Introduction
Endometrial cancer (EC) is the most common
gynecological cancer in developed countries and one of the most
important causes of cancer-related deaths in women worldwide.
Approximately 3% of women develop EC in their lifetime, and the
incidence is expected to increase further over the next 10 years
(1). EC is generally classified
into two types. Type I EC is the most common and is
estrogen-related, and in general this tumor is low grade and can be
diagnosed according to early bleed symptoms. Type II EC is much
more likely to be high grade (2).
The survival rate of patients with early-stage EC is relatively
high, but there is a poor prognosis for patients with advanced or
recurrent EC due to limited treatment options (3). In addition, for patients with
metastatic EC, the median survival time is only 7-12 months
(4). The standard treatment for EC
is surgery, platinum-based chemotherapy and radiotherapy where
recommended as adjuvant therapy for patients with high histologic
grade or metastasis (3). Estrogen
and progesterone receptors are expressed in Type 1 EC, which is
often responsive to endocrine treatment (5). However, endocrine therapy for advanced
stage and recurrent EC remains controversial as it has not been
revealed to widely improve long-term survival rates (6). For unresectable recurrent/metastatic
EC, chemotherapy is vital for EC patients, but currently the
options for therapeutic agents are limited, this may be in part due
to variations in the molecular and genetic characteristics of the
tumor. In addition, EC patients are presenting at a younger age
than in previous decades (7–9)
therefore more chemotherapeutic agents or individual targeting
drugs with lower toxicity need to be urgently investigated. One
option is to investigate potential therapeutic drugs from natural
compounds to improve EC treatment.
Garcinol, a polyisoprenylated benzophenone, is a
natural compound isolated from the fruit rinds of Garcinia
indica (10). In recent years,
garcinol has been reported to have antioxidative, anti-inflammatory
and anticancer effects (11–14).
The anticancer function of garcinol has been reported in several
types of cancer cells, such as colon, prostate, liver, lung,
breast, esophageal, pancreatic and oral cancer cells (12,15–18).
Zhao et al demonstrated that garcinol suppressed cervical
cancer cell proliferation but did not change the cell viability of
primary normal cervical cells (19). Studies have also revealed that
garcinol has an antitumor function in vivo and in
vitro (12,13). Therefore, garcinol may be a
promising agent for some types of cancers. However, it has not been
investigated whether garcinol has anticancer effects in EC.
The cell cycle is an important physiological process
regulating cell growth and proliferation. Cell cycle progression is
tightly controlled by cell cycle-related proteins, including cell
cycle promotors cyclin D/E/A/B and cyclin-dependent kinase (CDK)
1/2/4/6, and cell cycle inhibitors p15, p16, p21, p27 and p53
(20). These proteins exert their
functions at specific different phases of the cell cycle, although
the cell cycle has a circular control system. Cyclins are
synthesized and destroyed at particular times during the cell cycle
(21). Different members of the CDK
family are associated with different cyclins, which play an
important role in switching from one phase to the next throughout
the cell cycle (22). Numerous
anticancer drugs induce cell cycle arrest to inhibit tumor cell
proliferation (23–26). Studies have revealed that garcinol
can also induce cell cycle arrest to inhibit cell growth in lung
cancer (26), cervical (19), oral (24), and breast cancer (17). In lung cancer cells, garcinol
altered relative cell-cycle protein expression levels, upregulating
p53 and p21, and downregulating cyclin D, CDK2 and CDK4, to induce
cell cycle arrest (26). Garcinol
inhibited cervical cancer cells by delaying cell cycle progression
at the G0/G1 phase and downregulating cyclin D1 and CDK4, while
upregulating p21 and p53 (19).
However, whether garcinol can induce cell cycle arrest and inhibit
cell proliferation in EC is unknown. Therefore, the aim of the
present study was to investigate the effect(s) of garcinol on EC
cell proliferation and the cell cycle.
Materials and methods
Cell culture
The human EC cell lines, Ishikawa (ISH) and HEC-1B,
were purchased from FuHeng Cell Bank (FuHeng Biology; http://www.fudancell.com/). Both of these cell lines
were authenticated with DNA fingerprinting using short tandem
repeat (STR) methodology by the provider on 13 June 2017 and 24 May
2017, respectively. ISH and HEC-1B cells were cultured in RPMI-1640
medium (cat. no. SH30809; HyClone; Cytiva) with 10% fetal bovine
serum (FBS) (cat. no. SFBS-B; Bovogen Biologicals Pty, Ltd.), 1%
penicillin-streptomycin (cat. no. 15140122, Gibco; Thermo Fisher
Scientific, Inc.), and incubated in 5% CO2 humidified
atmosphere at 37°C. The medium was changed every 2 days.
Real-time cell proliferation
assay
The label-free real-time cellular analysis (RTCA)
system (ACE BioSciences, Inc.) was used to observe the effect of
garcinol on cell proliferation according to the manufacturer's
instructions. Culture medium (50 µl) was added to each well of a
16-well E-plate to plot the baseline prior to the addition of 5,000
cells in 100 µl medium containing various concentrations of
garcinol (0, 1, 5, 10 and 20 µM; cat. no. 2088-25; BioVision,
Inc.). The cells were placed at room temperature for 30 min to
attach to the E-plate before subsequent detection. Cells were
incubated in a 5% CO2 humidified atmosphere at 37°C for
72 h. The instrument recorded the impedance of the electron flow
caused by adherent cells using a non-unit parameter called cell
index. Data was recorded every 15 min. The assay was performed in
duplicate at least three times.
Cell counting assay
The cells (1×105) were seeded in 6-well
plates. After incubation in a 5% CO2 humidified
atmosphere at 37°C for 24 h, various concentrations of garcinol (0,
1, 5, 10 and 20 µM) were added to each well and the cells were
cultured in a 5% CO2 humidified atmosphere at 37°C.
After 48 h, the cells were digested at 37°C for 3 min by 0.25%
trypsin-EDTA solution (cat. no. 25200056; Gibco; Thermo Fisher
Scientific, Inc.) and resuspended to a single cell suspension, and
then the number of cells were counted by an automated cell counter
machine (Bio-Rad Laboratories, Inc.). The assay was performed three
times in triplicate.
Colony formation assay
A total of 500 cells/well were plated in 6-well
plates, after 24 h, and then 0, 1, 5, 10 and 20 µM garcinol was
added and the media changed every 48 h. After 14 days, the cells
were washed with phosphate-buffered saline (PBS) and fixed with
100% methanol at room temperature for 30 min, and then washed with
PBS. The plates were stained with 1% crystal violet for 1 h at room
temperature and washed with tap water. Plates were air-dried,
photographed and all colonies were counted. The assay was performed
three times in triplicate.
Western blot analysis
Control and garcinol-treated cells were washed twice
with cold phosphate-buffered saline (PBS) and then RIPA buffer was
added (cat. no. P0013B; Beyotime Institute of Biotechnology) with
PMSF (cat. no. ST506; Beyotime Institute of Biotechnology) and
protease inhibitor cocktail (cat. no. CW2383S; CWBIO) on ice. Cells
were detached with a scraper, and cell suspensions were transferred
to EP tubes and lysed on ice for 30 min. Cell lysates were
centrifuged at 12,000 × g at 4°C for 30 min and the supernatant was
collected and stored at −80°C until required for analysis. Protein
concentrations were determined by BCA protein assay kit (cat. no.
P0009; Beyotime Institute of Biotechnology). Total protein (20 µg)
was mixed with 5X loading buffer and denatured at 95°C for 8 min
before being separated on 10% SDS-PAGE (cat. no. 1610183; Bio-Rad
Laboratories) and electro-transferred onto polyvinylidene
difluoride (PVDF) membranes (cat. no. IPVH00010; EMD Millipore).
The membranes were blocked with 5% skimmed milk for 2 h at room
temperature and incubated with target antibodies on a shaking bed
overnight at 4°C. Primary antibodies used in the present study
were: Anti-p53 (cat. no. MA5-12557; mouse; 1:1,000) and anti-cyclin
D1 (cat. no. MA5-14512; rabbit; 1:800; both from Invitrogen; Thermo
Fisher Scientific, Inc.); anti-cyclin B1 (cat. no. GB11255; rabbit;
1:1,000; from Servicebio); anti-p21 (product code ab109520; mouse;
1:2,000; Abcam); anti-CDK2 (cat. no. sc-6248; mouse; 1:2,000; Santa
Cruz Biotechnology, Inc.); anti-β-actin (cat. no. A531; mouse;
1:5,000; Sigma-Aldrich; Merck KGaA); anti-SAPK/JNK (product no.
9252S; rabbit; 1:2,000); anti-c-JUN (product no. 9165S; rabbit;
1:1,000); phospho-SAPK/JNK (Thr183/Tyr185) (product no. 4668S;
rabbit; 1:1,000); and phospho-c-JUN (Ser73) (product no. 3270S;
rabbit; 1:1,000; all from Cell Signaling Technology, Inc.). After
removing the primary antibodies, membranes were washed with TBST
3×10 min. The membranes were then incubated with HRP-conjugated
secondary antibodies goat anti-rabbit IgG (H+L) (cat. no.
SA00001-2; 1:5,000) and goat anti-mouse IgG (H+L) (cat. no.
SA00001-1, 1:5,000; both from ProteinTech Group, Inc.) on a shaking
bed for 2 h at room temperature. After washing 3 times with TBST,
chemiluminescence substrate was used for development and signals
were captured by ChemiDoc XRS+ imaging system (Bio-Rad
Laboratories, Inc.). Data were analyzed by ImageJ 1.45 software
(NIH) and β-actin was used as the internal control. Each assay was
repeated 3 times.
Flow cytometric analysis
ISH and HEC-1B cells were seeded at 5×105
cells in 60 mm dishes. After 24 h, media with various
concentrations of garcinol (0, 10 and 20 µM) were added to cells to
treat for 48 h at 37°C. Cells were washed with PBS, digested to a
single cell suspension with 0.25% trypsin-EDTA solution for 3 min
at 37°C, and then centrifuged at 200 × g for 5 min at room
temperature to collect cells. After resuspension, cells were fixed
with chilled 75% alcohol at −20°C for 2 days. Cells were washed 3×
with ice cold PBS, stained with propidium iodide (PI) (50 µg/ml)
(cat. no. P4170) and RNase A (100 µg/ml) (cat. no. V900498; both
from Sigma-Aldrich; Merck KGaA) for 30 min at 37°C in the dark. A
FACS Aria II flow cytometer (BD Biosciences) was used to obtain
cell cycle data and data were analyzed by FLOWJO 7.6 (BD
Biosciences). Each experiment was repeated 3 separate times.
5-Ethynyl-2′-deoxyuridine (EdU)
incorporation assay
Cells (1.5×104) were seeded in 8-well
Nunc Lab-Tek II chamber slides (cat. no. 154534; Thermo Fisher
Scientific, Inc.) and treated with various concentrations of
garcinol (0, 10 and 20 µM) for 48 h at 37°C when 10 µM EdU
(BeyoClick™ EdU Cell Proliferation Kit with Alexa Fluor 488; cat.
no. C0071L, Beyotime, Shanghai, China) used according to the
manufacturer's instructions was added to the cell culture and the
cells were incubated for an additional 2 h at 37°C. The cells were
then fixed with 4% paraformaldehyde at room temperature for 30 min,
permeabilized with 0.3% Triton X-100 for 15 min, and then click
additive solution contained in the aforementioned kit was added and
the cells were incubated on a shaking bed for 30 min at 37°C in the
dark. The cell nucleus was stained with Hoechst 33342 (included in
the aforementioned kit) for 15 min at room temperature. Images were
captured by Leica DM4B Fluorescence Microsystems using a 20X
objective. Data are presented as the ratio of the
fluorescent-positive cells to total cells. Each experiment was
repeated 3 separate times.
Statistical analysis
Data are presented as the mean ± standard deviations
(SD). Statistical analysis was performed using the SPSS 16.0 (IBM
Corp.). One-way analysis of variation (ANOVA) with Dunnett's
multiple comparisons test was used to compare individual data with
the control values. P<0.05 was considered to indicate a
statistically significant difference. All experiments were repeated
at least three times.
Results
Garcinol inhibits EC cell
proliferation
To investigate the effect of garcinol on EC
proliferation, RTCA and a cell counting assay were performed with
ISH and HEC-1B cells. In the RTCA assay, garcinol inhibited cell
proliferation of both ISH and HEC-1B cells in a dose- and
time-dependent manner (Fig. 1A-D).
This result was confirmed in both cell lines by traditional cell
counting methodology (Fig. 1E and
F). Data revealed that ISH cells were more sensitive than
HEC-1B cells to garcinol treatment, since 5 µM garcinol could
significantly inhibit cell proliferation in ISH cells from 48 to 72
h, but that concentration of garcinol could only inhibit the
proliferation of HEC-1B cells at 60 h (Fig. 1B and D).
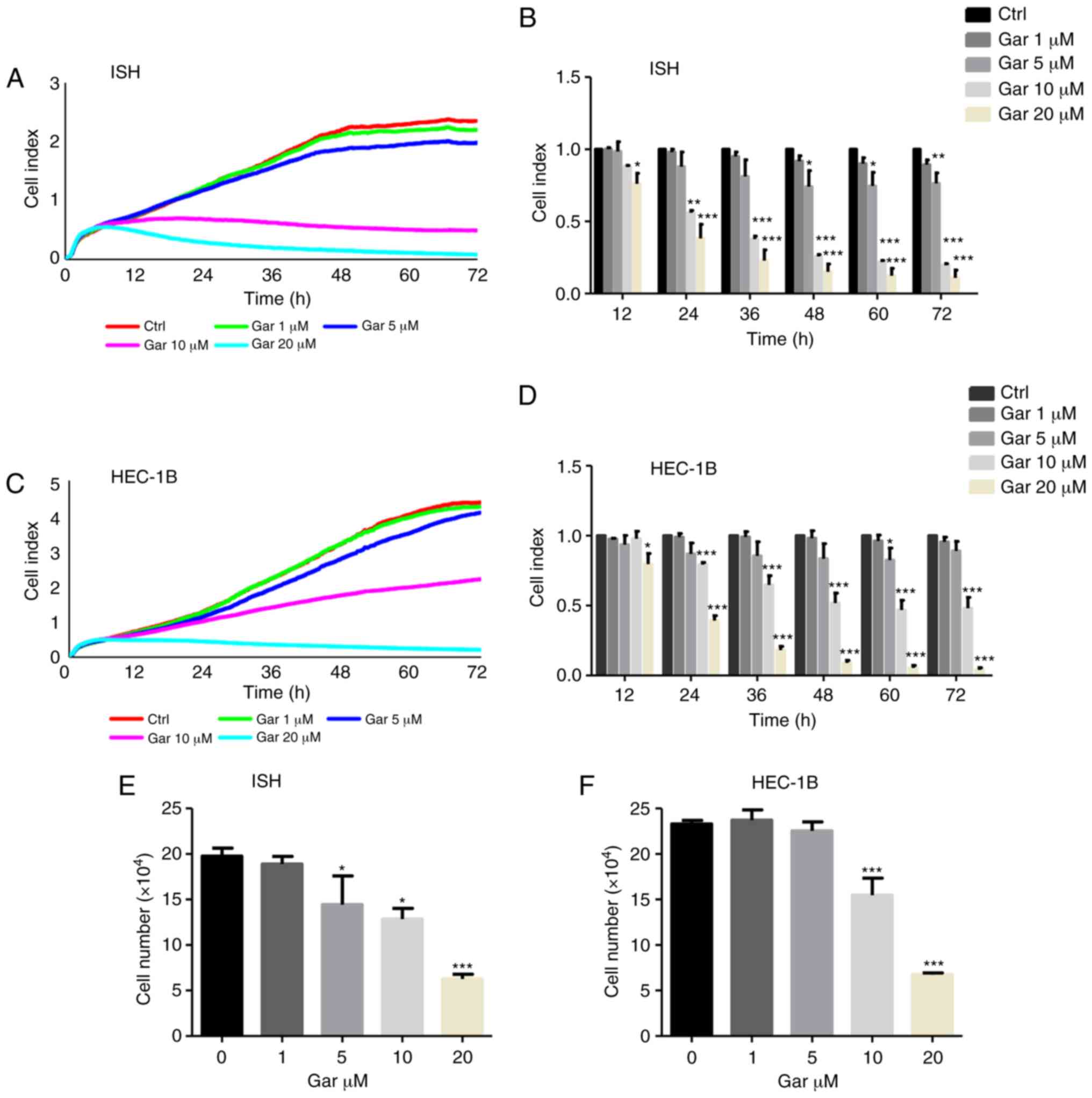 | Figure 1.Garcinol inhibits the proliferation
of EC cells. (A-D) Proliferation assays of EC (A and B) ISH cells
and (C and D) HEC-1B cells, exposed to 0, 1, 5, 10 and 20 µM
garcinol for 72 h and detected by label-free RTCA. Data are
presented as cell index curves, which were recorded by the RTCA
instrument for (A) ISH cells and (C) HEC-1B cells. Data at
time-points 12, 24, 36, 48, 60 and 72 h were analyzed separately
for (B) ISH cells and (D) HEC-1B cells. (E and F) Cell counting
results of (E) ISH cells and (F) HEC-1B cells, which were treated
with garcinol (0, 1, 5, 10 and 20 µM) for 48 h. Data are presented
as the mean ± SD, n=3. *P<0.05, **P<0.01, ***P<0.001 as
compared with the control group, indicate statistical significance
by one-way ANOVA with Dunnett's multiple comparisons test. EC,
endometrial cancer; ISH, Ishikawa; RTCA, real-time cellular
analysis; Gar, garcinol. |
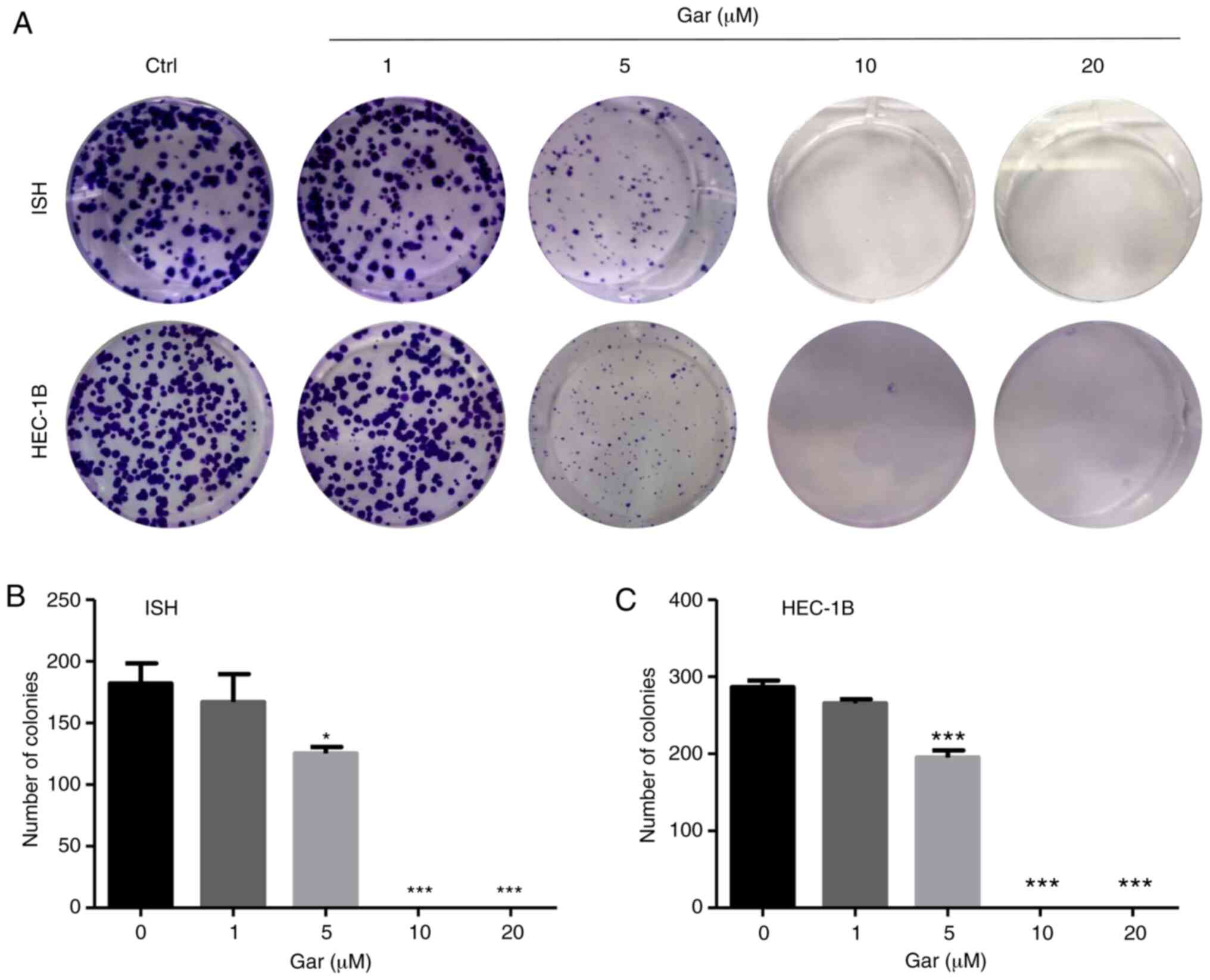
Garcinol inhibits the colony formation
ability of EC cells
After 14 days of continuous culture in various
concentrations of garcinol, colony formation of both ISH and HEC-1B
cells was significantly inhibited (Fig.
2). Treatment with 5 µM garcinol resulted in fewer, smaller
colonies in both cell lines compared with the control. Treatment
with 10 and 20 µM garcinol completely inhibited colony formation.
However, even after a long culture time, 1 µM garcinol could not
significantly decrease the ability of colony formation in both
types of cells.
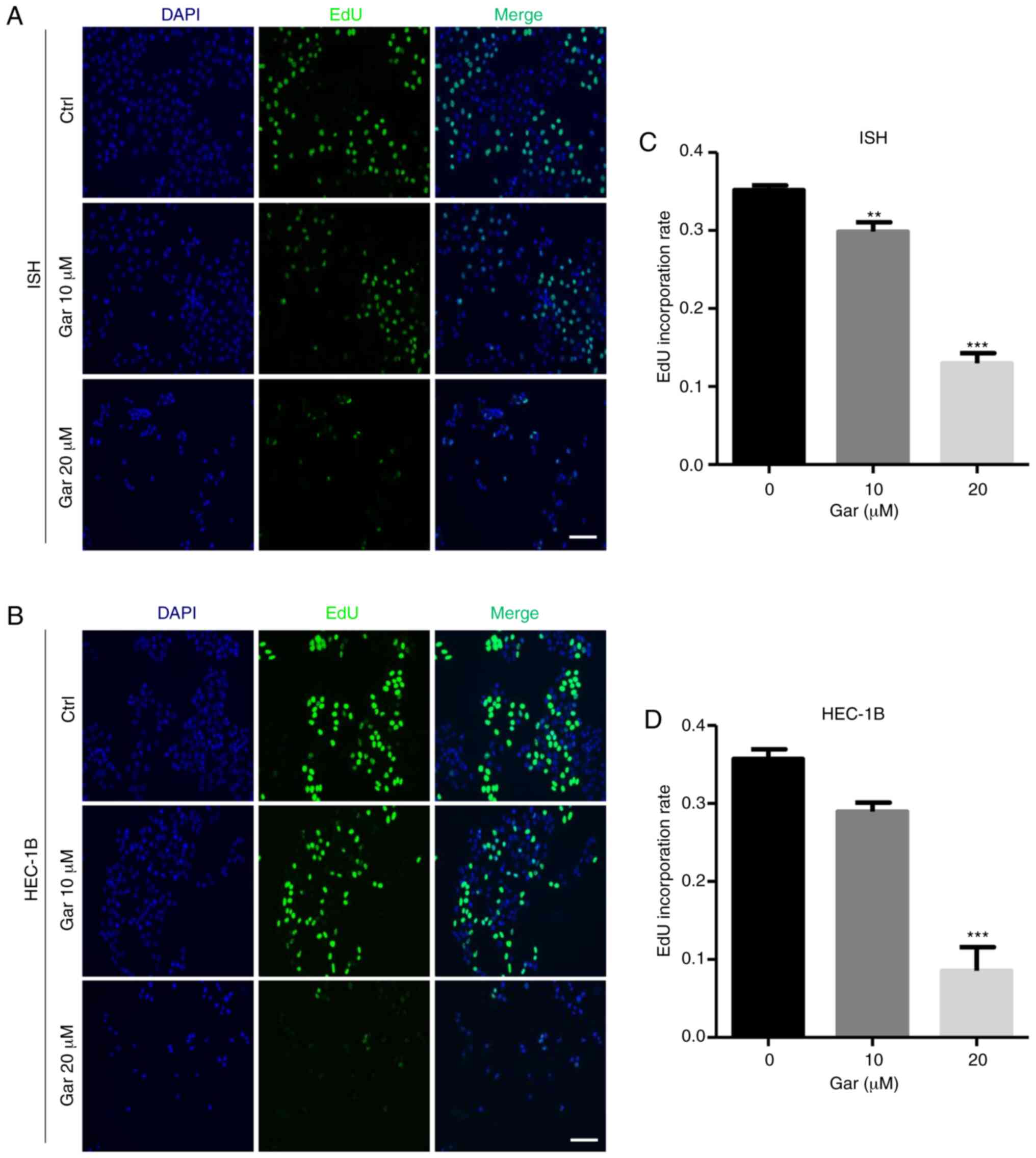
Garcinol attenuates S-phase DNA
synthesis in EC cells
To further assess the anti-proliferation function of
garcinol, an EdU incorporation assay was used to directly detect
DNA replication. After garcinol treatment, the percentage of
EdU-positive cells was significantly decreased in the 10 and 20 µM
garcinol-treated ISH cells and 20 µM HEC-1B cells compared with the
control of both types of EC cells (Fig.
3). The result demonstrated that garcinol can attenuate S-phase
DNA synthesis in EC cells to exert its anticancer effect.
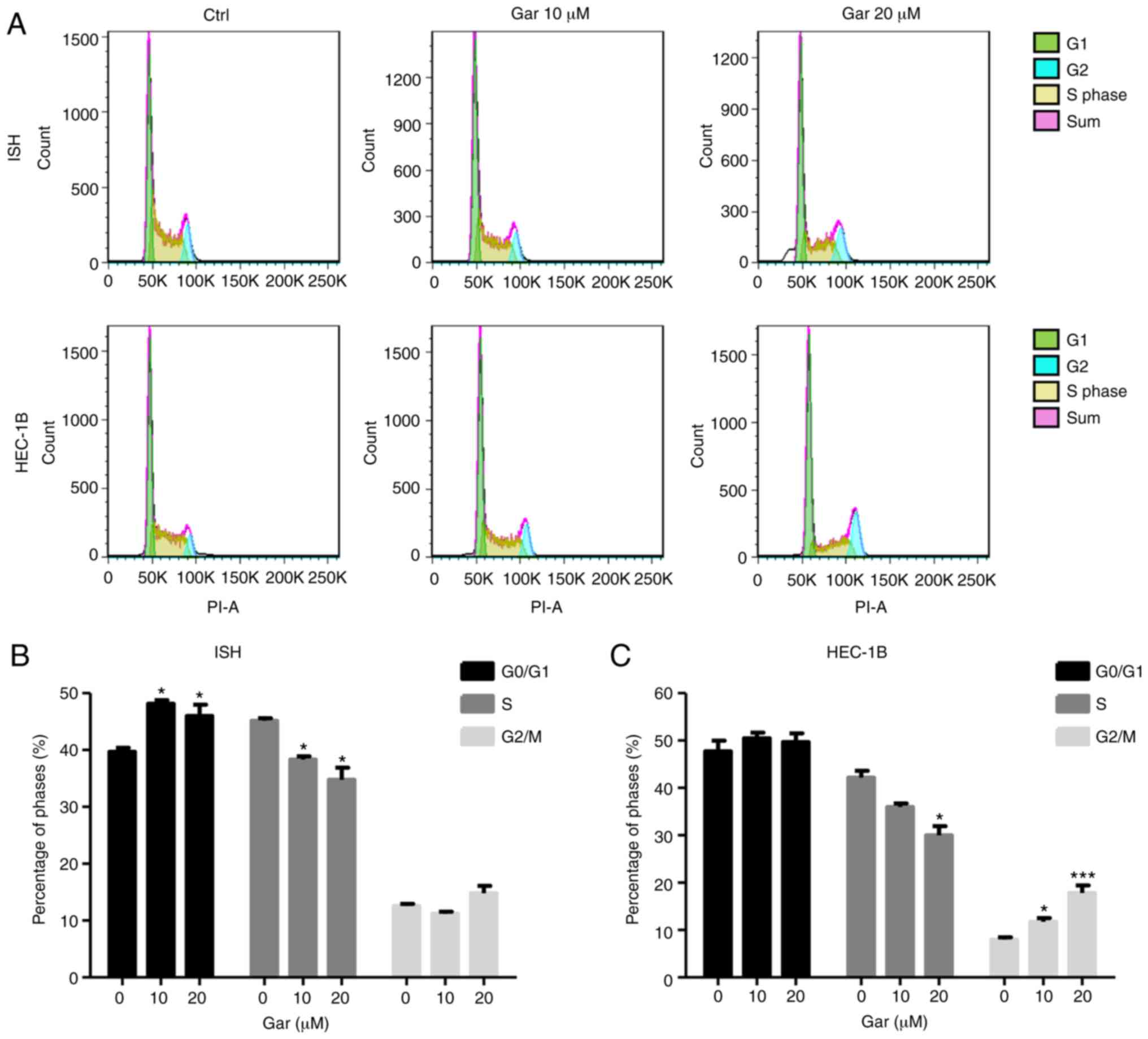
Garcinol induces cell cycle arrest in
ISH and HEC-1B cells
The cell cycle is an important physiological process
which controls proliferation, growth and survival of cells. Cell
cycle arrest can inhibit cancer cell proliferation, thus numerous
anticancer drugs use this mechanism for cancer therapy (20,27).
After garcinol treatment for 48 h, ISH cells were arrested in the
G1 phase, while HEC-1B cells were arrested in the G2 phase, and
garcinol reduced the number of cells in the S phase in both cell
types (Fig. 4). The cell cycle
results indicated that garcinol can inhibit cell proliferation by
inducing cell cycle arrest in EC, although the exact mechanism may
differ for different EC cell types.
Garcinol regulates the expression of
cell cycle-related genes in EC cells
Cell cycle progression is tightly controlled by cell
cycle-related genes (22,28). Therefore, examining the expression
of those genes/proteins may elucidate how garcinol regulates the
cell cycle in EC cells. Western blotting demonstrated that the
expression of p53 and p21 was significantly increased, while the
expression of CDK2, CDK4, cyclin D1 and cyclin B1 (22) was gradually decreased in a
dose-dependent manner in both ISH and HEC-1B cell lines (Fig. 5). Moreover, p53 expression was more
sensitive than the other genes to garcinol stimulation in ISH
cells, and even 1 µM garcinol could induce its expression.
Therefore, garcinol could induce cell cycle arrest through
regulating the expression of cell cycle-related proteins in EC
cells.
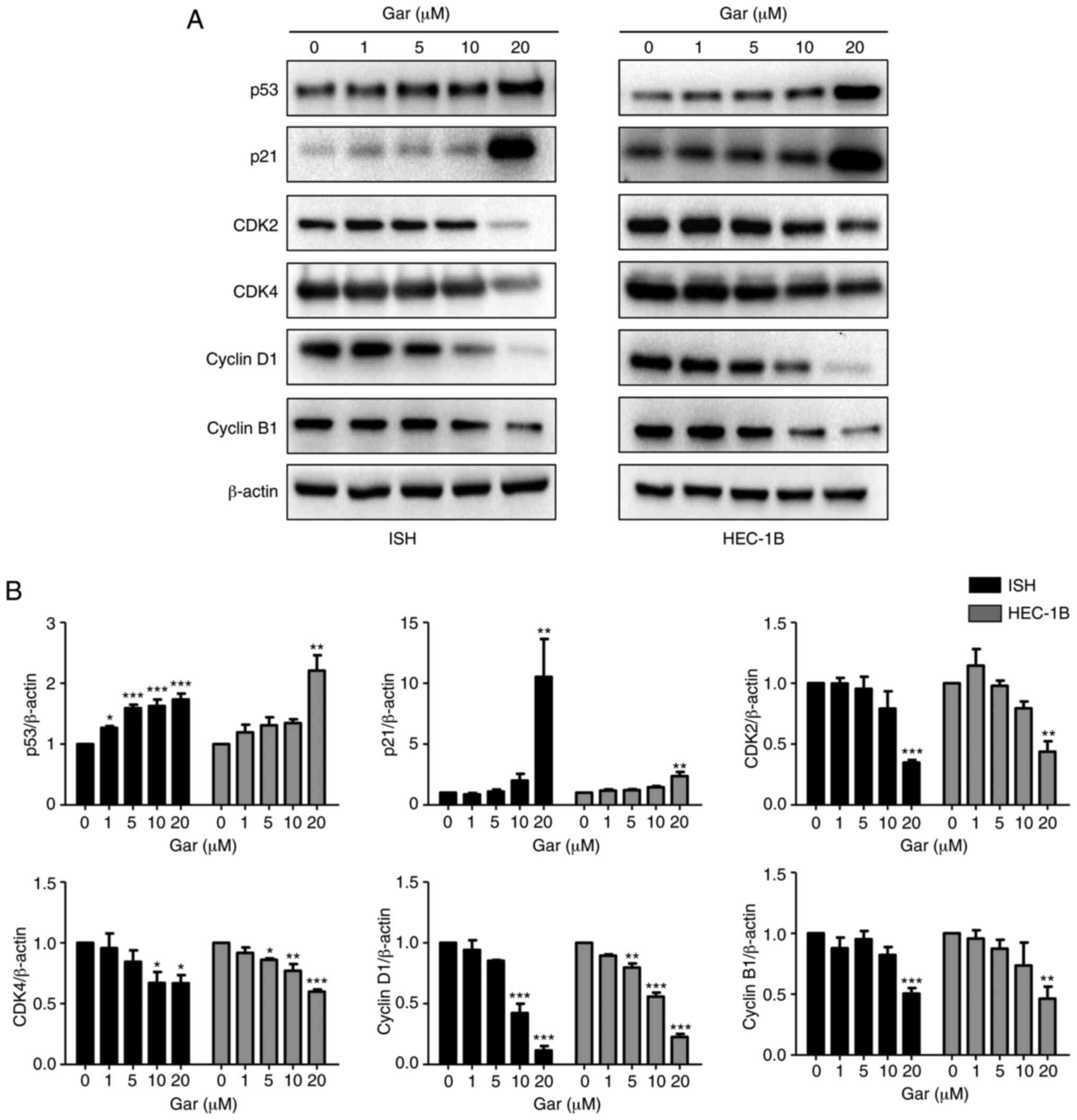 | Figure 5.Garcinol regulates the expression of
cell cycle-related proteins in EC cells. Effect of garcinol on the
expression of cell cycle-regulated genes in EC cells were detected
by western blotting. ISH and HEC-1B cells were cultured for 24 h
and then treated with and without garcinol (0, 1, 5, 10 and 20 µM)
for another 24 h. (A) Total protein was collected, and western blot
analyses were performed for p53, p21, cyclin D1, cyclin B1, CDK2,
CDK4. β-actin was used as α loading control. (B) Band density was
analyzed by ImageJ, and the results are expressed as the mean ± SD,
n=3. *P<0.05, **P<0.01 and ***P<0.001 compared with the
control, indicate statistical significance by one-way ANOVA with
Dunnett's multiple comparisons test. EC, endometrial cancer; ISH,
Ishikawa; Gar, garcinol. |
Garcinol may activate the JNK/c-JUN
signaling pathway in EC cells
The MAPK signaling pathway is a one of the key
pathways regulating cell proliferation, the cell cycle and
apoptosis. In the present study, ISH and HEC-1B cells were treated
with 20 µM garcinol for 30, 60, 90 and 120 min, and phosphorylation
levels of JNK and c-JUN were detected by western blotting. Garcinol
treatment increased phosphorylation levels of JNK and c-JUN, in
both of ISH and HEC-1B cells (Fig.
6). p-JNK was significantly increased at 60 and 90 min in ISH
cells, while it was significantly increased at 30, 90 and 120 min
in HEC-1B cells. p-c-JUN was significantly increased at 90 and 120
min in ISH cells, and for HEC-1B cells, it was significantly
increased from 30 to 120 min. Therefore, garcinol may increase
JNK/c-JUN signaling to regulate the cell cycle in EC.
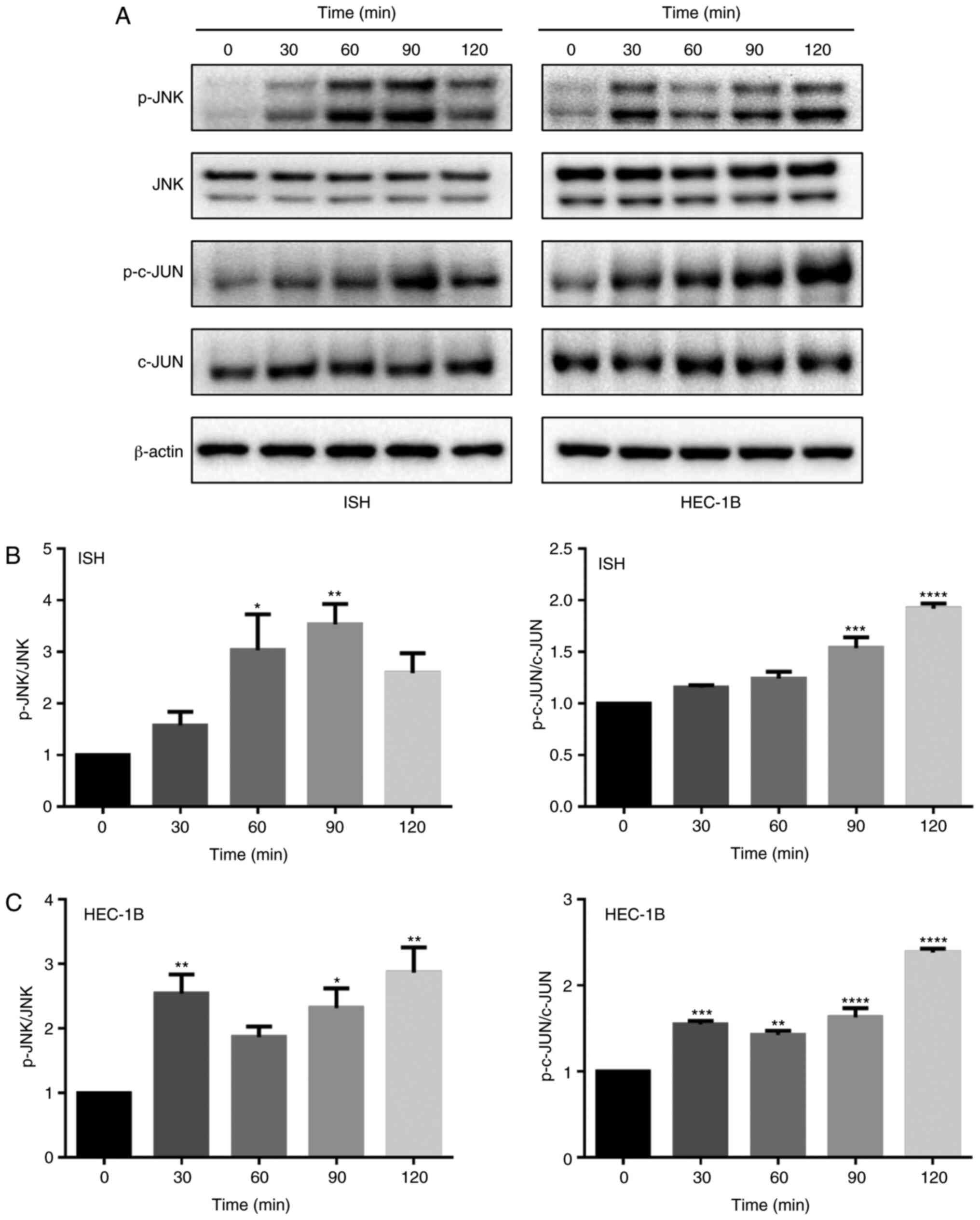 | Figure 6.Garcinol activates the JNK/c-JUN
signaling pathway in EC cells. (A) The effect of garcinol on JNK
and c-JUN expression, and their phosphorylation levels in EC cells.
ISH and HEC-1B cell were treated with garcinol (20 µM) for 0, 30,
60, 90 and 120 min. Western blot analyses were performed with
anti-p-JNK, JNK, p-c-JUN and c-JUN antibodies. β-actin was used as
a loading control. (B and C) Band density was analyzed by ImageJ,
and the results are expressed as the mean ± SD, n=3. *P<0.05,
**P<0.01, ***P<0.001 and ****P<0.0001 compared with the
control, indicate statistical significance by one-way ANOVA with
Dunnett's multiple comparisons test. JNK, c-JUN N-terminal kinase;
EC, endometrial cancer; ISH, Ishikawa; Gar, garcinol. |
Discussion
EC is the most common cancer of the female
reproductive system and an important cause of cancer-related deaths
in women worldwide (29). The
incidence and mortality of EC are increasing rapidly worldwide
(30). The 5-year survival rate is
more than 90% in patients with early-stage disease, treated with
surgery and then brachytherapy or external beam radiation therapy
(29,31). However, the prognosis of patients
with distant metastasis is poor, with the 5-year survival rate less
than 20% (32). The standard
treatment for EC is surgery, and platinum-based chemotherapy and
radiotherapy are recommended as adjuvant therapy for patients with
high histological grade or metastasis (3). However, it is becoming increasingly
important to develop new anticancer drugs. Previous studies
indicated that garcinol could inhibit cancer cell proliferation and
induce cell cycle arrest and apoptosis (12,24,26).
In the present study, it was demonstrated that garcinol could
inhibit the proliferation and cell cycle progression of ISH and
HEC-1B cells.
The cell cycle has three important checkpoints
G1-to-S, G2-to-M and M-to-G1. However, cell cycle arrest at any
point will finally lead to diminished cell proliferation. In cancer
cells, minimal blocks at the G1-S transition point drives cells
into the S phase, resulting in uncontrolled growth, therefore
investigation of agents which target the cell cycle is a viable
option for therapeutics (28).
Several previous studies have also reported that garcinol treatment
arrested the cell cycle at the G0/G1 phase in breast cancer MCF-7
cells, lung cancer cells, and cervical cancer cells (17,19,26,33).
Garcinol has also been revealed to arrest oral cancer cells at the
S phase (24), and 3T3-L1 cells at
the G2/M phase (34). The present
study also revealed that garcinol caused cell cycle arrest in EC,
although its effect was different in the two different cell lines
investigated. ISH cells were arrested at the G1 phase and HEC-1B
cells were arrested at the G2 phase, although both cell types had a
decrease in the number of cells in the S phase, which corresponded
to a decrease in S-phase DNA synthesis. The differences in
mechanism of cell cycle arrest between these two cell types may be
determined by their variant genetic characteristics. The ISH cell
line is derived from a well differentiated adenocarcinoma, with
high expression levels of ER-α, ER-β and PR, but low expression
levels of hMLH-1 and PTEN (35).
The HEC-1B cell line is derived from a moderately differentiated
adenocarcinoma, with low expression levels of ER-α, ER-β and PR,
but high expression levels of hMLH-1 and PTEN (35). However, whether these genetic
differences have effects, or how they are involved in the effects
of garcinol in EC remain to be further investigated. These studies
all support that garcinol decreases cancer cell proliferation by
targeting the cell cycle, although the exact mode of action may
differ depending on the cancer cell type.
The progression of the cell cycle is regulated by
cyclins and CDK complexes. CDKs are activated by cyclins and
suppressed by cyclin-dependent kinase inhibitors at cell cycle
checkpoints or specific phases (20,22).
Garcinol has been revealed to inhibit cervical cancer cell cycle
progression at the G0/G1 phase, via a mechanism associated with
downregulation of cyclin D1 and CDK4, and upregulation of p21 and
p53 (19). In lung cancer cells,
garcinol treatment downregulated the expression of CDK2, CDK4 and
cyclin D, leading to the H1299 lung cancer cells being arrested at
the G1 phase (26). In the present
study it was demonstrated that garcinol treatment upregulated the
expression of the tumor suppressor proteins p53 and p21, and
downregulated the expression of their target proteins CDK4, cyclin
D1 and cyclin B1 in ISH and HEC-1B cells. However, garcinol
arrested the cell cycle at different points in these two cell lines
suggesting some other, as yet unidentified, pathways may also be
involved in garcinol cell cycle arrest in different EC cell
lines.
The mitogen-activated protein kinase (MAPK)
signaling pathway is an important pathway in cancer, and has been
developed as a target for cancer treatment. It is known that JNK,
ERK and p38 are all members of the MAPK pathway which are involved
in cell proliferation, differentiation, cell cycle and apoptosis
(25). Numerous anticancer drugs
exert their effects through the p38 MAPK or ERK/MAPK signaling
pathways to control cell proliferation (23,25,26,36).
c-JUN is the major downstream target of JNK signaling, and
transcriptional regulation of c-JUN and its target genes is one of
the main functions of JNK (37).
The MAPK signaling pathway has been reported to play a role in
various types of cancer including prostate, colorectal and
pancreatic cancers and has been developed as a therapeutic target
(38). The present study revealed
that after garcinol treatment the expression of p53, a downstream
target of the JNK/c-JUN pathway, was increased in EC. Therefore, it
was determined whether garcinol inhibited EC cell proliferation
through the JNK/c-JUN signaling pathway. As a cell permeable drug,
garcinol may affect signaling pathways in cells to regulate the
proliferation and the cell cycle in EC cells. Garcinol increased
phosphorylation of both JNK and c-JUN in both EC cell types
investigated. The regulation of the JNK signaling pathway may be
one of the signaling mechanisms by which garcinol inhibits
proliferation of EC. The activation of p-JNK has also been found in
response to other anticancer agents such as huaier, aplidin and
adaphostin (25,37,39,40).
JNK in turn can phosphorylate and activate the transcription factor
activator protein-1 (AP-1), and activating transcription factor-2,
c-Myc, p53, and Bcl-2 family proteins, with these proteins
controlling various cellular responses, such as proliferation,
differentiation, apoptosis, the cell cycle and autophagy (25,37,39,40).
Since the JNK/c-JUN pathway was activated and its downstream target
gene p53 was significantly increased after garcinol treatment, the
JNK/c-JUN pathway may play important roles in garcinol-induced EC
cell cycle arrest. However, it is also possible that other pathways
may also be involved.
As a tumor suppressor gene, p53 induces cell cycle
arrest mainly through transcriptional activation of p21, which in
turn binds to cyclin D/CDK4 complexes and may cause G1 phase arrest
(41,42). Garcinol treatment induced H1299 lung
cancer cell G1 arrest through p21Waf1/Cip1 and
p27Kip1 activation, and subsequent inhibition of CDK
activity (26). p53 acts mainly
through the transcriptional activation of p21, and p21 binding to
cyclin D/CDK4 complexes to cause G1 phase arrest (42). Therefore, it was demonstrated that
garcinol may act by upregulating the expression of p21 and p53 and
downregulating cyclin D1 and CDK4 to induce ISH cell arrest in the
G1/S phase. However, p53 activation can not only arrest cells at
the G0/G1 checkpoint, but also at the G2/M checkpoint, because p21
can also inhibit cyclin B/CDK1 to inhibit cell-cycle progression,
and other p53 target genes such as 14-3-3σ GADD45, may interrupt
the interaction of Cdc2/cyclin B1 and participate in the arrest of
G2/M transition (43,44). In the present study it was
demonstrated that the expression of cyclin B1 was reduced, fitting
with cell cycle arrest in the G2/M phase in HEC-1B cells induced by
garcinol. Therefore, garcinol may act through activation of p53,
upregulation of p21 leading to inhibition of cyclin B1 and finally
causing HEC-1B cell arrest in the G2/M phase.
Previous studies have suggested that garcinol could
inhibit cancer cell proliferation through inhibition of STAT-3,
NF-κB and PI3K/AKT signaling pathways (16,19,24).
However, in the present study, one of the limitations was that we
only investigated the role of garcinol in the JNK/c-JUN pathway in
EC. In addition, garcinol is an inhibitor of the histone
acetyltransferases (HAT) p300 and Pcaf (45), with numerous roles in neurocytes
(46–49), immunocytes (50,51)
and cancer cells. Therefore, further in vitro studies should
also investigate other potential signaling pathways, as well as
other potential targets of garcinol activity including apoptosis,
autophagy and invasion. To further verify the potential use of
garcinol as a therapeutic for EC its effects on primary EC cells
should also be investigated in vitro, as well as in in
vivo animal models, particularly focusing on metastatic
disease.
In conclusion, in the present study evidence was
provided that garcinol can inhibit cell growth and induce cell
cycle arrest in EC, by regulating the expression of cell
cycle-related genes and the JNK/c-JUN signaling pathway. Therefore,
garcinol may be a promising candidate agent for EC
chemotherapy.
Acknowledgements
We appreciate Mrs. Shang Wang and Mr. Xiaobin Liu at
Guangzhou Women and Children's Medical Center (Guangzhou, China)
for their assistance with flow cytometry and fluorescence
microscopy, respectively.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81702567, 81671406
and 31871412), the Science and Technology Programs of Guangzhou
City (grant no. 201704030103), the Post-doc Initiation Fund of
Guangzhou (grant no. 3302), and the Post-doc Science Research
Initiation Fund of Guangzhou Women and Children's Medical Center
(grant no. 20160322).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
MZ and QL performed the cell culture, cell counting
assays and flow cytometric analysis. MZ performed the RTCA and EdU
assays. MZ, HH, DS, MC, FN, PW, YD, YP, and DW performed the
western blot experiments. MZ, QL and GEL designed the study and
performed the analyses. MZ and QL wrote the manuscript and GEL
revised the manuscript. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lee YC, Lheureux S and Oza AM: Treatment
strategies for endometrial cancer: Current practice and
perspective. Curr Opin Obstet Gynecol. 29:47–58. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sorosky JI: Endometrial cancer. Obstet
Gynecol. 120:383–397. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mitamura T, Dong P, Ihira K, Kudo M and
Watari H: Molecular-targeted therapies and precision medicine for
endometrial cancer. Jpn J Clin Oncol. 49:108–120. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oza AM, Elit L, Tsao MS, Kamel-Reid S,
Biagi J, Provencher DM, Gotlieb WH, Hoskins PJ, Ghatage P, Tonkin
KS, et al: Phase II study of temsirolimus in women with recurrent
or metastatic endometrial cancer: A trial of the NCIC clinical
trials group. J Clin Oncol. 29:3278–3285. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ushijima K: Hormonal treatment of
endometrial cancer. Nihon Rinsho. 70 (Suppl 4):388–393. 2012.(In
Japanese). PubMed/NCBI
|
|
6
|
Ito K, Utsunomiya H, Niikura H, Yaegashi N
and Sasano H: Inhibition of estrogen actions in human gynecological
malignancies: New aspects of endocrine therapy for endometrial
cancer and ovarian cancer. Mol Cell Endocrinol. 340:161–167. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miller KD, Fidler-Benaoudia M, Keegan TH,
Hipp HS, Jemal A and Siegel RL: Cancer statistics for adolescents
and young adults, 2020. CA Cancer J Clin. 17:Sep 17–2020.(Epub
ahead of print. doi.org/10.3322/caac.21637 2020.
|
|
8
|
Lindemann K, Eskild A, Vatten LJ and Bray
F: Endometrial cancer incidence trends in norway during 1953-2007
and predictions for 2008-2027. Int J Cancer. 127:2661–2668. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sung H, Siegel RL, Rosenberg PS and Jemal
A: Emerging cancer trends among young adults in the USA: Analysis
of a population-based cancer registry. Lancet Public Health.
4:e137–e147. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jayaprakasha GK and Sakariah KK:
Determination of organic acids in leaves and rinds of Garcinia
indica (Desr.) by LC. J Pharm Biomed Anal. 28:379–384. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schobert R and Biersack B: Chemical and
biological aspects of garcinol and isogarcinol: Recent
developments. Chem Biodivers. 16:e19003662019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu C, Ho PC, Wong FC, Sethi G, Wang LZ
and Goh BC: Garcinol: Current status of its anti-oxidative,
anti-inflammatory and anti-cancer effects. Cancer Lett. 362:8–14.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saadat N and Gupta SV: Potential role of
garcinol as an anticancer agent. J Oncol. 2012:6472062012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Padhye S, Ahmad A, Oswal N and Sarkar FH:
Emerging role of Garcinol, the antioxidant chalcone from
Garcinia indica Choisy and its synthetic analogs. J Hematol
Oncol. 2:382009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang L, Wang M, Guo H and Zhao H: Emerging
role of garcinol in targeting cancer stem cells of non-small cell
lung cancer. Curr Pharmacol Rep. 5:14–19. 2019. View Article : Google Scholar
|
|
16
|
Ahmad A, Sarkar SH, Aboukameel A, Ali S,
Biersack B, Seibt S, Li Y, Bao B, Kong D, Banerjee S, et al:
Anticancer action of garcinol in vitro and in vivo is in part
mediated through inhibition of STAT-3 signaling. Carcinogenesis.
33:2450–2456. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ye X, Yuan L, Zhang L, Zhao J, Zhang CM
and Deng HY: Garcinol, an acetyltransferase inhibitor, suppresses
proliferation of breast cancer cell line MCF-7 promoted by
17β-estradiol. Asian Pac J Cancer Prev. 15:5001–5007. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang J, Wu M, Zheng D, Zhang H, Lv Y,
Zhang L, Tan HS, Zhou H, Lao YZ and Xu HX: Garcinol inhibits
esophageal cancer metastasis by suppressing the p300 and TGF-β1
signaling pathways. Acta Pharmacol Sin. 41:82–92. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao J, Yang T, Ji J, Li C, Li Z and Li L:
Garcinol exerts anti-cancer effect in human cervical cancer cells
through upregulation of T-cadherin. Biomed Pharmacother.
107:957–966. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Malumbres M and Barbacid M: Mammalian
cyclin-dependent kinases. Trends Biochem Sci. 30:630–641. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Massagué J: G1 cell-cycle control and
cancer. Nature. 432:298–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ryu M, Sung CK, Im YJ and Chun C:
Activation of JNK and p38 in MCF-7 cells and the in vitro
anticancer activity of alnus hirsuta extract. Molecules.
25:10732020. View Article : Google Scholar
|
|
24
|
Aggarwal S and Das SN: Garcinol inhibits
tumour cell proliferation, angiogenesis, cell cycle progression and
induces apoptosis via NF-κB inhibition in oral cancer. Tumour Biol.
37:7175–7184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan L, Liu X, Yin A, Wei Y, Yang Q and
Kong B: Huaier aqueous extract inhibits cervical cancer cell
proliferation via JNK/p38 pathway. Int J Oncol. 47:1054–1060. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu SY, Liao CH, Chien MH, Tsai TY, Lin JK
and Weng MS: Induction of p21(Waf1/Cip1) by garcinol via
downregulation of p38-MAPK signaling in p53-independent H1299 lung
cancer. J Agric Food Chem. 62:2085–2095. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
George VC, Kumar DR and Kumar RA: Relative
in vitro potentials of parthenolide to induce apoptosis and cell
cycle arrest in skin cancer cells. Curr Drug Discov Technol.
13:34–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bertoli C, Skotheim JM and de Bruin RA:
Control of cell cycle transcription during G1 and S phases. Nat Rev
Mol Cell Biol. 14:518–528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Giannone G, Tuninetti V, Ghisoni E, Genta
S, Scotto G, Mittica G and Valabrega G: Role of cyclin-dependent
kinase inhibitors in endometrial cancer. Int J Mol Sci.
20:23532019. View Article : Google Scholar
|
|
30
|
Cai Z and Liu Q: Understanding the global
cancer statistics 2018: Implications for cancer control. Sci China
Life Sci. Aug 26–2019.(Epub ahead of print). doi:
10.1007/s11427-019-9816-1. View Article : Google Scholar
|
|
31
|
Signorelli M, Lissoni AA, Cormio G,
Katsaros D, Pellegrino A, Selvaggi L, Ghezzi F, Scambia G, Zola P,
Grassi R, et al: Modified radical hysterectomy versus extrafascial
hysterectomy in the treatment of stage I endometrial cancer:
Results from the ILIADE randomized study. Ann Surg Oncol.
16:3431–3441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dowdy SC: Improving oncologic outcomes for
women with endometrial cancer: Realigning our sights. Gynecol
Oncol. 133:370–374. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Collins HM, Abdelghany MK, Messmer M, Yue
B, Deeves SE, Kindle KB, Mantelingu K, Aslam A, Winkler GS, Kundu
TK and Heery DM: Differential effects of garcinol and curcumin on
histone and p53 modifications in tumour cells. BMC Cancer.
13:372013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hsu CL, Lin YJ, Ho CT and Yen GC:
Inhibitory effects of garcinol and pterostilbene on cell
proliferation and adipogenesis in 3T3-L1 cells. Food Funct.
3:49–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qu W, Zhao Y, Wang X, Qi Y, Zhou C, Hua Y,
Hou J and Jiang SW: Culture characters, genetic background,
estrogen/progesterone receptor expression, and tumorigenic
activities of frequently used sixteen endometrial cancer cell
lines. Clin Chim Acta. 489:225–232. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lim W, Jeong M, Bazer FW and Song G:
Coumestrol inhibits proliferation and migration of prostate cancer
cells by regulating AKT, ERK1/2, and JNK MAPK cell signaling
cascades. J Cell Physiol. 232:862–871. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kumar A, Singh UK, Kini SG, Garg V,
Agrawal S, Tomar PK, Pathak P, Chaudhary A, Gupta P and Malik A:
JNK pathway signaling: A novel and smarter therapeutic targets for
various biological diseases. Future Med Chem. 7:2065–2086. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sebolt-Leopold JS and Herrera R: Targeting
the mitogen-activated protein kinase cascade to treat cancer. Nat
Rev Cancer. 4:937–947. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cuadrado A, Garcia-Fernandez LF, Gonzalez
L, Suarez Y, Losada A, Alcaide V, Martinez T, Fernandez-Sousa JM,
Sanchez-Puelles JM and Munoz A: Aplidin induces apoptosis in human
cancer cells via glutathione depletion and sustained activation of
the epidermal growth factor receptor, Src, JNK, and p38 MAPK. J
Biol Chem. 278:241–250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yu C, Rahmani M, Almenara J, Sausville EA,
Dent P and Grant S: Induction of apoptosis in human leukemia cells
by the tyrosine kinase inhibitor adaphostin proceeds through a
RAF-1/MEK/ERK- and AKT-dependent process. Oncogene. 23:1364–1376.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Harvey M, Sands AT, Weiss RS, Hegi ME,
Wiseman RW, Pantazis P, Giovanella BC, Tainsky MA, Bradley A and
Donehower LA: In vitro growth characteristics of embryo fibroblasts
isolated from p53-deficient mice. Oncogene. 8:2457–2467.
1993.PubMed/NCBI
|
|
42
|
Chen J: The cell-cycle arrest and
apoptotic functions of p53 in tumor initiation and progression.
Cold Spring Harb Perspect Med. 6:a0261042016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Martín-Caballero J, Flores JM,
García-Palencia P and Serrano M: Tumor susceptibility of
p21(Waf1/Cip1)-deficient mice. Cancer Res. 61:6234–6238.
2001.PubMed/NCBI
|
|
44
|
Taylor WR and Stark GR: Regulation of the
G2/M transition by p53. Oncogene. 20:1803–1815. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Balasubramanyam K, Altaf M, Varier RA,
Swaminathan V, Ravindran A, Sadhale PP and Kundu TK:
Polyisoprenylated benzophenone, garcinol, a natural histone
acetyltransferase inhibitor, represses chromatin transcription and
alters global gene expression. J Biol Chem. 279:33716–33726. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang YW, Zhang X, Chen CL, Liu QZ, Xu JW,
Qian QQ, Li WY and Qian YN: Protective effects of Garcinol against
neuropathic pain-evidence from in vivo and in vitro studies.
Neurosci Lett. 647:85–90. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dunbar AB and Taylor JR: Garcinol blocks
the reconsolidation of multiple cocaine-paired cues after a single
cocaine-reactivation session. Neuropsychopharmacology.
42:1884–1892. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hao F, Jia LH, Li XW, Zhang YR and Liu XW:
Garcinol upregulates GABAA and GAD65 expression, modulates
BDNF-TrkB pathway to reduce seizures in pentylenetetrazole
(PTZ)-induced epilepsy. Med Sci Monit. 22:4415–4425. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Weng MS, Liao CH, Yu SY and Lin JK:
Garcinol promotes neurogenesis in rat cortical progenitor cells
through the duration of extracellular signal-regulated kinase
signaling. J Agric Food Chem. 59:1031–1040. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li W, Li H, Zhang M, Zhong Y, Wang M, Cen
J, Wu H, Yang Y and Wei Q: Isogarcinol extracted from Garcinia
mangostana L. ameliorates systemic lupus erythematosus-like
disease in a murine model. J Agric Food Chem. 63:8452–8459. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cui ZL, Gu W, Ding T, Peng XH, Chen X,
Luan CY, Han RC, Xu WG and Guo XJ: Histone modifications of Notch1
promoter affect lung CD4+ T cell differentiation in asthmatic rats.
Int J Immunopathol Pharmacol. 26:371–381. 2013. View Article : Google Scholar : PubMed/NCBI
|