Introduction
Acute myeloid leukemia (AML) is a common hematologic
malignancy with a statistical incidence rate of 38 case per million
individuals in the United States between 2001 and 2007 (1). In the last 20 years, no significant
improvement has been achieved in the prognosis or long-term
outcomes of AML, and a considerable number of patients still
experience induction failure or relapse after complete remission
(CR) (2). The interaction between
tumor cells and the tumor microenvironment is one of the key
mechanisms of chemoresistance in tumor cells, including AML
(3,4). Bone marrow (BM) mesenchymal stem cells
(BMMSCs) serve an important role in protecting AML cells from
chemotherapy-induced apoptosis (5,6).
Previous studies have reported that tumor cells recruit BMMSCs by
secreting cytokines or chemokines, and culturing BMMSCs to become
cancer-related fibroblasts (CAFs) supports the progression of
malignant cells (7,8). CAFs have been considered as central
components in the tumor microenvironment and serve vital roles in
tumor features, such as proliferation, angiogenesis, invasion and
metastasis (9–11). However, the exact origin of CAFs
remains unknown (12).
BMMSCs have recently been recognized as one of the
origins of CAFs (13,14). However, a precise molecular
definition of CAFs has not yet been elucidated. Previous studies
have revealed that CAFs express cell surface markers, including
fibroblast activation protein (FAPα) (15). Across a wide range of human cancer
types, such as gastric carcinoma, breast cancer and colon cancer,
the expression of FAPα has been reported to correlate with a higher
tumor grade and worse overall survival in solid tumors (16–20).
Considering the important role of FAPα in tumor progression and its
rare expression in healthy tissues, FAPα has become a key target in
tumor therapy (21,22). Previously, it was observed that
tumor cells can reprogram BMMSCs to evolve into CAFs, thereby
further promoting the progression of hematological malignancies
(23). The expression of FAPα in
BMMSCs moderately increases the number of tumor cells in
conditioned medium from myeloma cells and those co-cultured with
multiple myeloma (MM) cells (24).
Furthermore, knockdown of FAPα with small interfering (si)RNA
decreases the protection of bortezomib-induced apoptosis in MM
cells (24). However, it remains
unknown whether the expression of FAPα in BMMSCs is different
between patients with AML and healthy donors, nor has it been
elucidated whether FAPα serves an important role in mediating AML
cell features. Therefore, the aim of the present study was to
investigate the expression of FAPα in BMMSCs and BM, as well as to
identify the role of FAPα in BMMSCs in protecting AML cells from
apoptosis.
Materials and methods
Cell culture
The human AML cell line, Kasumi-1, was obtained from
the Department of Cancer Biology, Lerner Research Institute,
Cleveland Clinic. Cells were cultured in RPMI-1640 (Thermo Fisher
Scientific, Inc.) medium containing 10% heat-inactivated FBS
(Thermo Fisher Scientific, Inc.), 1% l-glutamine, 100 U/ml
penicillin and 100 µg/ml streptomycin at 37°C and 5%
CO2.
BMMSCs were cultured according to a previously
published paper (25). In total, 15
newly diagnosed patients with AML were selected, including six
males and nine females (median age 56 years; age range 26-78
years). The inclusion criteria for patients with AML were as
follows: i) Age ≥18 years old; ii) non-acute promyelocytic
leukemia; ii) the diagnostic criteria were based on the 2016 World
Health Organization diagnostic criteria for AML (26); and iv) no other malignant
tumors-present, except AML. The control group consisted of healthy
adults who were matched for age and sex with patients with AML.
After informed consent was obtained, in keeping with ethical
guidelines of The Second Affiliated Hospital of Nanchang University
and the Declaration of Helsinki, BMMNCs were obtained from
posterior superior iliac crest bone marrow of 15 healthy donors and
15 newly diagnosed patients with AML from The Second Affiliated
Hospital of Nanchang University between January 2018 and January
2020 via lymphocyte separation medium (Tianjin Hao Yang Biological
Manufacture Co., Ltd.), and were cultured. Half of the medium was
replaced every 3-7 days. After culture for 2-4 weeks, cells became
adherent and fibroblast-like, reaching >90% confluency. Then,
cells were digested with 0.25% trypsin-EDTA and subcultured in
RPMI-1640 (Thermo Fisher Scientific, Inc.) medium containing 10%
heat-inactivated FBS (Thermo Fisher Scientific, Inc), 1%
l-glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin at 37°C
and 5% CO2. MSCs of passage 2-6 were used in
experiments. Clinical and demographic parameters of 15 newly
diagnosed patients with AML and healthy donors are presented in
Table I.
 | Table I.Clinical and demographic parameters
of participants in the study. |
Table I.
Clinical and demographic parameters
of participants in the study.
| Parameter | Patients with acute
myeloid leukemia | Healthy donors | P-value |
|---|
| Number
(untreated) | 15 (8) | 15 (15) | >0.05 |
| Age, years | 56 (26–78) | 47 (25–65) | >0.05 |
| Sex |
|
| >0.05 |
|
Male | 6 | 7 |
|
|
Female | 9 | 8 |
|
| FAB system
classification |
|
|
|
| M2 | 9 | 0 |
|
| M4 | 2 | 0 |
|
| M5 | 3 | 0 |
|
| M6 | 1 | 0 |
|
| WBC,
109/l | 21.28
(1.03-39.04) | 5.67
(3.34-9.67) | <0.05 |
| HGB, g/l | 83 (36–150) | 128 (115–158) | <0.05 |
| PLT,
109/l | 32 (5–124) | 198 (119–279) | <0.05 |
| Myeloblasts in
marrow, % | 50±22.55 | – | – |
Identification of BMMSCs
BMMSCs were detached from the culture flasks using
Accutase™ solution (EMD Millipore). Cells were washed twice with
PBS, non-specific antigens were blocked with 5% goat serum (Beijing
Biosynthesis Biotechnology Co., Ltd.) for 1 h at room temperature,
and then incubated with anti-CD14-FITC (cat. no. 561712; BD
Pharmingen; BD Biosciences), anti-CD34-FITC (cat. no. 560942; BD
Pharmingen; BD Biosciences), anti-CD90-APC (cat. no. 561971; BD
Pharmingen; BD Biosciences), anti-CD105-PE (cat. no. 560839; BD
Pharmingen; BD Biosciences) and anti-CD45-PE-Cy7 (cat. no. 560915;
BD Pharmingen; BD Biosciences) antibodies for 15 min at 4°C
according to the manufacturer's instructions. Mouse IgG1K (cat. no.
562438; BD Pharmingen; BD Biosciences; 1:500) was incubated with
cells at 4°C as an isotype control. Cells were analyzed via flow
cytometry (Gallios; Beckman Coulter, Inc.) and the FlowJo software
program (FlowJo7.6; FlowJo LLC).
Detection of FAPα expression and
myeloblasts in marrow via flow cytometry
BMMSCs were detached from culture flasks using
Accutase™ solution, washed twice with PBS and non-specific antigens
were blocked with 5% goat serum (Beijing Biosynthesis Biotechnology
Co., Ltd.) for 1 h at room temperature. Then, cells were incubated
with mouse anti-FAPα (cat. no. sc-100528; Santa Cruz Biotechnology,
Inc.; 1:500) for 2 h at 4°C. Subsequently, cells were washed twice
with PBS and incubated with secondary antibody Alexa Fluor
488-labeled goat anti-mouse IgG (cat. no. A11001; Invitrogen;
Thermo Fisher Scientific, Inc.; 1:500) for 30 min at 4°C. Cells
were analyzed via flow cytometry (Gallios; Beckman Coulter, Inc.)
and the FlowJo7.6 software program (FlowJo LLC). The steps to
detect the myeloblasts in marrow were as follows: Leukemia-related
phenotype was assess using antibodies (CD7-FITC, cat. no. 561933;
CD117-PE, cat. no. 562407; CD19-APC, cat. no. 560727; HLA-DR-PE,
cat. no. 560651; CD15-FITC, cat. no. 560997; CD34-APC,cat. no.
560940; CD56-FITC, cat. no. 562794; CD13-PE, cat. no. 560998;
CD11b-APC,cat. no. 561690; CD64-PE, cat. no. 561926; CD14-FITC,
cat. no. 561712; CD33-APC, cat. no. 561817; cCD22-PE, cat. no.
563941; cCD3-APC, cat. no. 561800; BD Pharmingen; BD Biosciences;
1:500. MPO-FITC, cat. no. 130-107-177; Miltenyi Biotec; 1:500.),
followed by the addition of 50-100 µl bone marrow. This was mixed
well and incubated in dark at room temperature for 15-20 min. Then,
1 ml hemolysin was added and incubated in dark for 12 min.
Centrifugation with 200 × g at room temperature for 5 min was
conducted, the supernatant was removed and 300 µl PBS was added for
suspension. Finally, flow cytometry (Gallios; Beckman Coulter,
Inc.) was used for detection.
Immunofluorescence
Before use in experiments, BMMSCs were seeded (60%)
in 6-well plates and cultured for 24 h. Then, cells were washed
three times with PBS for 5 min each and fixed in 4%
paraformaldehyde for 30 min at room temperature. Cells were washed
three times with PBS and permeabilized within 0.3% Triton X-100 for
10 min. After being washed three times with PBS, non-specific
antigens were blocked with 5% goat serum (Beijing Biosynthesis
Biotechnology Co., Ltd.) for 1 h at room temperature. After being
washed three times with PBS, cells were and incubated with
anti-FAPα (cat. no. sc-100528; Santa Cruz Biotechnology, Inc.;
1:500) at 4°C overnight. Then, cells were incubated in the dark
with Alexa Fluor 488-labeled goat anti-mouse IgG (cat. no. A11001;
Invitrogen; Thermo Fisher Scientific, Inc.; 1:500) at 4°C for 30
min. After being washed three times with PBS, cells were incubated
with DAPI (1:500) for 5 min at room temperature in the dark. Then,
cells were visualized using a fluorescence microscope with ×10 and
×40 magnification and images were collected.
Immunohistochemistry
A small piece of cylindrical bone marrow tissue,
0.5-1 cm long, was collected from the puncture needle. After
sampling, tissues were immediately placed into 10% neutral buffered
formalin solution and fixed at room temperature for 6-24 h. Then,
4% bone marrow decalcification solution (Vignes; http://www.wexisgp.com/) was used for 2 h at 37°C. The
slices were prepared with a thickness of 3 µm and baked at 68°C for
1 h. The antigen was dewaxed with xylene and then repaired with
citric acid antigen repair solution (cat. no. G1202; Wuhan
Servicebio Technology Co., Ltd.). The slices were placed into 3%
hydrogen peroxide solution (cat. no. G0115; Wuhan Servicebio
Technology Co., Ltd.), incubated at room temperature and protected
from light for 25 min to block endogenous peroxidase. Then, 3% BSA
(cat. no. G5001, Wuhan Servicebio Technology Co., Ltd.) was added
to evenly cover the tissue and slices were sealed at room
temperature for 30 min. Slices were incubated overnight with
anti-FAPα antibody (cat. no. ab53066; Abcam; 1:100) at 4°C. Then,
slices were incubated with a secondary goat anti-rabbit antibody
(cat. no. GB23303; Wuhan Servicebio Technology Co., Ltd.; 1:200) at
room temperature for 50 min. After DAB (cat. no. G1211; Wuhan
Servicebio Technology Co., Ltd.) color development, re-dyeing
nucleus with hematoxylin for 3 min at room temperature, dehydration
and sealing with neutral gum, images were captured using Nikon E100
light microscope with ×200 and ×400 magnification and analyzed
using Image-pro plus 6.0 (Media Cybernetios, Inc; Version 6.0.0.260
for windows 2000/XP professional; serial no. 41M60032-00032)
[optical density (OD)/area].
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from BMMSCs using RNAiso™
Plus (Takara Bio, Inc.) according to the manufacturer's
instructions. cDNA was generated from 1 µg total RNA using Prime
Script™ RT reagent kit (Perfect Real Time; Takara Bio, Inc.) with
DNA Eraser according to the manufacturer's instructions. Duration
of RT was 15 min at 37°C. RT-qPCR was performed using
SYBR® Premix Ex Taq™ II (Tli RNaseH Plus; Takara Bio,
Inc.) following the manufacturers' instructions on an iQ5
Multicolor Real-Time PCR Detection System (Bio-Rad Laboratories,
Inc.). GAPDH served as an endogenous control. The primer sequences
for FAPα and GAPDH amplification were as follows: FAPα forward,
5′-GTATTTGGAGTTGCCACCTCTG-3′ and reverse,
5′-GAAGGGCGTAAGACAATGCAC-3′; and GAPDH forward,
5′-AAGGTGAAGGTCGGAGTCAAC-3′ and reverse,
5′-GGGGTCATTGATGGCAACAATA-3′. The thermocycling conditions for FAPα
and GAPDH amplification were as follows: Initial denaturation at
95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec and 60°C
for 30 sec. The primer specificity was verified using the melt
curve. The relative quantification of FAPα in BMMSCs was calculated
using the 2−ΔΔCq method (27).
Transfection and cell apoptosis
analysis via flow cytometry
The day before transfection, BMMSCs were seeded
(60%) in 12-orifice plates. When the cells reached ~90% confluency,
they were transfected with FAPα small interfering (si)RNA (forward,
5′-CGCCCUUCAAGAGUUCAUATT-3′ and reverse,
5′-UAUGAACUCUUGAAGGGCGTT-3′), FAPα-Mock (only lipo2000 and no siRNA
sequence) and NC siRNA (forward, 5′-UUCUCCGAACGUGUCACGUTT-3′ and
reverse, 5′-ACGUGACACGUUCGGAGAATT-3′) at room temperature using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The mass and concentration of transfected siRNA
was 5 µl and 100 pmol, respectively. After 6 h, the medium was
removed, and Kasumi-1 cells were added and co-cultured with
transfected BMMSCs. The co-culture ratio of AML cells and BMMSCs
was 1-2:1.
A total of 100 µM Cytosine arabinoside (Ara-C;
Pfizer, Inc.) was added to the well to induce apoptosis in Kasumi-1
cells (28) for 24 h at room
temperature. After 24 h, co-cultured Kasumi-1 cells were isolated
from the monolayer of BMMSCs, and gently transferred into the
monolayer with medium. Next, cells were stained with
FITC-conjugated Annexin V and PE-conjugated PI at 4°C for 30 min
following the Annexin V FITC Apop Dtec Kit I (BD Biosciences)
manufacturer's instructions. Annexin V-FITC and PI-PE positive
cells were considered apoptotic cells. The interfering effect was
determined via RT-qPCR and flow cytometry (Gallios; Beckman
Coulter, Inc.) and the FlowJo7.6 software program (FlowJoLLC).
Western blot analysis
After transfection of BMMSCs with FAPα-siRNA, BMMSCs
(2×105) and Kasumi-1 cells (5×105) were
cocultured in 6-well plates. After 48 h, Kasumi-1 cells were
collected and washed twice with PBS. Total cell lysates were
obtained using lysis buffer (Beyotime Institute of Biotechnology)
supplemented with 1 mM PMSF. The protein concentration was
determined using the BCA (Sangon Biotech Co., Ltd.) method
according to the manufacturer's guidelines. An equal amount of
protein (20-40 g) was separated via 10-12% SDS-PAGE and transferred
to PVDF membranes. Next, membranes were blocked with 5% non-fat
milk for 2 h at room temperature. PVDF membranes were incubated
with anti-actin and anti-β-catenin antibodies (cat. no. 2698S; Cell
Signaling Technology, Inc.; 1:500) overnight at 4°C. Subsequently,
membranes were washed and incubated with a horseradish
peroxidase-conjugated antibody (cat. no. 7074S; Cell Signaling
Technology, Inc.; 1:5,000) in 0.2% TBS-Tween at room temperature
for 2 h. After being washed, protein bands were visualized using an
ECL detection kit (Biological Industries). Semi-quantification of
the western blotting was performed using the Bio-Rad imaging system
and ImageJ software (29)
(Life-Line Fiji versions java8; http://imagej.net/Fiji/Downloads) to detect protein
expression.
Statistical analysis
All the experiments were repeated three times. All
analyses were performed using SPSS 22.0 software (IBM Corp.). The
χ2 test was used for categorical data. Measurement data
were presented as mean ± SD. The unpaired Students t-test was used
for two independent groups. Multiple comparisons were performed
using one-way ANOVA (Bonferroni) statistical analysis. Pearson
correlation test was used for correlation analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of BMMSCs
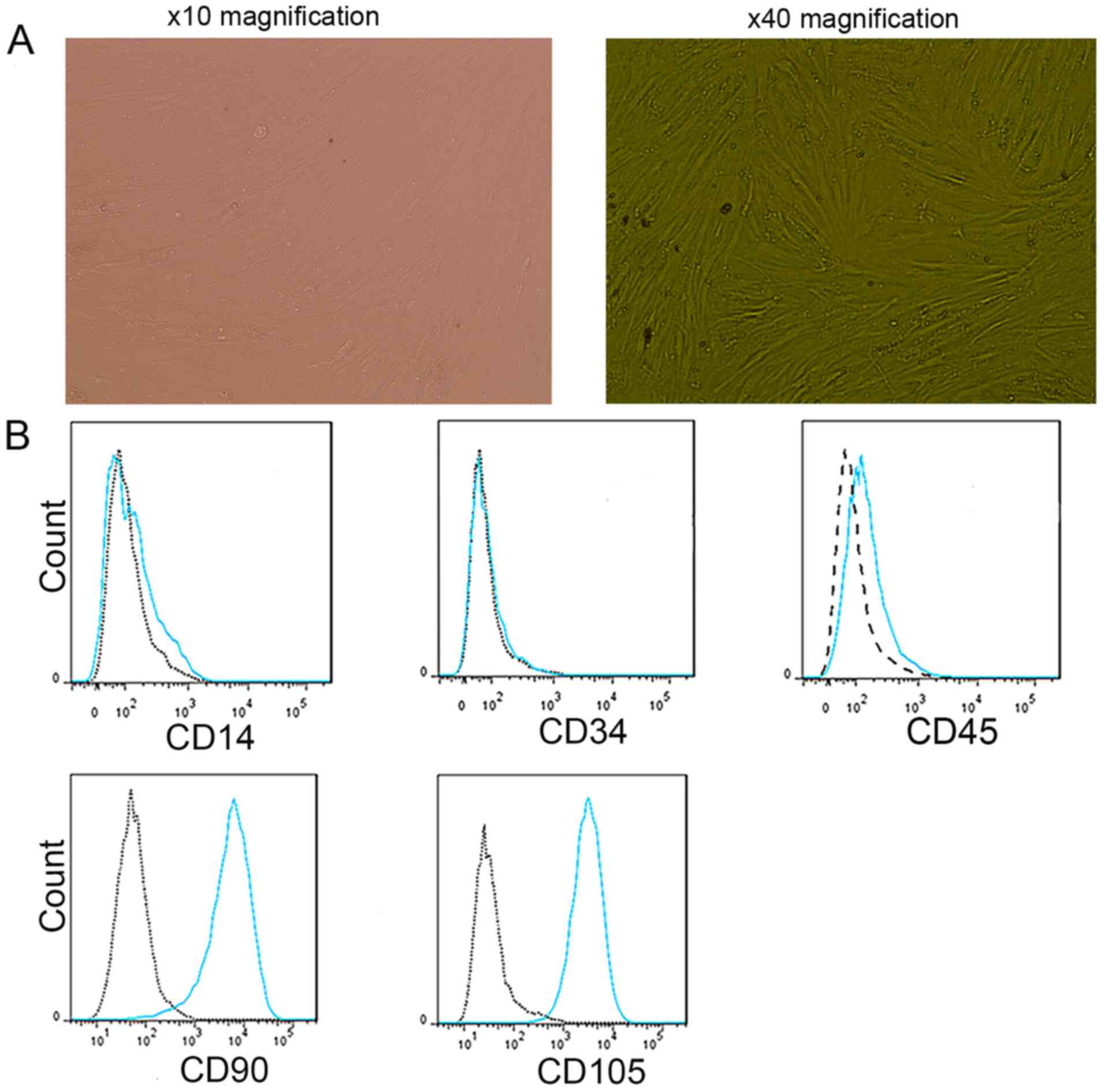
The experimental results demonstrated that cultured
BMMSCs adhered to the flasks and had a fibroblast-like morphology
(Fig. 1A). BMMSCs of passage 2 had
a low expression of CD14, CD34 and CD45, and a high expression of
CD90 and CD105 (n=3). Analysis of these CD molecular markers on
BMMSCs via flow cytometry is presented in Fig. 1B.
Expression of FAPα in BMMSCs of newly
diagnosed patients with AML and healthy donors
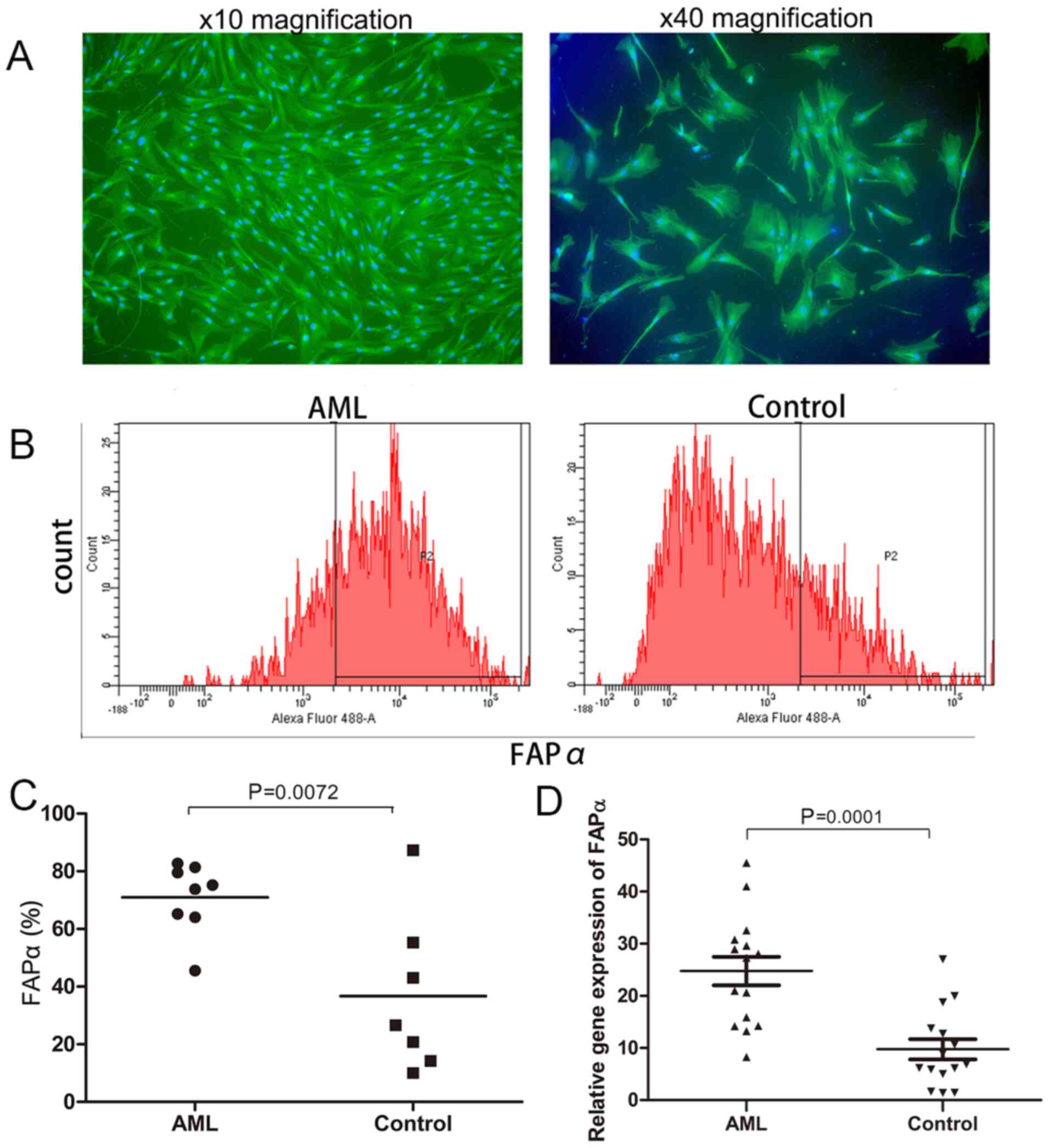
Significant differences in the expression of FAPα
between newly diagnosed patients with AML and healthy donors were
identified via flow cytometry (70.92±4.38 vs. 36.74±10.37%,
respectively; P=0.0072; Fig. 2B and
C), RT-qPCR (24.75±2.75 vs. 9.77±1.94, respectively; P=0.0001;
Fig. 2D) and immunofluorescence
(Fig. 2A).
Expression of FAPα in BM biopsy
samples of newly diagnosed patients with AML and healthy
donors
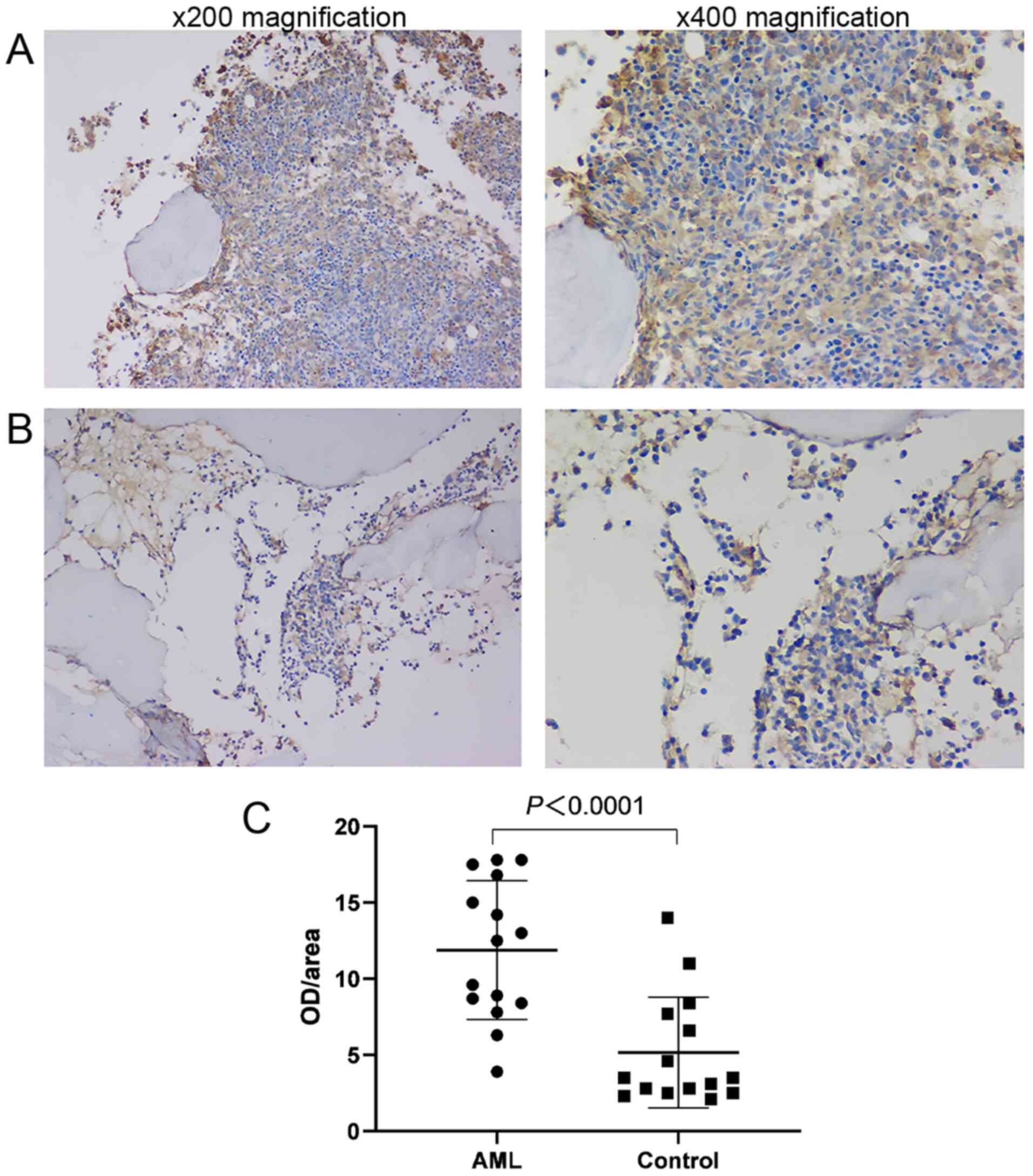
The expression of FAPα in BM biopsy samples between
newly diagnosed patients with AML and healthy donors was examined
via immunohistochemistry. FAPα expression was observed in the BM
mesenchymal matrix as indicated by the positively stained yellow
area (Fig. 3A and B). Image-pro
plus 6.0 software was used to estimate the percentage of the
stained area, represented as OD/area. The OD/area value was higher
in AML BM compared with BM from healthy donors (OD/area, 11.88±4.55
vs. 5.16±3.67; P<0.0001; n=15; Fig.
3C).
Correlation of FAPα expression in BM
with clinical parameters and survival time of newly diagnosed
patients with AML
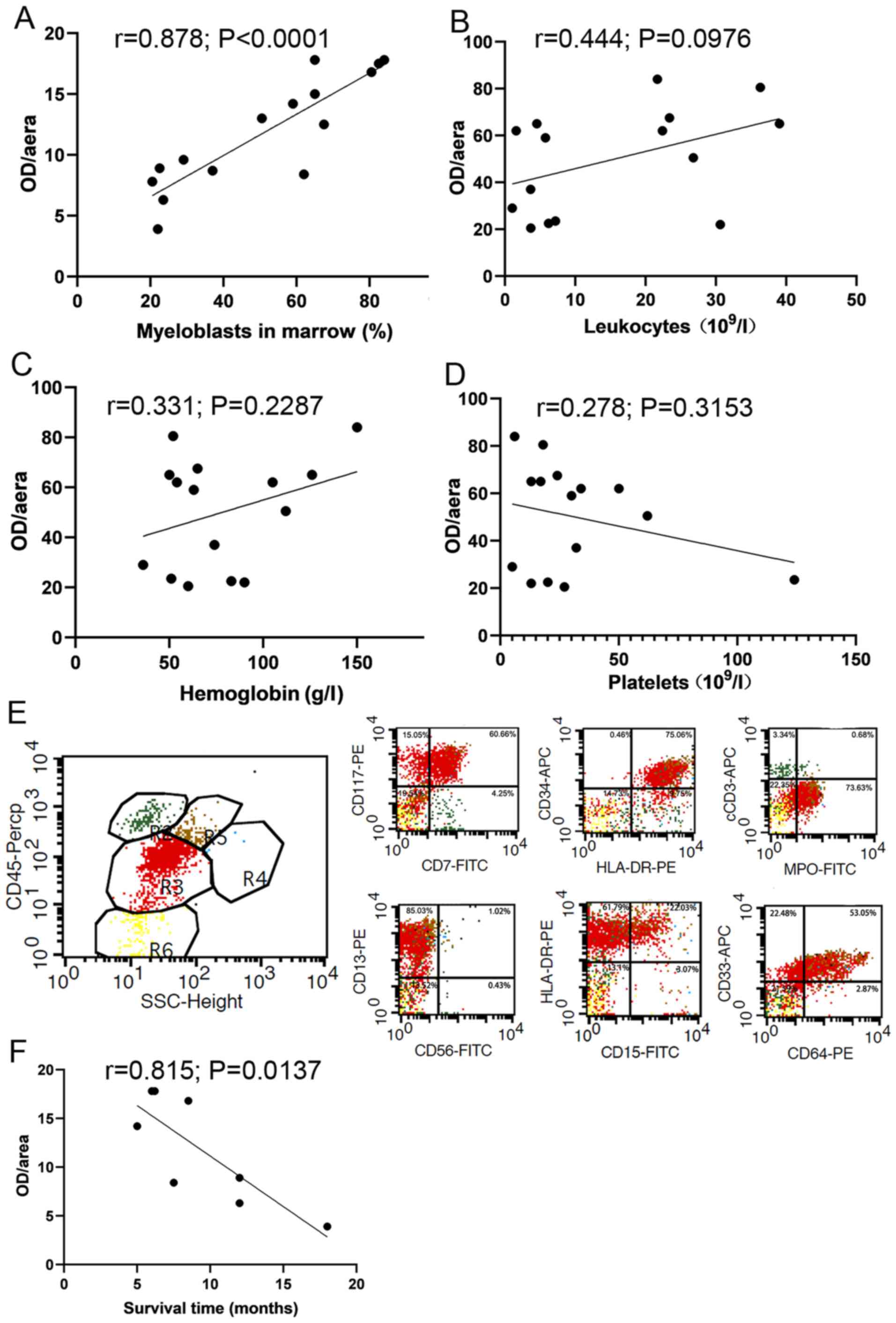
The correlation between FAPα expression in BM and
clinically parameters of newly diagnosed AML patients was analyzed,
and the results demonstrated that the OD/area value of FAPα was
positively correlated with the proportion of myeloblasts in marrow
detected via flow cytometry (r=0.878; P<0.0001; Fig. 4A). No significant correlation was
observed between the OD/area value of FAPα and leukocytes (r=0.444;
P=0.0976; Fig. 4B), hemoglobin
(r=0.331; P=0.2287; Fig. 4C) or
platelets (r=0.278; P=0.3153; Fig.
4D). Representative flow scatter plots of myeloblasts in marrow
is presented in Fig. 4E. The
OD/area value of FAPα was significantly and negatively correlated
with survival in eight patients who did not undergo chemotherapy
(r=0.815; P=0.0137; Fig. 4F).
BMMSCs protects AML cells from
Ara-C-induced apoptosis, which in part is contributed by FAPα
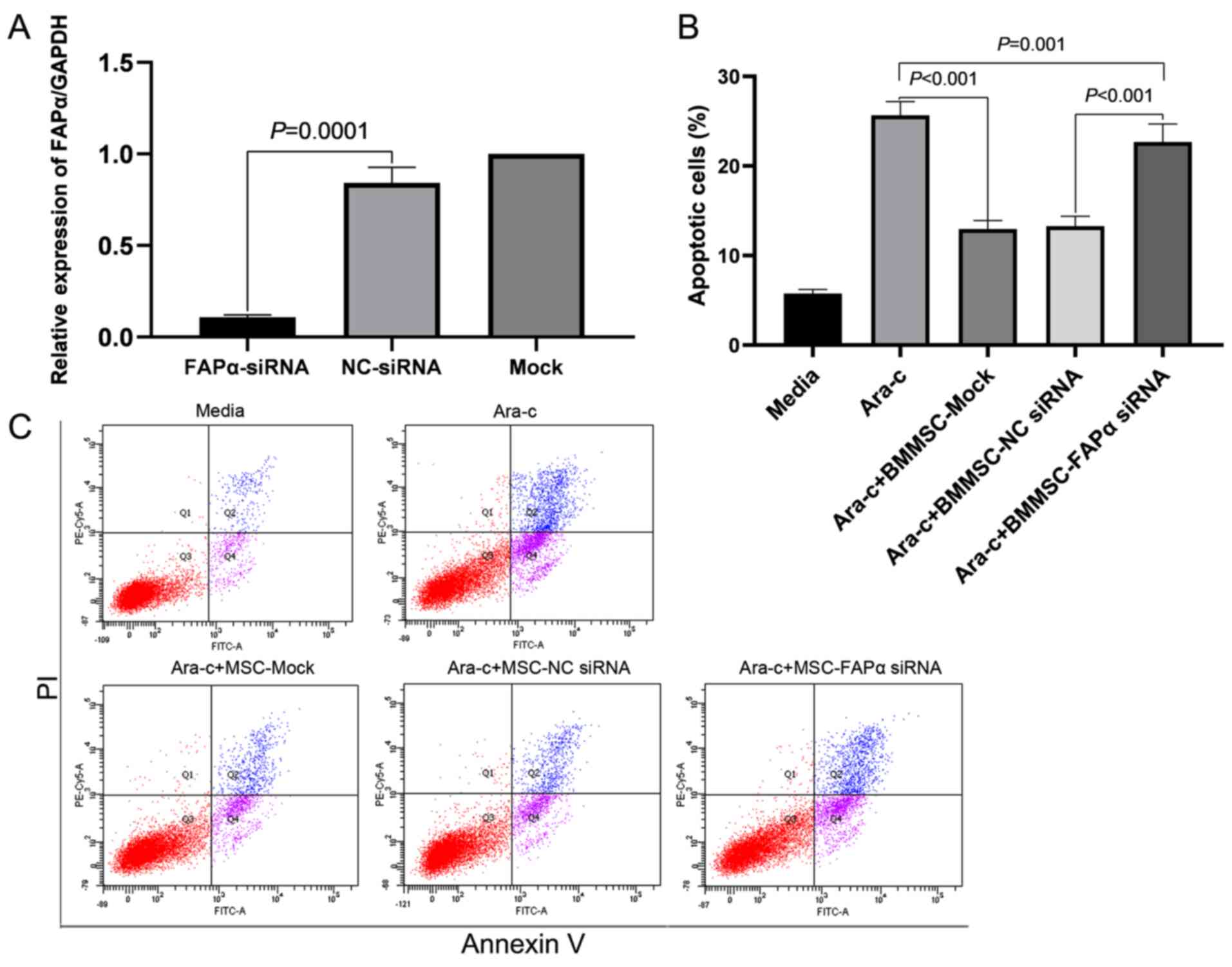
RT-qPCR was used to verify successful transfection
prior to detecting apoptosis. The relative expression of FAPα mRNA
in the FAPα-siRNA group was significantly lower compared with that
in negative control (NC)-siRNA group (0.11±0.01 vs. 0.84±0.08;
P=0.0001; Fig. 5A). Then, flow
cytometry results demonstrated that BMMSCs protected Kasumi-1 cells
from apoptosis, even in the presence of Ara-C (100 µM), and
knockdown of FAPα with siRNA decreased the protective effect of
BMMSCs (Fig. 5C). The proportion of
apoptotic cells in the MSC-Mock group was significantly lower
compared with that in the Ara-C group (12.96±0.95 vs. 25.66±1.54%;
P<0.001; n=9; Fig. 5B).
Knockdown of FAPα using siRNA decreased the protective effect. The
proportion of apoptotic cells in the FAPα-siRNA group was
significantly increased compared with the proportion in the
NC-siRNA group (22.69±1.99 vs. 13.29±1.10%; P<0.001; n=9;
Fig. 5B). The proportion of
apoptotic cells in FAPα-siRNA group was also lower compared with
that in the Ara-C group (22.69±1.99 vs. 25.66±1.54%; P=0.001;
Fig. 5B).
Possible mechanism of FAPα protecting
AML cells from apoptosis
The results demonstrated that Kasumi-1 cells
expressed β-catenin, which could be inhibited by Ara-C. The
expression of β-catenin in Kasumi-1 cells was significantly
increased when Kasumi-1 cells were cocultured with BMMSCs and
NC-siRNA for 48 h, and even in the presence of Ara-C the expression
of β-catenin is activated. Compared with NC-siRNA, knockdown of
FAPα using siRNA significantly suppressed the expression of
β-catenin (Fig. 6A). The experiment
was performed in triplicate, the relative expression of β-catenin
was calculated and statistical analyses were performed. In the
absence of BMMSCs, the expression of β-catenin was decreased from
113.20±2.77 to 44.02±0.06 in groups with and without Ara-C,
respectively (P<0.0001; Fig.
6B). In addition, in the presence of Ara-C and BMMSCs, the
expression of β-catenin in the NC-siRNA group and FAPα-siRNA group
was 116.84±0.40 and 82.11±1.26, respectively (P<0.0001; Fig. 6B).
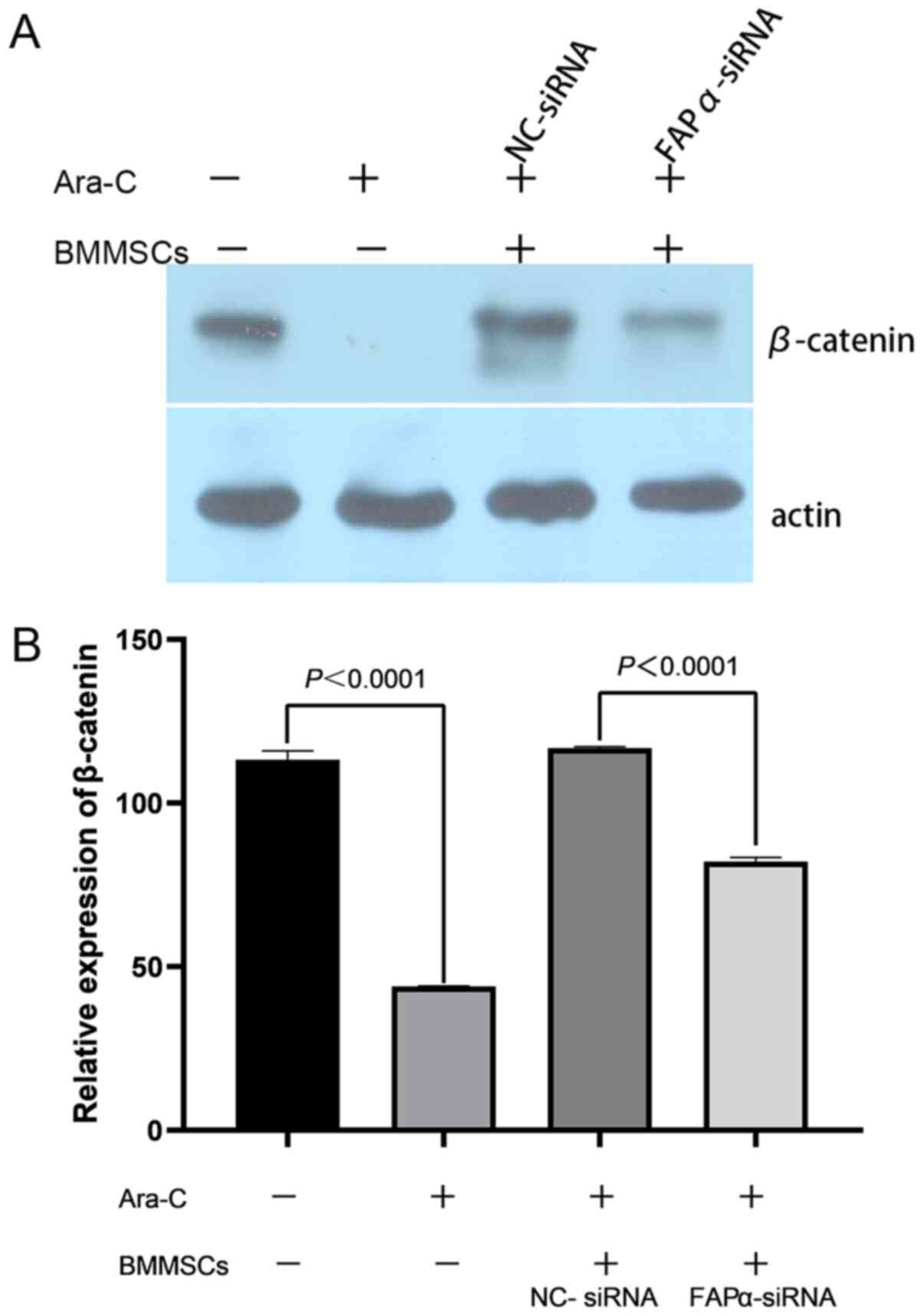 | Figure 6.Possible role of FAPα in
BMMSCs-mediated protection of AML cell line. (A) Kasumi-1 cells
expressed β-catenin, which can be inhibited by Ara-C. The
expression of β-catenin of Kasumi-1 cells was significantly
activated when Kasumi-1 cells cocultured with BMMSCs. Knockdown of
FAPα using siRNA significantly suppressed the expression of
β-catenin. (B) In the absence of BMMSCs, the expression of
β-catenin in groups with Ara-C was 113.20±2.77 and that without
Ara-C was 44.02±0.06 (P<0.0001). In addition, in the presence of
Ara-C and BMMSCs, the expression of β-catenin in the NC-siRNA group
and FAPα-siRNA group was 116.84±0.40 and 82.11±1.26, respectively
(P<0.0001). NC, negative control; siRNA, small interfering RNA;
BMMSCs, bone marrow mesenchymal stem cells; FAPα, fibroblast
activation protein α; Ara-C, Cytosine arabinoside. |
Discussion
In 1991, Caplan's pioneering work reported the
existence of BMMSCs (30).
Typically, MSCs are isolated by their ability to adhere to the
plastic surface of a culture dish, which demonstrates the ability
of the cells to expand in culture plates, and MSCs have specific
markers of immunological characteristics (31). The International Society for
Cellular Therapy suggests three minimal criteria for the
characterization of MSCs: i) Adherence to plastic; ii) expression
of markers associated with MCS (such as CD73, CD90 and CD105), and
the lack of hematopoietic-related cell expression (including CD34,
CD45, CD11b or CD14, CD19 or CD79 and human leukocyte antigen DR);
and iii) potential tri-lineage differentiation into adipocytes,
osteoblasts and chondrocytes (30).
In the present study, cultured BMMSCs adhered to culture flasks and
were fibroblast-like in morphology. In addition, BMMSCs of passage
2 had a low expression of CD14, CD34 and CD45, and high expression
of CD90 and CD105. Collectively, the current findings met the
minimal criteria for defining MSCs.
FAPα (also known as seprase) is a serine
oligopeptidase that was originally identified in 1986 as an
inducible cell surface glycoprotein F19 (32), and was renamed FAP in 1994 based
upon its abundance in activated fibroblasts (33). Human FAP is comprised of M(r) 95,000
(p95, FAPα) and M(r) 105,000 (p105, FAPβ) subunits, which are
conjugated by non-covalent, non-disulfide bonds (34). Immunoblot experiments have shown
that FAPα, but not FAPβ, carries an epitope that is bound by
monoclonal antibody F19 (35), and
so the F19 surface antigen was renamed to FAPα.
Most healthy adult tissues have little or no
detectable FAPα expression (36).
However, FAPα expression is highly upregulated during active tissue
remodeling of fibroblasts, including wound healing, fibrosis and at
cancer sites (37). FAPα is also
expressed in BMMSCs, and promotes the motility of human BMMSCs
(38). In the context of cancer,
FAPα has been widely considered as a marker of CAFs, which have
multiple pro-tumorigenic functions (38). Moreover, FAPα itself has been
reported to exert pro-tumorigenic activity, both via enzymatic and
non-enzymatic mechanisms (36,39).
Previous studies have revealed that tumor cell conditioned medium
can transform BMMSCs into CAFs, thereby promoting tumor progression
(40). Moreover, tumor cells can
reprogram BMMSCs and promote the evolution of BMMSCs into CAFs,
further facilitating the development of hematological malignancies
(41). Therefore, it was
hypothesized that the AML microenvironment may instruct BMMSCs to
become CAFs, to further increase the expression of FAPα in BMMSCs
compared with BMMSCs in a healthy environment.
The present study first evaluated the expression of
FAPα on BMMSCs between newly diagnosed patients with AML and
healthy donors using RT-qPCR and flow cytometry. The experimental
results demonstrated that the expression of FAPα in newly diagnosed
patients with AML was significantly higher compared with healthy
donors. However, the specific origin of FAPα within the BM remains
to be elucidated. To further confirm the expression of FAPα in BM,
the expression of FAPα in the BM mesenchymal matrix was positively
stained yellow. It was identified that the OD/area value of FAPα in
newly diagnosed patients with AML was significantly higher compared
with that of healthy donors. Previous studies have reported that
FAPα expression is associated with worse prognosis in solid tumors
(42). However, the relationship
between FAPα expression and the prognosis in hematologic tumors
remains unknown. The present study demonstrated that the OD/area
value of FAPα was positively correlated with the proportion of
myeloblasts in BM as detected via flow cytometry, and was
significantly negatively associated with survival time in eight
patients who did not undergo chemotherapy due to the costs and
risks. Collectively, these results suggested that FAPα serves an
important role in the AML BM microenvironment, and may be involved
in the development of AML. Thus, increased expression of FAPα in BM
may be a poor prognostic factor in AML.
The interaction between AML cells and BMMSCs, either
via direct contact or secreted cytokines, can promote AML
progression, survival, growth, chemotherapy resistance and
angiogenesis (43–46). However, little is known regarding
the function of FAPα expression in BMMSCs and how it affects the
features of AML cells. In the current study, flow cytometry results
demonstrated that BMMSCs protected Ara-C (100 µM)-induced Kasumi-1
cell apoptosis in a cocultured system. Furthermore, knockdown of
FAPα with siRNA decreased this protective effect. However, the
proportion of apoptotic cells in the FAPα-siRNA group was lower
compared with that in Ara-Cgroup. Thus, these results suggested
that BMMSCs in the BM microenvironment of patients with AML were
instructed to become CAFs to protect AML cells from apoptosis,
which, in part was contributed by FAPα. However, other factors may
be involved in the protective effect of BMMSCs on leukemia cell
apoptosis.
Previous studies have revealed that, via a variety
of mechanisms, BMMSCs inhibit AML cell apoptosis and promote their
survival, proliferation and chemotherapy resistance (46–51).
In addition, it has been reported that Wnt ligands are the key
drivers of most types of tissue stem cells in adult mammals, and
mutated Wnt pathway components are causative in the progression of
multiple growth-related pathologies and cancer (52). For instance, the Wnt/β-catenin
pathway is required for the development of leukemia stem cells in
AML (53). However, the current
understanding of the activation of the BMMSC-mediated Wnt/β-catenin
signaling pathway is limited. Furthermore, the relationship between
FAPα and Wnt/β-catenin for AML cellular features remains unknown.
The present study demonstrated that Kasumi-1 cells expressed
β-catenin, which could be inhibited by Ara-C. However, the
expression of β-catenin in Kasumi-1 cells was significantly
increased when Kasumi-1 cells were cocultured with BMMSCs, even in
the presence of Ara-C. The data also indicated that knockdown of
FAPα using siRNA significantly suppressed the expression of
β-catenin. Thus, it was suggested FAPα may be involved in
BMMSC-mediated apoptosis of AML cells induced by Ara-C, which may
be achieved via the β-catenin pathway.
In conclusion, the present results indicated that
FAPα may serve an important role in the AML BM microenvironment and
that it may be involved in the development of AML. Moreover, it was
suggested that increased expression of FAPα in BM may be a poor
prognostic factor in AML. Further studies demonstrated that BMMSCs
could protect AML cells from apoptosis, which in part was
contributed by FAPα, and likely occurs via the β-catenin signaling
pathway. However, whether the activation of β-catenin was dependent
on direct contact with BMMSCs or secretory of cytokines requires
further research. In addition, the quantity of the samples should
be expanded to clarify the relationship between FAPα expression and
the prognosis of AML, and additional studies should be performed
using other AML cell lines to further verify the anti-apoptotic
role of FAPα in AML cells.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81560030), the Natural
Science Foundation of Jiangxi Province (grant no. 20192BAB205034)
and the Science and Technology Plan of Health and Family Planning
Commission Jiangxi Province (grant no. 20181097).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SM, FZ and GC designed the research and analyzed the
data. SM, YZ and LY performed the cell function experiments. SM and
YZ collected the BM liquid and biopsy samples, and reviewed the
clinical features. SM prepared the images and drafted the original
manuscript. FZ and GC reviewed and edited the manuscript. All
authors read and approved the manuscript, and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The study was approved by the Medical Research
Ethics Committee of the Second Affiliated Hospital of Nanchang
University. All patients and healthy donors provided written
informed consent. The study in keeping with ethical guidelines of
the Second Affiliated Hospital of Nanchang University and the
Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dores GM, Devesa SS, Curtis RE, Linet MS
and Morton LM: Acute leukemia incidence and patient survival among
children and adults in the United States, 2001-2007. Blood.
119:34–43. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Montesinos P, Bergua J, Infante J, Esteve
J, Guimaraes JE, Sierra J and Sanz MÁ: Update on management and
progress of novel therapeutics for R/R AML: An Iberian expert panel
consensus. Ann Hematol. 98:2467–2483. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liao Z, Tan ZW, Zhu P and Tan NS:
Cancer-associated fibroblasts in tumor microenvironment -
Accomplices in tumor malignancy. Cell Immunol. 343:1037292019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shafat MS, Gnaneswaran B, Bowles KM and
Rushworth SA: The bone marrow microenvironment - Home of the
leukemic blasts. Blood Rev. 31:277–286. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Geyh S, Rodríguez-Paredes M, Jäger P,
Khandanpour C, Cadeddu RP, Gutekunst J, Wilk CM, Fenk R, Zilkens C,
Hermsen D, et al: Functional inhibition of mesenchymal stromal
cells in acute myeloid leukemia. Leukemia. 30:683–691. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
von der Heide EK, Neumann M, Vosberg S,
James AR, Schroeder MP, Ortiz-Tanchez J, Isaakidis K, Schlee C,
Luther M, Jöhrens K, et al: Molecular alterations in bone marrow
mesenchymal stromal cells derived from acute myeloid leukemia
patients. Leukemia. 31:1069–1078. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Augsten M: Cancer-associated fibroblasts
as another polarized cell type of the tumor microenvironment. Front
Oncol. 4:622014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chaturvedi P, Gilkes DM, Wong CC, Luo W,
Zhang H, Wei H, Takano N, Schito L, Levchenko A, Semenza GL and
Kshitiz: Hypoxia-inducible factor-dependent breast
cancer-mesenchymal stem cell bidirectional signaling promotes
metastasis. J Clin Invest. 123:189–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Raz Y, Cohen N, Shani O, Bell RE,
Novitskiy SV, Abramovitz L, Levy C, Milyavsky M, Leider-Trejo L,
Moses HL, et al: Bone marrow-derived fibroblasts are a functionally
distinct stromal cell population in breast cancer. J Exp Med.
215:3075–3093. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tao L, Huang G, Song H, Chen Y and Chen L:
Cancer associated fibroblasts: An essential role in the tumor
microenvironment. Oncol Lett. 14:2611–2620. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou P, Xiao N and Wang J, Wang Z, Zheng
S, Shan S and Wang J, Du J and Wang J: SMC1A recruits
tumor-associated-fibroblasts (TAFs) and promotes colorectal cancer
metastasis. Cancer Lett. 385:39–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Komohara Y and Takeya M: CAFs and TAMs:
Maestros of the tumour microenvironment. J Pathol. 241:313–315.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen X and Song E: Turning foes to
friends: Targeting cancer-associated fibroblasts. Nat Rev Drug
Discov. 18:99–115. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
LeBleu VS and Kalluri R: A peek into
cancer-associated fibroblasts: origins, functions and translational
impact. Dis Model Mech. 11:dmm0294472018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Higashino N, Koma YI, Hosono M, Takase N,
Okamoto M, Kodaira H, Nishio M, Shigeoka M, Kakeji Y and Yokozaki
H: Fibroblast activation protein-positive fibroblasts promote tumor
progression through secretion of CCL2 and interleukin-6 in
esophageal squamous cell carcinoma. Lab Invest. 99:777–792. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wen X, He X, Jiao F, Wang C, Sun Y, Ren X
and Li Q: Fibroblast activation protein-α-positive fibroblasts
promote gastric cancer progression and resistance to immune
checkpoint blockade. Oncol Res. 25:629–640. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu M, Qian C, Hu Z, Fei B and Zhou H:
Biomarkers in tumor Microenvironment? Upregulation of fibroblast
activation protein-α correlates with gastric cancer progression and
poor prognosis. OMICS. 21:38–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Byrling J, Sasor A, Nilsson J, Said
Hilmersson K, Andersson R and Andersson B: Expression of fibroblast
activation protein and the clinicopathological relevance in distal
cholangiocarcinoma. Scand J Gastroenterol. 55:82–89. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang L, Yang L, Xia ZW, Yang SC, Li WH,
Liu B, Yu ZQ, Gong PF, Yang YL, Sun WZ, et al: The role of
fibroblast activation protein in progression and development of
osteosarcoma cells. Clin Exp Med. 20:121–130. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pleshkan VV, Alekseenko IV, Tyulkina DV,
Kyzmich AI, Zinovyeva MV and Sverdlov ED: Fibroblast activation
protein (FAP) as a possible target of an antitumor strategy. Mol
Gen Mikrobiol Virusol. 34:90–97. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li M, Li M, Yin T, Shi H, Wen Y, Zhang B,
Chen M, Xu G, Ren K and Wei Y: Targeting of cancer associated
fibroblasts enhances the efficacy of cancer chemotherapy by
regulating the tumor microenvironment. Mol Med Rep. 13:2476–2484.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Raffaghello L, Vacca A, Pistoia V and
Ribatti D: Cancer associated fibroblasts in hematological
malignancies. Oncotarget. 6:2589–2603. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zi FM, He JS, Li Y, Wu C, Wu WJ, Yang Y,
Wang LJ, He DH, Yang L, Zhao Y, et al: Fibroblast activation
protein protects bortezomib-induced apoptosis in multiple myeloma
cells through β-catenin signaling pathway. Cancer Biol Ther.
15:1413–1422. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pittenger MF: Mesenchymal stem cells from
adult bone marrow. Mesenchymal Stem Cells. Springer; Basel: pp.
27–44. 2008, View Article : Google Scholar
|
|
26
|
Arber DA, Orazi A, Hasserjian R, Thiele J,
Borowitz MJ, Le Beau MM, Bloomfield CD, Cazzola M and Vardiman JW:
The 2016 revision to the World Health Organization classification
of myeloid neoplasms and acute leukemia. Blood. 127:2391–2405.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Y, Mallampati S, Sun B, Zhang J, Kim
SB, Lee JS, Gong Y, Cai Z and Sun X: Wnt pathway contributes to the
protection by bone marrow stromal cells of acute lymphoblastic
leukemia cells and is a potential therapeutic target. Cancer Lett.
333:9–17. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Abramoff MD, Magalhaes PJ and Ram SJ:
Image processing with ImageJ. Biophoton Int. 11:36–42. 2004.
|
|
30
|
Caplan AI: Mesenchymal stem cells. J
Orthop Res. 9:641–650. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop Dj and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The International Society for Cellular
Therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rettig WJ, Chesa PG, Beresford HR,
Feickert HJ, Jennings MT, Cohen J, Oettgen HF and Old LJ:
Differential expression of cell surface antigens and glial
fibrillary acidic protein in human astrocytoma subsets. Cancer Res.
46:6406–6412. 1986.PubMed/NCBI
|
|
33
|
Rettig WJ, Su SL, Fortunato SR, Scanlan
MJ, Raj BK, Garin-Chesa P, Healey JH and Old LJ: Fibroblast
activation protein: Purification, epitope mapping and induction by
growth factors. Int J Cancer. 58:385–392. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Scanlan MJ, Raj BK, Calvo B, Garin-Chesa
P, Sanz-Moncasi MP, Healey JH, Old LJ and Rettig WJ: Molecular
cloning of fibroblast activation protein alpha, a member of the
serine protease family selectively expressed in stromal fibroblasts
of epithelial cancers. Proc Natl Acad Sci USA. 91:5657–5661. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rettig WJ, Garin-Chesa P, Healey JH, Su
SL, Ozer HL, Schwab M, Albino AP and Old LJ: Regulation and
heteromeric structure of the fibroblast activation protein in
normal and transformed cells of mesenchymal and neuroectodermal
origin. Cancer Res. 53:3327–3335. 1993.PubMed/NCBI
|
|
36
|
Keane FM, Yao TW, Seelk S, Gall MG,
Chowdhury S, Poplawski SE, Lai JH, Li Y, Wu W, Farrell P, et al:
Quantitation of fibroblast activation protein (FAP)-specific
protease activity in mouse, baboon and human fluids and organs.
FEBS Open Bio. 4:43–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jacob M, Chang L and Puré E: Fibroblast
activation protein in remodeling tissues. Curr Mol Med.
12:1220–1243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chung KM, Hsu SC, Chu YR, Lin MY, Jiaang
WT, Chen RH and Chen X: Fibroblast activation protein (FAP) is
essential for the migration of bone marrow mesenchymal stem cells
through RhoA activation. PLoS One. 9:e887722014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hamson EJ, Keane FM, Tholen S, Schilling O
and Gorrell MD: Understanding fibroblast activation protein (FAP):
Substrates, activities, expression and targeting for cancer
therapy. Proteomics Clin Appl. 8:454–463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zi F, He J, He D, Li Y, Yang L and Cai Z:
Fibroblast activation protein α in tumor microenvironment: Recent
progression and implications (Review). Mol Med Rep. 11:3203–3211.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tian K, Yang S, Ren Q and Han Z, Lu S, Ma
F, Zhang L and Han Z: p38 MAPK contributes to the growth inhibition
of leukemic tumor cells mediated by human umbilical cord
mesenchymal stem cells. Cell Physiol Biochem. 26:799–808. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu F, Qi L, Liu B, Liu J, Zhang H, Che D,
Cao J, Shen J, Geng J, Bi Y, et al: Fibroblast activation protein
overexpression and clinical implications in solid tumors: A
meta-analysis. PLoS One. 10:e01166832015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Traer E, Martinez J, Javidi-Sharifi N,
Agarwal A, Dunlap J, English I, Kovacsovics T, Tyner JW, Wong M and
Druker BJ: FGF2 from marrow microenvironment promotes resistance to
FLT3 inhibitors in acute myeloid leukemia. Cancer Res.
76:6471–6482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fathi E, Sanaat Z and Farahzadi R:
Mesenchymal stem cells in acute myeloid leukemia: A focus on
mechanisms involved and therapeutic concepts. Blood Res.
54:165–174. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Brenner AK, Nepstad I and Bruserud Ø:
Mesenchymal stem cells support survival and proliferation of
primary human acute myeloid leukemia cells through heterogeneous
molecular mechanisms. Front Immunol. 8:1062017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li Q, Pang Y, Liu T, Tang Y, Xie J, Zhang
B and Chen H: Effects of human umbilical cord-derived mesenchymal
stem cells on hematologic malignancies. Oncol Lett. 15:6982–6990.
2018.PubMed/NCBI
|
|
47
|
Yang X, Sexauer A and Levis M: Bone marrow
stroma-mediated resistance to FLT3 inhibitors in FLT3-ITD AML is
mediated by persistent activation of extracellular regulated
kinase. Br J Haematol. 164:61–72. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tian C, Zheng G, Zhuang H, Li X, Hu D, Zhu
L, Wang T, You MJ and Zhang Y: MicroRNA-494 activation suppresses
bone marrow stromal cell-mediated drug resistance in acute myeloid
leukemia cells. J Cell Physiol. 232:1387–1395. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Guo F, Wang Y, Liu J, Mok SC, Xue F and
Zhang W: CXCL12/CXCR4: A symbiotic bridge linking cancer cells and
their stromal neighbors in oncogenic communication networks.
Oncogene. 35:816–826. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Malfuson JV, Boutin L, Clay D, Thépenier
C, Desterke C, Torossian F, Guerton B, Anginot A, de Revel T,
Lataillade JJ, et al: SP/drug efflux functionality of hematopoietic
progenitors is controlled by mesenchymal niche through VLA-4/CD44
axis. Leukemia. 28:853–864. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Battula VL, Benito J, Somanchi AG, Duvvuri
S, Hodgson L, Chen Y, Ma W, Davis RE, McNiece I, Shpall EJ, et al:
Acute myeloid leukemia cells acquire stem cell features in the bone
marrow microenvironment. Clin Lymphoma Myeloma Leuk. 14:S119–S120.
2014. View Article : Google Scholar
|
|
52
|
Nusse R and Clevers H: Wnt/β-catenin
signaling, disease, and emerging therapeutic modalities. Cell.
169:985–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wang Y, Krivtsov AV, Sinha AU, North TE,
Goessling W, Feng Z, Zon LI and Armstrong SA: The Wnt/beta-catenin
pathway is required for the development of leukemia stem cells in
AML. Science. 327:1650–1653. 2010. View Article : Google Scholar : PubMed/NCBI
|