Introduction
Pancreatic cancer (PaCa) is one of the most
aggressive types of cancer. In 2018, PaCa was the 4th most common
cause of all cancer-related deaths in Japan (1) and the third most common cause of death
among all cancer-related deaths in the USA (2). In 2019, over 56,000 patients were newly
diagnosed with PaCa and almost 46,000 patients died from PaCa in
the USA (2). After diagnosis, 24% of
patients survived for 1 year, and the 5-year overall survival rate
was only 9% in 2019, globally (3).
Furthermore, PaCa is highly resistant to chemotherapy (4); however, FOLFIRINOX and gemcitabine with
nab-paclitaxel have been found to improve the median survival time
to 11.1 months in France (2011) and 8.5 months in USA (2013),
respectively (5–7). Thus, there is an urgent requirement to
develop new therapeutic agents to reduce mortality rates in
patients with PaCa.
Numerous natural compounds, such as Curcuma longa
(turmeric) and Vitis vinifera (grape seed extract) have been
reported to have anticancer effects (8,9). Escin, a
natural triterpene saponin extracted from horse chestnuts
(Aesculus hippocastanum), has been widely used to treat
inflammation in Traditional Chinese Medicine in Korea, China, and
Japan (10). Previous studies have
found that escin had antitumor effects in various types of human
cancer cell, including glioblastoma, lung adenocarcinoma, melanoma,
hepatocellular carcinoma, PaCa, leukemia and osteosarcoma (10–14). In
addition, escin has been reported to inhibit migration and
invasion, and induced caspase-dependent apoptosis and autophagy
(9,13). Escin may inhibit NF-κB activation
(15,16); however, the specific mechanisms
involved are unknown.
NF-κB is a transcription factor that was first
discovered in 1986 and is reported that it plays essential roles in
carcinogenesis-related angiogenesis (17). NF-κΒ is also involved in the creation
of new blood vessels, which provides oxygen and nutrients to the
tumor cells, and in the progression of the growth of malignant
solid tumors (18). In solid tumors,
angiogenesis occurs via several steps, including extracellular
matrix remodeling, migration and proliferation of endothelial
cells, and capillary tube formation (19). New blood vessels formed in the tumor
allow tumor cells to circulate and metastasize to distant organs.
Our previous study reported that the liver metastatic potential of
PaCa cell lines was associated with angiogenesis and that NF-κB was
homeostatically activated in PaCa cells, with high metastatic
potential (19). These findings
indicated that agents blocking NF-κB activation can decrease
angiogenesis in PaCa, and additional studies have reported that
tumor growth and angiogenesis in several types of cancer, including
PaCa, were reduced by the inhibition of NF-κB activity (20,21). In
previous reports, interleukin (IL)-8 and vascular endothelial
growth factor (VEGF) have been identified as key mediators of
angiogenesis in PaCa (22–24). In addition, our previous study showed
that suppressing NF-κB activation decreased the secretion of both
IL-8 and VEGF in PaCa (18).
Furthermore, it was shown that several natural compounds inhibited
angiogenesis by suppressing NF-κB activity, and reducing VEGF and
IL-8 production (25,26). However, the mechanism of how escin
affects the angiogenesis of PaCa is not fully understood.
The present study aimed to clarify whether escin
inhibited angiogenesis in PaCa cells by suppressing nuclear
translocation of NF-κB.
Materials and methods
Reagents
Escin (C55H86O24;
CID 6476031) and dimethyl sulfoxide (DMSO) were purchased from
Sigma-Aldrich (Merck KGaA). Escin solution (50 mM) was prepared in
DMSO, stored as small amounts at −20°C, then thawed and diluted in
cell culture medium as required. Recombinant human tumor necrosis
factor (TNF)-α was purchased from R&D Systems Inc.
Cell lines and treatments
The human pancreatic adenocarcinoma cell lines,
BxPC-3, AsPC-1 and SW1990, and the immortalized human endothelial
cell line, EA.hy 926, were purchased from the American Type Culture
Collection. The BxPC-3 and AsPC-1 cell lines were maintained in
RPMI-1640 medium, while the SW1990 and EA.hy 926 cell lines were
maintained in DMEM (both from Sigma-Aldrich; Merck KGaA). Each
medium was replenished with 10% fetal bovine serum (FBS), 10 mg/ml
streptomycin, 10,000 U/ml penicillin, and 25 µg amphotericin B (all
from Gibco; Thermo Fisher Scientific, Inc.). All the cell lines
were cultured at 37°C in a humidified incubator with 5%
CO2.
Cytotoxicity assay
The cytotoxicity of escin was assessed with a Premix
WST-1 Cell Proliferation Assay System (Takara Bio, Inc.) according
to the manufacturer's protocols. Briefly, BxPC-3, AsPC-1 and SW1990
cell lines were seeded at 2×103 cells/100 µl/well in
96-well plates and cultured for 1 day. Then, various concentrations
of escin (0–30 µM) and DMSO (equivalent to the concentration
contained in 30 µM escin) were added to the cells. After incubation
for 72 h, the absorbance at 450 nm was measured using a SpectraMax
340 spectrophotometer (Molecular Devices, LLC).
Immunocytochemical analysis for NF-κB
p65 localization
The BxPC-3, AsPC-1 and SW1990 cell lines were
initially seeded at 1×104 cells/chamber in a 4-chamber
slide glass and cultured overnight. The cells were then treated
with escin (10 µM) for 2 h and stimulated with TNF-α (1 ng/ml) for
15 min before the end of the incubation. The cells that had not
been treated were used as controls. The cells were then washed with
PBS and fixed with 4% paraformaldehyde for 20 min at room
temperature. Next, the cells were washed and permeabilized with
0.1% Triton-X for 3 min and incubated with blocking buffer (3% BSA,
FUJIFILM Wako Pure Chemical Corporation) for 1 h at room
temperature. The cells were probed with anti-NF-κB p65 antibody
(cat. no. 8242T; Cell Signaling Technology, Inc.) overnight at 4°C.
Subsequently, the cells were washed and incubated with Alexa
Fluor® 488 goat anti-rabbit IgG (cat. no. ab150077;
Abcam) for 1 h at room temperature. Primary and secondary
antibodies were used at 1:400 and 1:500 dilution with 3% BSA,
respectively. The nuclei were visualized with DAPI staining at room
temperature for 10 min. Images of the stained slides were captured
using a BZ-X710 fluorescent microscope at ×100 (Keyence
Corporation).
Nuclear protein extraction and NF-κB
p65 activity assays
The BxPC-3, AsPC-1 and SW1990 cell lines were seeded
at 2×106 in 100-mm dishes with 10% FBS and cultured to
~80% confluence. The cells were then treated with escin (10 µM) in
5% FBS for 2 h and stimulated with or without TNF-α (5 ng/ml) for
30 min before the end of the incubation. Nuclear extracts were
obtained from the cells using a Nuclear Extraction kit (Active
Motif, Inc.). The concentrations of intranuclear proteins were
measured with a Pierce BCA protein assay kit (Thermo Fisher
Scientific, Inc.). Nuclear extracts were stored at −80°C until
further use. The NF-κB activity was determined using a Trans AM
NF-κB p65/p50 Transcription Factor Assay kit (cat. no. 40096;
Active Motif, Inc.) according to the manufacturer's protocols. A
total of 4 µg nuclear extract was used for the NF-κB activity
assays.
Western blot analysis of NF-κB p65 and
NF-κB phosphorylated (p)p65
The BxPC-3 cells were seeded at 2×106 in
100-mm dishes with 10% FBS and cultured to ~80% confluence. The
cells were then treated with escin (10 µM) in 5% FBS for 2 h and
stimulated with or without TNF-α (5 ng/ml) for 30 min before the
end of the incubation. Nuclear and cytoplasmic extracts were
obtained from the cells using a Nuclear Extraction kit (Active
Motif, Inc.). The concentrations of each protein were measured with
a Pierce BCA protein assay kit (Thermo Fisher Scientific, Inc.). A
total of 20 µg each protein extract was denatured at 90°C for 5 min
and separated on 10% Mini-PROTEAN TGX Precast gels (Bio-Rad
Laboratories, Inc.). The protein bands were transferred to
nitrocellulose membranes and blocked in iBind Flex Solution (iBind
Flex Buffer, iBind Flex Additive and distilled water; Thermo Fisher
Scientific, Inc.) for 15 min at room temperature. The primary and
secondary antibody reactions were performed using the iBind Flex
Western System (Thermo Fisher Scientific, Inc.) for 2.5 h at room
temperature according to the manufacturer's instructions. The
membranes were incubated with anti-p65 (1:1,000; cat. no. 8242S;
Cell Signaling Technology, Inc.), -p-p65 (1:1,000; cat. no. 3033S;
Cell Signaling Technology, Inc.), -GAPDH (1:2,000; cat. no.
SC-47724; Santa Cruz Biotechnology Inc.) and -TATA-box binding
protein (TBP; 1:1,000; cat. no. 22006-1-AP; ProteinTech Group,
Inc.) primary antibodies, then HRP-conjugated goat anti-rabbit
polyclonal secondary antibody (1:2,000; cat. no. P0448; Agilent
Technologies, Inc.). The protein-antibody complexes were visualized
with a SuperSignal West Pico Chemiluminescent Substrate,
SuperSignal West Femto Chemiluminescent Substrate, or Pierce ECL
Western Blotting Substrate (all from Thermo Fisher Scientific,
Inc.). The immunoreactive protein bands were detected using an
Amersham Imager 600 (Cytiva).
Reverse transcription-quantitative PCR
(RT-qPCR)
The BxPC-3, AsPC-1 and SW1990 cell lines were seeded
at 1×105 in 6-well plates with 10% FBS and cultured to
~80% confluence. Then, the cells were treated with or without 10 µM
escin and 5% FBS for 1 h and stimulated with TNF-α (1 ng/ml) for 15
min before the end of the incubation. Total RNA was extracted from
the cell pellets using a RNeasy Plus Mini kit (Qiagen GmbH)
according to the manufacturer's protocols and quantified using a
NanoDrop® 1000 (Thermo Fisher Scientific, Inc.). Total
RNA (1 µg) was reverse transcribed using a Super Script III
First-Strand Synthesis Super Mix for RT-qPCR (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocols.
RT-qPCR was performed using a Taqman Fast Advanced Master Mix and
Taqman Gene Expression Assays for VEGF (Hs00900055_m1),
IL-8 (Hs01553824_g1) and GAPDH (Hs99999905_m1) on a
7900HT Fast Real-Time PCR System (all from Applied Biosystems;
Thermo Fisher Scientific, Inc.). The following thermocycling
conditions were used: Initial denaturation at 95°C for 20 sec,
followed by 40 cycles at 95°C for 1 sec and 60°C for 20 sec. The
expression levels of VEGF and IL-8 were standardized
to those of GAPDH in each sample, using the relative
standard curve method (27).
ELISA
The BxPC-3, AsPC-1 and SW1990 cell lines were seeded
at 1×105 cells/well in a 6-well plate containing cell
specific medium as aforementioned, supplemented with 10% FBS and
incubated overnight at 37°C. The culture media were then changed,
and cells were incubated and stimulated with TNF-α (1 ng/ml) for 48
h in the presence of escin (10 µM). In addition, the BxPC-3 and
SW1990 cells were also incubated for an additional 48 h in the
presence of different doses of escin (0–10 µM) with 5% FBS after
culturing overnight as aforementioned. The culture media were then
collected and centrifuged at 400 × g for 5 min at 4°C to discard
particulates and stored at −80°C until further use. The
concentrations of IL-8 and VEGF were then measured using
appropriate ELISA kits (R&D Systems, Inc.) following the
manufacturer's protocols.
Tube formation assay for
angiogenesis
Tube formation was determined using the EA.hy 926
cell line and angiogenesis assays using Matrigel (Corning, Inc.).
The BxPC-3 and SW1990 cell lines were seeded at 1×105
cells/well in 6-well plates containing medium (RPMI-1640 and DMEM,
respectively) supplemented with 10% FBS and incubated overnight at
37°C. The culture media were then changed, and the cells were
incubated for an additional 48 h with or without escin (10 µM) in
2% FBS. The cell supernatants were then collected and centrifuged
at 400 × g for 5 min at 4°C to discard particulates. Matrigel was
added to a 96-well plate (50 µl/well) at 4°C and incubated for 30
min at 37°C for the Matrigel to solidify. The EA.hy 926 cells
(1.2×104 cells/well) were added on top of the Matrigel.
The cells were then incubated with mixed medium (50 µl of the
RPMI-1640 medium with 2% FBS and 50 µl of the aforementioned
supernatant per well) for 16 h to form capillary-like structures.
The cells incubated with RPMI medium with 2% FBS only were used as
the control. The EA.hy 926 cells (1.2×104 cells/well)
were also added on top of the Matrigel and incubated with RPMI
medium (2% FBS) containing 100 ng/ml recombinant IL-8 and VEGF
(both from R&D Systems Inc.), respectively. The number of
endotubes were counted under a confocal microscope (×40). A total
of 4 fields of view were analyzed per sample.
Statistical analysis
All experiments were performed in triplicate. All
the experimental data are represented as the mean ± SD. Comparisons
between two groups were assessed using unpaired t-tests, while
comparisons between multiple groups were determined using one-way
analysis of variance with Bonferroni's post hoc test for subsequent
comparison of individual groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Escin suppresses the proliferation of
the PaCa cells
The short-term effects of escin on the proliferation
of the PaCa cells (BxPC-3, AsPC-1 and SW1990) were examined using a
WST-1 assay after incubating the cells with various concentrations
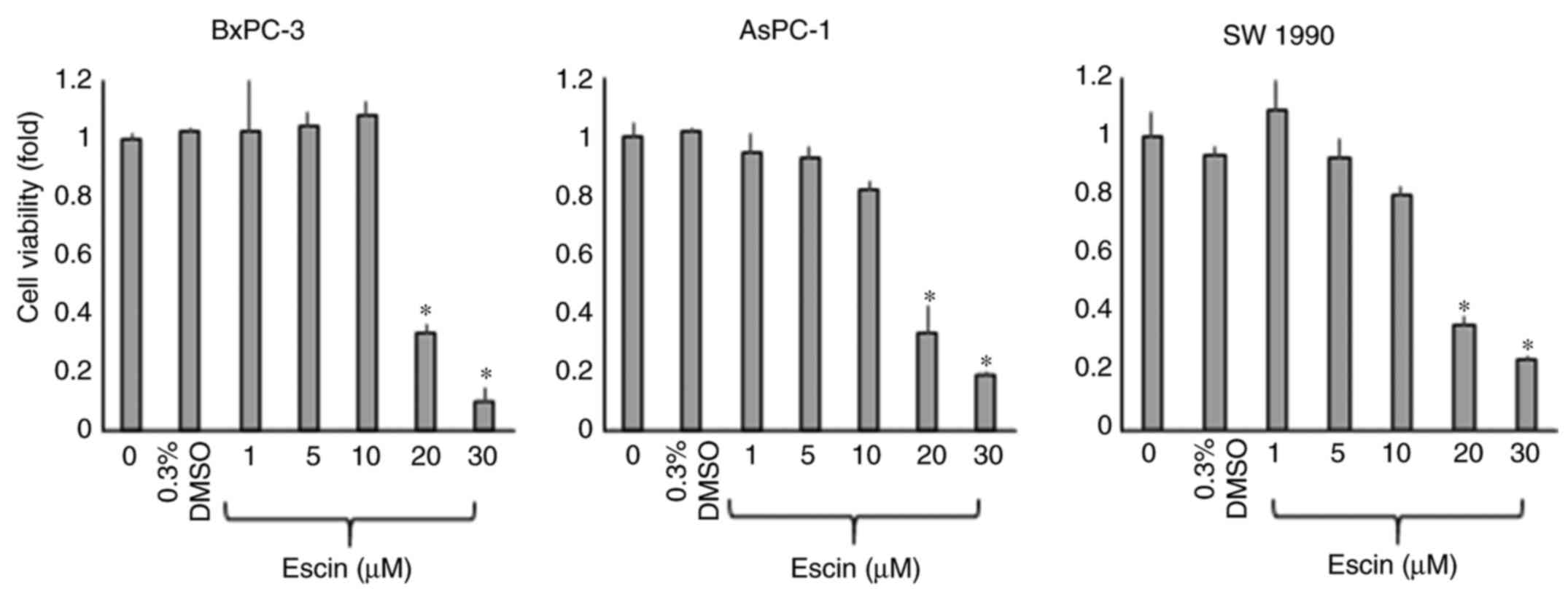
of escin for 72 h. The proliferation of all the PaCa cell lines was
significantly inhibited by >10 µM escin (Fig. 1). The half-maximal inhibitory
concentration (IC50) values were calculated from the
results of the WST-1 assays and were 17.2, 15.9 and 14.1 µM for the
BxPC-3, AsPC-1 and SW1990 cells, respectively. To avoid the effect
of cytotoxicity induced by escin, the concentration of escin was
set at less than the IC50 value in the subsequent
experiments.
Escin inhibits TNF-α-induced
translocation of NF-κB
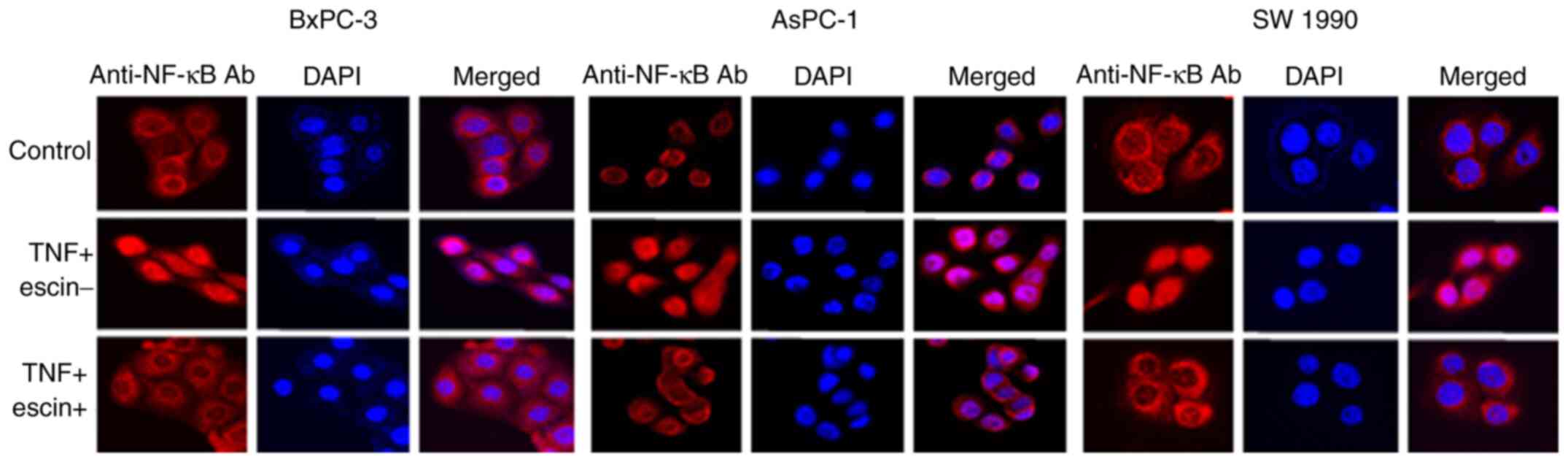
Immunocytochemical analysis was subsequently
performed to examine whether escin affected TNF-α-induced
translocation of p65 to the nucleus in the PaCa cells (Fig. 2). In cells treated with TNF-α alone,
p65 translocated into the nucleus, while in cells treated with
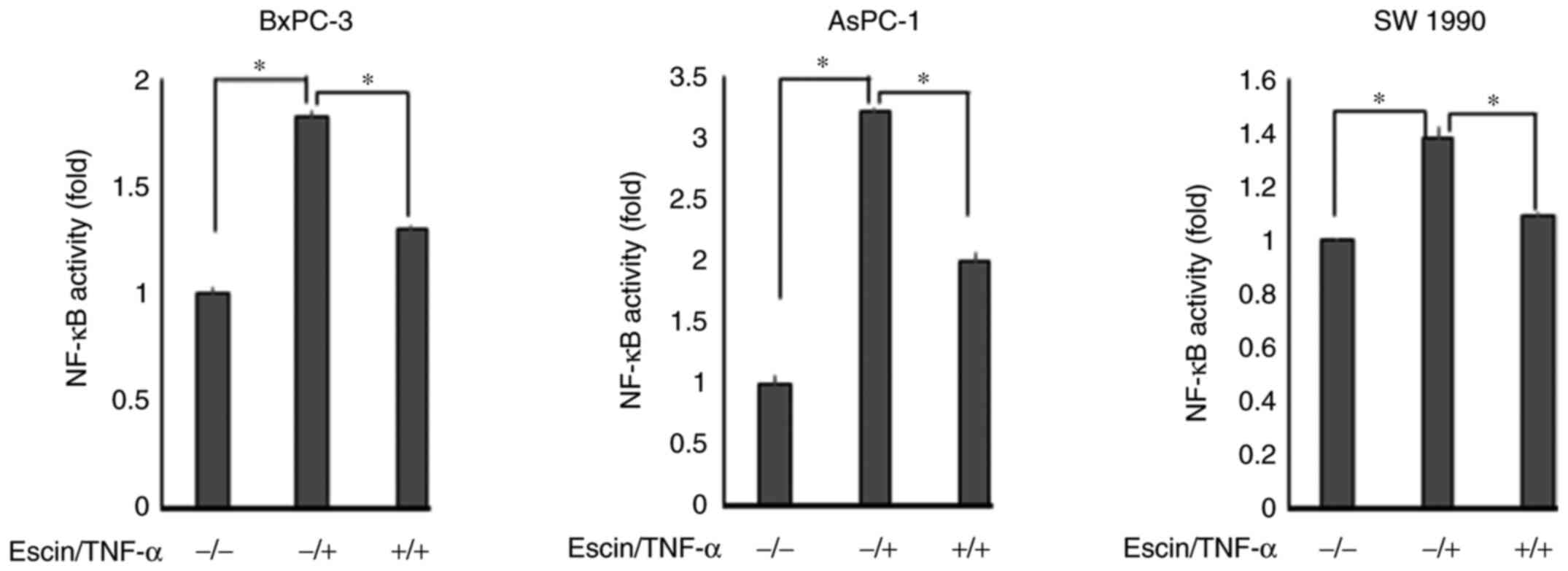
escin, p65 remained in the cytoplasm. The activity of p65
translocated into the nucleus was then determined using ELISA
(Fig. 3). The results demonstrated
that escin significantly reduced the TNF-α-induced activity of p65
in the nucleus. The activity of p65 was higher in cells treated
with both escin and TNF-α compared with that in cells that were
untreated; however, it is considered that the activation of p65 by
TNF-α stimulation was beyond the range of suppression by escin.
These results supported the hypothesis that escin inhibited the
translocation of p65. To further support the results that escin
inactivated NF-κB, western blot analysis was also performed to
determine the protein expression levels of p65 and p-p65 in the
cytoplasm and nucleus from the BxPC-3 cells, which were found to be
the most sensitive to escin in immunocytotochemical staining. Escin
notably reduced TNF-α-induced activation of both total p65 and
p-p65 in the cytoplasm and the nucleus (Fig. S1).
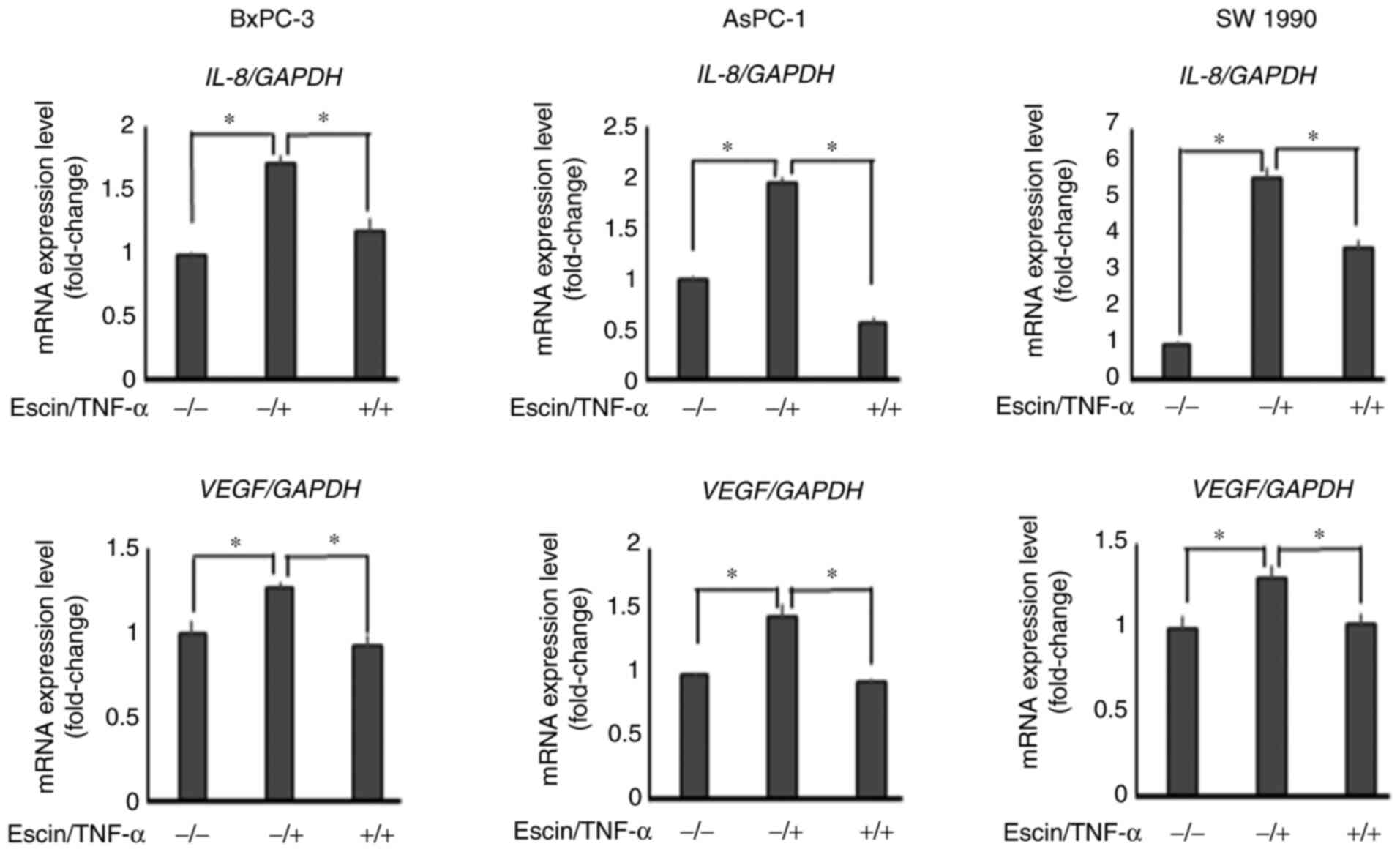
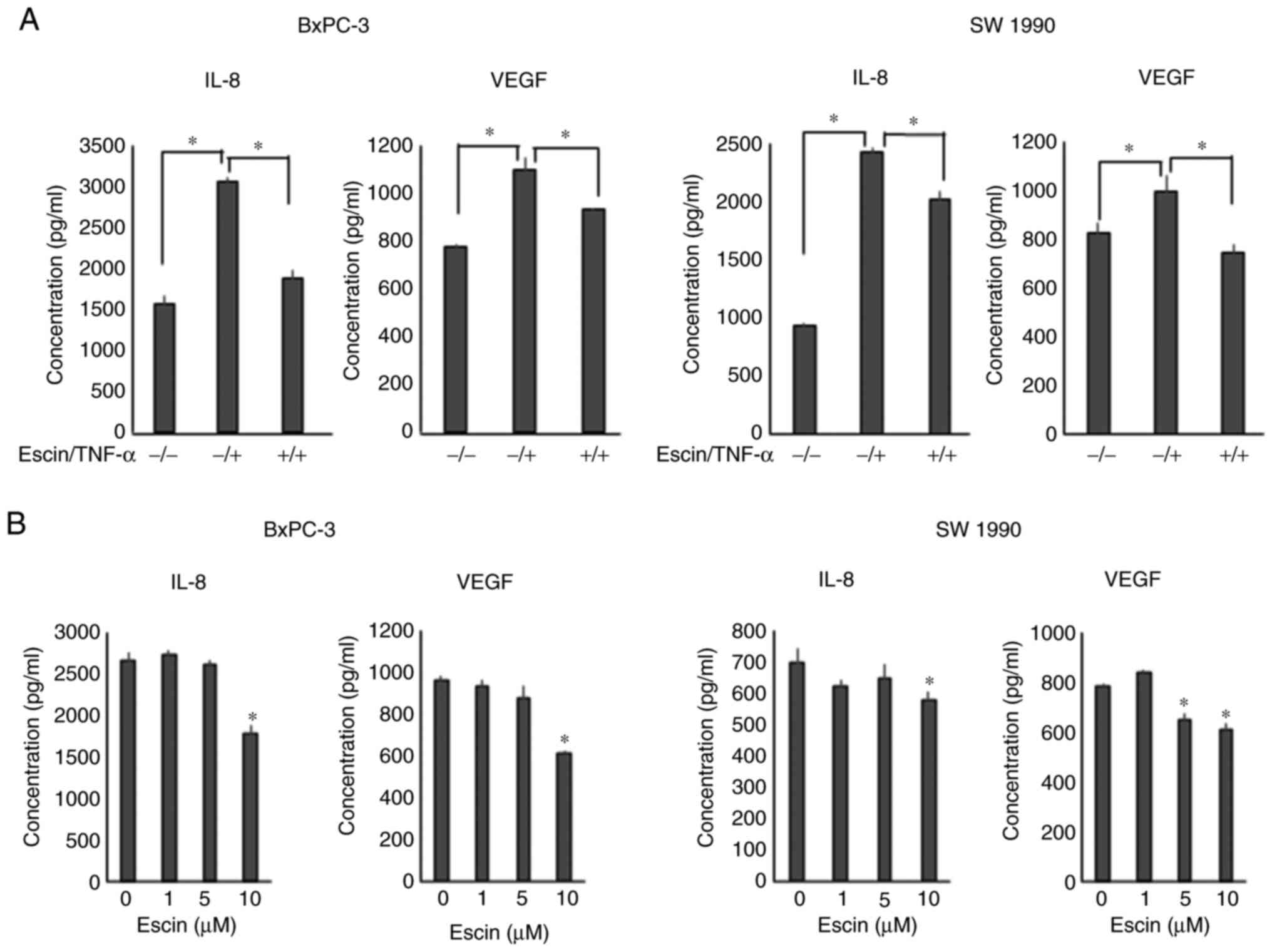
Escin downregulates TNF-α-induced mRNA
expression levels of IL-8 and VEGF
RT-qPCR revealed that escin (10 µM) significantly
decreased TNF-α-induced mRNA expression levels of IL-8 and
VEGF in PaCa cells (Fig.
4).
Escin suppresses the secretion of IL-8
and VEGF in the PaCa cells
Next, ELISA was performed to evaluate the protein
secretion levels of IL-8 and VEGF in PaCa cells. Escin (10 µM)
significantly suppressed TNF-α-induced protein secretion of IL-8
and VEGF in the PaCa cells (Fig. 5A).
With respect to the AsPC-1 cell line, secretion levels of both VEGF
and IL-8 were significantly enhanced by the treatment with TNF, and
the enhancement was significantly inhibited by escin (Fig. S2); however, the basal secretion
levels of both the angiogenic cytokines were lower compared with
that in the other cell lines. Therefore, the AsPC-1 cell line was
excluded from the next experiment. In addition, escin also
suppressed the secretion of these proteins in a dose-dependent
manner in the BxPC-3 and SW1990 cells (Fig. 5B). With respect to the inhibition of
IL-8 production, there were statistically significant differences;
however, the inhibition of TNF-α-induced IL-8 production in the
SW1990 cell line by escin was slightly lower compared with the
inhibition of VEGF production.
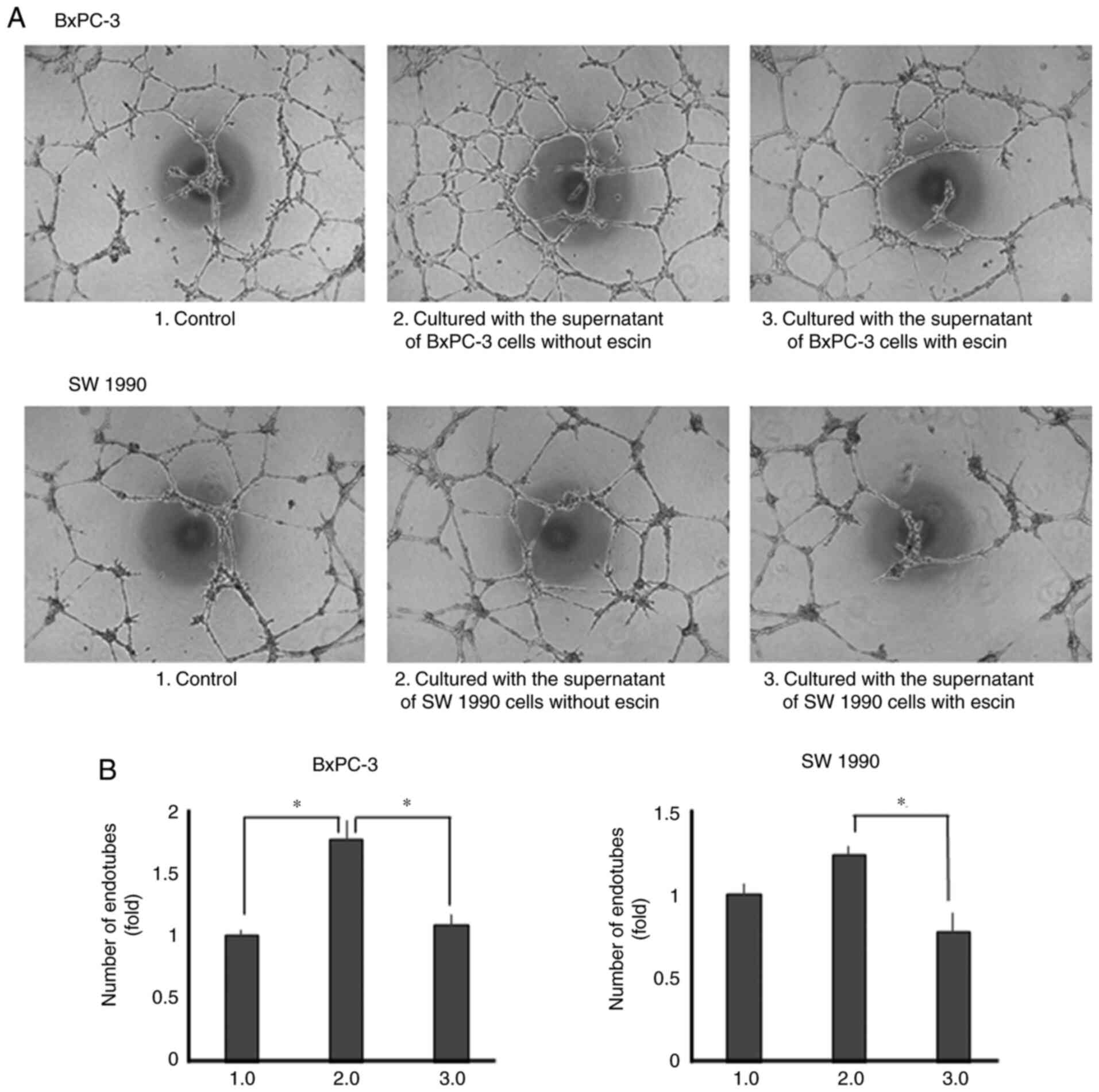
Escin inhibits tube formation in human
endothelial cells
Lastly, the effects of escin on tube formation in
human endothelial cells were determined. Tube formation was
enhanced when the cells were incubated with recombinant IL-8 and
VEGF (Fig. S3). Tube formation was
also enhanced when incubated with the supernatants from the
untreated PaCa cells. However, tube formation was significantly
decreased when the cells were incubated with supernatants from
escin-treated PaCa cells (Fig. 6A and
B).
Discussion
The aim of the present study was to clarify whether
escin, a natural compound extracted from horse chestnut, inhibited
the angiogenesis of PaCa by blocking NF-κB activation. The results
showed that escin suppressed cell proliferation and NF-κB
activation, and reduced the secretion of VEGF and IL-8 in several
human PaCa cell lines. In addition, escin suppressed human
endothelial cell line tube formation induced by PaCa cell
supernatants.
The progressive growth and metastasis of solid
malignant tumors, including PaCa, depend on angiogenic factors
released from the tumor and stromal cells (28). Various pro-angiogenic molecules,
including VEGF and IL-8, secreted by PaCa cells have been
identified and reported to mediate angiogenesis of PaCa (23,24,29). Our
previous study demonstrated that the higher the potential of liver
metastasis is, the more IL-8 is secreted in PaCa cell lines
(30).
In a previous study, NF-κB was reported to induce
the expression of various proteins (e.g. VEGF and IL-8) that are
involved in cell survival, apoptosis, proliferation, metastasis and
angiogenesis (31). Furthermore, ~70%
of PaCa cells exhibit constitutive activation of NF-κB (32), and the constitutive activity of NF-κB
in PaCa plays important roles in resistance to chemotherapy.
Several reports have demonstrated that high activity of the NF-κB
signaling pathway was involved in chemoresistance in PaCa (33–35).
In addition, our previous study demonstrated that
suppressing the activation of NF-κB using the proteasome inhibitor
MG132 blocked the production of pro-angiogenic molecules, such as
VEGF and IL-8 (18). These findings
were consistent with the results from the present study, that escin
reduced the production of pro-angiogenic molecules, including VEGF
and IL-8, by blocking NF-κB activation in PaCa cell lines.
Therefore, it logically follows that inhibiting the production of
these angiogenic factors by blocking NF-κB activity may suppress
angiogenesis in PaCa. Recently, NF-κB inhibitors, such as
bortezomib, have been administered to patients with multiple
myeloma (36). Unfortunately, these
inhibitors frequently have undesired side-effects, which prevent
their widespread use (36). Thus,
novel agents that suppress NF-κB activity with reduced toxicity are
required.
The present study has focused on the effects of
natural products, which are commonly considered safe and less
toxic. It has been reported that several natural products,
including curcumin, sesamin, zerumbone and escin, have anticancer
effects (9,26,37,38). In
PaCa, natural products exhibit anticancer effects by inhibiting
NF-κB (39). In addition, a previous
study showed that curcumin, which is a natural compound that also
inhibits NF-kB, similar to escin, had a synergistic effect on tumor
suppression when used in combination with gemcitabine (40). Therefore, it was hypothesized that
escin also has a similar effect by blocking NF-κB. Furthermore,
escin has already been widely used clinically to prevent
inflammatory edema due to inflammation caused by trauma, such as
surgery and fractures (41). Notably,
orally administered escin has been shown to be safe, whereas
injection of escin may cause phlebitis and allergies in animal
models (42).
To the best of our knowledge, this is the first
study associating escin with suppression of the production of
angiogenic factors, such as VEGF and IL-8, by inhibiting NF-κB
activity. When the study was started, it was hypothesized that
escin might have anti-angiogenic effects by reducing the production
of VEGF and IL-8 in the PaCa cells (at lower concentrations);
however, direct pro-apoptotic effects on PaCa cells (at higher
concentrations) were also found. At a concentration of 20 µM, escin
inhibited cell proliferation in several PaCa cell lines. To
eliminate the cytotoxicity of escin, all experiments were performed
at concentrations of escin below its IC50. No
significant cytotoxicity was observed at a concentration of 10 µM
by WST-1 assays. It was found that 10 µM escin reduced
TNF-α-induced NF-κB activation. It is conceivable that the pathway
via which TNF induces the production of IL-8 includes not only
NF-κB signaling, but also other pathways. For example, the
TNF/TRAF2 axis induces not only NF-κB activity, but also AP-1
activity in PaCa (43). AP-1 also
regulates IL-8 production in PaCa (44). In the present study, it was found that
escin significantly suppressed IL-8 and VEGF production; however,
it may be a partial suppression. Accordingly, it can be concluded
from the results that by inactivating NF-κB, escin caused
downregulation of VEGF and IL-8 production, affecting PaCa-induced
angiogenesis.
PaCa is generally referred to as an ischemic tumor
based on diagnostic imaging findings at the microscopic level;
however, there are numerous reports showing the associations
between microvessel density and the prognosis of PaCa (45–47). In
fact, some reports have demonstrated the effectiveness of
anti-angiogenic treatment for PaCa (48,49).
Furthermore, it was previously demonstrated by our research group
that new treatment targeting angiogenesis inhibited the tumor
growth of PaCa in vivo (25,50). Thus,
it was examined whether escin inhibited angiogenesis in human
endothelial cells. The EA.hy 926 cell line is an immortalized
endothelial cell line that has previously been used to estimate
angiogenic potential (51). The
results from the present study indicated that culturing EA.hy 926
cells with supernatants from the PaCa cell lines, which may contain
VEGF and IL-8, enhanced the angiogenic potential of the EA.hy 926
cells; however, culturing the cells with supernatants from
escin-treated PaCa cells, which may contain less VEGF and IL-8,
suppressed angiogenic potential. To the best of our knowledge, no
study has demonstrated the marked effects of escin on PaCa-induced
angiogenesis.
In conclusion, the results from the present study
indicated that a low concentration of escin inhibited angiogenesis
by reducing the secretion of VEGF and IL-8 by suppressing NF-κB
activation in PaCa. As escin did not affect the normal
proliferating function of the cells, escin may be a safer and less
toxic compound compared with other available treatments, such as
gemcitabine. Therefore, escin may have important applications as an
effective therapeutic agent for PaCa; however, further
investigation is required, such as in vivo experiments,
before escin can be used in a clinical setting. These animal
experiments with nude mice will be performed in future
investigations.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The current study was supported by Grants-in-Aid for
Scientific Research (Japan Society for the Promotion of Science;
grant no. 19K18157).
Availability of data and materials
The data generated or analyzed during this study are
included in the published article.
Authors' contributions
KO and YM contributed to the conception and design
of the study, analyzed and interpreted the data, and wrote and
reviewed the manuscript. KO, YM, GU, YH, HI, KS, KT, MM, HT and ST
designed the study. KO, YA, TK, YH, HI and GU acquired the data.
KO, YH and YM confirm the authenticity of all the raw data. YA, TK,
YH, MM and RO wrote the methods section of the manuscript. YM, HT,
RO and ST provided technical support (advising on determining
concentration and time) in performing the RT-PCR,
immunocytochemistry and angiogenesis assays. YM and ST supervised
the study. All authors read the final manuscript and are equally
responsible for all aspects of the study, ensuring that the
integrity or accuracy of all part of the study.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PaCa
|
pancreatic cancer
|
|
NF-κB
|
nuclear factor-κB
|
|
IL-8
|
interleukin-8
|
|
VEGF
|
vascular endothelial growth factor
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
IC50
|
half-maximal inhibitory
concentration
|
References
|
1
|
National Cancer Center Japan, Center for
Cancer Control and Information Services. 2020, https://ganjoho.jp/reg_stat/statistics/stat/summary.html
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rawla P, Sunkara T and Gaduputi V:
Epidemiology of pancreatic cancer: Global trends, etiology and risk
factors. World J Oncol. 10:10–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cao H, Le D and Yang LX: Current status in
chemotherapy for advanced pancreatic adenocarcinoma. Anticancer
Res. 33:1785–1791. 2013.PubMed/NCBI
|
|
5
|
Conroy T, Desseigne F, Ychou M, Bouché O,
Guimbaud R, Bécouarn Y, Adenis A, Raoul J, Gourgou-Bourgade S, de
la Fouchardière C, et al: FOLFIRINOX versus gemcitabine for
metastatic pancreatic cancer. N Engl J Med. 364:1817–1825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Von Hoff DD, Ervin T, Arena FP, hiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Torphy RJ, Fujiwara Y and Schulick RD:
Pancreatic cancer treatment: Better, but a long way to go. Surg
Today. 50:1117–1125. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Z, Dabrosin C, Yin X, Fuster MM,
Arreola A, Rathmell WK, Generali D, Nagaraju GP, El-Rayes B,
Ribatti D, et al: Broad targeting of angiogenesis for cancer
prevention and therapy. Semin Cancer Biol. 35 (Suppl 1):S224–S243.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gupta SC, Kim JH, Prasad S and Aggarwal
BB: Regulation of survival, proliferation, invasion, angiogenesis,
and metastasis of tumor cells through modulation of inflammatory
pathways by nutraceuticals. Cancer Metastasis Rev. 29:405–434.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kwak H, An H, Alam MB, Choi WS, Lee SY and
Lee SH: Inhibition of migration and invasion in melanoma cells by
β-escin via the ERK/NF-κB signaling pathway. Biol Pharm Bull.
41:1606–1610. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Çiftçi GA, Işcan A and Kutlu M: Escin
reduces cell proliferation and induces apoptosis on glioma and lung
adenocarcinoma cell lines. Cytotechnology. 67:893–904. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Harford-Wright E, Bidère N and Gavard J:
β-escin selectively targets the glioblastoma-initiating cell
population and reduces cell viability. Oncotarget. 7:66865–66879.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rimmon A, Vexler A, Berkovich L, Earon G,
Ron I and Lev-Ari S: Escin chemosensitizes human pancreatic cancer
cells and inhibits the nuclear factor-kappaB signaling pathway.
Biochem Res Int. 2013:2517522013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu J, Yu W, Liu B, Wang Y, Shao J, Wang
J, Xia K, Liang C, Fang W, Zhou C and Tao H: Escin induces
caspase-dependent apoptosis and autophagy through the ROS/p38 MAPK
signalling pathway in human osteosarcoma cells in vitro and in
vivo. Cell Death Dis. 8:e31132017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang YW, Wang SJ, Zhou YN, Pan SH and Sun
B: Escin augments the efficacy of gemcitabine through
down-regulation of nuclear factor-κB and nuclear
factor-κB-regulated gene products in pancreatic cancer both in
vitro and in vivo. J Cancer Res Clin Oncol. 138:785–797. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Harikumar KB, Sung B, Pandey MK, Guha S,
Krishnan S and Aggarwal BB: Escin, a pentacyclic triterpene,
chemosensitizes human tumor cells through inhibition of nuclear
factor-kappaB signaling pathway. Mol Pharmacol. 77:818–827. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pramanik K, Makena M, Bhowmick K and
Pandey M: Advancement of NF-κB signaling pathway: A novel target in
pancreatic cancer. Int J Mol Sci. 19:38902018. View Article : Google Scholar
|
|
18
|
Matsuo Y, Sawai H, Ochi N, Yasuda A,
Sakamoto M, Takahashi H, Funahashi H, Takeyama H and Guha S:
Proteasome inhibitor MG132 inhibits angiogenesis in pancreatic
cancer by blocking NF-kappaB activity. Dig Dis Sci. 55:1167–1176.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bussolino F, Mantovani A and Persico G:
Molecular mechanisms of blood vessel formation. Trends Biochem Sci.
22:251–256. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sosmitha G, Bano S, Javadi M, Lu F,
Clarissa EH, Frank A, Kwang Seok A, Gautam S and Ajaikumar BK:
Zerumbone as an anti-cancer agent. Molecules. 24:7342019.
View Article : Google Scholar
|
|
21
|
Meteoglu I, Erdogdu IH, Meydan N, Erkus M
and Barutca S: NF-KappaB expression correlates with apoptosis and
angiogenesis in clear cell renal cell carcinoma tissues. J Exp Clin
Cancer Res. 27:532008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Itakura J, Ishiwata T, Friess H, Fujii H,
Matsumoto Y, Büchler MW and Korc M: Enhanced expression of vascular
endothelial growth factor in human pancreatic cancer correlates
with local disease progression. Clin Cancer Res. 3:1309–1316.
1997.PubMed/NCBI
|
|
23
|
Shi Q, Abbruzzese JL, Huang S, Fidler IJ,
Xiong Q and Xie K: Constitutive and inducible interleukin 8
expression by hypoxia and acidosis renders human pancreatic cancer
cells more tumorigenic and metastatic. Clin Cancer Res.
5:3711–3721. 1999.PubMed/NCBI
|
|
24
|
Shi Q, Le X, Abbruzzese JL, Peng Z, Qian
CN, Tang H, Xiong Q, Wang B, Li XC and Xie K: Constitutive Sp1
activity is essential for differential constitutive expression of
vascular endothelial growth factor in human pancreatic
adenocarcinoma. Cancer Res. 61:4143–4154. 2001.PubMed/NCBI
|
|
25
|
Saito K, Matsuo Y, Imafuji H, Okubo T,
Maeda Y, Sato T, Shamoto T, Tsuboi K, Morimoto M, Takahashi H, et
al: Xanthohumol inhibits angiogenesis by suppressing nuclear
factor-κB activation in pancreatic cancer. Cancer Sci. 109:132–140.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tsuboi K, Matsuo Y, Shamoto T, Shibata T,
Koide S, Morimoto M, Guha S, Sung B, Aggarwal BB, Takahashi H and
Takeyama H: Zerumbone inhibits tumor angiogenesis via NF-κB in
gastric cancer. Oncol Rep. 31:57–64. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bustin S: Quantification of mRNA using
real-time reverse transcription PCR (RT-PCR): Trends and problems.
J Mol Endocrinol. 29:23–39. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Folkman J: Angiogenesis and angiogenesis
inhibition: An overview. Exs. 79:1–8. 1997.PubMed/NCBI
|
|
29
|
Brown LF, Berse B, Jackman RW, Tognazzi K,
Manseau EJ, Senger DR and Dvorak HF: Expression of vascular
permeability factor (vascular endothelial growth factor) and its
receptors in adenocarcinomas of the gastrointestinal tract. Cancer
Res. 53:4727–4735. 1993.PubMed/NCBI
|
|
30
|
Sawai H, Funahashi H, Okada Y, Matsuo Y,
Sakamoto M, Yamamoto M, Takeyama H and Manabe T: Interleukin-1alpha
enhances IL-8 secretion through p38 mitogen-activated protein
kinase and reactive oxygen species signaling in human pancreatic
cancer cells. Med Sci Monit. 11:BR343–BR350. 2005.PubMed/NCBI
|
|
31
|
Aggarwal BB: Nuclear factor-kappaB: The
enemy within. Cancer Cell. 6:203–208. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xiong HQ, Abbruzzese JL, Lin E, Wang L,
Zheng L and Xie K: NF-kappaB activity blockade impairs the
angiogenic potential of human pancreatic cancer cells. Int J
Cancer. 108:181–188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Arlt A and Schäfer H: NFkappaB-dependent
chemoresistance in solid tumors. Int J Clin Pharmacol Ther.
40:336–347. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sebens S, Arlt A and Schäfer H: NF-kappaB
as a molecular target in the therapy of pancreatic carcinoma.
Recent Results Cancer Res. 177:151–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Holcomb B, Yip-Schneider M and Schmidt CM:
The role of nuclear factor kappaB in pancreatic cancer and the
clinical applications of targeted therapy. Pancreas. 36:225–235.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen D, Frezza M, Schmitt S, Kanwar J and
Dou QP: Bortezomib as the first proteasome inhibitor anticancer
drug: Current status and future perspectives. Curr Cancer Drug
Targets. 11:239–253. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aggarwal BB, Van Kuiken ME, Iyer LH,
Harikumar KB and Sung B: Molecular targets of nutraceuticals
derived from dietary spices: Potential role in suppression of
inflammation and tumorigenesis. Exp Biol Med (Maywood).
234:825–849. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Harikumar KB, Sung B, Tharakan ST, Pandey
MK, Joy B, Guha S, Krishnan S and Aggarwal BB: Sesamin manifests
chemopreventive effects through the suppression of
NF-kappaB-regulated cell survival, proliferation, invasion, and
angiogenic gene products. Mol Cancer Res. 8:751–761. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yue Q, Gao G, Zou G, Yu H and Zheng X:
Natural products as adjunctive treatment for pancreatic cancer:
Recent trends and advancements. Biomed Res Int. 2017:84125082017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kunnumakkara AB, Guha S, Krishnan S,
Diagaradjane P, Gelovani J and Aggarwal BB: Curcumin potentiates
antitumor activity of gemcitabine in an orthotopic model of
pancreatic cancer through suppression of proliferation,
angiogenesis, and inhibition of nuclear factor-kappaB-regulated
gene products. Cancer Res. 67:3853–3861. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gallelli L: Escin: A review of its
anti-edematous, anti-inflammatory, and venotonic properties. Drug
Des Devel Ther. 13:3425–3437. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li M, Lu C, Zhang L, Zhang J, Du Y, Duan
S, Wang T and Fu F: Oral administration of escin inhibits acute
inflammation and reduces intestinal mucosal injury in animal
models. Evid Based Complement Alternat Med.
2015:5036172015.PubMed/NCBI
|
|
43
|
Trauzold A, Röder C, Sipos B, Karsten K,
Arlt A, Jiang P, Martin-Subero JI, Siegmund D, Müerköster S,
Pagerols-Raluy L, et al: CD95 and TRAF2 promote invasiveness of
pancreatic cancer cells. FASEB J. 19:620–622. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sclabas GM, Fujioka S, Schmidt C, Li Z,
Frederick WA, Yang W, Yokoi K, Evans DB, Abbruzzese JL, Hess KR, et
al: Overexpression of tropomysin-related kinase B in metastatic
human pancreatic cancer cells. Clin Cancer Res. 11:440–449.
2005.PubMed/NCBI
|
|
45
|
Stipa F, Lucandri G, Limiti MR, Bartolucci
P, Cavallini M, Di Carlo V, D'Amato A, Ribotta G and Stipa S:
Angiogenesis as a prognostic indicator in pancreatic ductal
adenocarcinoma. Anticancer Res. 22:445–449. 2002.PubMed/NCBI
|
|
46
|
Benckert C, Thelen A, Cramer T, Weichert
W, Gaebelein G, Gessner R and Jonas S: Impact of microvessel
density on lymph node metastasis and survival after curative
resection of pancreatic cancer. Surg Today. 42:169–176. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Amin Z, Theis B, Russell RC, House C,
Novelli M and Lees WR: Diagnosing pancreatic cancer: The role of
percutaneous biopsy and CT. Clin Radiol. 61:996–1002. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Annese T, Tamma R, Ruggieri S and Ribatti
D: Angiogenesis in pancreatic cancer: Pre-clinical and clinical
studies. Cancers (Basel). 11:3812019. View Article : Google Scholar
|
|
49
|
Zhang Z, Ji S, Zhang B, Liu J, Qin Y, Xu J
and Yu X: Role of angiogenesis in pancreatic cancer biology and
therapy. Biomed Pharmacother. 108:1135–1140. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Matsuo Y, Raimondo M, Woodward TA, Wallace
MB, Gill RA, Tong Z, Burdick MD, Yang Z, Strieter RM, Hoffman RM
and Guha S: CXC-chemokine/CXCR2 biological axis promotes
angiogenesis in vitro and in vivo in pancreatic cancer. Int J
Cancer. 125:1027–1037. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang HL, Chang HC, Lin SW, Kumar KJS, Liao
CH, Wang HM, Lin KY and Hseu YC: Antrodia salmonea inhibits
TNF-α-induced angiogenesis and atherogenesis in human endothelial
cells through the down-regulation of NF-κB and up-regulation of
Nrf2 signaling pathways. J Ethnopharmacol. 151:394–406. 2014.
View Article : Google Scholar : PubMed/NCBI
|