Introduction
Oral squamous cell carcinoma (OSCC) is a major
burden to human health according to a 2019 epidemiological
investigation of cancer (1). The
major carcinogenic factors of OSCC are tobacco, alcohol and
ultraviolet rays. The typical clinical features of OSCC are obscure
boundaries, bleeding and ulcers that affect the chewing, speech and
appearance of patients (2). The
biological behavior of OSCC cells, including rapid proliferation,
resistance to apoptosis, rapid invasion and epithelial-mesenchymal
transition, is related to the OSCC clinical phenotype (3). With the rapid development of molecular
biology, it has been revealed that the expression of some
intracellular molecules is correlated with the biological behavior
of OSCC (4,5), and it has been demonstrated that high
expression of Yes-associated protein (YAP) results in a rapid cell
proliferation rate and poor differentiation of OSCC (6,7). Our
previous study also revealed that YAP could be transported into the
nucleus, driving the transcription of the c-Myc and
Bcl-2 genes, which impairs apoptosis and facilitates the
proliferation of OSCC cells (8). In
addition, it has also been reported that overexpression of YAP is
related to the drug resistance and epithelial to mesenchymal
properties of OSCC (9,10). Another study also indicated that
cell growth and invasion could be inhibited by the suppression of
YAP in OSCC (11,12). Therefore, YAP may be a tumor
promoter and could be used as a potential anticancer drug target
for OSCC (13).
Metformin, which is extracted from French lilac and
easily produced, is an effective hypoglycemic drug with low
toxicity (14). It has been
revealed that metformin reduces the cancer risk and mortality of
diabetic patients compared with that of nonusers via meta-analysis
and epidemiology studies (15,16).
Therefore, as metformin is widely used, researchers have revealed
that metformin can effectively reduce the risk of cancer in people
with diabetes, and a dose-response relationship was proposed
(16). In addition, previous
studies have also revealed that metformin improves the survival of
patients who suffer from head and neck squamous cell carcinoma
(including oral cancer) (17,18).
Recently, it has been demonstrated that metformin suppresses the
growth of OSCC cells by promoting cell apoptosis and decreasing the
malignant potential of OSCC (19,20).
Metformin can also increase the sensitivity of OSCC cells to
cisplatin and disulfiram (21,22).
Thus, metformin may become a promising anticancer drug for OSCC.
However, it has been confirmed that metformin inhibits OSCC by
inhibiting the AMPK/mTOR (19),
NF-κB/HIF-1α (21) and LSF/Aurora-A
(23) pathways in cells. The
relationship between metformin and YAP in OSCC, according to our
knowledge, remains uncertain.
Metformin exerts its anticancer effect by reducing
mTOR and c-Myc protein expression in cancer cells (24–27),
and YAP expression is related to mTOR and c-Myc expression. YAP
translocates from the cytoplasm to the nucleus once the Hippo
signaling pathway is inactivated, which leads to increased
expression of mTOR in tuberous sclerosis complex cells and
colorectal cancer cells (28,29).
Hansen et al also explained that YAP promotes the expression
of mTOR protein via the high-affinity leucine transporter LAT1
(30). Moreover, it has been
verified that the protein expression of c-Myc in gastric cancer
cells and liver cancer cells is upregulated by the overexpression
of YAP (31–34). Our previous research also indicated
that YAP translocates to the nucleus, driving c-Myc gene
transcription in OSCC cells (8).
Thus, we hypothesized that metformin can downregulate the protein
expression of c-Myc and mTOR via a YAP-dependent mechanism in OSCC
cells.
In the present study, the relationship between
metformin and YAP in OSCC and the regulatory mechanism by which YAP
regulates the protein expression of c-Myc and mTOR in
metformin-treated cells were investigated. The present study thus
revealed a novel molecular mechanism of metformin in OSCC.
Materials and methods
Cell lines
Human tongue squamous cell carcinoma CAL27 and SCC25
cell lines were purchased from ATCC. CAL27 cells were cultured in
high-glucose culture medium (DMEM; HyClone; Cytiva) containing 10%
fetal bovine serum (FBS; Biological Industries) and 1%
penicillin-streptomycin (HyClone; Cytiva) . SCC25 cells were
cultured in high-glucose culture medium (DMEM F12; HyClone; Cytiva)
containing 10% FBS and 1% penicillin-streptomycin.
Reagents and antibodies
Metformin (item no. D9351) was purchased from
Solarbio Life Sciences, and Verteporfin (CL 318952) was purchased
from Sigma-Aldrich; Merck KGaA. Cell Counting Kit-8 (CCK-8) was
purchased from Dojindo Molecular Technologies, Inc. Both cell cycle
assay kit and Annexin V-PI cell apoptosis assay kit were purchased
from Hangzhou Multi Sciences (Lianke) Biotech Co., Ltd. APC-7AAD
cell apoptosis assay kit was purchased from Sungene Biotech Co.,
Ltd. Antibodies against CDK4 (D9G3E; product no. 12790), CDK6
(D4S8S; product no. 13331), p21 (12D1; product no. 2947), Bax
(D2E11; product no. 5023), Bcl-2 (D55G8; product no. 4223),
mammalian STE20-like kinase 1 (MST1) (D8B9Q; product no. 14946),
phosphorylated (p)-MST (Thr183; E7UD1; product no. 49332), large
tumor suppressor 1 (LATS1) (C66B5; product no. 3477), p-LATS1
(Thr1097; D57D3; product no. 8654), YAP (D8H1X; product no. 14074),
p-YAP (Ser127; D9W2I; product no. 13008), mTOR (7C10; product no.
2983), p-mTOR (Ser2448; D9C2; product no. 5536), c-Myc (D84C12;
product no. 5605), SAV1 (D6M6X; product no. 13301) and MOB1 (E1N9D;
product no. 13730) were obtained from Cell Signaling Technologies,
Inc.
Cell proliferation assay
A Cell Counting Kit-8 (CCK-8) assay was used to
detect changes in cell proliferation. First, the cells were seeded
at 2,500 cells/well in 96-well plates and cultured in a 37°C carbon
dioxide incubator for 24 h. Next, 0, 5, 15 and 30 mmol/l metformin
was added to 96-well plates, and the cells were cultured for 24, 48
and 72 h. Then, 10 µl of CCK-8 reagent was added to each well and
incubated for 2.5 h. Subsequently, a microplate reader was used to
detect the absorbance at 450 nm.
Cells treated without metformin or YAP
overexpression lentiviral vector served as the control group. The
experimental groups were as follows: Cells treated with metformin;
cells treated with YAP overexpression lentiviral vector; and cells
treated with metformin and YAP overexpression lentiviral vector. A
15-mmol/ml concentration of metformin was used in the relevant
experimental groups. The cell viability of each group was detected
by CCK-8 assay after 24 h of drug incubation, and a microplate
reader was used to detect the absorbance at 450 nm.
Cell cycle assay
Firstly, cells (2.5×106) were treated
with 0, 5, 15 and 30 mmol/l metformin for 24 h. Furthermore, cells
treated without metformin and overexpression YAP gene lentiviral
vector served as the control group; cells treated with metformin,
overexpression YAP gene lentiviral vector as well as metformin and
overexpression YAP gene lentiviral vector served as the
experimental groups. In addition, cells treated without metformin
and verteporfin served as the control group; cells treated with
verteporfin, as well as metformin and verteporfin served as the
experimental groups. A concentration of 15 mmol/l metformin or 1 µM
verteporfin was used in the experimental groups. After 24-h drug
incubation, 70% ethanol was used to fix cells for 12 h at 4°C,
after washing the cells with PBS. The cells were treated with cell
cycle detection kit (item no. CA1510; Solarbio Life Sciences)
according to the manufacturer's instructions and the proportion of
each cell cycle was analyzed using a flow cytometer (FACSCanto II;
BD Biosciences).
Cell apoptosis assay
Cells treated with 0 mmol/l metformin served as the
control group and cells respectively treated with 5, 15 and 30
mmol/l metformin served as the experimental groups. In addition,
cells treated without metformin and overexpression YAP gene
lentiviral vector served as the control group while cells treated
with metformin, overexpression YAP gene lentiviral or cells treated
with metformin combined with overexpression YAP gene lentiviral
vector served as the experimental groups. Similarly, cells treated
without metformin or verteporfin served as the control group while
cells treated with verteporfin, metformin or metformin combined
with verteporfin served as the experimental groups. The
concentration of metformin was 15 mmol/l and the concentration of
verteporfin was 1 µl in the experimental groups. After 24 h of drug
incubation, cells were stained using a green fluorescent-labeled
Annexin V-APC staining kit (Sungene Biotech Co., Ltd.) according to
the manufacturer's instructions. Cells (2×105) were
collected from each group of samples. Secondly, 500 µl of 1X
binding buffer and 8 µl of Annexin V-fluorescein APC were used to
suspend cells for 10 min. Finally, the cells were incubated with 5
µl of 7-AAD solution for 5 min. Similarly, Annexin V-fluorescein
isothiocyanate/propidium iodide (FITC/PI) (Hangzhou Multi Sciences
(Lianke) Biotech Co., Ltd.) was used to detect cells with no
staining. Finally, the proportion of apoptotic cells was calculated
by flow cytometry (FACSCanto II; BD Biosciences).
Western blotting
Cells (8×106) were cultured in a culture
plate and were collected, and the total protein in cells was
extracted with an ice-cold lysate containing a phosphatase
inhibitor and a protease inhibitor (Solarbio Life Sciences). The
protein concentration was determined by a bicinchoninic acid assay
(Boster Biological Technology, Co., Ltd.). The proteins (20 µg
total protein/lane) were separated using 10% SDS-PAGE.
Electrophoresis and transfer were performed according to the
standard protocol of western blotting. After blocking the PVDF
membrane for 1 h at 4°C with 5% non-fat milk in Tris-buffered
saline (TBST) containing 0.1% Tween-20, the membrane was probed
with a primary antibody (1:1,000) overnight at 4°C. Then the
membrane was incubated with HRP-conjugated secondary antibody
(1:20,000; product no. 7074; Cell Signaling Technology, Inc.) for
40 min at 25°C. Finally, protein bands were detected using the
Immobilon Western Chemiluminescent HRP Substrate Kit (Vazyme
Biotech Co., Ltd.). Densitometric analysis was performed using
ImageJ v1.52 (National Institutes of Health).
Construction of YAP-overexpressing
cell lines
A YAP overexpression lentiviral vector
(1×108 TU/ml) was constructed, and an empty lentiviral
vector was used as a control. Both vectors were prepared by the
Shanghai Genechem Co., Ltd. and transfected into the CAL27 and
SCC25 cell lines for 72 h at 27°C. Subsequently, 24 h later the
transfection efficiency was detected by western blotting and
RT-qPCR.
Reverse transcription quantitative
(RT-q)PCR analysis
Cells treated with lentiviral blank vector served as
the control group while cells treated with overexpressed YAP vector
served as the experimental group. Then, 72 h later, total RNA in
cells was extracted by TRIzol total RNA isolation (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. The purity of the obtained total RNA samples was
detected by NanoDrop spectrophotometer (Thermo Fisher Scientific,
Inc.). Total RNA (1 µg) was reverse transcribed into cDNA using
PrimeScript™ RT II reagent kit (Takara Bio, Inc.) on Biometra PCR
thermal cycler (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Then, 2 µl RNase-free water (Takara Bio, Inc.), 2 µl primers
(Sangon Biotech Co., Ltd.), 1 µl obtained cDNA samples and 5 µl
SYBR-Green fluorescent dye (Takara Bio, Inc.) were mixed into a
10-µl reaction system. The forward primer of YAP was
5′-TCTTACACCGTGCTGCCATT-3′ and the reverse primer was
5′-AGCACCTGTCCAGGTATCAC-3′. The forward primer of GAPDH was
5′-GCACCGTCAAGGCTGAGAAC-3′ and the reverse primer was
5′-TGGTGAAGACGCCAGTGGA-3′. Finally, the gene expression was
detected by Light Cycler Roche 480 RT-qPCR instrument (Roche
Diagnostics). The results of fold-changes in mRNA levels were
calculated by using a 2−ΔΔCq method (35).
Statistical analysis
One-way analysis of variance (ANOVA) followed by
Tukey's post hoc test were used to analyze the differences between
several research groups, and the difference between two groups was
analyzed by two-tailed t-test (P<0.05 and P<0.01 were
considered to indicate statistically significant differences). All
the statistical results were analyzed by SPSS software 20.0 (IBM
Corp.).
Results
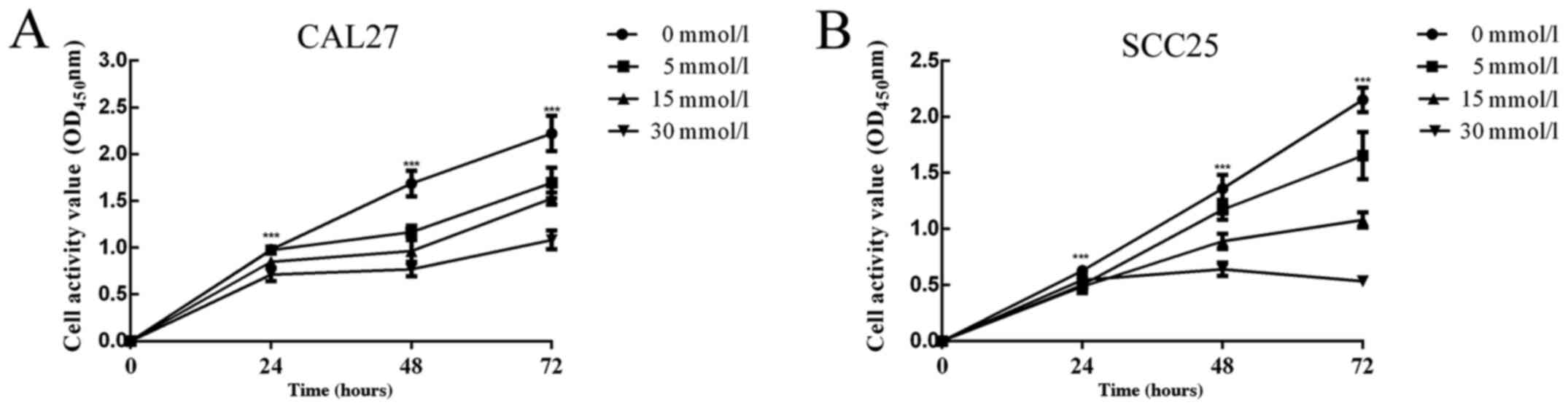
Metformin inhibits cell proliferation
and promotes cell apoptosis in OSCC
To illustrate the effect of metformin on the growth
of CAL27 and SCC25 cells, cells were first treated with 0, 5, 15
and 30 mmol/l metformin, and the changes in cell proliferation were
assessed at 24, 48 and 72 h by CCK-8 assay. It was evident that
cell proliferation decreased with increasing metformin
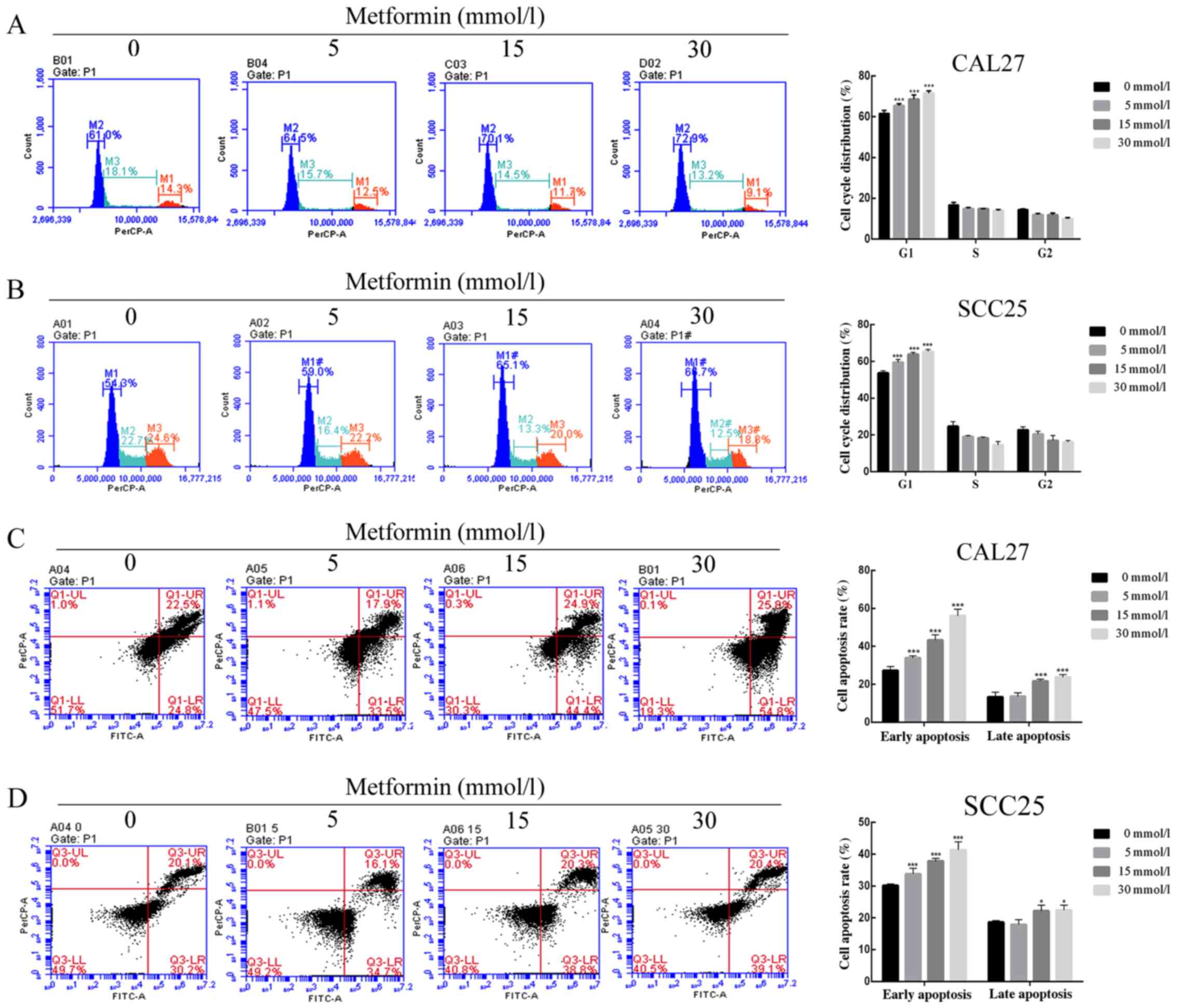
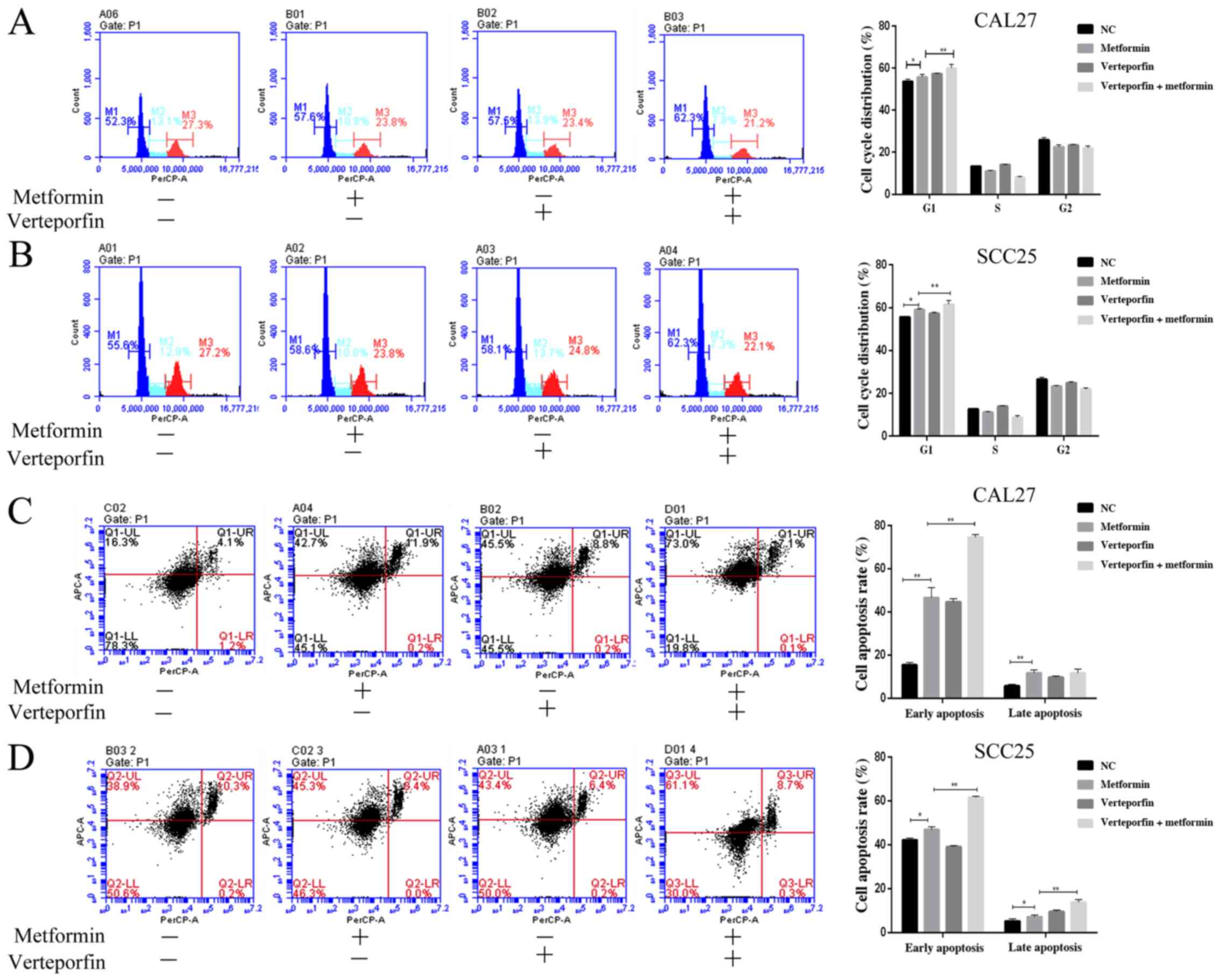
concentration and increasing dosing time (Fig. 1). The results of flow cytometric
assays revealed that the proportion of cells arrested in the G0/G1
phase was increased, while the proportion of cells in the S phase
and G2 phase were reduced in a dose-dependent manner (Fig. 2A and B). Then, FITC/PI staining and
flow cytometry were used to detect the cell apoptosis induced by
metformin and it was revealed that the early apoptosis rate and
late apoptosis rate were significantly increased with metformin
treatment compared with the control treatment (Fig. 2C and D). In addition, biomarkers of
proliferation and apoptosis were detected by western blotting. The
protein expression levels of CDK4 and CDK6, which are involved in
the G0/G1 phase (36), were
markedly decreased in metformin-treated cells compared to control
cells. p21 has the ability to inhibit the transition from the G1 to
S phase in the cell cycle (37),
and its expression was increased accordingly (Fig. 3A). The decreased protein expression
of Bcl-2 and the increased expression of the proapoptotic protein
Bax in the metformin group vs. the control group confirmed that
metformin inhibited cell growth by promoting apoptosis in the CAL27
and SCC25 cell lines (Fig. 3A). The
results revealed that metformin arrested the cell cycle in the G1/S
phase and promoted cell apoptosis in vitro.
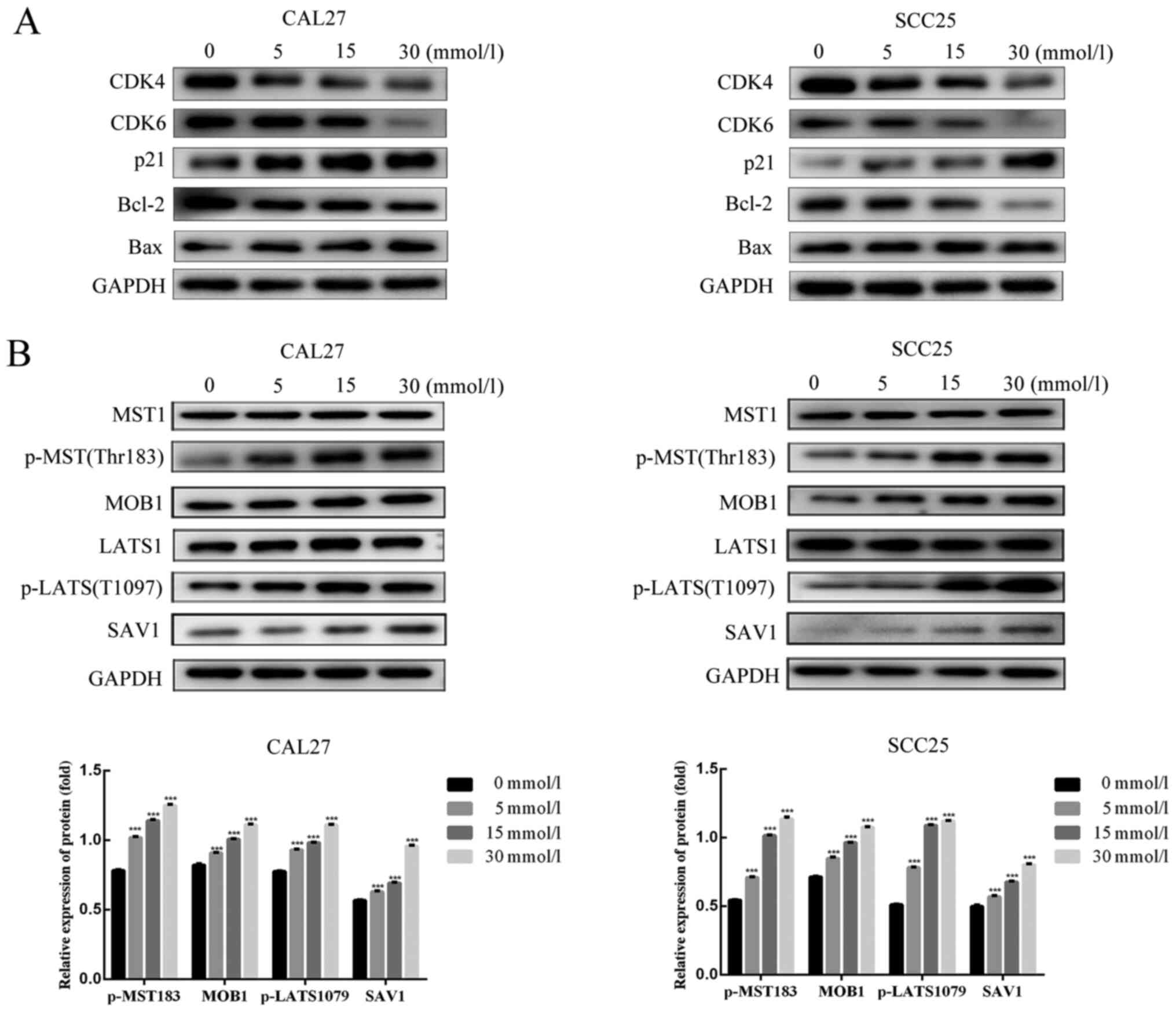 | Figure 3.Effect of metformin on protein
expression in OSCC cells. CAL27 and SCC25 cells were treated with
0, 5, 15 and 30 mmol/l metformin for 24 h. Total protein was
extracted from cells. Then, proteins related to the cell cycle and
cell apoptosis were detected by western blotting. The experiment
was repeated three times. (A) CDK4, CDK6, p21, Bcl-2 and Bax
protein expression in CAL27 and SCC25 cells following 24 h of
metformin treatment. (B) MST1, p-MST1, MOB1, LATS1,
p-LATS1SAV1protein expression in CAL27 and SCC25 cells following 24
h of metformin treatment. GAPDH served as a loading control. Data
are presented as the mean SEM. ***P<0.001 denote significance
for comparisons between the untreated control group and the
treatment group. OSCC, oral squamous cell carcinoma; MST1,
mammalian STE20-like kinase 1; p-, phosphorylated; LATS1, large
tumor suppressor 1; YAP, Yes-associated protein. |
Metformin stimulates the Hippo
signaling pathway in OSCC cells
YAP is one of the transcription coactivators
inhibited by the Hippo pathway and is related to OSCC cell growth
(6). The signaling molecules
related to the Hippo pathway rely on a phosphorylation cascade.
MST1 can be phosphorylated at Thr183 as a result of activation of
the Hippo pathway. Then, phosphorylated MST1 can bind with SAV1 to
phosphorylate LATS1 at T1097, which can bind MOB1. Next, upstream
activation of the Hippo pathway can result in the phosphorylation
of Yap at S127. Subsequently, YAP is susceptible to proteasomal
degradation (38). In the present
results, metformin increased the phosphorylation of MST1 (Thr183)
and LATS1 (T1097), and the expression of total protein was not
markedly altered as the dose of metformin increased. The protein
expression of SAV1 and MOB1 also increased with metformin treatment
(Fig. 3B).
Metformin inhibits OSCC cell growth by
decreasing YAP
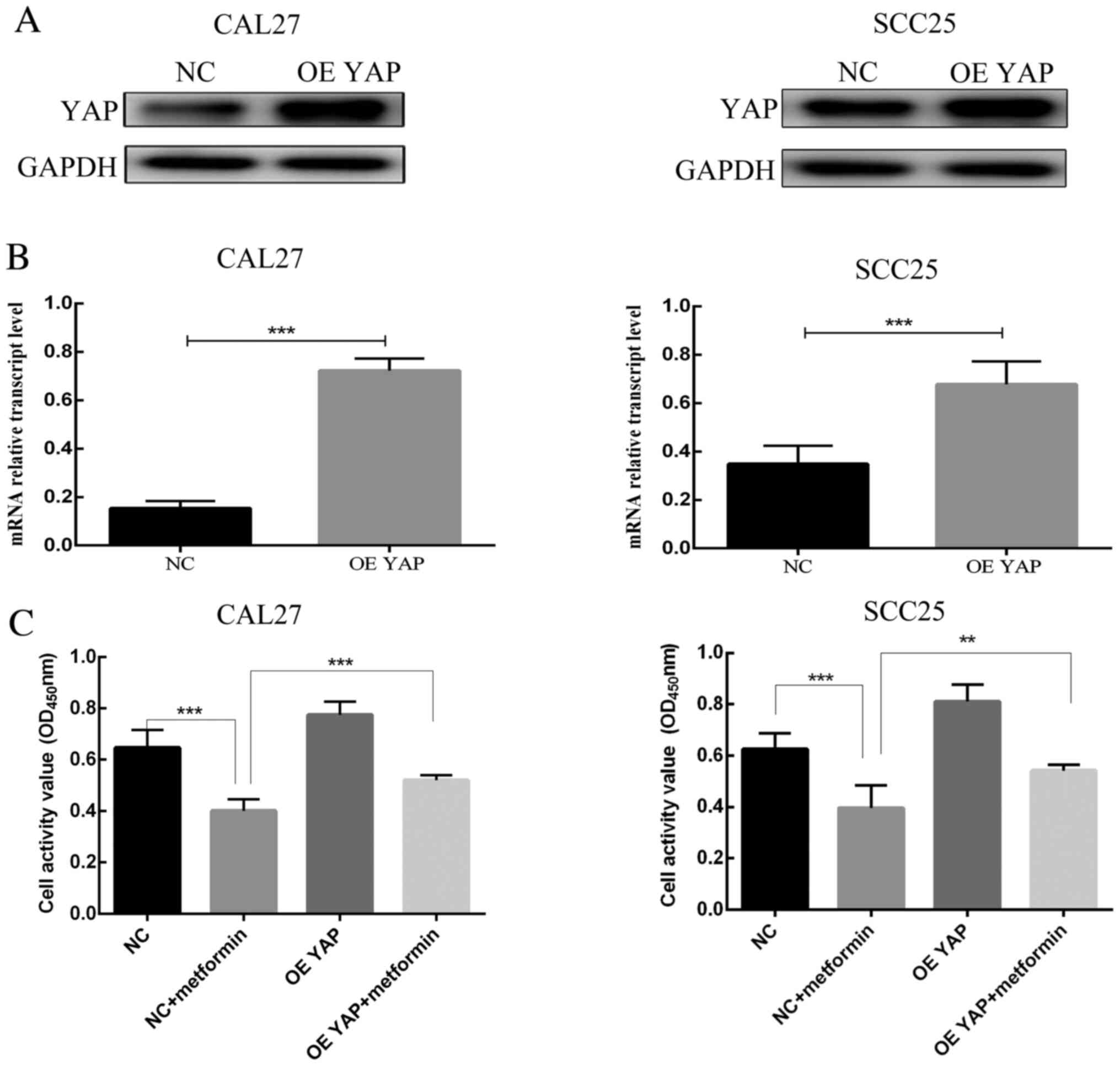
The YAP expression level was enhanced in CAL27 and
SCC25 cells by using YAP overexpression lentiviral particles. YAP
protein expression was confirmed to be higher in the overexpression
group than in the control group (Fig.
4A). The mRNA expression of YAP in the YAP overexpression group
was also higher than that in the control group (Fig. 4B). In contrast to transfection with
empty lentiviral vector, transfection with YAP overexpression
vector significantly weakened the effects of metformin on OSCC
cells. The optical density (OD) value at 450 nm in the CCK-8 assay
of YAP-overexpressing cells was higher than that of empty
lentivirus-transfected cells after metformin treatment for 24 h
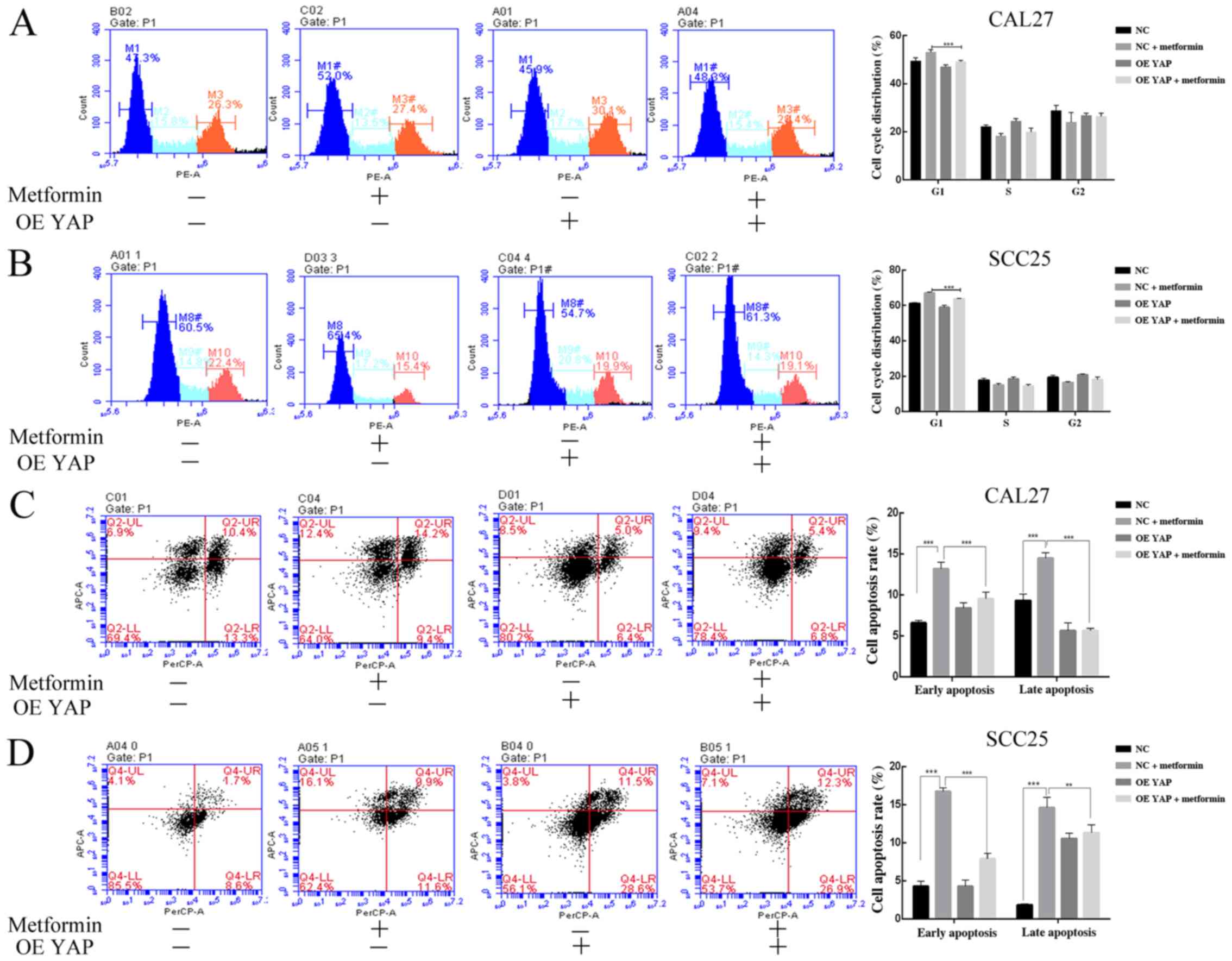
(Fig. 4C). The proportion of cells
arrested in G1 phase of the cell cycle in the YAP overexpression
group was lower than that in the empty lentivirus group after
metformin treatment for 24 h (Fig. 5A
and B). The cell apoptosis rate in the YAP overexpression group
was lower than that in the empty lentivirus group after metformin
treatment for 24 h (Fig. 5C and D).
To further identify the role of YAP in metformin treatment,
metformin was combined with the YAP inhibitor verteporfin. The
proportion of cells at the G1 phase (Fig. 6A and B) and the proportion of
apoptotic cells in the metformin and verteporfin treated group were
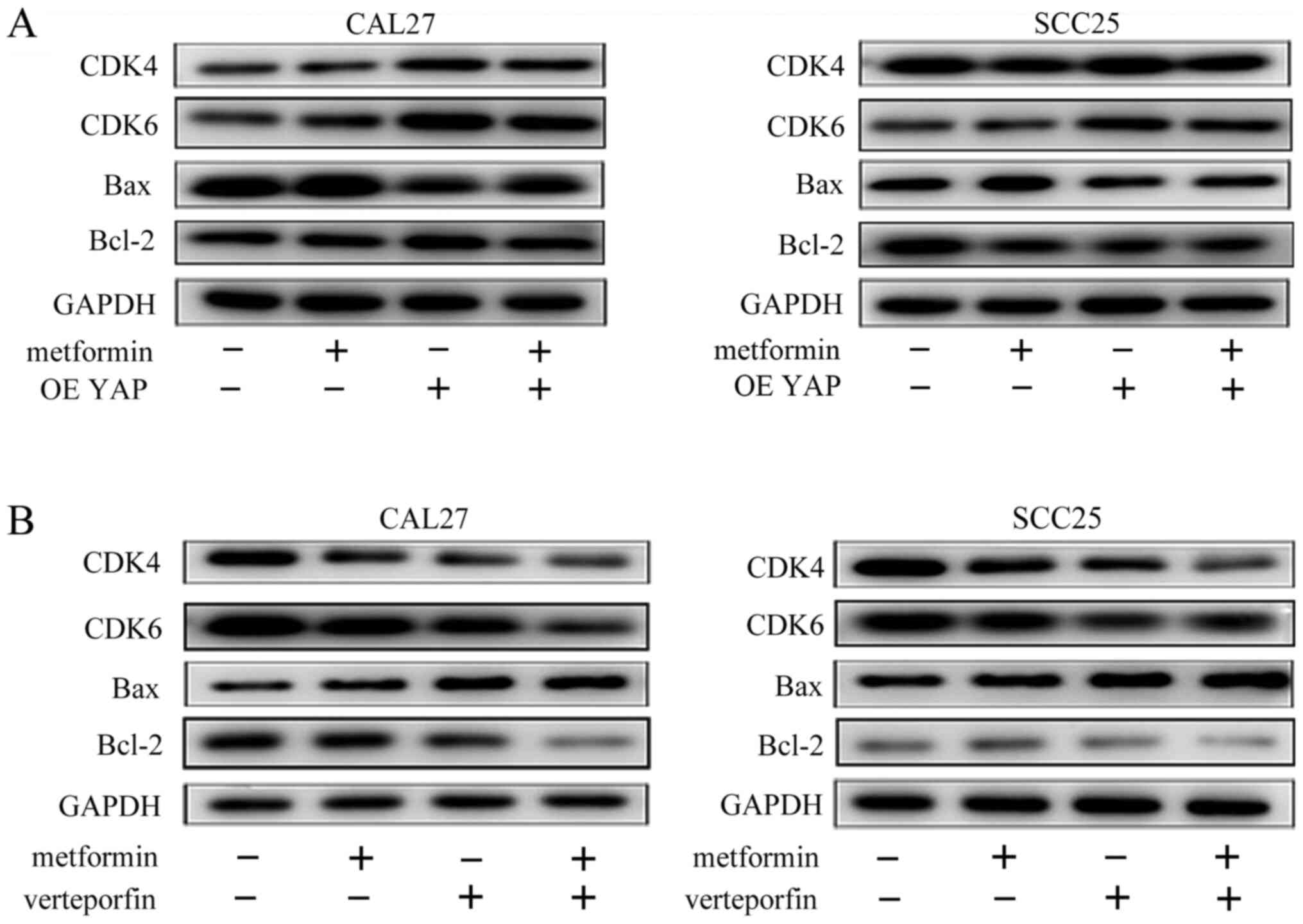
higher than those in the metformin-treated group (Fig. 6C and D). The YAP overexpression
group exhibited higher expression of CDK4, CDK6 and Bcl-2 and lower
expression of Bax than the empty lentivirus group after metformin
treatment for 24 h (Fig. 7A).
However, the CDK4, CDK6, and Bcl-2 protein expression was lower and
the Bax expression was higher in the verteporfin and metformin
combination treatment group than in the metformin treatment group
(Fig. 7B). These results clearly
indicated that metformin had an inhibitory effect on the
proliferation of OSCC cells by decreasing YAP protein.
Metformin decreases mTOR and c-Myc
through the downregulation of YAP
Other studies have revealed that YAP promotes the
expression of mTOR in hepatocellular cells and colorectal cancer
cells (39,40). Our previous study also revealed that
YAP facilitated the protein expression of c-Myc (8). However, few studies have reported that
metformin inhibits the expression of mTOR and c-Myc through a
YAP-dependent mechanism in OSCC cells (24–27).
To further illustrate the molecular relationship between metformin
and YAP in OSCC cells, western blotting was used to detect changes
in mTOR and c-Myc expression in OSCC cells. As anticipated, a
gradual reduction in mTOR expression, mTOR phosphorylation at
Ser2448 and c-Myc expression was observed as the metformin
concentration increased (Fig. 8A).
Overexpression of YAP facilitated mTOR and c-Myc protein expression
in CAL27 and SCC25 cells treated with metformin (Fig. 8B). The metformin and verteporfin
combination group exhibited a decreased expression level of mTOR
and c-Myc protein than the metformin treatment groups (Fig. 8C). These results indicated that YAP
inhibition was associated with the metformin-induced decrease in
mTOR and c-Myc expression in OSCC cells.
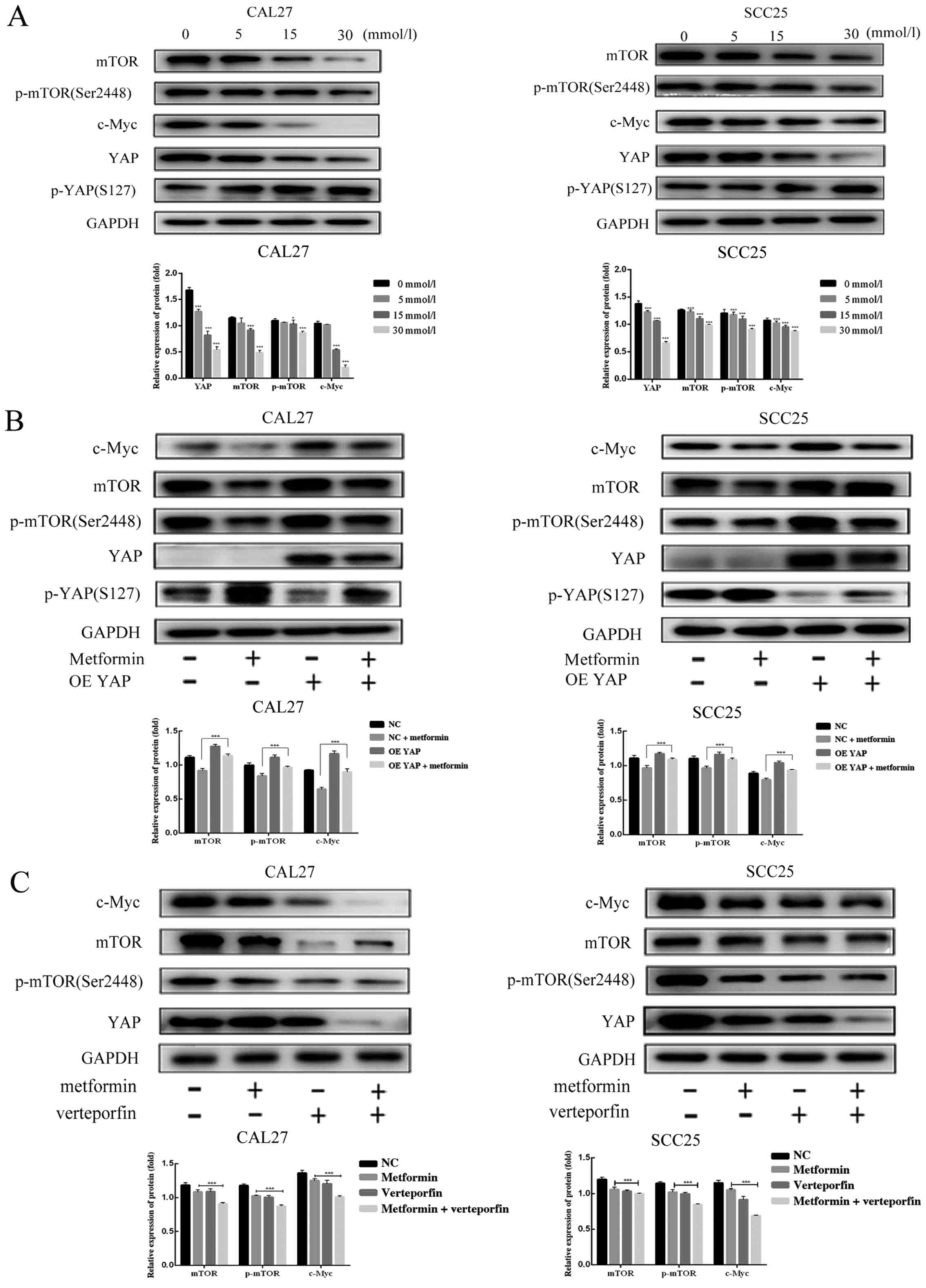 | Figure 8.Effect of metformin on mTOR, p-mTOR
and c-Myc protein expression in OSCC cells. CAL27 and SCC25 cells
were treated with 0, 5, 15 and 30 mmol/l metformin for 24 h. To
verify the effect of YAP protein on the protein expression of mTOR,
p-mTOR and c-Myc, the protein expression of YAP was changed by
lentivirus infection or verteporfin treatment. (A) The protein
expression of mTOR, p-mTOR and c-Myc in CAL27 and SCC25 cells upon
treatment with different concentrations of metformin. (B) The
protein expression of mTOR, p-mTOR and c-Myc in the control group,
metformin group, YAP overexpression group and the combined
treatment group. (C) The protein expression of mTOR, p-mTOR and
c-Myc in the control group, metformin group, verteporfin group and
the combined treatment group. GAPDH served as the loading control.
The data are presented as the mean SEM. *P<0.05 and
***P<0.001 denote significance for comparisons between the
metformin group and the combined treatment group. YAP,
Yes-associated protein; mTOR, mammalian target of rapamycin; p-,
phosphorylated; OSCC, oral squamous cell carcinoma; OE,
overexpression; NC, negative control |
Discussion
Metformin, discovered from French lilac, has been
widely studied for its role in cancer treatment since it has an
antitumor ability with characteristics of low toxicity and low cost
production (41). It has been
revealed that metformin-treated head and neck squamous cell
carcinoma patients had a higher survival rate than those not
treated with metformin, according to a meta-analysis (17,18).
Previous studies have indicated that metformin promotes the toxic
effects of disulfiram, 5-fluorouracil and cisplatin on OSCC cells
since metformin enhances the inhibitory effects of these
traditional chemotherapeutics, even at low concentrations (21,22,42,43).
In addition, it has also been demonstrated that metformin
suppresses cell proliferation and increases cell apoptosis in OSCC
in vivo and in vitro (19). Although some studies have elucidated
the effects of metformin on OSCC, the molecular mechanism of
metformin on OSCC cells remains undefined.
Prolonged proliferation and insensitivity to
apoptosis are two main types of biological characteristics
typically featured in human tumors (44). Therefore, controlling cancer cell
proliferation and apoptosis has a certain significance for the
clinical treatment of cancer. YAP, as a vital carcinogenic factor,
is important in OSCC growth, and high expression of YAP protein may
accelerate the progression of OSCC (6). YAP was demonstrated to enhance the
proliferation rate of cells and reduce apoptosis by promoting c-Myc
and Bcl-2 protein expression in OSCC (8). Moreover, cell growth and invasion were
inhibited by the suppression of YAP in OSCC (11). Inhibition of YAP protein also
decreased the drug resistance of cisplatin in OSCC cells (9). Thus, to determine whether metformin
has an inhibitory effect on OSCC, the YAP protein was
inhibited.
The protein binding between YAP and TEAD4 has been
demonstrated to accelerate the cell proliferation rate by markedly
promoting the transition of the cell cycle from the G1 to S phase
and reducing cell apoptosis (8). In
addition, the protein expression of CDK4/6 and p21 is involved in
the transition of the cell cycle from the G1 to S phase. CDK4, CDK6
and other cyclin-dependent kinases can accelerate the transition of
the cell cycle from the G1 to the S phase (36). p21 is an inhibitor of
cyclin-dependent kinases, and it has a negative regulatory effect
on the cell cycle transition from the G1 to the S phase (37). The Bcl-2 protein family can control
the apoptosis process of cells by influencing the outer
mitochondrial membrane potential. Bcl-2 protein and Bax protein
belong to the Bcl-2 protein family. The former blocks the process
of cell apoptosis, while the latter stimulates the processes of
cell apoptosis (45). As
anticipated, in the present study, metformin inhibited the
transition of the cell cycle from the G1 to S phase and increased
cell apoptosis in CAL27 and SCC25 cells. The protein expression of
YAP in CAL27 and SCC25 cells was also decreased with metformin
treatment compared to control treatment. The protein expression of
CDK4, CDK6, p21, Bcl-2 and Bax also paralleled the phenotypic
changes upon metformin exposure.
Next, YAP overexpression lentiviral particles or
empty lentiviral vectors were transfected into CAL27 and SCC25
cells. The cells were divided into a YAP overexpression group and a
negative control group. The protein and mRNA expression levels of
YAP in the YAP overexpression group were higher than those of the
empty vector group. In fact, metformin inhibited the transition of
the cell cycle from the G1 to S phase and increased cell apoptosis
in the control group. However, overexpression of YAP reversed the
inhibitory effects of metformin on the cell cycle and attenuated
metformin-induced cell apoptosis. The protein expression of CDK4,
CDK6, p21, Bcl-2 and Bax also paralleled the phenotypic changes in
the YAP overexpression group.
To further confirm that metformin affected the cell
cycle and apoptosis by decreasing YAP protein, cells were treated
with both metformin and verteporfin, which is an inhibitor of YAP
protein. Verteporfin is a photosensitizer that can be used for
photodynamic therapy to treat macular degeneration with limited
toxicity (46). Recent studies have
illustrated that verteporfin effectively inhibits the development
of cancer by decreasing the expression of YAP protein in cancer
cells (47,48). For example, verteporfin inhibits the
translocation of YAP protein into the nucleus by inhibiting the
binding of YAP and TEAD (49) or
promotes the expression of 14-3-3 protein so that YAP remains in
the cytoplasm (50). Our previous
experiments also verified that the expression of YAP protein in
OSCC cells decreased with increasing verteporfin drug concentration
(8). Therefore, verteporfin was
combined with metformin. As anticipated, the effects of the
combined use of metformin and verteporfin on the cell cycle and
apoptosis were greater than those of a single medication. The
protein expression of CDK4, CDK6, Bcl-2 and Bax also paralleled the
phenotypic changes in the metformin and verteporfin combination
group. These data clarified that metformin has an inhibitory effect
on OSCC by inhibiting the YAP protein.
YAP is a main downstream factor of the Hippo
signaling pathway and can be inhibited by the Hippo signaling
pathway (6). MST1, LATS1, SAV1 and
MOB1 constitute the upstream regulatory signal transduction of the
Hippo signaling pathway. It is known that MST1 can be
phosphorylated at Thr183 as a result of activation of the Hippo
pathway. Then, phosphorylated MST1 can bind with SAV1 to
phosphorylate LATS1 at T1097, which can bind MOB1. Subsequently,
the upstream Hippo pathway is activated, which could result in the
phosphorylation of YAP at S127. Phosphorylated YAP is susceptible
to proteasomal degradation (51).
Phosphorylation of S127 results in YAP sequestration in the
cytoplasm via interaction with 14-3-3 (11) or SCFβ-TrCP E3 ubiquitin ligase
(52). The present results
demonstrated that metformin increased the phosphorylation of MST1
(Thr183) and LATS1 (T1097), with virtually no change in the
expression level of total protein as metformin concentration
increased. The protein expression of SAV1 and MOB1 also increased
with metformin treatment. A change in YAP phosphorylation (S127)
and a progressive decrease in YAP expression followed the
activation of the upstream signaling factor in the Hippo pathway.
These results indicated that metformin may reduce the protein
expression of YAP by activating the Hippo pathway.
The mTOR signaling pathway transmits information
concerning synthesis or catabolism in cells and affects the
metabolism of proteins, lipids and nucleotides in cells, thereby
providing sufficient substances for cancer growth (53). Observation of patients with tongue
squamous cell carcinoma revealed high expression levels of mTOR and
p-mTOR proteins in patient tissue sections (54). It has been demonstrated that
circular RNA hsa-circ-0007059 or activated C kinase 1 receptor
promotes the proliferation of OSCC cells by increasing the
expression of mTOR protein (55,56).
YAP serves as a vital association between the Hippo pathway and the
PI(3)K-mTOR pathway. It has been confirmed that SAV1 inhibits the
protein expression of mTOR by inactivating the YAP protein in
colorectal cancer (29). It has
also been illustrated that mTOR blocks the inhibitory effect of
AMOTL2 on YAP to promote cell growth and invasion in glioblastoma
(57). In addition, YAP has been
revealed to increase the expression level of miR-29, which
indirectly increases the protein expression of mTOR in cells
(40). In the present study,
metformin decreased the protein expression of mTOR in a
dose-dependent manner. YAP overexpression inhibited the ability of
metformin to decrease the protein expression of mTOR. In contrast,
the combination of metformin with verteporfin enhanced the ability
of metformin to decrease the protein expression of mTOR.
The Myc genes are a family of
proto-oncogenes. MYC family proteins are involved in regulating the
proliferation, differentiation and apoptosis of cancer cells and
are closely related to cell division activities (58). A study of OSCC patients revealed
that the level of c-Myc protein expression was closely related to
disease severity and survival rate (59). Our previous study revealed that YAP
is transported into the nucleus, where it drives the transcription
of the c-Myc gene to accelerate OSCC cell growth (8). Other studies also demonstrated by RNA
sequencing that c-Myc serves as a definitive target of YAP and
mediates the ability of YAP to promote cancer growth (31,33).
Thus, it was investigated whether metformin has an inhibitory
effect on c-Myc in OSCC cells by inhibiting the YAP protein. In the
present study, metformin decreased the protein expression of c-Myc
in a dose-dependent manner. YAP overexpression inhibited the
ability of metformin to decrease the protein expression of c-Myc.
Conversely, the combination of metformin with verteporfin enhanced
the ability of metformin to decrease the protein expression of
c-Myc.
In conclusion, the present experiments demonstrated
that metformin prevented OSCC by reducing the expression of YAP
protein. Furthermore, it was revealed that YAP-mediated metformin
also had regulatory effects on c-Myc and mTOR proteins. This
provides a new reference for the anticancer mechanism of metformin.
However, this study still has some limitations. For example, this
molecular mechanism has not been verified by in vivo
experiments in the present study. If conditions permit, further
experiments will be designed to explore and verify this related
hypothesis in the future. The present study thus revealed that YAP
may be recognized as a potential target for metformin treatment in
OSCC.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Shandong Provincial Natural Science Foundation (grant no.
ZR2018MH018), the Young Scholars Program of Shandong University
(grant no. 2015WLJH53), the China Postdoctoral Science Foundation
(grant no. 2017M610432), the Shandong Postdoctoral Innovation
Project (grant no. 201703071) and the Construction Engineering
Special Fund of Taishan Scholars (grant no. ts201511106).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YWa, YWe, WG and YZ designed and executed the study.
YZ, XF, HT and XF analyzed the data. WY wrote the study. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Feller L and Lemmer J: Oral squamous cell
carcinoma: Epidemiology, clinical presentation and treatment. J
Cancer Ther. 3:263–268. 2012. View Article : Google Scholar
|
|
3
|
Rodrigues MFSD, Miguita L, De Andrade NP,
Heguedusch D, Rodini CO, Moyses RA, Toporcov TN, Gama RR, Tajara EE
and Nunes FD: GLI3 knockdown decreases stemness, cell proliferation
and invasion in oral squamous cell carcinoma. Int J Oncol.
53:2458–2472. 2018.PubMed/NCBI
|
|
4
|
Zhang L, Meng X, Zhu XW, Yang DC, Chen R,
Jiang Y and Xu T: Long non-coding RNAs in oral squamous cell
carcinoma: Biologic function, mechanisms and clinical implications.
Mol Cancer. 18:1022019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sakata J, Hirosue A, Yoshida R, Kawahara
K, Matsuoka Y, Yamamoto T, Nakamoto M, Hirayama M, Takahashi N,
Nakamura T, et al: HMGA2 contributes to distant metastasis and poor
prognosis by promoting angiogenesis in oral squamous cell
carcinoma. Int J Mol Sci. 20:202019. View Article : Google Scholar
|
|
6
|
Hiemer SE, Zhang L, Kartha VK, Packer TS,
Almershed M, Noonan V, Kukuruzinska M, Bais MV, Monti S and Varelas
X: A YAP/TAZ-regulated molecular signature is associated with oral
squamous cell carcinoma. Mol Cancer Res. 13:957–968. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li SY, Hu JA and Wang HM: Expression of
Yes-associated protein 1 gene and protein in oral squamous cell
carcinoma. Chin Med J (Engl). 126:655–658. 2013.PubMed/NCBI
|
|
8
|
Chen X, Gu W, Wang Q, Fu X, Wang Y, Xu X
and Wen Y: C-MYC and BCL-2 mediate YAP-regulated tumorigenesis in
OSCC. Oncotarget. 9:668–679. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoshikawa K, Noguchi K, Nakano Y, Yamamura
M, Takaoka K, Hashimoto-Tamaoki T and Kishimoto H: The Hippo
pathway transcriptional co-activator, YAP, confers resistance to
cisplatin in human oral squamous cell carcinoma. Int J Oncol.
46:2364–2370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zeng G, Xun W, Wei K, Yang Y and Shen H:
MicroRNA-27a-3p regulates epithelial to mesenchymal transition via
targeting YAP1 in oral squamous cell carcinoma cells. Oncol Rep.
36:1475–1482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dong C, Wei KJ, Zhang WB, Sun H, Pan HY
and Zhang L: LATS2 induced by TNF-alpha and inhibited cell
proliferation and invasion by phosphorylating YAP in oral squamous
cell carcinoma. J Oral Pathol Med. 44:475–481. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fan H, Tian H, Cheng X, Chen Y, Liang S,
Zhang Z, Liao Y and Xu P: Aberrant Kank1 expression regulates YAP
to promote apoptosis and inhibit proliferation in OSCC. J Cell
Physiol. 235:1850–1865. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nakatani K, Maehama T, Nishio M, Goto H,
Kato W, Omori H, Miyachi Y, Togashi H, Shimono Y and Suzuki A:
Targeting the Hippo signalling pathway for cancer treatment. J
Biochem. 161:237–244. 2017.PubMed/NCBI
|
|
14
|
Witters LA: The blooming of the French
lilac. J Clin Invest. 108:1105–1107. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Decensi A, Puntoni M, Goodwin P, Cazzaniga
M, Gennari A, Bonanni B and Gandini S: Metformin and cancer risk in
diabetic patients: A systematic review and meta-analysis. Cancer
Prev Res (Phila). 3:1451–1461. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Landman GW, Kleefstra N, van Hateren KJ,
Groenier KH, Gans RO and Bilo HJ: Metformin associated with lower
cancer mortality in type 2 diabetes: ZODIAC-16. Diabetes Care.
33:322–326. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rêgo DF, Pavan LM, Elias ST, De Luca Canto
G and Guerra EN: Effects of metformin on head and neck cancer: A
systematic review. Oral Oncol. 51:416–422. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang PH, Yeh KY, Wang CH, Chen EY, Yang
SW, Chou WC and Hsieh JC: Impact of metformin on patients with
advanced head and neck cancer undergoing concurrent
chemoradiotherapy. Head Neck. 39:1573–1577. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luo Q, Hu D, Hu S, Yan M, Sun Z and Chen
F: In vitro and in vivo anti-tumor effect of metformin as a novel
therapeutic agent in human oral squamous cell carcinoma. BMC
Cancer. 12:5172012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen CH, Tsai HT, Chuang HC, Shiu LY, Su
LJ, Chiu TJ, Luo SD, Fang FM, Huang CC and Chien CY: Metformin
disrupts malignant behavior of oral squamous cell carcinoma via a
novel signaling involving Late SV40 factor/Aurora-A. Sci Rep.
7:13582017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qi X, Xu W, Xie J, Wang Y, Han S, Wei Z,
Ni Y, Dong Y and Han W: Metformin sensitizes the response of oral
squamous cell carcinoma to cisplatin treatment through inhibition
of NF-κB/HIF-1α signal axis. Sci Rep. 6:357882016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jivan R, Peres J, Damelin LH, Wadee R,
Veale RB, Prince S and Mavri-Damelin D: Disulfiram with or without
metformin inhibits oesophageal squamous cell carcinoma in vivo.
Cancer Lett. 417:1–10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harada K, Ferdous T, Harada T and Ueyama
Y: Metformin in combination with 5-fluorouracil suppresses tumor
growth by inhibiting the Warburg effect in human oral squamous cell
carcinoma. Int J Oncol. 49:276–284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alalem M, Ray A and Ray BK: Metformin
induces degradation of mTOR protein in breast cancer cells. Cancer
Med. 5:3194–3204. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ben Sahra I, Regazzetti C, Robert G,
Laurent K, Le Marchand-Brustel Y, Auberger P, Tanti JF,
Giorgetti-Peraldi S and Bost F: Metformin, independent of AMPK,
induces mTOR inhibition and cell-cycle arrest through REDD1. Cancer
Res. 71:4366–4372. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Akinyeke T, Matsumura S, Wang X, Wu Y,
Schalfer ED, Saxena A, Yan W, Logan SK and Li X: Metformin targets
c-MYC oncogene to prevent prostate cancer. Carcinogenesis.
34:2823–2832. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Javeshghani S, Zakikhani M, Austin S,
Bazile M, Blouin MJ, Topisirovic I, St-Pierre J and Pollak MN:
Carbon source and myc expression influence the antiproliferative
actions of metformin. Cancer Res. 72:6257–6267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liang N and Pende M: YAP enters the mTOR
pathway to promote tuberous sclerosis complex. Mol Cell Oncol.
2:e9981002015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang J, Chang W, Fu Y, Gao Y, Zhao C,
Zhang X and Zhang S: SAV1 represses the development of human
colorectal cancer by regulating the Akt-mTOR pathway in a
YAP-dependent manner. Cell Prolif. 50:502017. View Article : Google Scholar
|
|
30
|
Hansen CG, Ng YL, Lam WL, Plouffe SW and
Guan KL: The Hippo pathway effectors YAP and TAZ promote cell
growth by modulating amino acid signaling to mTORC1. Cell Res.
25:1299–1313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Choi W, Kim J, Park J, Lee DH, Hwang D,
Kim JH, Ashktorab H, Smoot D, Kim SY, Choi C, et al: YAP/TAZ
initiates gastric tumorigenesis via upregulation of MYC. Cancer
Res. 78:3306–3320. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cai J, Song X, Wang W, Watnick T, Pei Y,
Qian F and Pan D: A RhoA-YAP-c-Myc signaling axis promotes the
development of polycystic kidney disease. Genes Dev. 32:781–793.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xiao W, Wang J, Ou C, Zhang Y, Ma L, Weng
W, Pan Q and Sun F: Mutual interaction between YAP and c-Myc is
critical for carcinogenesis in liver cancer. Biochem Biophys Res
Commun. 439:167–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Turato C, Cannito S, Simonato D, Villano
G, Morello E, Terrin L, Quarta S, Biasiolo A, Ruvoletto M, Martini
A, et al: SerpinB3 and Yap Interplay Increases Myc Oncogenic
Activity. Sci Rep. 5:177012015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lulla AR, Slifker MJ, Zhou Y, Lev A,
Einarson MB, Dicker DT and El-Deiry WS: miR-6883 family miRNAs
target CDK4/6 to induce G1 phase cell-cycle arrest in colon cancer
cells. Cancer Res. 77:6902–6913. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gulappa T, Reddy RS, Suman S, Nyakeriga AM
and Damodaran C: Molecular interplay between cdk4 and p21 dictates
G0/G1 cell cycle arrest in prostate cancer cells. Cancer Lett.
337:177–183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ren F, Zhang L and Jiang J: Hippo
signaling regulates Yorkie nuclear localization and activity
through 14-3-3 dependent and independent mechanisms. Dev Biol.
337:303–312. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park YY, Sohn BH, Johnson RL, Kang MH, Kim
SB, Shim JJ, Mangala LS, Kim JH, Yoo JE, Rodriguez-Aguayo C, et al:
Yes-associated protein 1 and transcriptional coactivator with
PDZ-binding motif activate the mammalian target of rapamycin
complex 1 pathway by regulating amino acid transporters in
hepatocellular carcinoma. Hepatology. 63:159–172. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tumaneng K, Schlegelmilch K, Russell RC,
Yimlamai D, Basnet H, Mahadevan N, Fitamant J, Bardeesy N, Camargo
FD and Guan KL: YAP mediates crosstalk between the Hippo and
PI(3)K-TOR pathways by suppressing PTEN via miR-29. Nat Cell Biol.
14:1322–1329. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Safe S, Nair V and Karki K:
Metformin-induced anticancer activities: Recent insights. Biol
Chem. 399:321–335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mynhardt C, Damelin LH, Jivan R, Peres J,
Prince S, Veale RB and Mavri-Damelin D: Metformin-induced
alterations in nucleotide metabolism cause 5-fluorouracil
resistance but gemcitabine susceptibility in oesophageal squamous
cell carcinoma. J Cell Biochem. 119:1193–1203. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Damelin LH, Jivan R, Veale RB, Rousseau AL
and Mavri-Damelin D: Metformin induces an intracellular reductive
state that protects oesophageal squamous cell carcinoma cells
against cisplatin but not copper-bis(thiosemicarbazones). BMC
Cancer. 14:3142014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Czabotar PE, Lessene G, Strasser A and
Adams JM: Control of apoptosis by the BCL-2 protein family:
Implications for physiology and therapy. Nat Rev Mol Cell Biol.
15:49–63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Henney JE: From the Food and Drug
Administration. JAMA. 283:27792000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang B, Shao W, Shi Y, Liao J, Chen X and
Wang C: Verteporfin induced SUMOylation of YAP1 in endometrial
cancer. Am J Cancer Res. 10:1207–1217. 2020.PubMed/NCBI
|
|
48
|
Lui JW, Xiao S, Ogomori K, Hammarstedt JE,
Little EC and Lang D: The efficiency of verteporfin as a
therapeutic option in pre-clinical models of melanoma. J Cancer.
10:1–10. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wei H, Wang F, Wang Y, Li T, Xiu P, Zhong
J, Sun X and Li J: Verteporfin suppresses cell survival,
angiogenesis and vasculogenic mimicry of pancreatic ductal
adenocarcinoma via disrupting the YAP-TEAD complex. Cancer Sci.
108:478–487. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang C, Zhu X, Feng W, Yu Y, Jeong K, Guo
W, Lu Y and Mills GB: Verteporfin inhibits YAP function through
up-regulating 14-3-3σ sequestering YAP in the cytoplasm. Am J
Cancer Res. 6:27–37. 2015.PubMed/NCBI
|
|
51
|
Meng Z, Moroishi T and Guan KL: Mechanisms
of Hippo pathway regulation. Genes Dev. 30:1–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao B, Li L, Tumaneng K, Wang CY and Guan
KL: A coordinated phosphorylation by Lats and CK1 regulates YAP
stability through SCF(beta-TRCP). Genes Dev. 24:72–85. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mossmann D, Park S and Hall MN: mTOR
signalling and cellular metabolism are mutual determinants in
cancer. Nat Rev Cancer. 18:744–757. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Li SH, Chien CY, Huang WT, Luo SD, Su YY,
Tien WY, Lan YC and Chen CH: Prognostic significance and function
of mammalian target of rapamycin in tongue squamous cell carcinoma.
Sci Rep. 7:81782017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Su W, Wang Y, Wang F, Zhang B, Zhang H,
Shen Y and Yang H: Circular RNA hsa_circ_0007059 indicates
prognosis and influences malignant behavior via AKT/mTOR in oral
squamous cell carcinoma. J Cell Physiol. 234:15156–15166. 2019.
View Article : Google Scholar
|
|
56
|
Zhang X, Liu N, Ma D, Liu L, Jiang L, Zhou
Y, Zeng X, Li J and Chen Q: Receptor for activated C kinase 1
(RACK1) promotes the progression of OSCC via the AKT/mTOR pathway.
Int J Oncol. 49:539–548. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Artinian N, Cloninger C, Holmes B,
Benavides-Serrato A, Bashir T and Gera J: Phosphorylation of the
Hippo pathway component AMOTL2 by the mTORC2 kinase promotes YAP
signaling, resulting in enhanced glioblastoma growth and
invasiveness. J Biol Chem. 290:19387–19401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gallant P and Steiger D: Myc's secret life
without Max. Cell Cycle. 8:3848–3853. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Pérez-Sayáns M, Suárez-Peñaranda JM,
Padín-Iruegas E, Gayoso-Diz P, Reis-De Almeida M, Barros-Angueira
F, Gándara-Vila P, Blanco-Carrión A and García-García A:
Quantitative determination of c-myc facilitates the assessment of
prognosis of OSCC patients. Oncol Rep. 31:1677–1682. 2014.
View Article : Google Scholar : PubMed/NCBI
|