Introduction
Pancreatic cancer (PC) is an aggressive malignancy
and was predicted to be the second leading cause of cancer-related
deaths in 2020 (1,2). Pancreatic ductal adenocarcinoma is the
most frequent histological subtype. Despite advances in cancer
treatment in recent decades, PC has continued to have poor clinical
outcomes due to its late presentation with nonspecific symptoms and
early metastatic nature (3). The
overall 5-year survival rate for PC is 8%, which is the lowest
compared with all other solid tumors (4). Given the high mortality, novel therapies
to maximize life length and quality of life of patients with PC are
urgently required. Thus, a thorough investigation of the mechanisms
underlying the progression of PC is warranted.
Long noncoding RNAs (lncRNAs) are a class of
noncoding transcripts longer than 200 nucleotides that have been
recognized to play important roles in various physiological and
pathological processes, such as cell proliferation and apoptosis,
cell growth and senescence, and immune activation or inactivation
(5–7).
One well-defined mechanism by which lncRNAs act is by competition
with endogenous RNAs (ceRNAs) to sponge microRNAs (miRNAs), thus
affecting their regulatory function (8). miRNAs are a class of noncoding
transcripts of approximately 22 nucleotides, which are widely known
to participate in posttranscriptional regulation of gene expression
via binding to the 3′-untranslated regions (3′-UTRs) of target
messenger RNAs (mRNAs) and thus are deeply involved in a variety of
biological activities (9,10). The sophisticated regulatory network
involving lncRNA, miRNA, and mRNA is attracting increasing research
attention worldwide.
LncRNA GATA3-antisense RNA 1 (GATA3-AS1) is a
divergent lncRNA neighboring GATA3. GATA3-AS1 has been reported to
share a promoter region with GATA3, and it is necessary for
efficient transcription of GATA3 (11). One study demonstrated that GATA3 was
markedly overexpressed in PC samples (12). Moreover, GATA3-AS1 was recently
recognized as an oncogene in hepatocellular carcinoma (HCC)
(13). Therefore, based on existing
research, it was hypothesized that GATA3-AS1 is dysregulated and
acts as a modulating molecular mechanism in the development of PC.
The aim of the present study was to characterize the dysregulation
and the biological roles of GATA3-AS1 in PC and to clarify the
possible miRNA-mRNA regulatory axis involved.
Materials and methods
Patient samples
Cancer tissues and adjacent noncancerous tissues
(normal tissues) were collected from 30 patients with PC at the
First Affiliated Hospital of Xi'an Jiaotong University who
underwent curative resection for PC between August 2017 and October
2019. Among them, 17 were males, and 13 were females. Their ages
were between 35 and 71 years, and the mean age was 56 years. None
of the subjects had received any biotherapy or chemotherapy before
recruitment to this study. Informed consent was obtained from all
participants, and the study protocol was approved by the Ethics
Committee of the First Affiliated Hospital of Xi'an Jiaotong
University.
Cell culture
The human immortalized pancreatic duct epithelial
cell line HPDE6-C7 and PC cell lines BxPC-3, Capan-2, SW1990,
PANC-1, and AsPC-1 were obtained from American Type Culture
Collection and cultured in Dulbecco's modified Eagle's medium
(DMEM) or RPMI-1640 medium supplemented with 10% fetal bovine serum
(FBS) and 100 U/ml penicillin/streptomycin (all from Gibco; Thermo
Fisher Scientific, Inc.), in a humidified atmosphere with 5%
CO2 at 37°C.
Cell transfection
Small interfering RNAs (siRNAs) specifically
targeting GATA3-AS1 (sequence: 5′-UCUCCGCGCGUCAAUCGA-3′), as well
as the corresponding negative control (NC; sequence:
5′-CUACACCGUAUUCUACUACUA-3′), miR-30b-5p mimic (sequence:
5′-UGUAAACAUCCUACACUCAGCU-3′) and mimic-NC (sequence:
5′-UUCUCCGAACGUGUCACGUTT-3′) were obtained from Shanghai GenePharma
Co., Ltd. pcDNA3.1-GATA3-AS1 and pcDNA3.1-Tex10 plasmids were
constructed by Shanghai Sangon Biotech Co., Ltd. In a 24-well
plate, cell transfection with siRNAs or si-NC (30 pmol/well),
mimics or mimic-NC (30 pmol/well), or plasmids (0.5 µg/well) was
performed using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions at
room temperature. Unless otherwise specified, subsequent
experimentation was performed after 48 h of transfection. An
untransfected (normal cultured cells) group was identified as the
control group.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from cancer tissues and cell
lines using Trizol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) in accordance with the manufacturer's instructions.
Complementary DNA (cDNA) was reverse transcribed using SuperScript
First Strand cDNA System (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's instructions. The levels of
GATA3-AS1 and mRNA expression of Tex10 were assessed using SYBR
Premix Ex Taq kit (Takara Biotechnology Co., Ltd.) following the
manufacturer's instructions. GAPDH was used as an internal control.
The levels of miR-30b-5p were assessed using TaqMan microRNA assays
(Applied Biosystems; Thermo Fisher Scientific, Inc.) following the
manufacturer's instructions. U6 was used as an internal control.
qPCR was performed on an Applied Biosystems 7500 Real-Time PCR
System (Applied Biosystems; Thermo Fisher Scientific, Inc.) under
the following thermocycling conditions: Initial denaturation at
95°C for 5 min followed by 40 cycles at 95°C for 15 sec
(denaturation), 58°C for 30 sec (annealing) and 72°C for 30 sec
(elongation), and final extension at 72°C for 5 min. The relative
expression was calculated using the 2−ΔΔCq method
(14). The primer sequences are
presented in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Primer sequences
(5′-3′) |
|---|
| GATA3-AS1 | F:
AAGTTGAGCGGGGTATGT |
|
| R:
TTTCTGGCCTTTGGTGTC |
| miR-30b-5p | F:
ACACTCCAGCTGGGTGTAAACATCCTACAC |
|
| R:
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGCTGAGT |
| Tex10 | F:
TTGCAACTTGCTCATCTTGG |
|
| R:
AGAGTCTGCAGGGAGAACCA |
| GAPDH | F:
AGAACGGGAAGCTTGTCATC |
|
| R:
CATCGCCCCACTTGATTTTG |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
Cell viability assay
Cell viability was assessed using Cell Counting Kit
(CCK)-8 (Beyotime Institute of Biotechnology) at 0, 24, 48, 72 and
96 h post transfection according to the manufacturer's
instructions. Briefly, 10 µl of CCK-8 solution was added to each
well (4×104 cells) of 96-well plates, and the cells were
incubated with CCK-8 solution for 4 h at 37°C. The absorbance value
was measured at 450 nm using a microplate reader (Thermo Fisher
Scientific, Inc.).
Colony formation assay
Cells (1×106) were seeded into 6-well
plates and cultured in DMEM or RPMI-1640 medium with 10% FBS at
37°C. After 2 weeks, the cells were fixed using 4% paraformaldehyde
for 20 min, and then stained using 1% crystal violet dye for 20 min
at room temperature, and then the number of colonies was
counted.
Cell apoptosis assay
Cell apoptosis was assessed by flow cytometry 48 h
post transfection using an Annexin V-fluorescein isothiocyanate
(FITC) apoptosis detection kit (Beyotime Institute of
Biotechnology) according to the manufacturer's instructions.
Briefly, 1×106 cells were seeded into 6-well plates and
48 h post-transfection, the cells were collected, washed, and
resuspended in binding buffer. Then, the cells were incubated with
5 µl of Annexin V-FITC and 10 µl of propidium iodide at room
temperature in the dark for 20 min. Cell apoptosis was assessed
using a FACSCalibur (BD Biosciences) within 1 h. FlowJo software
Ver.10 (Tree Star, Inc.) was used for analysis.
Transwell assay
The invasion assay was performed using 24-well
Transwell chambers with 8-µm pores (Corning, Inc.); the chambers
were precoated with Matrigel for 30 min at 37°C. Cells were
harvested at 24 h post transfection and then 200 µl of cell
suspension in serum-free medium (4×104 cells) was seeded
in the upper chamber of Transwell plates, while 600 µl of medium
containing 20% FBS was added to the lower chamber. After 48 h of
incubation (72 h post transfection), cells remaining in the upper
chamber were removed by cotton swabs, while the cells on the filter
surface were fixed using 4% paraformaldehyde for 20 min and then
stained using 0.1% crystal violet dye for 20 min at room
temperature, and then counted in five random fields under an
optical inverted microscope (magnification, ×200).
Spheroid formation assay
Cells (1×103) were seeded in Ultra Low
Attachment 24-well plates (Corning, Inc.) and cultured in
serum-free DMEM/F12 medium (Invitrogen; Thermo Fisher Scientific,
Inc.), supplemented with 2% B27 (Gibco; Thermo Fisher Scientific,
Inc.), 20 ng/ml epidermal growth factor, and 10 ng/ml basic
fibroblast growth factor (PeproTech, Inc.) containing 100 U/ml
penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
Seven days later, the formed spheroids were counted under an
optical inverted microscope (magnification, ×200).
Dual-luciferase reporter assay
The starBase v2.0 (http://starbase.sysu.edu.cn/) was used in the
bioinformatics analysis to predict the binding sites between
GATA3-AS1 and miR-30b-5p, and between miR-30b-5p and Tex10
(15). The sequences of GATA3-AS1
wild-type (wt, chr10:8,092,655-8,092,876) and Tex10 wt
(chr9:103,064,259-103,064,483), as well as their corresponding
mutant sequences were synthesized and inserted into the pGL3
vectors by Shanghai Sangon Biotech Co., Ltd., and named as
pGL3-GATA3-AS1 wt, pGL3-GATA3-AS1 mut, pGL3-Tex10 wt, and
pGL3-Tex10 mut plasmids, respectively. Cells (4×104)
were co-transfected with indicated pGL3 plasmids and miR-30b-5p
mimic or mimic NC, along with pRL-TK plasmids (Promega
Corporation), using Lipofectamine 2000 according to the
manufacturer's instructions. The luciferase activity was monitored
48 h post transfection using Dual-Glo Luciferase assay (Promega
Corporation) according to the manufacturer's instructions. The
relative firefly luciferase activity was normalized to
Renilla luciferase activity.
Western blotting
Total protein was extracted from PC tissues and cell
lines using RIPA buffer (Beyotime Institute of Biotechnology)
according to the manufacturer's instructions. The BCA method was
used for protein quantification. Equal amounts (30 µg) of proteins
were separated by 10% SDS-PAGE and transferred onto nitrocellulose
membranes (EMD Millipore), followed by blocking with 5% nonfat milk
for 1 h at room temperature. Then, the membranes were incubated
with anti-Tex10 (1:1,000; cat. no. 720257; Invitrogen; Thermo
Fisher Scientific, Inc.), anti-Wnt1 (1:500; ab15251; Abcam) or
anti-β-catenin (1:500; ab16051; Abcam) at 4°C overnight and then
incubated with horseradish-peroxidase-conjugated secondary antibody
(1:10,000; cat. no. BA1055; Boster Biological Technology) at room
temperature for 1 h. The protein bands were visualized using an
enhanced chemiluminescence detection kit (Amersham Biosciences;
Cytiva). GAPDH was used as an internal control. The density of the
bands was quantified by ImageJ 1.52a (National Institutes of
Health).
Lentiviral vectors and animal
experiments
Lentiviral vectors encoding short hairpin
(sh)-GATA3-AS1 or shRNA NC were constructed and packaged by
Shanghai GenePharma Co., Ltd. Briefly, the 2nd generation system
was used. Lentiviral plasmid (20 µg), and two packaging components
(15 µg or 5 µg, respectively) were co-transfected into 293T cells
using Lipofectamine 2000 according to the manufacturer's protocol.
At 8 h post-transfection, the medium was replaced with medium
containing 10% serum, and the cells were cultured for an additional
48-72 h. Subsequently, the supernatant containing lentiviral
particles was harvested and concentrated by ultracentrifugation to
obtain a high-titer lentivirus concentration. Then the virus
solution was added to the cells at the multiplicity of infection
(MOI) of 40 for 72 h. Finally, after 5-7 days of purinomycin
screening, the stable cells were collected and passaged at least
three times for subsequent animal experiments.
A total of 16 BALB/c nude mice (male; aged 4-6
weeks) were purchased from the Chengdu DOSSY Experimental Animals
Co., Ltd. All the mice were housed under 20-25°C in a humidified
room (humidity, 40-55%) with a 12-h light-dark cycle and free
access to food and water. The PANC-1 cells or AsPC-1 cells infected
with lentivirus-shRNA NC or lentivirus-sh-GATA3-AS1 were
subcutaneously injected into the axilla of nude mice, respectively
(n=4 per group). Animal health was monitored every day throughout
the study, and the mice were to be euthanized whenever they
exhibited persistent lethargy and loss of appetite. However, no
mice exhibited the aforementioned humane endpoints and the 16
tumor-bearing mice all survived throughout the present study. Every
3 days, the tumor size was monitored and the maximum tumor size was
≤1 cm3. Two weeks later, the mice were euthanized by
cervical dislocation. Both, no fluctuation in the chest cavity
(respiratory arrest) and no breathing and heartbeat sound (cardiac
arrest) confirmed the sacrifice of the mice. Then the tumors were
isolated and weighed. The animal study was approved by the Animal
Care and Use Committee of the First Affiliated Hospital of Xi'an
Jiaotong University.
Statistical analysis
Statistical analysis was carried out with GraphPad
Prism 8.0 (GraphPad Software, Inc). All data are expressed as the
mean ± standard deviation of at least three independent
experiments. The differences were analyzed by paired t-test
between two groups, and one-way analysis of variance followed by
Bonferroni's post hoc test among more than two groups. Correlation
of molecular expression was analyzed by linear regression.
P<0.05 was considered to indicate a statistically significant
difference.
Results
LncRNA GATA3-AS1 expression is
upregulated in human PC tissues and cell lines
The expression level of lncRNA GATA3-AS1 was first
assessed in PC tissues and cell lines. As revealed in Fig. 1A, the expression of GATA3-AS1 in the
cancer tissues from patients with PC was significantly upregulated,
compared with the normal tissues. As revealed in Fig. 1B, the expression of GATA3-AS1 in the
PC cell lines BxPC-3, Capan-2, SW1990, PANC-1, and AsPC-1 was also
significantly upregulated, compared with the normal human
pancreatic duct epithelial cell line HPDE6-C7. To assess the
biological effects of GATA3-AS1 upregulation, its expression was
knocked down in PANC-1 and AsPC-1 cells by transfecting them with
siRNAs. As revealed in Fig. 1C and D,
compared with the siRNA NC group, the interference efficiency
exhibited no obvious difference between si-GATA3-AS1-1 and
si-GATA3-AS1-2, and both decreased the expression of GATA3-AS1 in
PANC-1 and AsPC-1 cells. In the following experiments,
si-GATA3-AS1-2 was selected to interfere with GATA3-AS1
expression.
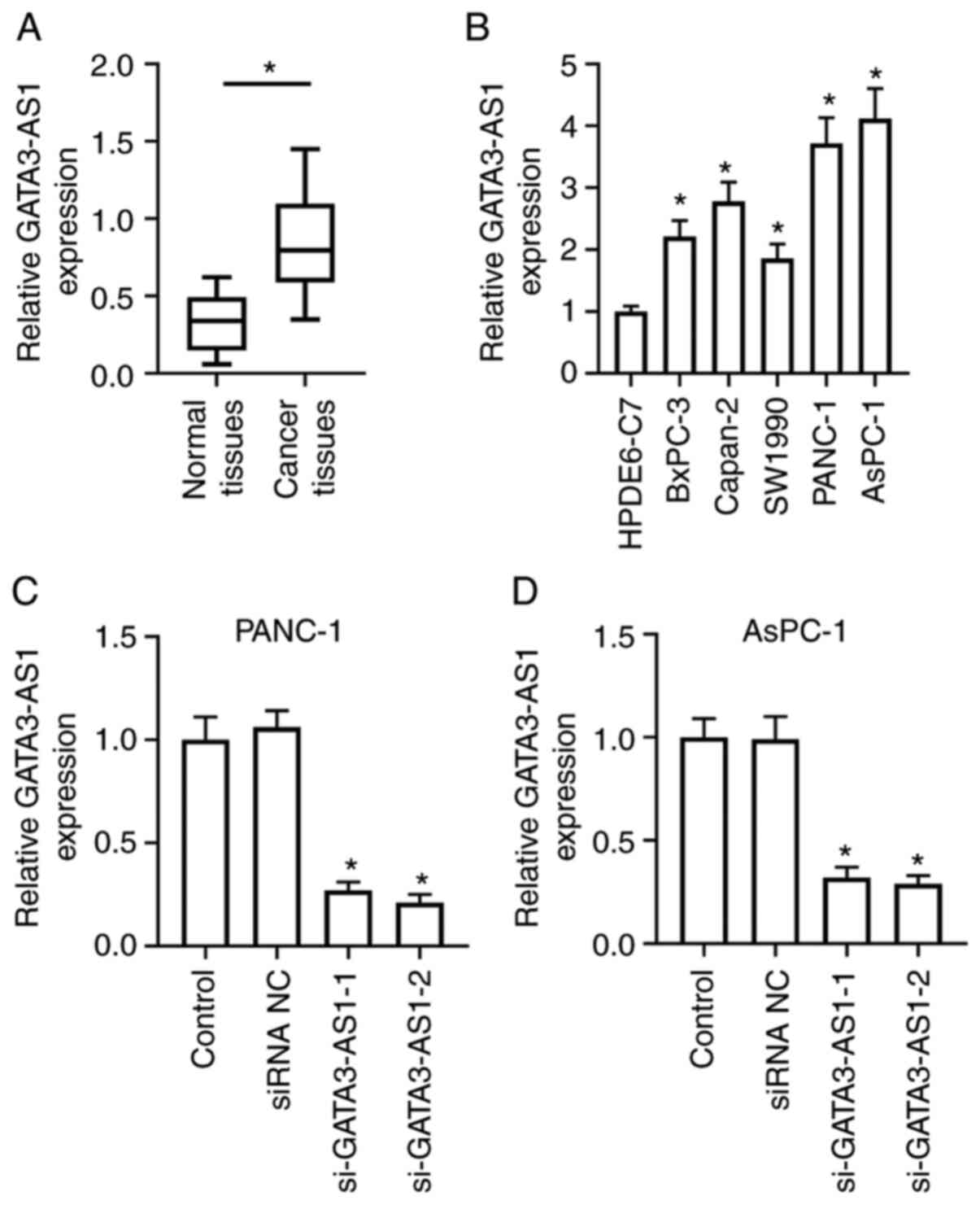 | Figure 1.LncRNA GATA3-AS1 expression is
upregulated in human PC tissues and cell lines. (A) The expression
of GATA3-AS1 in 30 pairs of PC tissues and matched normal tissues
was assessed by qPCR. *P<0.05, compared with the normal tissues.
(B) The expression of GATA3-AS1 in normal human pancreatic duct
epithelial cell line HPDE6-C7, and in PC cell lines BxPC-3,
Capan-2, SW1990, PANC-1, and AsPC-1 was assessed by qPCR.
*P<0.05, compared with the HPDE6-C7 group. (C and D) Two
GATA3-AS1 siRNAs (si-GATA3-AS1-1 and si-GATA3-AS1-2) and negative
control siRNA (siRNA NC) were transfected into PANC-1 or AsPC-1
cells, respectively. Then, the expression of GATA3-AS1 was detected
by qPCR. *P<0.05, compared with the siRNA NC group. LncRNA, long
noncoding RNA; GATA3-AS1, GATA3-antisense RNA 1; PC, pancreatic
cancer; qPCR, quantitative polymerase chain reaction; siRNA or si-,
small interfering RNA; NC, negative control. |
Knockdown of GATA3-AS1 affects the
cell biology and stem cell-like characteristics of PC cells
As revealed in Fig. 2A and
B, the CCK-8 assay revealed that the OD (450 nm) in the
si-GATA3-AS1 group was significantly decreased compared with that
in the siRNA NC group, indicating that the cell viability of both
PANC-1 and AsPC-1 cells was reduced by the knockdown of GATA3-AS1.
Furthermore, compared with the siRNA NC group, the si-GATA3-AS1
group exhibited decreased proliferation ability (Fig. 2C), increased cell apoptosis (Fig. 2D), and reduced invasion ability
(Fig. 2E) in both PANC-1 and AsPC-1
cells. In addition, the spheroid formation ability in the
si-GATA3-AS1 group was also significantly reduced compared with the
siRNA NC group (Fig. 2F). Moreover,
the in vivo tumorigenicity experiment revealed that the
tumor volumes and weights of nude mice in the sh-GATA3-AS1 group
were significantly decreased compared with the nude mice in the
shRNA NC group (Fig. 2G).
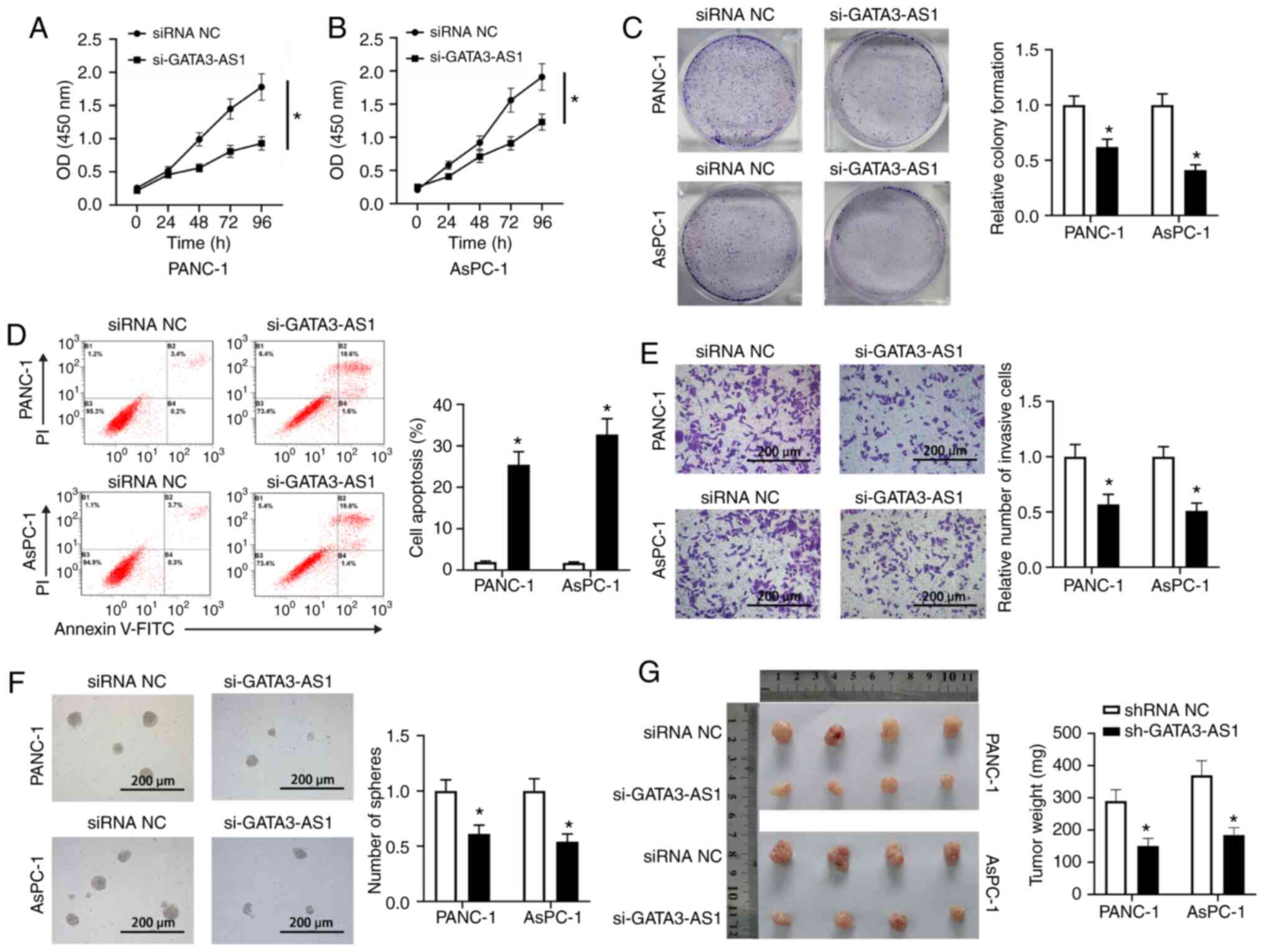 | Figure 2.Knockdown of GATA3-AS1 affects the
cell biology and stem cell-like characteristics of PC cells. (A and
B) Cell Counting Kit-8 assay at 0, 24, 48, 72 and 96 h post
transfection. (C) Cell proliferation of PANC-1 and AsPC-1 cells was
assessed by colony formation assay. (D) Cell apoptosis of PANC-1
and AsPC-1 cells was assessed by flow cytometry. (E) Cell invasion
of PANC-1 and AsPC-1 cells was assessed by Transwell assay. (F) The
stemness of PANC-1 and AsPC-1 cells was assessed by spheroid
formation assay. *P<0.05, compared with the siRNA NC group. (G)
The PANC-1 or AsPC-1 cells infected with lentivirus-shRNA NC or
lentivirus-sh-GATA3-AS1 were subcutaneously injected into the
axilla of nude mice, respectively. Two weeks later, the primary
tumors were weighed. *P<0.05, compared with the shRNA NC group.
GATA3-AS1, GATA3-antisense RNA 1; PC, pancreatic cancer; siRNA or
si-, small interfering RNA; shRNA or sh-, short hairpin RNA; NC,
negative control. |
GATA3-AS1 acts as a ceRNA of
miR-30b-5p
The bioinformatics analysis revealed that GATA3-AS1
contained a miR-30b-5p binding site (Fig.
3D). The expression of miR-30b-5p in the PC tissues and cell
lines was then assessed. As revealed in Fig. 3A, the expression of miR-30b-5p in the
cancer tissues from patients with PC was significantly
downregulated, compared with the normal tissues. In addition, as
revealed in Fig. 3B, compared with
the HPDE6-C7 cells, the expression of GATA3-AS1 in BxPC-3, Capan-2,
SW1990, PANC-1, and AsPC-1 cell lines was also significantly
downregulated. A negative correlation was revealed between the
expression of GATA3-AS1 and miR-30b-5p in the PC tissues (Fig. 3C). Furthermore, the dual-luciferase
reporter assay revealed that, in comparison with the mimic-NC
group, miR-30b-5p overexpression significantly reduced the firefly
luciferase activity of the pGL3-GATA3-AS1 wt plasmids, but it had
no effect on the firefly luciferase activity of pGL3-GATA3-AS1 mut
plasmids in either PANC-1 or AsPC-1 cells (Fig. 3E and F). Moreover, as revealed in
Fig. 3G, compared with the mimic NC
group, the expression of GATA3-AS1 had no obvious difference when
miR-30b-5p was overexpressed in both PANC-1 and AsPC-1 cells. In
addition, compared with the siRNA NC group, the expression of
miR-30b-5p was significantly upregulated in the si-GATA3-AS1 group;
compared with the pcDNA 3.1 group, the expression of miR-30b-5p was
significantly downregulated in the GATA3-AS1 overexpression group
(Fig. 3H and I).
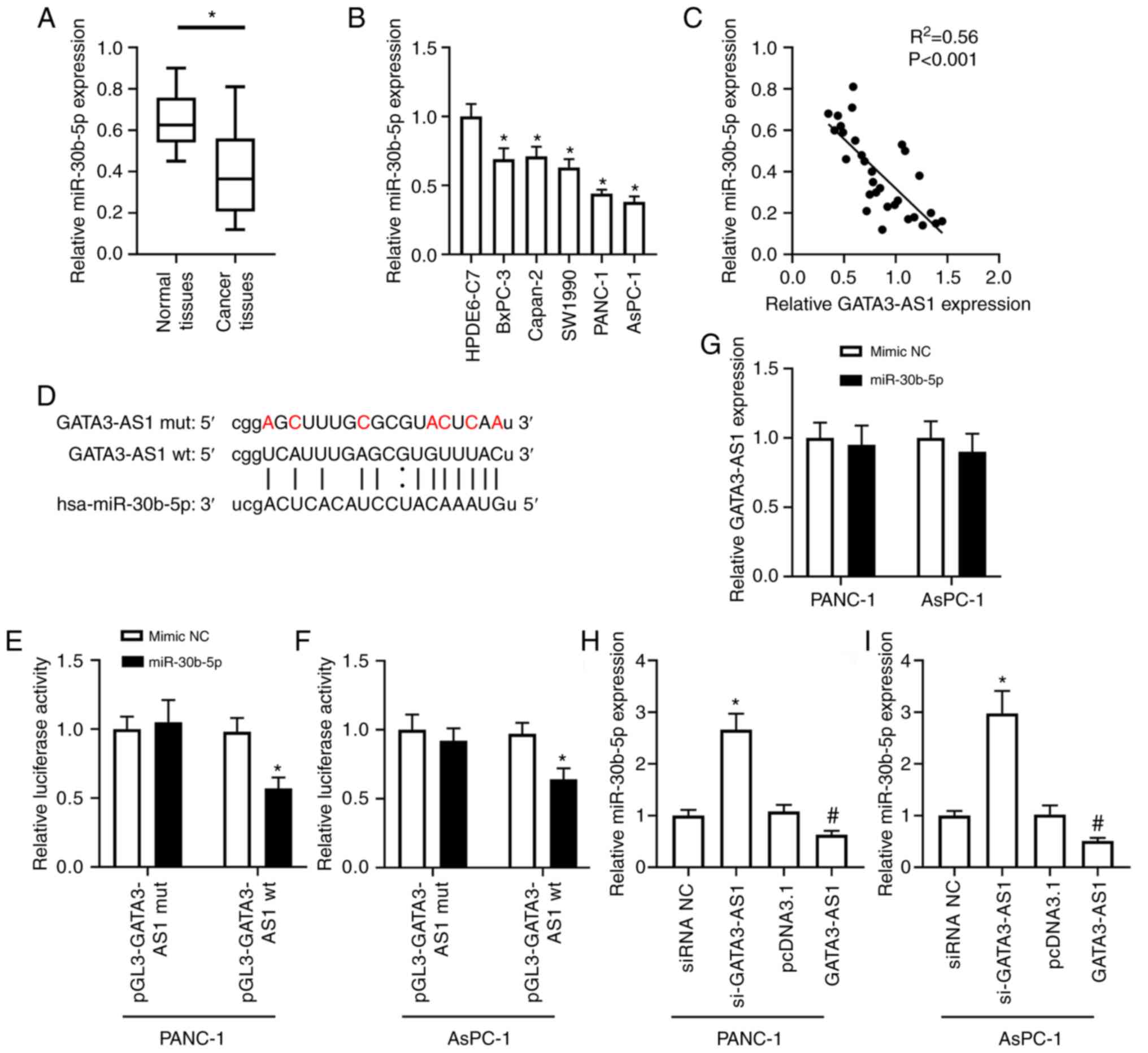 | Figure 3.GATA3-AS1 acts as a ceRNA of
miR-30b-5p. (A) The expression of miR-30b-5p in 30 pairs of PC
tissues and matched normal tissues was assessed by qPCR.
*P<0.05, compared with the normal tissues. (B) The expression of
miR-30b-5p in HPDE6-C7, BxPC-3, Capan-2, SW1990, PANC-1, and AsPC-1
cell lines was assessed by qPCR. *P<0.05, compared with the
HPDE6-C7 group. (C) The negative correlation between the expression
of GATA3-AS1 and miR-30b-5p in PC tissues. (D) The wild-type and
mutant sequences of the binding site between GATA3-AS1 and
miR-30b-5p. (E and F) The target relationship between GATA3-AS1 and
miR-30b-5p was confirmed by dual-luciferase reporter assay in
PANC-1 and AsPC-1 cells. *P<0.05, compared with the mimic-NC
group. (G) PANC-1 and AsPC-1 cells were transfected with mimic NC
or miR-30b-5p mimic. Then, the expression of GATA3-AS1 was assessed
by qPCR. (H and I) PANC-1 and AsPC-1 cells were transfected with
si-GATA3-AS1 or pcDNA 3.1-GATA3-AS1 plasmids to knockdown or
overexpress GATA3-AS1. Then, the expression of miR-30b-5p was
assessed by qPCR. *P<0.05, compared with the siRNA NC group;
#P<0.05, compared with the pcDNA 3.1 group.
GATA3-AS1, GATA3-antisense RNA 1; ceRNA; competing endogenous RNA;
miR-30b-5p, microRNA-30-5p; PC, pancreatic cancer; qPCR,
quantitative polymerase chain reaction; siRNA or si-, small
interfering RNA; NC, negative control; wt, wild-type; mut,
mutant. |
miR-30b-5p directly targets the 3′-UTR
of Tex10 to suppress its expression
The bioinformatics analysis revealed that Tex10 also
contained a miR-30b-5p binding site (Fig.
4E). The expression of Tex10 was then assessed in the PC
tissues and cell lines. As revealed in Fig. 4A, compared with the normal tissues,
the mRNA expression of Tex10 in the cancer tissues from patients
with PC was significantly upregulated. In addition, as revealed in
Fig. 4B, compared with the HPDE6-C7
cells, the mRNA expression of Tex10 in BxPC-3, Capan-2, SW1990,
PANC-1, and AsPC-1 cell lines was also significantly upregulated. A
positive correlation was revealed between the expression of
GATA3-AS1 and Tex10 (Fig. 4C), and a
negative correlation between the expression of miR-30b-5p and Tex10
(Fig. 4D) in the PC tissues.
Furthermore, the dual-luciferase reporter assay revealed that
compared with the mimic-NC group, miR-30b-5p overexpression
significantly reduced the firefly luciferase activity of the
pGL3-Tex10 wt plasmids, but it had no effect on the firefly
luciferase activity of pGL3-Tex10 mut plasmids in either PANC-1 or
AsPC-1 cells (Fig. 4F and G).
Moreover, as revealed in Fig. 4H and
I, compared with the mimic NC group, the mRNA and protein
expression of Tex10 in the miR-30b-5p overexpression group were
markedly downregulated in both PANC-1 and AsPC-1 cells. In
addition, compared with the siRNA NC group, the mRNA and protein
expression of Tex10 were significantly downregulated in the
si-GATA3-AS1 group; compared with the pcDNA 3.1 group, the
expression of Tex10 was significantly upregulated in the GATA3-AS1
overexpression group in the PANC-1 cells (Fig. 4J and K). Similar results were observed
in the AsPC-1 cells (Fig. 4L and
M).
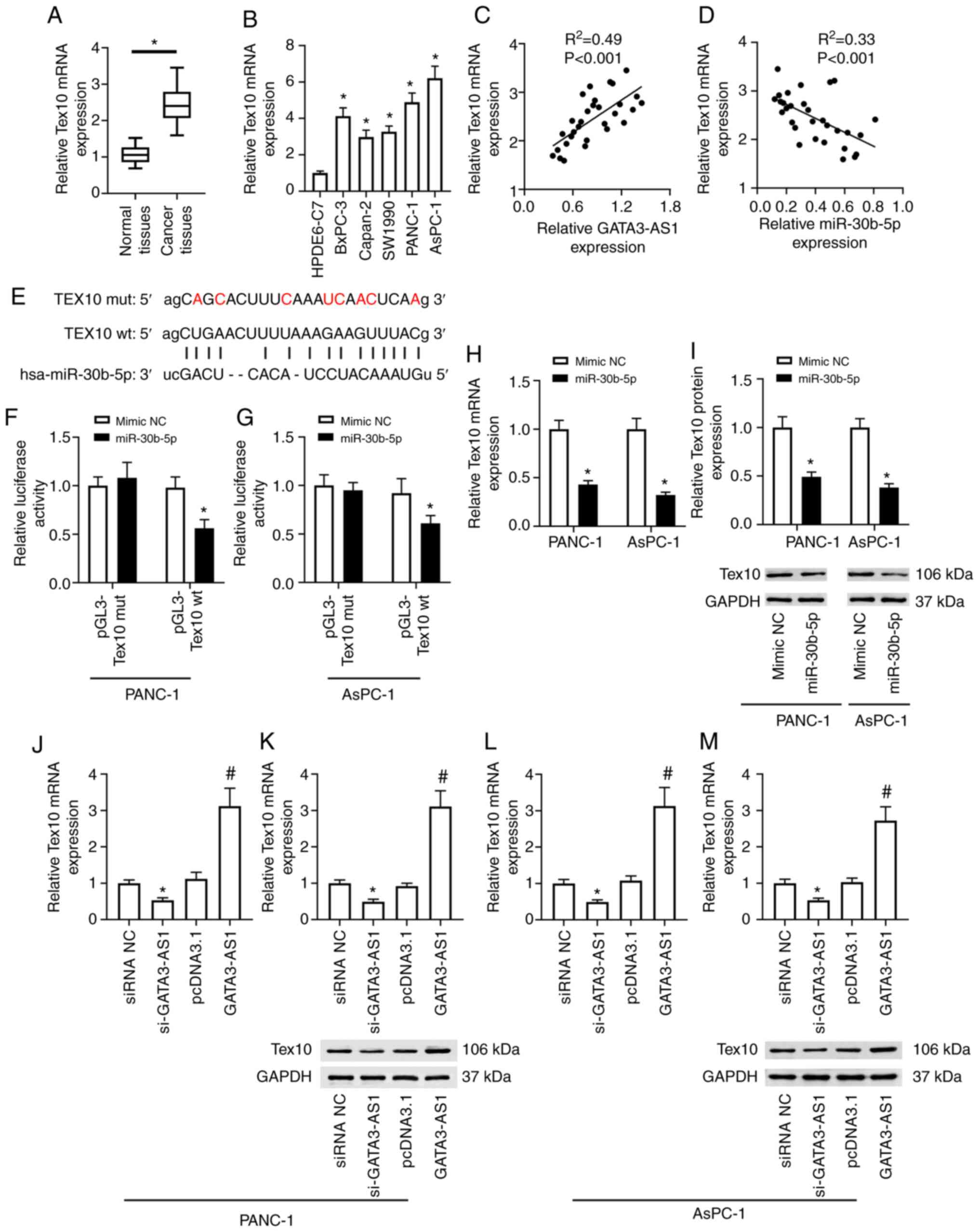 | Figure 4.miR-30b-5p directly targets the
3′-UTR of Tex10 to suppress its expression. (A) The mRNA expression
of Tex10 in 30 pairs of PC tissues and matched normal tissues was
assessed by qPCR. *P<0.05, compared with the normal tissues. (B)
The mRNA expression of Tex10 in HPDE6-C7, BxPC-3, Capan-2, SW1990,
PANC-1, and AsPC-1 cell lines was assessed by qPCR. *P<0.05,
compared with the HPDE6-C7 group. (C) The positive correlation
between the expression of GATA3-AS1 and Tex10 in PC tissues. (D)
The negative correlation between the expression of miR-30b-5p and
Tex10 in PC tissues. (E) The wild-type and mutant sequences of the
binding site between miR-30b-5p and Tex10. (F and G) The target
relationship between miR-30b-5p and Tex10 was confirmed by
dual-luciferase reporter assay in PANC-1 and AsPC-1 cells.
*P<0.05, compared with the mimic-NC group. (H and I) PANC-1 and
AsPC-1 cells were transfected with mimic NC or miR-30b-5p mimic.
The mRNA and protein expression levels of Tex10 were assessed by
qPCR and western blotting. *P<0.05, compared with the mimic-NC
group. PANC-1 and AsPC-1 cells were transfected with si-GATA3-AS1
or pcDNA 3.1-GATA3-AS1 plasmids. (J and K) The mRNA and protein
expression levels of Tex10 in PANC-1 cells were assessed by qPCR
and western blotting. (L and M) The mRNA and protein expression
levels of Tex10 in AsPC-1 cells were assessed by qPCR and western
blotting. *P<0.05, compared with the siRNA NC group,
#P<0.05, compared with the pcDNA 3.1 group.
miR-30b-5p, microRNA-30-5p; Tex10, testis-expressed protein 10; PC,
pancreatic cancer; qPCR, quantitative polymerase chain reaction;
GATA3-AS1, GATA3-antisense RNA 1; siRNA or si-, small interfering
RNA; NC, negative control; wt, wild-type; mut, mutant. |
miR-30b-5p-Tex10 axis partially
mediates the effects of GATA3-AS1
To determine whether the miR-30b-5p-Tex10 axis was
involved in the regulation of GATA3-AS1 bioactivities, pcDNA 3.1
empty plasmids, pcDNA 3.1-GATA3-AS1 plasmids, a combination of
pcDNA 3.1-GATA3-AS1 plasmids and miR-30b-5p mimic, or a combination
of pcDNA 3.1-GATA3-AS1 plasmids, miR-30b-5p mimic and
pcDNA3.1-Tex10 plasmids were transfected into AsPC-1 cells. As
revealed in Fig. 5A, compared with
the GATA3-AS1 group, the upregulated protein expression of Tex10
was significantly suppressed in the GATA3-AS1 + miR-30b-5p group.
However, this was reversed and it was upregulated again in the
GATA3-AS1 + miR-30b-5p + Tex10 group. The cell viability, cell
proliferation, cell apoptosis and invasion, and stem cell
characteristics were further assessed. Compared with the pcDNA 3.1
group, increased cell viability and proliferation, invasion, and
spheroid formation abilities were observed in the GATA3-AS1 group;
all of these were suppressed in the GATA3-AS1 + miR-30b-5p group.
Furthermore, compared with the GATA3-AS1 + miR-30b-5p group, the
cell viability and proliferation, invasion, and spheroid formation
abilities once more exhibited a rising trend, and cell apoptosis
exhibited a decreasing trend in the GATA3-AS1 + miR-30b-5p + Tex10
group (Fig. 5B-F). In addition,
compared with the pcDNA 3.1 group, the protein expression levels of
Wnt1 and β-catenin were upregulated in the GATA3-AS1 group;
compared with the GATA3-AS1 group, the protein expression levels of
Wnt1 and β-catenin were downregulated in the GATA3-AS1 + miR-30b-5p
group but upregulated in the GATA3-AS1 + miR-30b-5p + Tex10 group
(Fig. 5G and H).
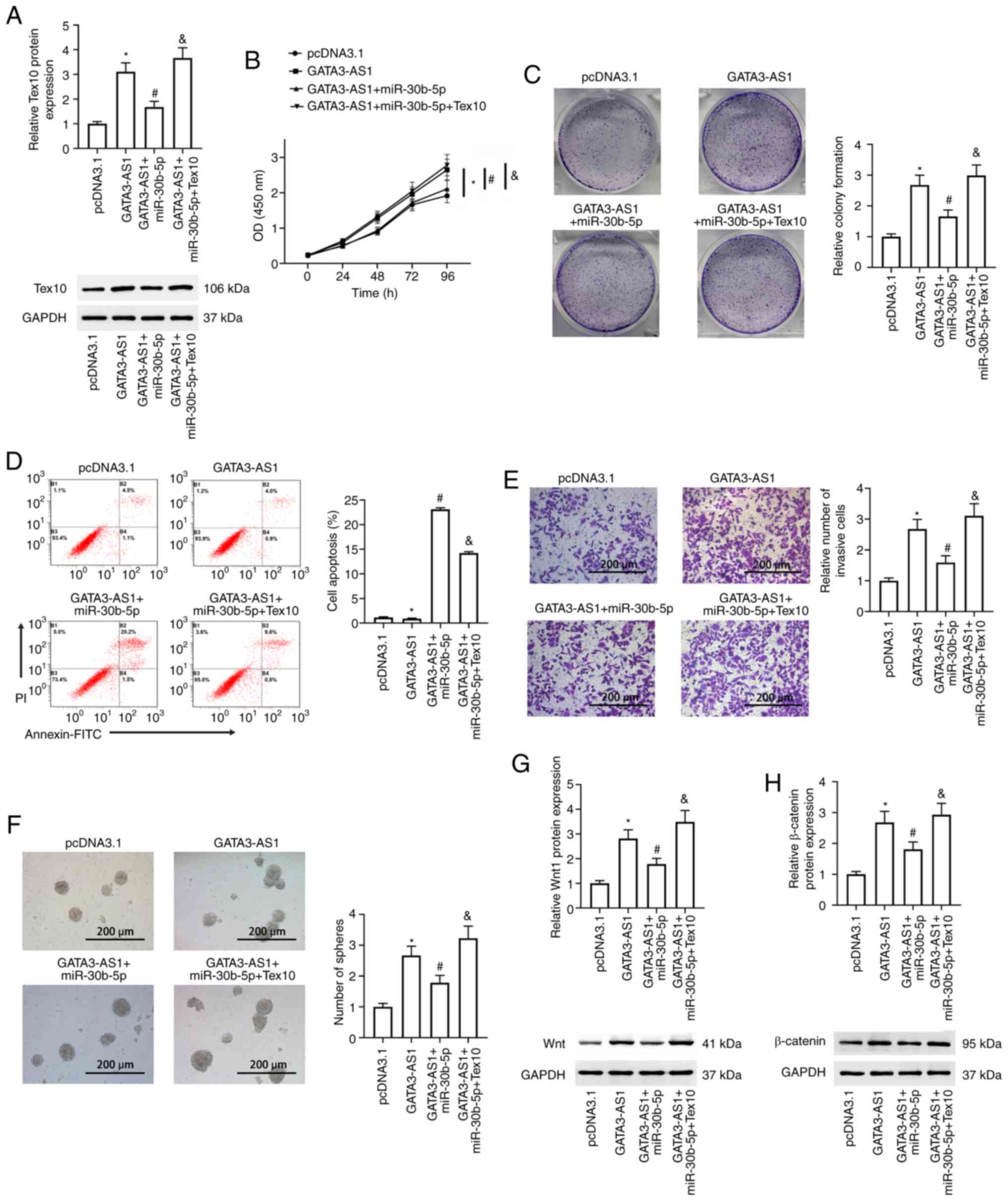 | Figure 5.miR-30b-5p-Tex10 axis partially
mediates the effects of GATA3-AS1. AsPC-1 cells were transfected
with pcDNA 3.1 empty plasmids, pcDNA 3.1-GATA3-AS1 plasmids, a
combination of pcDNA 3.1-GATA3-AS1 plasmids and miR-30b-5p mimic,
or a combination of pcDNA 3.1-GATA3-AS1 plasmids, miR-30b-5p mimic,
and pcDNA3.1-Tex10 plasmids. (A) The protein expression of Tex10
was assessed by western blotting. (B) Cell viability was assessed
by Cell Counting Kit-8 assay at 0, 24, 48, 72 and 96 h post
transfection. (C) Cell proliferation was assessed by colony
formation assay. (D) Cell apoptosis was assessed by flow cytometry.
(E) Cell invasion was assessed by Transwell assay. (F) The stemness
was assessed by spheroid formation assay. The protein expression of
(G) Wnt1 and (H) β-catenin was assessed by western blotting.
*P<0.05, compared with the pcDNA 3.1 group;
#P<0.05, compared with the GATA3-AS1 group;
&P<0.05, compared with the GATA3-AS1 + miR-30b-5p
group. miR-30b-5p, microRNA-30-5p; Tex10, testis-expressed protein
10; GATA3-AS1, GATA3-antisense RNA 1. |
Discussion
During the past few decades, it has become widely
recognized that the noncoding portion of the mammalian
transcriptome is more complex than the protein-coding genes. Among
these ncRNAs, 18% of lncRNAs have been revealed to be associated
with the progression of cancers, compared with 9% of protein-coding
genes (16,17), indicating that lncRNAs could have a
major role in modulating carcinogenesis. Some well-defined lncRNAs
have been recognized as oncogenes or suppressors in various cancers
(7), such as H19 (18,19),
HOTAIR (20,21), MALAT1 (22,23), UCA1
(24,25), and MEG3 (26,27). In
the present study, it was demonstrated that GATA3-AS1 may function
as an oncogene in the progression of PC.
GATA3-AS1 is a newly discovered antisense lncRNA,
which was first identified from human CD4+ T-cell
subsets (28). Subsequently, Zhu
et al (29) revealed that
GATA3-AS1 was overexpressed in human bladder cancer tissues.
Recently, Luo et al (13)
revealed that GATA3-AS1 was significantly upregulated in HCC
tissues. In the present study, GATA3-AS1 expression in the PC
tissues and cell lines was significantly upregulated, indicating
that the aberrant expression may contribute to the progression of
PC, similar to its oncogenic roles in HCC. To better understand the
biological roles of GATA3-AS1 in PC, in vitro
loss-of-function experiments were performed. Knockdown of GATA3-AS1
in PANC-1 and AsPC-1 cells obviously inhibited cell viability,
proliferation, and invasion and increased cell apoptosis.
Furthermore, knockdown of GATA3-AS1 in PANC-1 and AsPC-1 cells
resulted in reduced stemness, which was characterized by suppressed
spheroid formation ability. Moreover, decreased volumes and weights
of primary tumors by GATA3-AS1 knockdown were revealed in the in
vivo tumorigenicity experiment. Collectively, these data
demonstrated that GATA3-AS1 also acts as an oncogene to accelerate
the progression of PC.
Accumulating evidence has demonstrated that lncRNAs
act as endogenous ceRNAs for specific miRNAs and thus regulate gene
expression. For instance, Gong et al (30) have revealed that UCA1 interacts with
miR-203, resulting in the release of miR-203-targeted transcripts
ZEB2, and thus promotes metastatic gastric cancer. Xu et al
(31) have shown that MALAT1
facilitates colorectal cancer cell proliferation, invasion, and
migration by sponging the miR-145-SOX9 regulatory axis. Chang et
al (32) have shown that HOTAIR
regulates CCND1 and CCND2 expression by sponging miR-206 to
stimulate the proliferation and invasion of ovarian cancer cells.
Ding et al (33) have shown
that the H19-miR-29b-3p-PGRN axis promotes epithelial-mesenchymal
transition of colorectal cancer cells. In short, the previous
studies collectively indicated that the lncRNA-miRNA-mRNA axis is
an important regulatory mechanism in cancer progression. In order
to gain insights into the underlying regulatory mechanism of
GATA3-AS1, noteworthy miR-30b-5p, which has a binding site with
GATA3-AS1, was predicted by bioinformatics analysis. Past studies
have demonstrated that the miR-30 family plays a crucial regulatory
role in the pathogenesis of different diseases, which indicates it
may be a promising regulator (34,35).
miR-30b was reported to be downregulated in PC, serving as a novel
therapeutic agent for the treatment of PC (36,37). In
the present study, consistent with the aforementioned result,
miR-30b expression was significantly decreased in PC tissues and
cell lines. Furthermore, a negative correlation between the
expression of GATA3-AS1 and miR-30b-5p in the PC tissues was
revealed, implying that GATA3-AS1 could directly bind to miR-30b-5p
to sponge it. This implication was confirmed by dual-luciferase
reporter assay. These data indicated that GATA3-AS1 acts as a ceRNA
of miR-30b-5p in PC cells.
Notable Tex10, which shares a binding site with
miR-30b-5p, was further predicted by bioinformatics analysis.
Tex10, a component of the five friends of methylated Chtop and Rix
complexes, is deeply involved in transcriptional regulation and
ribosome biogenesis, as well as the cell cycle (38,39). Ding
et al (40) identified Tex10
as an evolutionarily conserved key pluripotency factor that is
required for embryonic stem cell self-renewal and pluripotency.
Recently, Xiang et al (41,42)
demonstrated that Tex10 is upregulated and plays a potent
carcinogenic role in both HCC and esophageal squamous cell
carcinoma. In the present study, similar to the aforementioned
result, Tex10 was significantly upregulated in PC tissues and cell
lines. The direct target relationship between miR-30b-5p and Tex10
was confirmed by dual-luciferase reporter assay. Furthermore, a
positive correlation between the expression of GATA3-AS1 and Tex10
and a negative correlation between the expression of miR-30b-5p and
Tex10 was revealed in PC tissues. In addition, GATA3-AS1 knockdown
upregulated the expression of miR-30b-5p, but downregulated the
expression of Tex10, whereas GATA3-AS1 overexpression had the
opposite effects. Collectively, these data indicated that GATA3-AS1
acts as a ceRNA of miR-30b-5p, leading to the release of
miR-30b-5p-targeted transcript Tex10 in PC cells.
The function of the GATA3-AS1/miR-30b-5p/Tex10 axis
involved in PC was further addressed. The increased cell viability
and proliferation, cell invasion, and stemness induced by GATA3-AS1
overexpression were all suppressed by the co-overexpression of
GATA3-AS1 and miR-30b-5p; whereas the cell survival revealed an
increasing trend with co-overexpression of GATA3-AS1, miR-30b-5p,
and Tex10. These data indicated that the biological roles of
GATA3-AS1 in PC are at least in part mediated by the
miR-30b-5p-Tex10 axis. The aberrant activation of Wnt/β-catenin
signaling has been reported to contribute to carcinogenesis and
tumor progression of several cancers, including PC (43,44).
Furthermore, the effects of miR-30b-5p and Tex10 have been revealed
to be associated with Wnt/β-catenin signaling (42,45).
Therefore, it was further assessed whether Wnt/β-catenin signaling
participates in the regulation of the GATA3-AS1/miR-30b-5p/Tex10
axis. It was revealed that the upregulated expression of Wnt1 and
β-catenin induced by GATA3-AS1 overexpression was suppressed by the
co-overexpression of GATA3-AS1 and miR-30b-5p, but was upregulated
once more by the co-overexpression of GATA3-AS1, miR-30b-5p, and
Tex10. These results indicated that the GATA3-AS1/miR-30b-5p/Tex10
axis modulated Wnt/β-catenin signaling in PC cells.
In conclusion, the present findings demonstrated a
significant increase of GATA3-AS1 expression in both PC tissues and
cell lines. In vitro loss- or gain-of-function experiments
revealed that the GATA3-AS1-miR-30b-5p-Tex10 axis modulated cell
proliferation, invasion, apoptosis, and stemness in PC, which may
be associated with the Wnt/β-catenin signaling pathway. The present
study may provide some novel insights into identifying an
informative biomarker and treatment targets for PC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available from the corresponding author upon
reasonable request.
Authors' contributions
YL and LL conceived and designed the study. YL and
GX performed the experiments, and analyzed the data. YL wrote the
paper. YL, GX and LL reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All patients were informed before their inclusion;
written consent of the patients was obtained. All experimental
protocols were approved by the Ethics Committee of The First
Affiliated Hospital of Xi'an Jiaotong University (Xi'an, China).
Animal protocols were approved by the Animal Care and Use Committee
of the First Affiliated Hospital of Xi'an Jiaotong University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing of
interests.
References
|
1
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Henriksen A, Dyhl-Polk A, Chen I and
Nielsen D: Checkpoint inhibitors in pancreatic cancer. Cancer Treat
Rev. 78:17–30. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kopp F and Mendell JT: Functional
classification and experimental dissection of long noncoding RNAs.
Cell. 172:393–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jarroux J, Morillon A and Pinskaya M:
History, discovery, and classification of lncRNAs. Adv Exp Med
Biol. 1008:1–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Z, Wang P, Zhang L, Huang C, Gao J,
Li Y and Yang B: Identification of key genes and long noncoding
RNA-associated competing endogenous RNA (ceRNA) networks in
early-onset preeclampsia. Biomed Res Int.
2020:16734862020.PubMed/NCBI
|
|
9
|
Vishnoi A and Rani S: MiRNA Biogenesis and
regulation of diseases: An overview. Methods Mol Biol. 1509:1–10.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gibbons HR, Shaginurova G, Kim LC, Chapman
N, Spurlock CF III and Aune TM: Divergent lncRNA GATA3-AS1
regulates GATA3 transcription in T-Helper 2 cells. Front Immunol.
9:25122018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gulbinas A, Berberat PO, Dambrauskas Z,
Giese T, Giese N, Autschbach F, Kleeff J, Meuer S, Büchler MW and
Friess H: Aberrant gata-3 expression in human pancreatic cancer. J
Histochem Cytochem. 54:161–169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo X, Zhou N, Wang L, Zeng Q and Tang H:
Long noncoding RNA GATA3-AS1 promotes cell proliferation and
metastasis in hepatocellular carcinoma by suppression of PTEN,
CDKN1A, and TP53. Can J Gastroenterol Hepatol. 2019:13896532019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42((Database issue)): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Khachane AN and Harrison PM: Mining
mammalian transcript data for functional long non-coding RNAs. PLoS
One. 5:e103162010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei W, Liu Y, Lu Y, Yang B and Tang L:
LncRNA XIST promotes pancreatic cancer proliferation through
miR-133a/EGFR. J Cell Biochem. 118:3349–3358. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kallen AN, Zhou XB, Xu J, Qiao C, Ma J,
Yan L, Lu L, Liu C, Yi JS, Zhang H, et al: The imprinted H19 lncRNA
antagonizes let-7 microRNAs. Mol Cell. 52:101–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ren J, Fu J, Ma T, Yan B, Gao R, An Z and
Wang D: LncRNA H19-elevated LIN28B promotes lung cancer progression
through sequestering miR-196b. Cell Cycle. 17:1372–1380. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang Q and Hann SS: HOTAIR: An oncogenic
long non-coding RNA in human cancer. Cell Physiol Biochem.
47:893–913. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chang YT, Lin TP, Tang JT, Campbell M, Luo
YL, Lu SY, Yang CP, Cheng TY, Chang CH, Liu TT, et al: HOTAIR is a
REST-regulated lncRNA that promotes neuroendocrine differentiation
in castration resistant prostate cancer. Cancer Lett. 433:43–52.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim J, Piao HL, Kim BJ, Yao F, Han Z, Wang
Y, Xiao Z, Siverly AN, Lawhon SE, Ton BN, et al: Long noncoding RNA
MALAT1 suppresses breast cancer metastasis. Nat Genet.
50:1705–1715. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li S, Mei Z, Hu HB and Zhang X: The lncRNA
MALAT1 contributes to non-small cell lung cancer development via
modulating miR-124/STAT3 axis. J Cell Physiol. 233:6679–6688. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang H, Guan Z, He K, Qian J, Cao J and
Teng L: LncRNA UCA1 in anti-cancer drug resistance. Oncotarget.
8:64638–64650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yao F, Wang Q and Wu Q: The prognostic
value and mechanisms of lncRNA UCA1 in human cancer. Cancer Manag
Res. 11:7685–7696. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Uroda T, Anastasakou E, Rossi A, Teulon
JM, Pellequer JL, Annibale P, Pessey O, Inga A, Chillón I and
Marcia M: Conserved pseudoknots in lncRNA MEG3 are essential for
stimulation of the p53 pathway. Mol Cell. 75:982–995.e9. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang W, Xie Y, Chen F, Liu X, Zhong LL,
Wang HQ and Li QC: LncRNA MEG3 acts a biomarker and regulates cell
functions by targeting ADAR1 in colorectal cancer. World J
Gastroenterol. 25:3972–3984. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang H, Nestor CE, Zhao S, Lentini A,
Bohle B, Benson M and Wang H: Profiling of human CD4+
T-cell subsets identifies the TH2-specific noncoding RNA GATA3-AS1.
J Allergy Clin Immunol. 132:1005–1008. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu YP, Bian XJ, Ye DW, Yao XD, Zhang SL,
Dai B, Zhang HL and Shen YJ: Long noncoding RNA expression
signatures of bladder cancer revealed by microarray. Oncol Lett.
7:1197–1202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gong P, Qiao F, Wu H, Cui H, Li Y, Zheng
Y, Zhou M and Fan H: LncRNA UCA1 promotes tumor metastasis by
inducing miR-203/ZEB2 axis in gastric cancer. Cell Death Dis.
9:11582018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu Y, Zhang X, Hu X, Zhou W, Zhang P,
Zhang J, Yang S and Liu Y: The effects of lncRNA MALAT1 on
proliferation, invasion and migration in colorectal cancer through
regulating SOX9. Mol Med. 24:522018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang L, Guo R, Yuan Z, Shi H and Zhang D:
LncRNA HOTAIR Regulates CCND1 and CCND2 expression by sponging
miR-206 in ovarian cancer. Cell Physiol Biochem. 49:1289–1303.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ding D, Li C, Zhao T, Li D, Yang L and
Zhang B: LncRNA H19/miR-29b-3p/PGRN axis promoted
epithelial-mesenchymal transition of colorectal cancer cells by
acting on wnt signaling. Mol Cells. 41:423–435. 2018.PubMed/NCBI
|
|
34
|
Mao L, Liu S, Hu L, Jia L, Wang H, Guo M,
Chen C, Liu Y and Xu L: MiR-30 family: A promising regulator in
development and disease. Biomed Res Int. 2018:96234122018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang SJ, Yang SY, Wang DD, Chen X, Shen
HY, Zhang XH, Zhong SL, Tang JH and Zhao JH: The miR-30 family:
Versatile players in breast cancer. Tumour Biol.
39:10104283176922042017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiong Y, Wang Y, Wang L, Huang Y, Xu Y, Xu
L, Guo Y, Lu J, Li X, Zhu M and Qian H: MicroRNA-30b targets Snail
to impede epithelial-mesenchymal transition in pancreatic cancer
stem cells. J Cancer. 9:2147–2159. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Azmi AS, Li Y, Aboukameel A, Muqbil I,
Philip PA and Mohammad RM: DNA-methylation-caused downregulation of
miR-30 contributes to the high expression of XPO1 and the
aggressive growth of tumors in pancreatic ductal adenocarcinoma.
Cancers (Basel). 11:11012019. View Article : Google Scholar
|
|
38
|
Fanis P, Gillemans N, Aghajanirefah A,
Pourfarzad F, Demmers J, Esteghamat F, Vadlamudi RK, Grosveld F,
Philipsen S and van Dijk TB: Five friends of methylated chromatin
target of protein-arginine-methyltransferase[prmt]-1 (chtop), a
complex linking arginine methylation to desumoylation. Mol Cell
Proteomics. 11:1263–1273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Castle CD, Cassimere EK and Denicourt C:
LAS1L interacts with the mammalian Rix1 complex to regulate
ribosome biogenesis. Mol Biol Cell. 23:716–728. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ding J, Huang X, Shao N, Zhou H, Lee DF,
Faiola F, Fidalgo M, Guallar D, Saunders A, Shliaha PV, et al:
Tex10 coordinates epigenetic control of super-enhancer activity in
pluripotency and reprogramming. Cell Stem Cell. 16:653–668. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiang X, Deng L, Xiong R, Xiao D, Chen Z,
Yang F, Liu K and Feng G: Tex10 is upregulated and promotes cancer
stem cell properties and chemoresistance in hepatocellular
carcinoma. Cell Cycle. 17:1310–1318. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xiang X, Xiong R, Yu C, Deng L, Bie J,
Xiao D, Chen Z, Zhou Y, Li X, Liu K and Feng G: Tex10 promotes
stemness and EMT phenotypes in esophageal squamous cell carcinoma
via the Wnt/β-catenin pathway. Oncol Rep. 42:2600–2610.
2019.PubMed/NCBI
|
|
43
|
Shang S, Hua F and Hu ZW: The regulation
of beta-catenin activity and function in cancer: Therapeutic
opportunities. Oncotarget. 8:33972–33989. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ram Makena M, Gatla H, Verlekar D,
Sukhavasi S, K Pandey M and C Pramanik K: Wnt/β-Catenin signaling:
The culprit in pancreatic carcinogenesis and therapeutic
resistance. Int J Mol Sci. 20:42422019. View Article : Google Scholar
|
|
45
|
Jiang S, Miao D, Wang M, Lv J, Wang Y and
Tong J: MiR-30-5p suppresses cell chemoresistance and stemness in
colorectal cancer through USP22/Wnt/β-catenin signaling axis. J
Cell Mol Med. 23:630–640. 2019. View Article : Google Scholar : PubMed/NCBI
|














