shoshan-Barmatz
V: The expression level of the voltage-dependent anion channel
controls life and death of the cell. Proc Natl Acad Sci USA.
103:5787–5792. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Su L, Liu G, Hao X, Zhong N, Zhong D, Liu
X and Singhal S: Death receptor 5 and cellular FLICE-inhibitory
protein regulate pemetrexed-induced apoptosis in human lung cancer
cells. Eur J Cancer. 47:2471–2478. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chacko AD, Liberante F, Paul I, Longley DB
and Fennell DA: Voltage dependent anion channel-1 regulates death
receptor mediated apoptosis by enabling cleavage of caspase-8. BMC
Cancer. 10:3802010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li L and Yang XJ: Tubulin acetylation:
Responsible enzymes, biological functions and human diseases. Cell
Mol Life Sci. 72:4237–4255. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park MA, Zhang G, Mitchell C, Rahmani M,
Hamed H, Hagan MP, Yacoub A, Curiel DT, Fisher PB, Grant S and Dent
P: Mitogen-activated protein kinase kinase 1/2 inhibitors and
17-allylamino-17-demethoxygeldanamycin synergize to kill human
gastrointestinal tumor cells in vitro via suppression of c-FLIP-s
levels and activation of CD95. Mol Cancer Ther. 7:2633–2648. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shida T, Cueva JG, Xu Z, Goodman MB and
Nachury MV: The major alpha-tubulin K40 acetyltransferase alphaTAT1
promotes rapid ciliogenesis and efficient mechanosensation. Proc
Natl Acad Sci USA. 107:21517–21522. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Eshun-Wilson L, Zhang R, Portran D,
Nachury MV, Toso DB, Löhr T, Vendruscolo M, Bonomi M, Fraser JS and
Nogales E: Effects of α-tubulin acetylation on microtubule
structure and stability. Proc Natl Acad Sci USA. 116:10366–10371.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
No M, Choi EJ and Kim IA: Targeting HER2
signaling pathway for radiosensitization: Alternative strategy for
therapeutic resistance. Cancer Biol Ther. 8:2351–2361. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Burke PJ: Mitochondria, bioenergetics and
apoptosis in cancer. Trends Cancer. 3:857–870. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Krüger V, Becker T, Becker L,
Montilla-Martinez M, Ellenrieder L, Vögtle FN, Meyer HE, Ryan MT,
Wiedemann N, Warscheid B, et al: Identification of new channels by
systematic analysis of the mitochondrial outer membrane. J Cell
Biol. 216:3485–3495. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reif MM, Fischer M, Fredriksson K, Hagn F
and Zacharias M: The N-terminal segment of the voltage-dependent
anion channel: A possible membrane-bound intermediate in pore
unbinding. J Mol Biol. 431:223–243. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Böhm R, Amodeo GF, Murlidaran S, Chavali
S, Wagner G, Winterhalter M, Brannigan G and Hiller S: The
structural basis for low conductance in the membrane protein VDAC
upon β-NADH binding and voltage gating. Structure. 28:206–214.e4.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Urbani A, Giorgio V, Carrer A, Franchin C,
Arrigoni G, Jiko C, Abe K, Maeda S, Shinzawa-Itoh K, Bogers JFM, et
al: Purified F-ATP synthase forms a Ca2+-dependent
high-conductance channel matching the mitochondrial permeability
transition pore. Nat Commun. 10:43412019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cosentino K and García-Sáez AJ: Bax and
Bak pores: Are we closing the circle? Trends Cell Biol. 27:266–275.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chin HS, Li MX, Tan IKL, Ninnis RL, Reljic
B, Scicluna K, Dagley LF, Sandow JJ, Kelly GL, Samson AL, et al:
VDAC2 enables BAX to mediate apoptosis and limit tumor development.
Nat Commun. 9:49762018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zalk R, Israelson A, Garty ES,
Azoulay-Zohar H and Shoshan-Barmatz V: Oligomeric states of the
voltage-dependent anion channel and cytochrome c release from
mitochondria. Biochem J. 386:73–83. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rosano C: Molecular model of hexokinase
binding to the outer mitochondrial membrane porin (VDAC1):
Implication for the design of new cancer therapies. Mitochondrion.
11:513–519. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Scharstuhl A, Mutsaers HA, Pennings SW,
Russel FG and Wagener FA: Involvement of VDAC, Bax and ceramides in
the efflux of AIF from mitochondria during curcumin-induced
apoptosis. PLoS One. 4:e66882009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu Z, Schaedel L, Portran D, Aguilar A,
Gaillard J, Marinkovich MP, Théry M and Nachury MV: Microtubules
acquire resistance from mechanical breakage through intralumenal
acetylation. Science. 356:328–332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Taschner M, Vetter M and Lorentzen E:
Atomic resolution structure of human α-tubulin acetyltransferase
bound to acetyl-CoA. Proc Natl Acad Sci USA. 109:19649–19654. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang Q and Liu X: The dual functions of
α-tubulin acetylation in cellular apoptosis and autophage induced
by tanespimycin in lung cancer cells. Cancer Cell Int. 20:3692020.
View Article : Google Scholar : PubMed/NCBI
|














