High hydrostatic pressure (HHP) is a traditional
technology used to produce steel, alloys, ceramics, and synthetic
materials (1). Over the past few
decades, HHP has also been used for non-heat pasteurization of
processed foods, designed to extend the storage time of foods, such
as juice, milk, and canned products (2). As researchers in different scientific
fields continue to explore HHP, some new applications of the
technology are emerging (3,4). Since most practical applications of HHP
technology are subjecting biological systems to hydrostatic
pressure (HP), the uniqueness of this method is currently being
investigated at various levels, ranging from viruses, enzymes,
microorganisms, mammalian cells, and tissues (5–7). Usually,
all pressures are causing a reduction in the volume of the system,
which can lead to changes in its structure and disturb the
equilibrium of chemical reactions (8). Excessive pressure may lead to the
destruction of cell structure (9).
For numerous thermophilic microorganisms, HP inhibits cell growth
in the range of tens of megapascal (MPa) and completely inhibits
cell growth at approximately 50 MPa (10). Pressure greater than 200 MPa can
annihilate most microorganisms (11).
Cancer poses a huge threat to human health.
Currently, the main cancer treatments are surgery, radiation
therapy (RT), and chemotherapy (CT). Multimodal treatment
strategies may be effective in reducing tumor size, regressing
solid tumors, and extending patient life (12). However, the recurrence and metastasis
of tumors gives rise to a poor prognosis for numerous patients
(13). Therefore, the goal of
oncology treatment is not only to exterminate tumor cells of the
primary origin but also to obtain long-lasting antitumor effects in
order to control metastatic and recurrent tumor cells. However,
these treatments currently leave much to be desired in terms of
providing sustained antitumor effects and limiting tumor metastasis
and recurrence. In addition to these disadvantages, these
treatments have numerous toxic side effects that affect normal
tissues (14). Combining the
antitumor effects of the body with the host immune system to
generate an effective antitumor immune response is an attractive
therapeutic approach.
Tumor vaccines are designed to specifically activate
the immune system of patients (15).
Therefore, it is necessary to activate the immune response of
patients to the tumor. The immune system must be trained to control
dormancy and metastasis of residual tumor cells (16). Tumor vaccines may bypass the complex
processes of defining individual antigens. There are numerous
associated antigens on the surface and inside the tumor cells that
prevent tumors from escaping immunity (17). To enhance the antitumor immune
response, additional application of immune adjuvants is beneficial
(18). The combination of
conventional therapies with immunotherapy may improve the overall
patient survival. In addition, immunotherapy may be more
appropriate for oncology patients because of its lower toxicity
compared to CT.
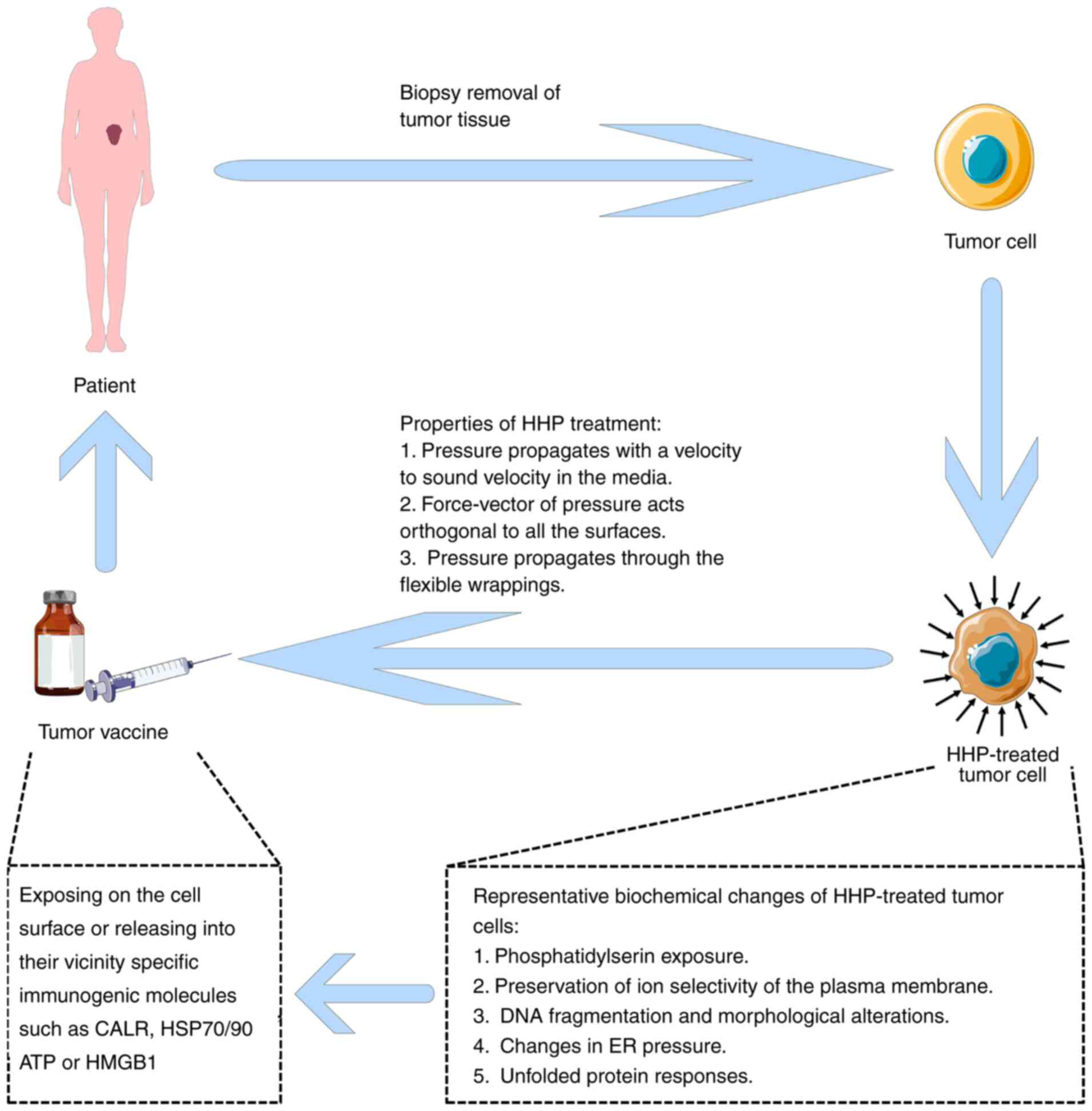
HHP technology is an effective approach to the
production of tumor vaccines. Helmstein reported the application of
HHP for the treatment of bladder cancer in 1972 (18). The patients were treated with
hydrostatic bladder dilation. Subsequently, some authors reported
that vaccination of HHP-treated tumor cells treated with the
chemical cross-linker adenosine dialdehyde alone or in combination
with the reducing agent N-acetyl-L-cysteine induced antitumor
immunity in vivo (19) and
in vitro (20). In this
review, we summarized the latest knowledge on the relevance of
hydrostatic hypertension for immunotherapy of biomolecules and
tumors and discussed possible future directions for the development
of HHP tumor vaccines.
The physical property of pressure is defined as the
force per unit area acting on the surface in a direction
perpendicular to the surface: P=F/A in which P represents the
pressure, F represents the normal force applied to the surface, and
A represents the area of the surface. The official unit of pressure
is the Pascal (Pa) (1 Pa=1 N/1 m2=10−5 bar).
The Newton represents a small force, while 1 m2
corresponds to a large surface, thus the Pascal is a very small
unit of pressure. Therefore, MPa (1 MPa=106 Pa=10 atm)
is a common unit of pressure used in HP research. The conversion
from MPa to other units of pressure is presented in Table I (21).
The first research on HP traces back to the late
19th century and was carried out mainly by oceanographers and
physiologists (22). HP exists in all
the explored biological environments. The pressure extends from 0.1
MPa (atmospheric pressure) at the sea level to 110 MPa at the
deepest part of the ocean in the Mariana Trench, 11 km below sea
level (23). The average depth of the
ocean is 3.8 km. The average pressure on various marine organisms
is approximately 38 MPa, which is 380 times the atmospheric
pressure (24).
Over the past few decades, a growing number of
disciplines have begun to explore the potential of exposing a
variety of complex biological units to HHP, including proteins,
lipids, nucleic acids, eukaryotic cells, and multicellular tissues
(41). In general, all pressure
effects correspond to a reduction in the volume of the biological
unit and the acquisition of a more compact structure (8). The aforementioned will be discussed in
more detail in the later sections.
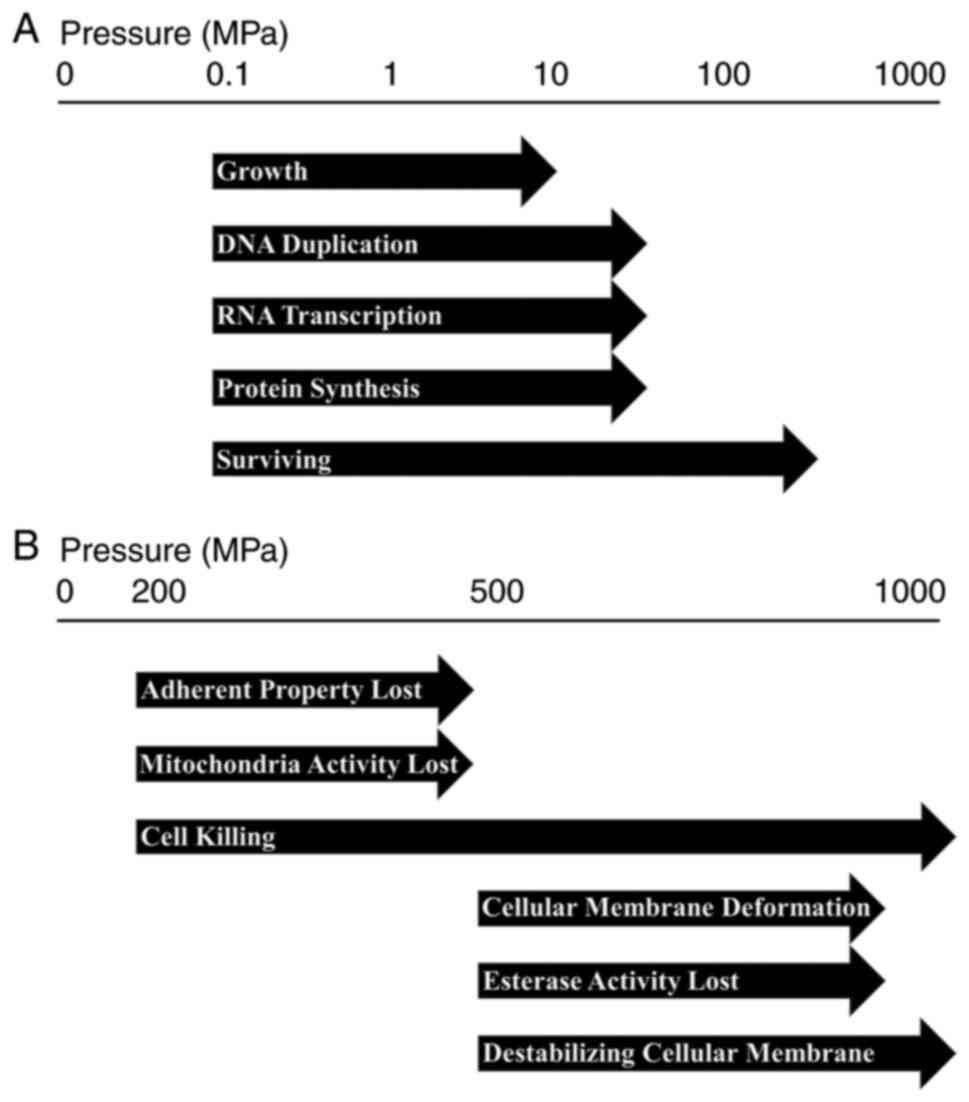
HP is one of the physical factors that affect
cellular physiology. Inappropriate pressure can lead to inhibition
of cell growth, structural destruction of cells, and cell death.
HHP between 1 and 100 MPa is considered non-lethal, which leads to
reversible morphological changes and slight stress response. HHP
between 100 and 150 MPa can induce the apoptosis of murine cells.
HHP between 150 and 250 MPa can affect the viability of human
cells, while pressures between 300 and 400 MPa can lead to cell
necrosis (41,42). During HHP treatment inactivation, the
pressure is instantaneously and uniformly distributed throughout
the non-toxic media and can be transmitted through all flexible
packaging materials. Each part of the treated sample is subjected
to the same pressure simultaneously (43). Finally, each treated cell in the
system is exactly subjected to the same stress and extremely high
reproducibility can be achieved (44). Pressure will penetrate the cell
instantly and completely, thus applying it to the entire
intracellular components (44). It is
assumed that exposing cells to pressures above a certain threshold
will result in a progressive increase in membrane rigidity and
protein denaturation, which will eventually lead to cell death
(43) (Fig.
1).
Some studies have reported apoptosis as well as
necrosis of cells after non-physiological HHP exposure, with the
specific mechanism of cell death mainly depended on the sensitivity
of the cell type and the pressure level (45,46). Cell
death occurs through apoptosis at pressures of approximately 200
MPa (47), and cell necrosis occurs
at pressures >300 MPa (23,24,47).
Liu et al used HHP to inactivate B16-F10 melanoma cells at
different pressures (≥50 MPa) and for different durations (≥1 min)
(48). Their results suggested that
HHP may be an effective measure for the preparation of melanoma
vaccines when the pressure was ≥200 MPa and the treatment duration
was ≥30 min. Seitz et al demonstrated that in vitro
treatment at 200 MPa or higher completely inhibited the formation
of tumor cell colonies and that HHP produced inactivated tumor
cells that could be used as a tumor vaccine (49). Similarly, they demonstrated synergy
between tumor cell-based vaccines and RT, significantly hindering
tumor growth by generating a favorable antitumor immune
microenvironment.
HHP-induced apoptosis occurs through the activation
of caspase-3 by extrinsic and intrinsic pathways. The extrinsic
pathway is characterized by the binding of the Fas ligands to the
cell death receptor Fas on the cell surface (50). Cytochrome c is frequently
released from mitochondria into the cytoplasm when the intrinsic
pathway is activated (47). Apoptosis
leads to cell death through cell shrinkage, loss of microvilli, and
chromatin condensation (47). The
clearance of apoptotic cells is mediated by ‘find-me’ signals
released by apoptotic cells to facilitate the clearance of
apoptotic cells by phagocytes (51).
Phagocytes recognize the ‘eat me’ signals on the surface of
apoptotic cells and rapidly clear them. Clearance of apoptotic
cells stimulates activated phagocytes, which secrete
anti-inflammatory signals such as transforming growth factor-β
(TGF-β) and interleukin-10 (IL-10) (52). However, it has been revealed that
apoptosis may also exhibit immune-stimulatory features under
certain circumstances, especially when treated with γ-irradiation
or certain CT drugs (anthracyclines) (53). Cell necrosis occurs in HHP above 300
MPa (24). The onset of cellular
necrosis is not dependent on the activation of caspases. Cellular
necrosis leads to cell swelling, organelle degradation, especially
irreversible mitochondrial damage, and changes in intracellular
ionic concentrations. These changes ultimately lead to the damage
of cell membranes and the release of inflammatory cellular
inclusions (47). However, it is not
entirely clear to what extent the molecular nature of the danger
signals of passive exposure of necrotic tumor cells overlaps with
immunogenic apoptosis.
Apoptotic cells in the physiological state are
immune-silent or tolerogenic. They are part of the physiological
processes that maintain homeostasis in multicellular organisms
(54). Apoptosis is characterized by
some cellular morphological and biochemical features, such as
blistering, chromatin condensation, and DNA fragmentation (55). In contrast to apoptosis, cell necrosis
is associated with inflammation, which is controlled by
pathological processes (56).
Extracellular high mobility group box 1 (HMGB1) and heat shock
proteins (HSPs) are typical examples of such released immune
activator proteins (57). In
addition, apoptotic and necrotic cells can also release some other
danger signals (58). The loss of
cell membrane integrity leads to the release of danger signals,
which can lead to the activation and maturation of immune cells and
often generate inflammation (58). It
must be taken into consideration that in the case of apoptosis, the
danger signals are modified before the secretion, resulting in the
opposite immunological outcome (59).
For instance, HMGB1 is generally oxidized by reactive oxygen
species (ROS) during apoptosis and thus loses its immune potency
(60). This suggests that dying cells
and their microenvironment determine whether the immune activation
or the immunosuppression is induced.
The forms of cellular death are manifold and
sometimes indistinguishable. Some studies have also focused on the
cell death pattern after HHP treatment. Dead cells are potent
modulators of the immune system, and the immunogenicity of the
treated cells depends largely on the death-inducing stimulus
(61,62). In the syngeneic condition, live and
normal apoptotic cells do not drive immune responses or lead to
anti-inflammation (52). In stark
contrast, necrotic cells induce immune responses (63). However, inactivation processes that
induce necrosis in vitro usually do not exhibit favorable
immunogenic responses in vivo. The immunogenicity of treated
cells is highly dependent on the death-initiating stimulus and must
be analyzed separately (61).
Finally, the treatment processes must be carried out in highly
repeatable manners and meet all legal requirements.
Pressure, similar to temperature, is an important
thermodynamic parameter that can profoundly affect molecular
systems (64). HHP represents a
distinctive form of stress associated with volume changes that
alter numerous physiological and biochemical processes, such as
protein unfolding and dissociation, lipid bilayer phase transition,
and ligand binding (65,66). Pressure studies on biomolecules are
generally performed between 0.1 and 1,500 MPa (23). In cells, HHP mainly affects the
non-covalent bonds of biomacromolecules, such as proteins and
lipids (23). The HHP of
approximately 200 MPa mainly alters the tertiary and quaternary
structure of proteins, which leads to disruption of the enzymatic
function. Under this pressure, the primary structures of proteins
can remain intact. When the HHP is >400 MPa, the protein is
rapidly denatured (67). HHP
transforms the conformation of phospholipid bilayers from
fluid-crystalline type to gel-like type (68). Mitochondrial activity is associated
with important functions for cell growth. For example, the
polymerization of actin filaments and the conversion of adenosine
triphosphate (ATP) (69). Therefore,
HHP treatment of approximately 200 MPa will induce cell killing by
inactivating mitochondrial activity (7). Furthermore, DNA may be resistant to
non-physiological HHP at approximately 1,000 MPa because its
secondary structure is mainly stabilized by H-bonds, which are
practically insensitive to HHP in biotechnology (70). Only HHP >1,000 MPa can induce the
transition from double-stranded DNA to single-stranded DNA. The
effects of different pressure levels on cells and biomacromolecules
are presented in Fig. 2.
The absorption of amino acids is also sensitive to
HHP. Effects of HHP on amino acid uptake is attributed to the
impairment of amino acid permeases and transport proteins at the
cell surfaces (75). Generally
speaking, 200 MPa HHP treatment completely induced cell killing by
inactivating the enzymatic activity in mitochondria (76). As is well known, the mitochondria are
associated with the polymerization of actin filaments and the
supply of cellular energy (77).
Significant advances have been made in our knowledge of cellular
piezoelectric physiology (78,79). Given
its effect on proteins, it is not surprising to reveal that
sublethal HHP can induce the expression of HSPs in both prokaryotic
and eukaryotic cells (80).
The ribosomes are decomposed by HHP due to the
decomposition of polymorphs caused by pressure (70S → 30S + 50S)
(81,82). The dissociation of uncharged ribosomes
is accompanied by a large negative volume change. The dissociation
of ribosomes appears to be one of the main factors leading to cell
death by HHP treatment, as cells cannot survive when the quantity
of functional ribosomes drops below a threshold (81).
Lipid membranes are the most pressure-sensitive
biological components due to their high compressibility and
fluidity (83). The lipid components
of biological membranes have usually been visualized as a physical
homogeneous system. However, there is growing evidence that lipid
domains of different compositions and structures may exist in
biological membranes even at the physiological temperature
(84,85). Changes in pressure will lead to
changes in the structures and sizes of these domains as well as in
the functional state of the membrane. The low pressures can change
the local composition of biological membrane lipid domains, which
have significant biochemical implications (86). In general, the maintenance of basic
physiological functions of biological membranes requires the
dynamic and structural properties of cell membranes. Manisegaran
et al reported that HHP and low temperature have a strong
influence on lipid biological membranes, mainly because they both
affect the fluidity of biological membranes to a large extent,
leading to tight packing and limiting acyl-chain movement (86).
The compression of the phospholipid bilayer is
usually anisotropic. Under HHP conditions, the acyl-chain
straightening leads to lateral contracting and increased thickness.
This phenomenon is also accompanied by a phase transition from the
liquid crystal to the gel phase (87). As pressure increases, lipid bilayers
lose fluidity and rapidly become impermeable to water and other
molecules, while protein-lipid interactions essential for optimal
functions of the biological membranes become weaker (87). In addition to altering fluidity, HHP
also changes the compositions of lipid membranes by increasing the
content of unsaturated fatty acids. This corresponds to an adaptive
mechanism of increased fluidity, which is thought to be because
unsaturated fatty acids require less carbon and energy to achieve
similar results in membrane fluidity compared to saturated fatty
acids (88).
Due to the sensitivity of lipids to HHP, these
biological components are usually considered as the main targets of
cellular and microbial stress inactivation (87). Membrane potential also decreases with
increasing pressure until 400 MPa (89). These results support that the
properties of cellular membranes are related to the cell-killing
activity of the treatment. However, cell membrane damage induced by
HHP may not be the critical factor for the cell killing, as 400–500
MPa of pressure is required to induce the damage of the cell
membranes, but 200 MPa of pressure is sufficient to kill cells.
Pressure treatment decreases the enzymatic and metabolic activity
(90). It is considered that deep-sea
creatures have evolved special membranes and membrane proteins to
adapt to such extreme circumstances. A variety of organisms can
compensate for the packing effects of biological membranes by
modifying compositions of fatty acids (86). Cold adaptation is usually associated
with the binding of the unsaturated bonds within the acyl chains
(91). Acyl chains on biological
membranes containing unsaturated bonds have larger conformations
than their saturated counterparts, thus allowing greater
conformational freedom and less packing of the biological membrane.
Consequently, the biological membrane becomes more fluid. Sinensky
reported that this adaptation by creatures living in cold and
high-pressure environments has been known as homeoviscous
adaptation (92). Whether
homeoviscous adaptation universally occurs in deep-sea creatures
remains unknown, as the properties of natural biological membranes
responding to HHP have not been fully studied.
HHP may change the fluidity of membranes and
indirectly affect the binding or conformation of signaling
molecules (93). HHP can also change
forces within the membranes by increasing the bending rigidity to
produce biological forces sufficient to trigger the
mechano-chemical responses (94). The
direct lethal effects of HHP may include biological membrane damage
and other undefined fast-acting responses, and ROS production due
to biological membrane damage may continue even after the
treatment. Atmospheric oxygen may promote ROS production by
oxidases located in the damaged biological membrane of HHP-treated
cells. HHP can trigger metabolic disorders that generate ROS in
treated cells, which can eventually damage their viability
(95).
Typically, HHP below 1,000 MPa cannot affect DNA,
while pressure treatments above 1,000 MPa may result in DNA
alterations (24). As stress
increases, double-stranded DNA becomes more stable, so processes
such as replication, transcription, and translation become more
difficult. The transition from double-stranded to single-stranded
is essential for cell survival (96).
However, Macgregor revealed that the clear transition from the
double-stranded DNA to the single-stranded DNA can be observed
below 1,000 MPa (96).
Cancer immunotherapy (CI), particularly when used in
combination with other therapies such as RT and CT, is a promising
avenue to cancer treatment. Shi et al have demonstrated the
synergistic effects of the combination of CI and CT in the clinical
study (103). CI stands out in the
second-line treatment for recurrent tumors and metastases by
activating the immune systems of patients to trigger an antitumor
response (103). Since tumor
recurrence and metastasis remain the main reasons for tumor-related
deaths, the identification of tumor cell specificity and
persistence will be the focus of future research. To date, the
number and type of specific T cells required for the efficient
antitumor therapy are unknown (104). Although different subtypes of immune
cells are suspected to have various effects on tumor progression,
the infiltration of immune cells is usually associated with the
prognosis of most solid tumors (105,106).
Therefore, CI is a suitable adjuvant to standard tumor therapies
because it is designed to activate the immune system of the patient
against tumor cells (107).
When developing multimodal concepts for tumor
therapy, the treatment methods of tumor vaccines should be
considered (108). Vaccination is an
agent that causes the host to receive treated autologous tumor
cells in order for a large number of defined tumor antigens to
simultaneously stimulate the immune system of the host (109). HHP inactivation of tumor cells can
be performed in a highly repeatable manner according to Good
Manufacturing Practices (GMP) and legal requirements. These
vaccines must also fulfill the major requirements for all
cell-based therapeutic tumor vaccines, including i) complete
inactivation of tumor cells, ii) maintenance of immunogenicity, and
iii) compliance with statutory provisions (110). Physical (X-ray and freeze-thawing)
and chemical methods have been used to inactivate tumor cells in
vaccination experiments (46).
However, these methods usually have some restrictions. Tumor cells
cannot be inactivated safely by a mild treatment, or the
inactivated cells are weakly immunogenic (46). HHP fulfills these specifications for
clinical vaccine: It inactivates tumor cells effectively, is
non-toxic, does not wreck the immunogenicity of the tumor cells,
and can comply with GMP and legal requirements. In additiom, it is
a repeatable and easy to apply method (111). Therefore, HHP treatment is superior
to other methods, such as freeze-thaw, radiation, or heat killing
methods. Furthermore, the HHP approach cannot mix other chemicals
into tumor vaccines, unlike the chemical methods (49). These advantages suggest that HHP is a
promising method for generating tumor vaccines (112).
The use of HHP in tumor inactivation and tumor
vaccine development has been studied since the 1970s (113). In 1972, HHP was reported as the
tumor treatment method and was investigated for the treatment of
bladder cancer by using the hydrostatic bladder dilatation method
(18). Later, Deckmann et al
reported that treatment of leukemic cells with 150 MPa HHP resulted
in enhanced immunogenicity (114).
Since then, HHP has been used to inactivate tumor cells to develop
vaccines. Eisenthal et al treated tumor cells with HHP (120
MPa) in the presence of the biocompatible crosslinker, adenosine
dialdehyde (115). Treated cells
were potent immunogens because HHP increases the antigenic
presentation by rearranging the cell surface proteins into
immunogenic clusters (118). This
study indicated that HHP-killed tumor cells can trigger antitumor
immune responses. This syngeneic tumor vaccine that mimics the
autologous vaccine from their tumor cells should contain all
relevant tumor-associated antigens (TAAs) that may target a
specific patient (116).
The purpose of tumor vaccines is to train the immune
system to actively develop lasting immune memory to fight the
metastasis and recurrence of tumors (117). Weiss et al concluded that the
inactivation of intact tumor cells induced by HHP, the degradation
of the nucleus, and the preservation of the immunogenic potential
of these dead tumor cells facilitate the use of this technology for
the production of tumor vaccines (46). Frey et al have also revealed
that HHP-treated tumor cells can preserve their shapes for more
than a few weeks, which appears to be important for the production
of potent vaccines (24). HHP
treatment also caused a marked increase in cytoplasmic viscosity of
the treated cells, which would lead to a slow and sustained release
of cell-derived antigens and danger signals (45). These features are critical for the
phagocytosis of tumor cells and subsequent presentation of cognate
antigens by dendritic cells (DCs), and further expand the prospects
of applying HHP technology to produce tumor vaccines (118).
Notably, HHP-induced cell necrosis occurs
simultaneously with cytoplasmic gelation, and these cellular
particles appear to maintain the relevant immunogenicity (42). Frey et al have proposed the use
of HHP-treated tumor cells as the whole-cell-based tumor vaccines,
due to the preservation of the antigenicity (24). Moserova et al revealed that HHP
treatment could induce immunogenic cell death in tumor cell lines
and revealed some molecular mechanisms associated with this
phenomenon (119). Apoptosis induced
by HHP treatment was controlled by the overproduction of ROS, which
caused a rapid establishment of an integrated stress response and
subsequent activation of caspase-2, caspase-3, and caspase-8
activation in dying tumor cells (119). Traditionally, apoptosis is
considered immunologically silent, but specific immunogenic
molecules, such as calreticulin, adenosine-triphosphate (ATP), HSP
70/90, or HMGB1, are released at or near the cell surfaces, thereby
activating immune cells to enhance their antitumor activity
(119,120).
Processing of tumor cells by HHP promotes the
production of ROS. The processing of cells by pressure triggers
downstream signaling pathways, such as cleavage of caspase-2,
caspase-3, and caspase-8 (121).
Moserova et al reported the application of various ROS
scavengers and indicated that for HHP-induced calreticulin, the
production of ROS was one of the prerequisites (119). More specifically, HHP-mediated ROS
production may affect endoplasmic reticulum (ER) homeostasis,
further triggered the phosphorylation of extension initiation
factor (eIF)-2α and the cleavage of caspase-2, which was important
for HHP-induced danger signaling involving calreticulin induction
(122). Sandow et al have
revealed that ER stress and ROS production may or may not lead to
caspase-2 cleavage, depending on the prevailing environment
(123). In addition, specific
antibody blockade of calreticulin or depletion of caspase-2
significantly inhibited DC phagocytosis (119). While it remained unclear how
caspase-2 regulated the exposure of calreticulin, the localization
of this caspase in the ER and Golgi systems suggested the possible
involvement in regulating the transit mechanisms (124). However, inhibition of ROS production
was not sufficient to eliminate HHP-induced calreticulin exposure,
suggesting that ROS-independent mechanisms may also be involved in
this process. Therefore, exogenous calreticulin is important for
the phagocytosis of tumor cells and the induction of specific
immune responses in vaccine patients (125).
In previous studies, HHP technology was identified
as an inducer of antitumor immunity in a variety of tumor cell
lines (126–128). Due to its immunogenicity, this
physical modality has been standardized and validated for
incorporation into the manufacturing process of tumor immunotherapy
products (129). In numerous cases,
it was not possible to obtain autologous malignant cells from
patients due to the inoperability of the tumor. The number of
malignant cells in the biopsy or resected tumors was too small to
produce a vaccine for the repeated administration during the
immunotherapy. The HHP-killed tumor cell lines also retained an
abundance of specific or tumor-associated antigens, which
represented an advantage of this approach over numerous vaccines,
where the number of antigens is often limited (130). Autologous tumor cells are a suitable
source of antigens for vaccination. Providing an antigen array on
the cell surface reduces the risk of tumor immune escape and
eliminates the need to define individual antigens (131).
The advantages of vaccines based on autologous whole
tumor cells are that they do not have to prospectively identify
target antigens and can provide numerous TAAs that aberrantly
express autoantigens. In contrast to neoantigens, the latter should
only activate remaining low-affinity T cells and have to overcome
the self-tolerance (132). Several
additional methods have been developed to address the barrier, such
as the addition of adjuvants, repeated vaccination, or
co-stimulation (104). HHP-treated
whole-cell-tumor vaccines have the advantage of providing multiple
antigens and therefore lead to better results. This approach has
been demonstrated in clinical trials in multiple myeloma (133) or renal cell carcinoma (134). Therefore, the inactivated form of
whole tumor cells used as a vaccine, as well as the cell death
induced in the primary tumor by standard methods, is important for
triggering the effective antitumor immunity.
Large numbers of treated apoptotic tumor cells have
been shown to trigger an effective antitumor immune response in
mice (135). Galluzzi et al
reported two morphologically equivalent but immunologically
distinct subcategories of apoptosis, immunogenic and
non-immunogenic apoptosis, thus introducing the concept of
immunogenic cell death (ICD) (112).
ICD was primarily mediated by defined spatiotemporal release or
exposure of relevant danger signals or the damage-associated
molecular patterns (DAMPs), which could act as adjuvants or
associated danger signals to the innate immune system to trigger
host protective antitumor immunity (136). Recently, several DAMPs have been
associated with ICDs, where surface exposure of the ER-resident
chaperone calreticulin was one of the main checkpoints for
determining cellular immunogenicity (137). McDaniel et al reported that
tumor cell ICDs were characterized by induction of ER stress
response, production of ROS, and release of danger-related
molecules, such as calreticulin, HSP, HMGB1, or ATP (138). Several tumor chemotherapies and cell
physical death-inducing modalities have been described to induce
ICD of tumor cells (139).
DCs play a key role in the immune response because
they can capture antigens bound to the pattern recognition
receptors, process and present them to the naive T cells, thus
inducing the T-cell activation and thus creating an important
bridge between the innate and adaptive immune systems (143). DCs have been demonstrated to play a
crucial role in the induction of antitumor immune responses
(119). DC-based immunotherapy is
safe and induces antitumor immunity in patients with advanced
melanoma (144). De Sanctis et
al revealed that in an orthotopic mouse model of prostate
cancer, the experimental group demonstrated that DC-based vaccines
were as effective as CT in slowing tumor growth (145). Mikyskova et al demonstrated
that DC-based vaccines were a reasonable tool for treating human
prostate cancer (126). Hradilova
et al reported important preclinical data from phase I/II
clinical trials in non-small cell lung cancer (NSCLC) using
DC-based active cellular immunotherapy in combination with CT and
immune boosters for the treatment of NSCLC (127). Urbanova et al also reported
important preclinical data from an ongoing phase I/II clinical
trial in NSCLC involving the use of DC-based active cellular
immunotherapy to produce immunogenically-killed lung cancer cells
(130). These studies demonstrated
that a DC vaccine can lead to long-lasting tumor immunity, a
process that requires three steps (146). In the first step, DCs must obtain
the relevant TAAs. Secondly, DCs have to mature and induce T-cell
responses. In DC-based cellular immunotherapy, the enhanced ability
of DCs to co-stimulate naive T cells during maturation may be
important. The final step is to allow T cells to overcome the
immunosuppression of the primary solid tumor and enter the tumor
bed (146).
The success of DC-based tumor immunotherapy depends
on the range of TAAs presented by DCs and the ability of DCs to
produce cytokines such as IL-12p70 and provide the co-stimulation
to T cells (147). Immature DCs are
constantly migrating in the tissues and blood, scanning the
environment for danger signals or potential pathogens (148). These pathogens and signals can
activate innate immunity and interact with pattern recognition
receptors, purinergic receptors, and phagocytosis-related receptors
expressed by DCs and stimulate the presentation of tumor antigens
to T cells (148). Typically,
autologous DC-based vaccines use in vitro cultures of DCs
loaded with the tumor antigens and promote the maturation of the
DCs (149). Tumor cells are
phagocytosed after being recognized by DCs, which then undergo
antigenic processing. DCs must reach the mature stage to induce an
effective immune response because semi-mature DCs have tolerogenic
features (146). The maturation of
DCs is accompanied by a decrease in antigen assimilation and an
increase in migration capacity. DCs move to the lymph nodes (LN),
where the complex of peptide and MHC-class II is presented to the
antigen-specific T-cell receptor (TCR) on the naive CD4+
T-cell. This again indicates the enhanced DCs function in tumor
patients (150).
Tumor cells can be inactivated by different methods
and the choice of the optimal inactivation method is crucial for
the DC vaccine optimization (15).
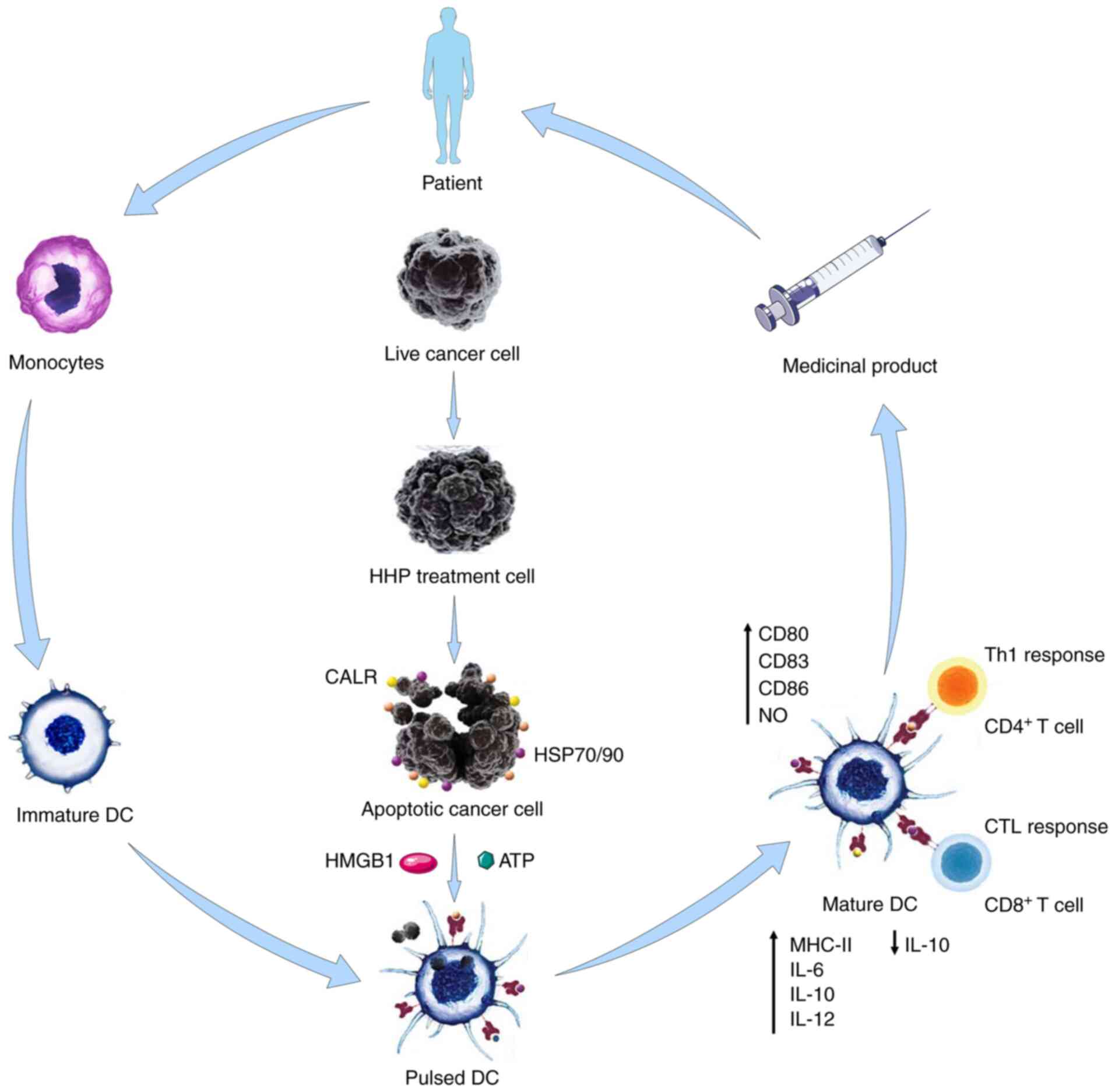
HHP treatment has been revealed to maintain immunogenic tumor cell
inactivation. HHP-treated tumor cells can induce the
monocyte-derived DC maturation. DCs cultured with HHP-treated tumor
cells can also induce the activation of T cells in vitro
(49). Phagocytosis of HHP-killed
tumor cells by DC stimulates the expression of
maturation-associated molecules on DCs and induces the production
of proinflammatory cytokines. The tendency of increased numbers of
the CD8+ T cells and the natural killer (NK) cells in
the spleens of the experimental animals are detected when DCs are
pulse-stimulated with HHP-treated tumor cells (126,130).
HHP treatment for tumor cells can be standardized
according to GMP requirements and incorporated into manufacturing
protocols for DC-based cellular CI (130). More importantly, DCs loaded with
HHP-treated tumor cells exhibited enhanced phagocytosis, expressed
high levels of co-stimulatory molecules, and stimulated a large
number of specific T lymphocytes, whereas no T regulatory cells
were induced in the absence of the additional immunostimulatory
agents (140). Mature DCs express
high levels of co-stimulatory molecules and peptide-bound MHC class
I and II molecules and produce pro-inflammatory cytokines, which
are essential for efficient stimulation of tumor antigen-specific
T-cell responses (152). These
interactions effectively activate DCs to phagocytose dying tumor
cells and allow them to mature and acquire an immunostimulatory
phenotype (152). A schematic
diagram of DC-based vaccine preparation using immunogenic
HHP-treated tumor cells, which can be applied to other physical
tumor cell death-inducing modalities is presented in Fig. 3.
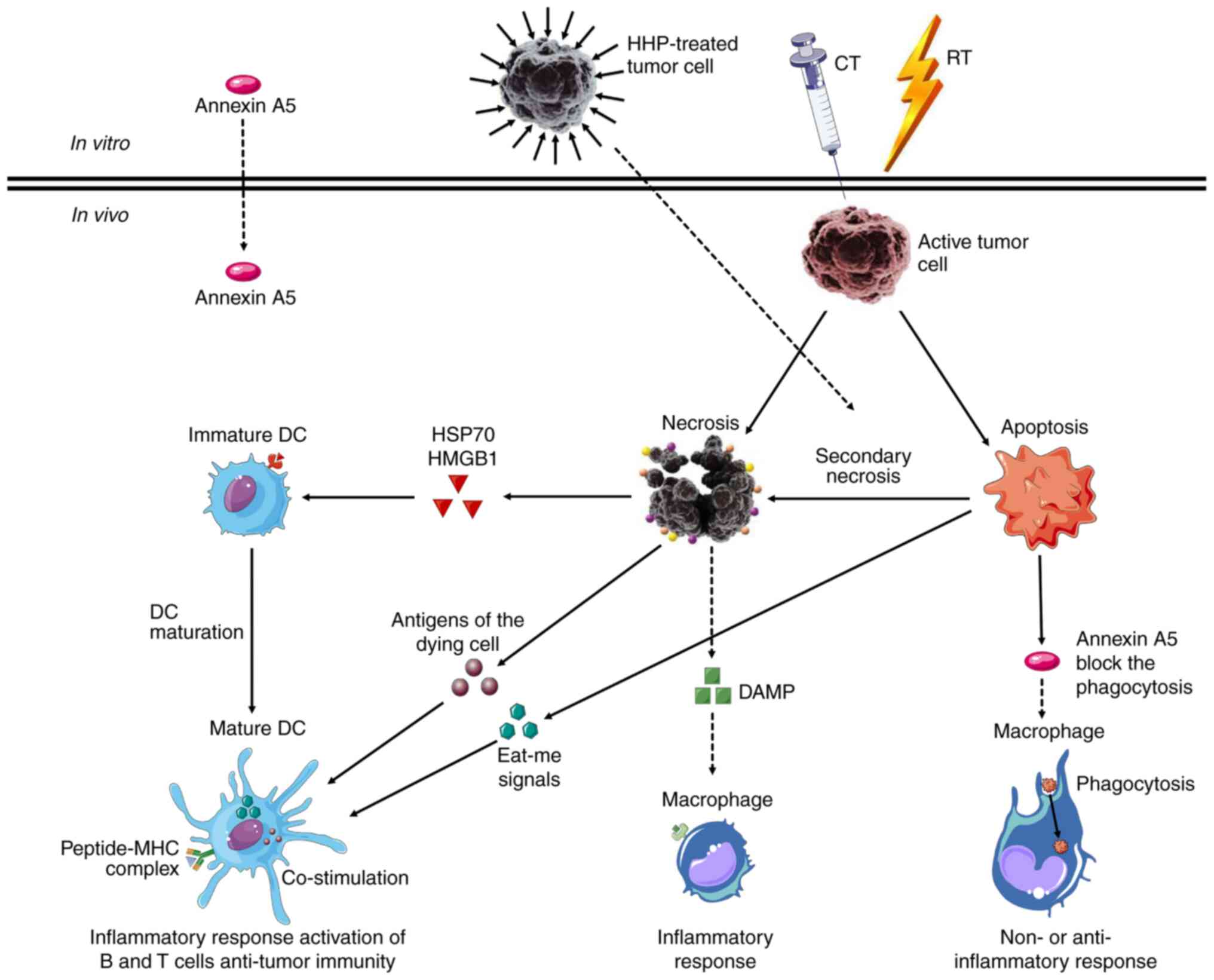
Phospholipids, such as lysophosphatidylcholine
(LPC) and phosphatidylserine (PS), are involved in the clearance of
apoptotic and necrotic cells. Apoptotic exposure of PS is one of
the main ‘eat me’ signals for macrophages (153). Annexin A5 (AnxA5), a high specific
ligand for PS, is an important modulator of immune responses
against the PS-exposed particles (154). AnxA5 is also the focus of
therapeutic applications for the delivery of drugs to the relevant
cells expressing PS on the cell surface (154). AnxA5 binds to phospholipids in a
Ca2+ dependent manner and it blocks the phagocytosis of
dying tumor cells by macrophages, but not DCs. Thus, the clearance
of dying tumor cells is transferred from macrophages to DCs
(155).
PS also expose the luminal surfaces of the vascular
endothelium of tumors. In animal models, antibody-targeting of PS
damages tumor vasculature and induces antitumor immune responses
(156). In addition in vivo,
the provided AnxA5 decreased the uptake of apoptotic cells by the
peritoneal macrophages and increased their uptake by DCs (157). In summary, AnxA5 promotes DC uptake
by interfering with the macrophage-mediated clearance of apoptotic
and necrotic tumor cells. In the presence of AnxA5, the
microenvironment becomes inflammatory and leads to the regression
of allogeneic tumors as well as the rejection and cure of the
syngeneic tumors. The influence of AnxA5 on clearance of apoptotic
tumor cells and antitumor immunity is presented in Table III.
In addition, DAMPs may also directly activate cells
of the innate immune systems, such as NK cells (163). Inhibition of apoptotic cell
clearance by macrophages with AnxA5, or induction of abundant
apoptotic cells in a multimodal treatment setting, can promote the
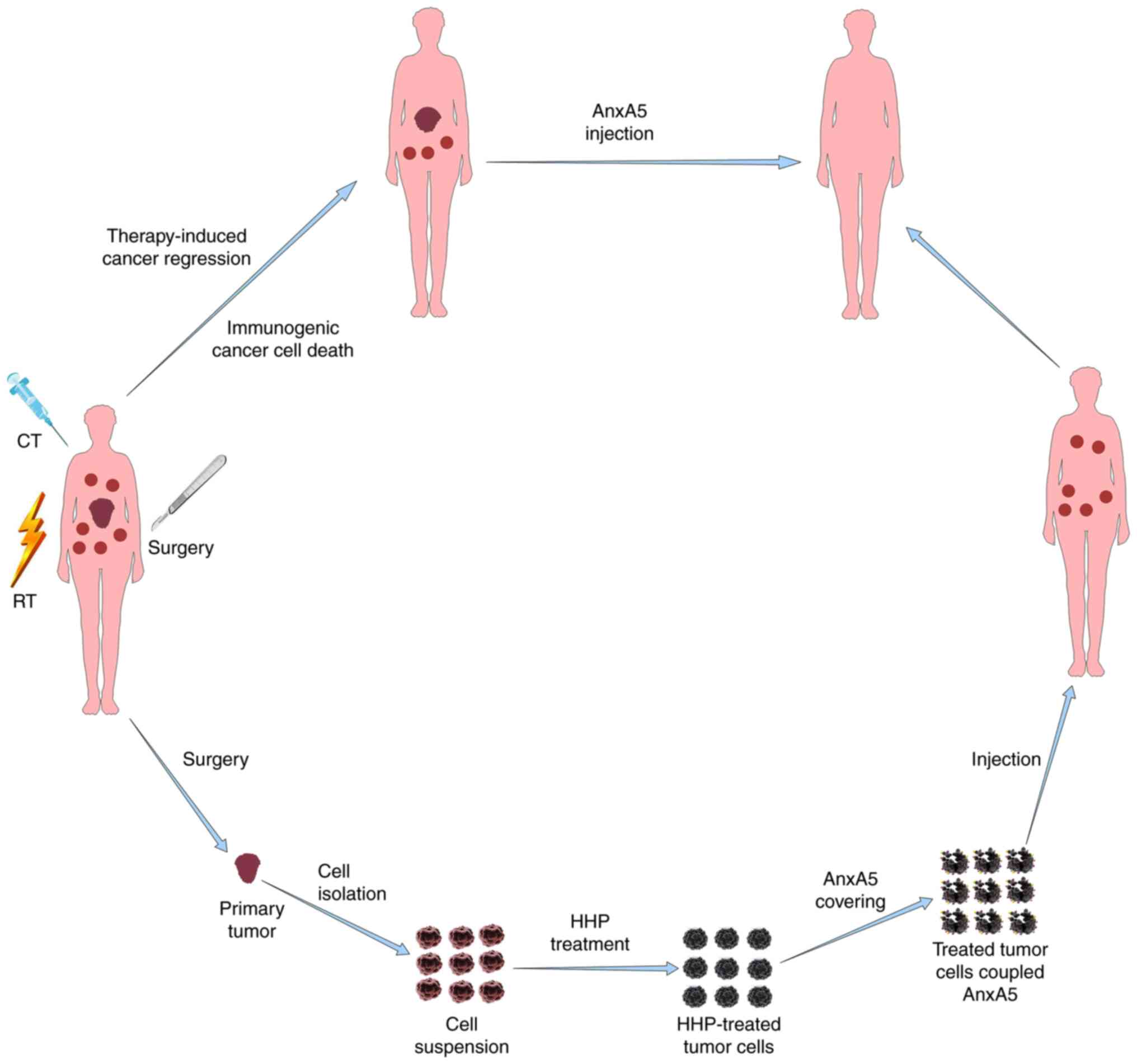
necrotic immune form of tumor cells (164). Immunogenic forms of tumor cell death
can also be induced by killing biopsy-derived fresh tumor cells
in vitro, resulting in complete cell death by increasing the
immunogenicity (165). Fig. 5 summarizes the principle of multimodal
treatments to induce tumor cell death leading to antitumor
immunity.
The purpose of this short review was to summarize
the knowledge and applications of HHP technology in the development
of tumor vaccines and to envisage new possible research directions
and applications. HHP technology has great potential for the
development of tumor vaccines and provides a new treatment scheme
for cancer patients (Table IV).
Achieving the sustained antitumor response is a major limitation of
most current therapeutic methods of solid tumors and additional and
multimodal treatment approaches are required. HHP technology is an
effective method for producing whole-cell vaccines or DC-based
antitumor vaccines. However, to date, studies of both vaccines
remain in the preclinical phase (41). Therefore, there is still a need to
develop new vaccines for treatment with HHP that can avoid the
disadvantages of existing HHP vaccines and activate the immune
system to produce sustained antitumor immunity (Table V).
Tumor vaccines require activation of T cells to
resist the immunosuppressive microenvironment (166). Progressive tumors usually promote
tumor growth by promoting infiltration of tumor-bearing M2-like
macrophages, myeloid-derived suppressor cells (MDSC), and
regulatory T (T-Reg) cells, thereby inhibiting the local expansion
and effector functions of CD4+ T helper cells and
cytotoxic CD8+ T cells (167). Reluctant T-cell transfer vaccines
with or without costimulatory antibodies, particularly against
CD27, CD40, and CD137, can expand the tumor-specific T-cell pool
(168). In most patients, T
cell-mediated immunity in the tumor microenvironment is affected by
several mechanisms of suppressive immune cell use within the tumor
as well as by T-cell checkpoint suppression (169). This converts T cells into
lymphocytes, which may have a transient but modest effect on the
tumor. Specialized and selective regulation of the microenvironment
may lead to temporary tumor shrinkage and render intra-tumor T
cells resistant to the tumor (170).
This may be achieved by inducing an acute inflammatory response
using pattern recognition receptor (PRR) agonists or by removing or
inhibiting regulatory mechanisms that modulate immunity [T-Reg
cells, MDSC, and/or M2 tumor-associated macrophages (TAM)]
(171). If there is no additional
activation of a strong tumor-specific T-cell response, there is no
significant effect on tumor growth in most patients (172). If a tumor-specific T-cell response
has been ignited, checkpoints against cytotoxic
T-lymphocyte-associated protein 4, programmed cell death protein 1,
lymphocyte activation gene 3, antibody to T-cell immunoglobulin
mucin receptor 3, or natural killer cell receptor A (or against
their respective ligands) can help maintain the full effector
function of T cells within the tumor (173). In some patients, this will lead to
tumor destruction, while in others, immunosuppression of cells in
the microenvironment may prevail (174). In situations where immunosuppression
is alleviated and the tumor is sufficiently immunogenic, activation
of DCs and M1-like TAMs will promote the attraction and activation
of tumor-specific T cells and maintain the antitumor activity over
time (175). They may lead to tumor
eradication.
Several studies have observed in groups of patients
with different types of tumors that patients have improved outcomes
with immunotherapy when B cells compose a cluster of cells called
tertiary lymphoid structures (TLS) within the tumor (176,177).
Tumor-infiltrating B lymphocytes have been found in some tumor
tissues and are an important component of TLS in tumor tissues
(178). Tertiary lymphoid structures
are ectopic lymphoid organs formed in non-lymphoid tissues during
chronic inflammatory as well as tumor formation and consist of T
cells, B cells, follicular dendritic cells, as well as other cells
(179). TLS can be present in tumor
tissue in various states of maturation, with the highest level
forming germinal center structures (180). The impact of tumor-infiltrating B
cells and TLS on tumor formation and the efficacy of immunotherapy
have also received attention (180),
but their specific roles in tumors and their underlying mechanisms
are not fully understood. These results also indicate new
directions for subsequent research, combining T cell-mediated
immunotherapy with approaches using B cells, which may lead to more
effective antitumor therapies for more patients.
Inactivation technologies used to prepare tumor
cell vaccines should be aimed at inducing immunogenic malignant
cell death forms. If tumor cell vaccines are prepared by the
inactivation of whole tumor cells, the immunogenicity of the dead
tumor cells should be enhanced or at least maintained by this
procedure. The main focus of future oncology treatment concepts
should be to combine classical antitumor therapy with immunotherapy
to achieve the synergistic antitumor effects of both
modalities.
Not applicable.
This work was supported by the Funds of Jilin
Provincial Finance Department (grant no. JLSCZD2019-002).
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
KL and BL conceived and designed the study and
prepared the manuscript. SY and JL were responsible for the
literature search, data visualization, and analysis. LM and WT
retrieved the relevant literature and revised the manuscript for
important intellectual content. All authors read and approved the
final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Hoover DG and Farka DF: Biological effects
of high hydrostatic pressure on food microorganisms. Food Technol.
43:99–107. 1989.
|
|
2
|
Komora N, Maciel C, Pinto CA, Ferreira V,
Brandão TRS, Saraiva JMA, Castro SM and Teixeira P: Non-thermal
approach to Listeria monocytogenes inactivation in milk: The
combined effect of high pressure, pediocin PA-1 and bacteriophage
P100. Food Microbiol. 86:1033152020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin T, O'Keefe S, Duncan S and
Fernández-Fraguas C: Manipulation of the dry bean (Phaseolus
vulgaris L.) matrix by hydrothermal and high-pressure
treatments: Impact on in vitro bile salt-binding ability. Food
Chem. 310:1256992020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li X, Xue YM, Guo HM, Deng CY, Peng DW,
Yang H, Wei W, Liu Y, Liu FZ, Wang ZY, et al: High hydrostatic
pressure induces atrial electrical remodeling through upregulation
of inflammatory cytokines. Life Sci. 242:1172092020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kurosaka G, Uemura S, Mochizuki T, Kozaki
Y, Hozumi A, Suwa S, Ishii R, Kato Y, Imura S, Ishida N, et al: A
novel ER membrane protein Ehg1/May24 plays a critical role in
maintaining multiple nutrient permeases in yeast under
high-pressure perturbation. Sci Rep. 9:183412019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guillou S and Membre JM: Inactivation of
Listeria monocytogenes, Staphylococcus aureus, and
Salmonella enterica under high hydrostatic pressure: A
quantitative analysis of existing literature data. J Food Prot.
82:1802–1814. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Romek M, Kucia M, Gajda B, Krzysztofowicz
E and Smorag Z: Effect of high hydrostatic pressure on
mitochondrial activity, reactive oxygen species level and
developmental competence of cultured pig embryos. Theriogenology.
140:99–108. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Azeem M, Mu TH and Zhang M: Effects of
high hydrostatic pressure and soaking solution on proximate
composition, polyphenols, anthocyanins, β-carotene, and antioxidant
activity of white, orange, and purple fleshed sweet potato flour.
Food Sci Technol Int. 26:388–402. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang C, Xue Y, Yousaf L, Hu J and Shen Q:
Effects of high hydrostatic pressure on the ordered structure
including double helices and V-type single helices of rice starch.
Int J Biol Macromol. 144:1034–1042. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bartlett DH: Pressure effects on in vivo
microbial processes. Biochim Biophys Acta. 1595:367–381. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Knorr D, Heinz V and Buckow R: High
pressure application for food biopolymers. Biochim Biophys Acta.
1764:619–631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ott O, Rödel C, Weiss C, Wittlinger M,
Krause F, Dunst J, Fietkau R and Sauer R: Radiochemotherapy for
bladder cancer. Clin Oncol (R Coll Radiol). 21:557–565. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamashita H, Noguchi S, Murakami N, Toda
M, Uchino S, Watanabe S and Kawamoto H: Extracapsular invasion of
lymph node metastasis. A good indicator of disease recurrence and
poor prognosis in patients with thyroid microcarcinoma. Cancer.
86:842–849. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pastushenko I and Blanpain C: EMT
transition states during tumor progression and metastasis. Trends
Cell Biol. 29:212–226. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vandenberk L, Belmans J, Van Woensel M,
Riva M and Van Gool SW: Exploiting the immunogenic potential of
cancer cells for improved dendritic cell vaccines. Front Immunol.
6:6632016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Markovsky E, Budhu S, Samstein RM, Li H,
Russell J, Zhang Z, Drill E, Bodden C, Chen Q, Powell SN, et al: An
antitumor immune response is evoked by partial-volume single-dose
radiation in 2 murine models. Int J Radiat Oncol Biol Phys.
103:697–708. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang Y, Wang FM, Wang T, Wang YJ, Zhu ZY,
Gao YT and Du Z: Tumor-infiltrating FoxP3+ Tregs and
CD8+ T cells affect the prognosis of hepatocellular
carcinoma patients. Digestion. 86:329–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Helmstein K: Treatment of bladder
carcinoma by a hydrostatic pressure technique. Report on 43 cases.
Br J Urol. 44:434–450. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Goldman Y, Peled A and Shinitzky M:
Effective elimination of lung metastases induced by tumor cells
treated with hydrostatic pressure and N-acetyl-L-cysteine. Cancer
Res. 60:350–358. 2000.PubMed/NCBI
|
|
20
|
Shinitzky M and Goldman Y: Immunotherapy
of cancer with pressure modified cells. Isr Med Assoc J. 2:615–620.
2000.PubMed/NCBI
|
|
21
|
Rivalain N, Roquain J and Demazeau G:
Development of high hydrostatic pressure in biosciences: Pressure
effect on biological structures and potential applications in
biotechnologies. Biotechnol Adv. 28:659–672. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bridgman PW: The measurement of high
hydrostatic pressure. I. A simple primary gauge. Proc Am Acad Arts
Sci. 44:201–217. 1909. View Article : Google Scholar
|
|
23
|
Aertsen A, Meersman F, Hendrickx ME, Vogel
RF and Michiels CW: Biotechnology under high pressure: Applications
and implications. Trends Biotechnol. 27:434–441. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Frey B, Janko C, Ebel N, Meister S,
Schlucker E, Meyer-Pittroff R, Fietkau R, Herrmann M and Gaipl US:
Cells under pressure-treatment of eukaryotic cells with high
hydrostatic pressure, from physiologic aspects to pressure induced
cell death. Curr Med Chem. 15:2329–2336. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Roger H: Action des hautes pressions sur
quelques bactéries. Arch Physiol Norm Path. 7:12–17. 1895.
|
|
26
|
Hite BH: The effect of pressure in the
preservation of milk: A preliminary report. West Virginia
Agricultural Experiment Station. 1899.
|
|
27
|
Hite BH, Weakley CE and Giddings NJ: The
effect of pressure on certain micro-organisms encountered in the
preservation of fruits and vegetables. West Virginia University
Agricultural Experiment Station. 1914.
|
|
28
|
Basset J and Macheboeuf MA: Etude sur les
effets biologiques des ultrapressions: Résistance des bactéries,
des diastases et des toxines aux pressions très élevées. Comp Rend
Acad Sci. 195:1431–1433. 1932.
|
|
29
|
Atanasiu P, Barbu E and Basset J: Effect
of very high pressure on Newcastle virus. I. Dissociation of
infectious power and of hemagglutination. Ann Inst Pasteur (Paris).
81:340–343. 1951.(In Undetermined Language). PubMed/NCBI
|
|
30
|
Basset J, Lepine P and Chaumont L: Effects
of high pressures on the poliomyelitis virus (Lansing strain). Ann
Inst Pasteur (Paris). 90:575–593. 1956.(In French). PubMed/NCBI
|
|
31
|
Basset J, Macheboeuf M and Sandor G:
Etudes sur les effets biologiques des ultra-pressions. Action des
pressions très élevées sur les protéides. CR Hebd Acad Sci.
197:796–798. 1933.
|
|
32
|
Basset J, Wollman E, Macheboeuf MA and
Bardach M: Etudes sur les effets biologiques des ultra-pressions:
action des pressions élevées sur les tumeurs. CR Hebd Acad Sci.
200:2001935.
|
|
33
|
Johnson FH, Brown D and Marsland D: A
basic mechanism in the biological effects of temperature, pressure
and narcotics. Science. 95:200–203. 1942. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Marsland D and Jaffee O: Effects of
pressure on the cleaving eggs of the frog (Rana pipiens). J
Cell Comp Physiol. 34:439–450. 1949. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wolpert L, Marsland D and Hirshfield M:
The effect of high hydrostatic pressure on the mechanical
properties of the surface of the sea-urchin egg. J Cell Sci.
8:87–92. 1971.PubMed/NCBI
|
|
36
|
Bourns B, Franklin S, Cassimeris L and
Salmon ED: High hydrostatic pressure effects in vivo: Changes in
cell morphology, microtubule assembly, and actin organization. Cell
Motil Cytoskeleton. 10:380–390. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Haskin CL, Athanasiou KA, Klebe R and
Cameron IL: A heat-shock-like response with cytoskeletal disruption
occurs following hydrostatic pressure in MG-63 osteosarcoma cells.
Biochem Cell Biol. 71:361–371. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Parkkinen JJ, Ikonen J, Lammi MJ,
Laakkonen J, Tammi M and Helminen HJ: Effects of cyclic hydrostatic
pressure on proteoglycan synthesis in cultured chondrocytes and
articular cartilage explants. Arch Biochem Biophys. 300:458–465.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chawla R, Patil GR and Singh AK: High
hydrostatic pressure technology in dairy processing: A review. J
Food Sci Technol. 48:260–268. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang HW, Hsu CP and Wang CY: Healthy
expectations of high hydrostatic pressure treatment in food
processing industry. J Food Drug Anal. 28:1–13. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Adkins I, Hradilova N, Palata O, Sadilkova
L, Palova-Jelinkova L and Spisek R: High hydrostatic pressure in
cancer immunotherapy and biomedicine. Biotechnol Adv. 36:577–582.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Korn A, Frey B, Sheriff A, Gaipl US, Franz
S, Meyer-Pittroff R, Bluemelhuberh G and Herrmann M: High
hydrostatic pressure inactivated human tumour cells preserve their
immunogenicity. Cell Mol Biol (Noisy-le-grand). 50:469–477.
2004.PubMed/NCBI
|
|
43
|
Patterson MF: Microbiology of
pressure-treated foods. J Appl Microbiol. 98:1400–1409. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jacobo-Velázquez DA and Hernández-Brenes
C: Biochemical changes during the storage of high hydrostatic
pressure processed avocado paste. J Food Sci. 75:S264–S270. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Frey B, Franz S, Sheriff A, Korn A,
Bluemelhuber G, Gaipl US, Voll RE, Meyer-Pittroff R and Herrmann M:
Hydrostatic pressure induced death of mammalian cells engages
pathways related to apoptosis or necrosis. Cell Mol Biol
(Noisy-le-grand). 50:459–467. 2004.PubMed/NCBI
|
|
46
|
Weiss EM, Meister S, Janko C, Ebel N,
Schlücker E, Meyer-Pittroff R, Fietkau R, Herrmann M, Gaipl US and
Frey B: High hydrostatic pressure treatment generates inactivated
mammalian tumor cells with immunogeneic features. J Immunotoxicol.
7:194–204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yamaguchi T, Hashiguchi K, Katsuki S,
Iwamoto W, Tsuruhara S and Terada S: Activation of the intrinsic
and extrinsic pathways in high pressure-induced apoptosis of murine
erythroleukemia cells. Cell Mol Biol Lett. 13:49–57. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu K, Yan S, Ma Z and Liu B: Effective
pressure and treatment duration of high hydrostatic pressure to
prepare melanoma vaccines. Oncol Lett. 20:1135–1142. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Seitz C, Rückert M, Deloch L, Weiss EM,
Utz S, Izydor M, Ebel N, Schlücker E, Fietkau R, Gaipl US and Frey
B: Tumor Cell-Based vaccine generated with high hydrostatic
pressure synergizes with radiotherapy by generating a favorable
anti-tumor immune microenvironment. Front Oncol. 9:8052019.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nakamura-López Y, Sarmiento-Silva RE,
Moran-Andrade J and Gómez-García B: Staurosporine-induced apoptosis
in P388D1 macrophages involves both extrinsic and intrinsic
pathways. Cell Biol Int. 33:1026–1031. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ravichandran KS and Lorenz U: Engulfment
of apoptotic cells: Signals for a good meal. Nat Rev Immunol.
7:964–974. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Voll RE, Herrmann M, Roth EA, Stach C,
Kalden JR and Girkontaite I: Immunosuppressive effects of apoptotic
cells. Nature. 390:350–351. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
53
|
Locher C, Conforti R, Aymeric L, Ma Y,
Yamazaki T, Rusakiewicz S, Tesnière A, Ghiringhelli F, Apetoh L,
Morel Y, et al: Desirable cell death during anticancer
chemotherapy. Ann NY Acad Sci. 1209:99–108. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fraccaroli L, Alfieri J, Larocca L,
Calafat M, Mor G, Leirós CP and Ramhorst R: A potential tolerogenic
immune mechanism in a trophoblast cell line through the activation
of chemokine-induced T cell death and regulatory T cell modulation.
Hum Reprod. 24:166–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Griffith TS and Ferguson TA: Cell death in
the maintenance and abrogation of tolerance: The five Ws of dying
cells. Immunity. 35:456–466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Golstein P and Kroemer G: Cell death by
necrosis: Towards a molecular definition. Trends Biochem Sci.
32:37–43. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lavric M, Miranda-Garcia MA, Holzinger D,
Foell D and Wittkowski H: Alarmins firing arthritis: Helpful
diagnostic tools and promising therapeutic targets. Joint Bone
Spine. 84:401–410. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rahmani M, Reese E, Dai Y, Bauer C, Kramer
LB, Huang M, Jove R, Dent P and Grant S: Cotreatment with
suberanoylanilide hydroxamic acid and 17-allylamino
17-demethoxygeldanamycin synergistically induces apoptosis in
Bcr-Abl+ ψells sensitive and resistant to STI571
(imatinib mesylate) in association with down-regulation of Bcr-Abl,
abrogation of signal transducer and activator of transcription 5
activity, and Bax conformational change. Mol Pharmacol.
67:1166–1176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shin SA, Moon SY, Park D, Park JB and Lee
CS: Apoptotic cell clearance in the tumor microenvironment: A
potential cancer therapeutic target. Arch Pharm Res. 42:658–671.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Paudel YN, Angelopoulou E, Piperi C,
Balasubramaniam VRMT, Othman I and Shaikh MF: Enlightening the role
of high mobility group box 1 (HMGB1) in inflammation: Updates on
receptor signalling. Eur J Pharmacol. 858:1724872019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kepp O, Tesniere A, Schlemmer F, Michaud
M, Senovilla L, Zitvogel L and Kroemer G: Immunogenic cell death
modalities and their impact on cancer treatment. Apoptosis.
14:364–375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Munoz LE, Frey B, Pausch F, Baum W,
Mueller RB, Brachvogel B, Poschl E, Rodel F, von der Mark K,
Herrmann M and Gaipl US: The role of Annexin A5 in the modulation
of the immune response against dying and dead cells. Curr Med Chem.
14:271–277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sachet M, Liang YY and Oehler R: The
immune response to secondary necrotic cells. Apoptosis.
22:1189–1204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Gross M and Jaenicke R: Proteins under
pressure. The influence of high hydrostatic pressure on structure,
function and assembly of proteins and protein complexes. Eur J
Biochem. 221:617–630. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Avagyan S, Vasilchuk D and Makhatadze GI:
Protein adaptation to high hydrostatic pressure: Computational
analysis of the structural proteome. Proteins. 88:584–592. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Matsuki H, Kato K, Okamoto H, Yoshida S,
Goto M, Tamai N and Kaneshina S: Ligand partitioning into lipid
bilayer membranes under high pressure: Implication of variation in
phase-transition temperatures. Chem Phys Lipids. 209:9–18. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Perreault V, Hénaux L, Bazinet L and Doyen
A: Pretreatment of flaxseed protein isolate by high hydrostatic
pressure: Impacts on protein structure, enzymatic hydrolysis and
final hydrolysate antioxidant capacities. Food Chem. 221:1805–1812.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ding W, Palaiokostas M, Shahane G, Wang W
and Orsi M: Effects of high pressure on phospholipid bilayers. J
Phys Chem B. 121:9597–9606. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Guo K, Xiao W and Qiu D: Polymerization of
actin filaments coupled with adenosine triphosphate hydrolysis:
Brownian dynamics and theoretical analysis. J Chem Phys.
135:1051012011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Harrison SL, Barbosa-Cánovas GV and
Swanson BG: Pulsed electric field and high hydrostatic pressure
induced leakage of cellular material from saccharomyces cerevisiae.
Pulsed Electric Fields in Food Processing, Fundamental Aspects and
Applications. 183–191. 2019. View Article : Google Scholar
|
|
71
|
Bridgman PW: The coagulation of albumen by
pressure. J Biol Chem. 19:511–512. 1914. View Article : Google Scholar
|
|
72
|
Grant EA, Dow RB and Franks WR:
Denaturation of egg albumin by pressure. Science. 94:6161941.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Franck M, Perreault V, Suwal S, Marciniak
A, Bazinet L and Doyen A: High hydrostatic pressure-assisted
enzymatic hydrolysis improved protein digestion of flaxseed protein
isolate and generation of peptides with antioxidant activity. Food
Res Int. 115:467–473. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Charlier C, Alderson TR, Courtney JM, Ying
J, Anfinrud P and Bax A: Study of protein folding under native
conditions by rapidly switching the hydrostatic pressure inside an
NMR sample cell. Proc Natl Acad Sci USA. 115:E4169–E4178. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Barba FJ, Poojary MM, Wang J, Olsen K and
Orlien V: Effect of high pressure processing and storage on the
free amino acids in seedlings of Brussels sprouts. Innov Food Sci
Emerg Technol. 41:188–192. 2017. View Article : Google Scholar
|
|
76
|
Gederaas OA, Rasch MH, Berg K, Lagerberg
JW and Dubbelman TM: Photodynamically induced effects in colon
carcinoma cells (WiDr) by endogenous photosensitizers generated by
incubation with 5-aminolaevulinic acid. J Photochem Photobiol B.
49:162–170. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kobayashi M, Ohata K and Yoshida A:
Mitochondria membrane potential and developmental ability of mouse
oocytes were enhanced by high hydrostatic pressure treatment. Human
Reproduction. Oxford Univ Press Great Clarendon St; Oxford: Ox2
6dp. 2017
|
|
78
|
Shuai C, Liu G, Yang Y, Yang W, He C, Wang
G, Liu Z, Qi F and Peng S: Functionalized BaTiO3
enhances piezoelectric effect towards cell response of bone
scaffold. Colloids Surf B Biointerfaces. 185:1105872020. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Ribeiro S, Puckert C, Ribeiro C, Gomes AC,
Higgins MJ and Lanceros-Méndez S: Surface charge-mediated
cell-surface interaction on piezoelectric materials. ACS Appl Mater
Interfaces. 12:191–199. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tölgyesi FG and Böde C: Pressure and heat
shock proteins. Comparative High Pressure Biology. CRC Press; pp.
57–74. 2016
|
|
81
|
Nguyen HTM, Akanuma G, Hoa TTM, Nakai Y,
Kimura K, Yamamoto K and Inaoka T: Ribosome reconstruction during
Recovery from High-Hydrostatic-Pressure-Induced injury in Bacillus
subtilis. Appl Environ Microbiol. 86:e01640–19. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Mota MJ, Lopes RP, Delgadillo I and
Saraiva JA: Microorganisms under high pressure-adaptation, growth
and biotechnological potential. Biotechnol Adv. 31:1426–1434. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Al-Ayoubi SR, Schinkel PKF, Berghaus M,
Herzog M and Winter R: Combined effects of osmotic and hydrostatic
pressure on multilamellar lipid membranes in the presence of PEG
and trehalose. Soft matter. 14:8792–8802. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Mato JM: Lipid components of cellular
membranes. Phospholipid Metabolism in Cellular Signaling; CRC
Press; pp. 1–8. 2018, PubMed/NCBI
|
|
85
|
Nickels JD, Chatterjee S, Stanley CB, Qian
S, Cheng X, Myles DA, Standaert RF, Elkins JG and Katsaras J: The
in vivo structure of biological membranes and evidence for lipid
domains. PLoS Biol. 15:e20022142017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Manisegaran M, Bornemann S, Kiesel I and
Winter R: Effects of the deep-sea osmolyte TMAO on the temperature
and pressure dependent structure and phase behavior of lipid
membranes. Phys Chem Chem Phys. 21:18533–18540. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Winter R and Jeworrek C: Effect of
pressure on membranes. Soft Matter. 5:3157–3173. 2009. View Article : Google Scholar
|
|
88
|
Valentine RC and Valentine DL: Omega-3
fatty acids in cellular membranes: A unified concept. Prog Lipid
Res. 43:383–402. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ritz M, Tholozan JL, Federighi M and Pilet
MF: Morphological and physiological characterization of Listeria
monocytogenes subjected to high hydrostatic pressure. Appl Environ
Microbiol. 67:2240–2247. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Mahara A, Morimoto N, Sakuma T, Fujisato T
and Yamaoka T: Complete cell killing by applying high hydrostatic
pressure for acellular vascular graft preparation. Biomed Res Int.
2014:3796072014. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Gibson OR, Taylor L, Watt PW and Maxwell
NS: Cross-adaptation: Heat and cold adaptation to improve
physiological and cellular responses to hypoxia. Sports Med.
47:1751–1768. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Sinensky M: Homeoviscous adaptation-a
homeostatic process that regulates the viscosity of membrane lipids
in Escherichia coli. Proc Natl Acad Sci USA. 71:522–525.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Montagne K, Uchiyama H, Furukawa KS and
Ushida T: Hydrostatic pressure decreases membrane fluidity and
lipid desaturase expression in chondrocyte progenitor cells. J
Biomech. 47:354–359. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Purushothaman S, Cicuta P, Ces O and
Brooks NJ: Influence of high pressure on the bending rigidity of
model membranes. J Phys Chem B. 119:9805–9810. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kimura K, Morimatsu K, Inaoka T and
Yamamoto K: Injury and recovery of Escherichia coli
ATCC25922 cells treated by high hydrostatic pressure at 400–600
MPa. J Biosci Bioeng. 123:698–706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Macgregor RB: The interactions of nucleic
acids at elevated hydrostatic pressure. Biochim Biophys Acta.
1595:266–276. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Girard E, Prange T, Dhaussy AC,
Migianu-Griffoni E, Lecouvey M, Chervin JC, Mezouar M, Kahn R and
Fourme R: Adaptation of the base-paired double-helix molecular
architecture to extreme pressure. Nucleic Acids Res. 35:4800–4808.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lin X, Long L, Shan X, Zhang S, Shen S and
Liu B: In planta mobilization of mPing and its putative autonomous
element Pong in rice by hydrostatic pressurization. J Exp Bot.
57:2313–2323. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Long L, Lin X, Zhai J, Kou H, Yang W and
Liu B: Heritable alteration in DNA methylation pattern occurred
specifically at mobile elements in rice plants following
hydrostatic pressurization. Biochem Biophys Res Commun.
340:369–376. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Aertsen A and Michiels CW: Mrr instigates
the SOS response after high pressure stress in Escherichia
coli. Mol Microbiol. 58:1381–1391. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Kroemer G, Galluzzi L, Vandenabeele P,
Abrams J, Alnemri ES, Baehrecke EH, Blagosklonny MV, El-Deiry WS,
Golstein P, Green DR, et al: Classification of cell death:
Recommendations of the nomenclature committee on cell death 2009.
Cell Death Differ. 16:3–11. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Abe F: Exploration of the effects of high
hydrostatic pressure on microbial growth, physiology and survival:
Perspectives from piezophysiology. Biosci Biotechnol Biochem.
71:2347–2357. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Shi G, Yang Q, Zhang Y, Jiang Q, Lin Y,
Yang S, Wang H, Cheng L, Zhang X, Li Y, et al: Modulating the tumor
microenvironment via oncolytic viruses and CSF-1R inhibition
synergistically enhances Anti-PD-1 immunotherapy. Mol Ther.
27:244–260. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Hollingsworth RE and Jansen K: Turning the
corner on therapeutic cancer vaccines. NPJ Vaccines. 4:72019.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Saldanha G, Flatman K, Teo KW and Bamford
M: A novel numerical scoring system for melanoma tumour
infiltrating lymphocytes has better prognostic value than standard
scoring. Am J Surg Pathol. 41:906–914. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Vano YA, Petitprez F, Giraldo NA, Fridman
WH and Sautes-Fridman C: Immune-based identification of cancer
patients at high risk of progression. Curr Opin Immunol. 51:97–102.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Qiao Y, Choi JE, Vo JN, Tien JC, Wang L,
Xiao L, Simko SA, Delekta AD, Hodge NB, Desai P, et al: Therapeutic
targeting autophagy to sensitize cancer immunotherapy in various
cancer types. Cancer Res. 79 (Suppl 13):S41532019.
|
|
108
|
Sahin U and Türeci Ö: Personalized
vaccines for cancer immunotherapy. Science. 359:1355–1360. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hanna MG Jr: Autologous tumor vaccines and
methods. Google Patents. 2018.
|
|
110
|
Buckanovich RJ, Coukos G and Facciabene A:
Methods and compositions for treating solid tumors and enhancing
tumor vaccines. Google Patents. 2019.
|
|
111
|
Weiss EM, Wunderlich R, Ebel N, Rubner Y,
Schlücker E, Meyer-Pittroff R, Ott OJ, Fietkau R, Gaipl US and Frey
B: Selected anti-tumor vaccines merit a place in multimodal tumor
therapies. Front Oncol. 2:1322012. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Galluzzi L, Buqué A, Kepp O, Zitvogel L
and Kroemer G: Immunogenic cell death in cancer and infectious
disease. Nat Rev Immunol. 17:97–111. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Basset J, Lépine P and Chaumont L: Effets
des hautes pressions sur le virus de la poliomyelite
(souche-lansing). Annales de l institut pasteur; Paris: 1956
|
|
114
|
Deckmann M, Haimovitz R and Shinitzky M:
Selective release of integral proteins from human erythrocyte
membranes by hydrostatic pressure. Biochim Biophys Acta.
821:334–340. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Eisenthal A, Gonenne A, Skornick Y,
Gelfand A, Kaver I, Yelin A, Yehoshua H, Lifschitz-Mercer B and
Shinitzky M: Th2 to Th1 cytokine shift in autologous peripheral
blood mononuclear cells responding to autologous human tumor cells
modified by hydrostatic pressure and cross-linking (PCL) in vitro.
Annual Meeting of the American Association of Cancer Research.
Heidelberg SB: 37. Springer; Washington, DC: pp. ppA32321996
|
|
116
|
Chene L, Sader CD, Magalhaes J, Strozzi F,
Tibaldi L, Mendez C, Baeriswyl S, Laveissiere A and Bonny C:
Microbiome derived peptides stimulate strong immune response
against tumor associated antigens and trigger in vivo tumor
regression after vaccination. Cancer Res. 79 (Suppl
13):S14752019.
|
|
117
|
Weir G, Quinton T, Hutchins JT, Freimark
BD and Stanford M: Phosphatidylserine-targeting antibodies enhance
anti-tumor activity of a tumor vaccine in a HPV-induced tumor
model. Cancer Res. 77 (Suppl 13):S36572017.
|
|
118
|
Xu L, Kwak M, Zhang W, Zeng L, Lee PC and
Jin JO: Rehmannia glutinosa polysaccharide induces toll-like
receptor 4 dependent spleen dendritic cell maturation and
anti-cancer immunity. Oncoimmunology. 6:e13259812017. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Moserova I, Truxova I, Garg AD, Tomala J,
Agostinis P, Cartron PF, Vosahlikova S, Kovar M, Spisek R and
Fucikova J: Caspase-2 and oxidative stress underlie the immunogenic
potential of high hydrostatic pressure-induced cancer cell death.
Oncoimmunology. 6:e12585052016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Kroemer G, Galluzzi L, Kepp O and Zitvogel
L: Immunogenic cell death in cancer therapy. Annu Rev Immunol.
31:51–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Lin CF, Chen CL, Chang WT, Jan MS, Hsu LJ,
Wu RH, Tang MJ, Chang WC and Lin YS: Sequential caspase-2 and
caspase-8 activation upstream of mitochondria during ceramideand
etoposide-induced apoptosis. J Biol Chem. 279:40755–40761. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Garg AD, Krysko DV, Verfaillie T,
Kaczmarek A, Ferreira GB, Marysael T, Rubio N, Firczuk M, Mathieu
C, Roebroek AJ, et al: A novel pathway combining calreticulin
exposure and ATP secretion in immunogenic cancer cell death. EMBO
J. 31:1062–1079. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Sandow JJ, Dorstyn L, O'reilly LA, Tailler
M, Kumar S, Strasser A and Ekert PG: ER stress does not cause
upregulation and activation of caspase-2 to initiate apoptosis.
Cell Death Differ. 21:475–480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Azevedo AP, Silva SN, Reichert A, Lima F,
Júnior E and Rueff J: The role of caspase genes polymorphisms in
genetic susceptibility to philadelphia-negative myeloproliferative
neoplasms in a Portuguese population. Pathol Oncol Res. 25:961–969.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zappasodi R, de Braud F and Di Nicola M:
Lymphoma immunotherapy: Current status. Front Immunol. 6:4482015.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Mikyskova R, Indrova M, Stepanek I,
Kanchev I, Bieblova J, Vosahlikova S, Moserova I, Truxova I,
Fucikova J, Bartunkova J, et al: Dendritic cells pulsed with tumor
cells killed by high hydrostatic pressure inhibit prostate tumor
growth in TRAMP mice. Oncoimmunology. 6:e13625282017. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Hradilova N, Sadilkova L, Palata O,
Mysikova D, Mrazkova H, Lischke R, Spisek R and Adkins I:
Generation of dendritic cell-based vaccine using high hydrostatic
pressure for non-small cell lung cancer immunotherapy. PLoS One.
12:e01715392017. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Sakamoto M, Morimoto N, Jinno C, Mahara A,
Ogino S, Suzuki S, Kusumoto K and Yamaoka T: Melanin pigments in
the melanocytic nevus regress spontaneously after inactivation by
high hydrostatic pressure. PLoS One. 12:e01869582017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Liang ZL, Mao QY, Wang YP, Zhu FC, Li JX,
Yao X, Gao F, Wu X, Xu M and Wang JZ: Progress on the research and
development of inactivated EV71 whole-virus vaccines. Hum Vaccin
Immunother. 9:1701–1705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Urbanova L, Hradilova N, Moserova I,
Vosahlikova S, Sadilkova L, Hensler M, Spisek R and Adkins I: High
hydrostatic pressure affects antigenic pool in tumor cells:
Implication for dendritic cell-based cancer immunotherapy. Immunol
Lett. 187:27–34. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Kammer GM, Kurrasch R and Scillian JJ:
Capping of the surface OKT3 binding molecule prevents the T-cell
proliferative response to antigens: Evidence that this molecule
conveys the activation signal. Cell Immunol. 87:284–294. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Kast WM, Levitsky H and Marincola FM:
Synopsis of the 6th Walker's cay colloquium on cancer vaccines and
immunotherapy. J Transl Med. 2:202004. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Rosenblatt J and Avigan D: Cellular
immunotherapy for multiple myeloma. Best Pract Res Clin Haematol.
21:559–577. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Jocham D, Richter A, Hoffmann L, Iwig K,
Fahlenkamp D, Zakrzewski G, Schmitt E, Dannenberg T, Lehmacher W,
von Wietersheim J and Doehn C: Adjuvant autologous renal tumour
cell vaccine and risk of tumour progression in patients with
renal-cell carcinoma after radical nephrectomy: Phase III,
randomised controlled trial. Lancet. 363:594–599. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Pfeffer CM and Singh ATK: Apoptosis: A
target for anticancer therapy. Int J Mol Sci. 19:4482018.
View Article : Google Scholar
|
|
136
|
Garg AD, Galluzzi L, Apetoh L, Baert T,
Birge RB, Bravo-San Pedro JM, Breckpot K, Brough D, Chaurio R,
Cirone M, et al: Molecular and translational classifications of
DAMPs in immunogenic cell death. Front Immunol. 6:5882015.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Gomes-da-Silva LC, Zhao L, Arnaut LG,
Kroemer G and Kepp O: Redaporfin induces immunogenic cell death by
selective destruction of the endoplasmic reticulum and the Golgi
apparatus. Oncotarget. 9:311692018. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
McDaniel A, Anand S, Alzubier A, Berlin J,
Hartman H, Uecker D and Nuccitelli R: Profiling the immunogenic
cell death (ICD) mechanisms induced by Nano-Pulse Stimulation (NPS)
treatment in mouse B16-F10 melanoma tumors using NanoString
technology. Pulse Biosciences London; 2017
|
|
139
|
Wang Q, Ju X, Wang J, Fan Y, Ren M and
Zhang H: Immunogenic cell death in anticancer chemotherapy and its
impact on clinical studies. Cancer Lett. 438:17–23. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Fucikova J, Moserova I, Truxova I,
Hermanova I, Vancurova I, Partlova S, Fialova A, Sojka L, Cartron
PF, Houska M, et al: High hydrostatic pressure induces immunogenic
cell death in human tumor cells. Int J Cancer. 135:1165–1177. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Fucikova J, Koci L, Pokorna K, Truxova I,
Moserova I, Rozkova D and Spisek R: Cryopreservation of apoptotic
cancer cells for use in immunotherapy against cancer. Google
Patents. 2019.
|
|
142
|
Iribarren K, Buque A, Mondragon L, Xie W,
Lévesque S, Pol J, Zitvogel L, Kepp O and Kroemer G: Anticancer
effects of anti-CD47 immunotherapy in vivo. Oncoimmunology.
8:15506192019. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
van Panhuys N: TCR signal strength alters
T-DC activation and interaction times and directs the outcome of
differentiation. Front Immunol. 7:62016. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Gross S, Erdmann M, Haendle I, Voland S,
Berger T, Schultz E, Strasser E, Dankerl P, Janka R, Schliep S, et
al: Twelve-year survival and immune correlates in dendritic
cell-vaccinated melanoma patients. JCI Insight. 2:e914382017.
View Article : Google Scholar
|
|
145
|
De Sanctis F, Sandri S, Martini M,
Mazzocco M, Fiore A, Trovato R, Garetto S, Brusa D, Ugel S and
Sartoris S: Hyperthermic treatment at 56°C induces tumour-specific
immune protection in a mouse model of prostate cancer in both
prophylactic and therapeutic immunization regimens. Vaccine.
36:3708–3716. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Mellman I, Coukos G and Dranoff G: Cancer
immunotherapy comes of age. Nature. 480:480–489. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Constantino J, Gomes C, Falcão A, Cruz MT
and Neves BM: Antitumor dendritic cell-based vaccines: Lessons from
20 years of clinical trials and future perspectives. Transl Res.
168:74–95. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Tjoa B, Boynton A, Kenny G, Ragde H,
Misrock SL and Murphy G: Presentation of prostate tumor antigens by
dendritic cells stimulates T-cell proliferation and cytotoxicity.
Prostate. 28:65–69. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Ning Y, Shen K, Wu Q, Sun X, Bai Y, Xie Y,
Pan J and Qi C: Tumor exosomes block dendritic cells maturation to
decrease the T cell immune response. Immunol Lett. 199:36–43. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Van den Eertwegh AJ, Versluis J, Van den
Berg HP, Santegoets SJ, Van Moorselaar RJA, Van der Sluis TM, Gall
HE, Harding TC, Jooss K, Lowy I, et al: Combined immunotherapy with
granulocyte-macrophage colony-stimulating factor-transduced
allogeneic prostate cancer cells and ipilimumab in patients with
metastatic castration-resistant prostate cancer: A phase 1
dose-escalation trial. Lancet Oncol. 13:509–517. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Mikyšková R, Štěpánek I, Indrová M,
Bieblová J, Šímová J, Truxová I, Moserová I, Fučíková J, Bartůňková
J, Špíšek R and Reiniš M: Dendritic cells pulsed with tumor cells
killed by high hydrostatic pressure induce strong immune responses
and display therapeutic effects both in murine TC-1 and TRAMP-C2
tumors when combined with docetaxel chemotherapy. Int J Oncol.
48:953–964. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Neves BM, Cruz MT, Francisco V,
Garcia-Rodriguez C, Silvestre R, Cordeiro-da-Silva A, Dinis AM,
Batista MT, Duarte CB and Lopes MC: Differential roles of
PI3-kinase, MAPKs and NF-kappaB on the manipulation of dendritic
cell T(h)1/T(h)2 cytokine/chemokine polarizing profile. Mol
Immunol. 46:2481–2492. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Budai Z, Ujlaky-Nagy L, Kis GN, Antal M,
Bankó C, Bacsó Z, Szondy Z and Sarang Z: Macrophages engulf
apoptotic and primary necrotic thymocytes through similar
phosphatidylserine-dependent mechanisms. FEBS Open Bio. 9:446–456.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Bolean M, Borin IA, Simão AMS, Bottini M,
Bagatolli LA, Hoylaerts MF, Millán JL and Ciancaglini P:
Topographic analysis by atomic force microscopy of proteoliposomes
matrix vesicle mimetics harboring TNAP and AnxA5. Biochim Biophys
Acta Biomembr. 1859:1911–1920. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Cai Y, Ran W, Zhai Y, Wang J, Zheng C, Li
Y and Zhang P: Recent progress in supramolecular peptide assemblies
as virus mimics for cancer immunotherapy. Biomater Sci.
8:1045–1057. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
He J, Yin Y, Luster TA, Watkins L and
Thorpe PE: Antiphosphatidylserine antibody combined with
irradiation damages tumor blood vessels and induces tumor immunity
in a rat model of glioblastoma. Clin Cancer Res. 15:6871–6880.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Bondanza A, Zimmermann VS, Rovere-Querini
P, Turnay J, Dumitriu IE, Stach CM, Voll RE, Gaipl US, Bertling W,
Pöschl E, et al: Inhibition of phosphatidylserine recognition
heightens the immunogenicity of irradiated lymphoma cells in vivo.
J Exp Med. 200:1157–1165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Hodge JW, Guha C, Neefjes J and Gulley JL:
Synergizing radiation therapy and immunotherapy for curing
incurable cancers: Opportunities and challenges. Oncology
(Williston Park). 22:1064–1070. 2008.PubMed/NCBI
|
|
159
|
Guo X, Song J, Zhao J, Wang B, Yang Z, Sun
P and Hu M: Impact of ANXA5 polymorphisms on glioma risk and
patient prognosis. J Neurooncol. 142:11–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Cossarizza A, Cooper EL, Suzuki MM,
Salvioli S, Capri M, Gri G, Quaglino D and Franceschi C: Earthworm
leukocytes that are not phagocytic and cross-react with several
human epitopes can kill human tumor cell lines. Exp Cell Res.
224:174–182. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Pouwels SD, Zijlstra GJ, van der Toorn M,
Hesse L, Gras R, Ten Hacken NH, Krysko DV, Vandenabeele P, de Vries
M, van Oosterhout AJ, et al: Cigarette smoke-induced necroptosis
and DAMP release trigger neutrophilic airway inflammation in mice.
Am J Physiol Lung Cell Mol Physiol. 310:L377–L386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Kurosaka K, Watanabe N and Kobayashi Y:
Potentiation by human serum of anti-inflammatory cytokine
production by human macrophages in response to apoptotic cells. J
Leukoc Biol. 71:950–956. 2002.PubMed/NCBI
|
|
163
|
Fischer U, Koppang EO and Nakanishi T:
Teleost T and NK cell immunity. Fish Shellfish Immunol. 35:197–206.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Peng B, Liu S, Guo C, Sun X and Sun MZ:
ANXA5 level is linked to in vitro and in vivo tumor malignancy and
lymphatic metastasis of murine hepatocarcinoma cell. Future Oncol.
12:31–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Yang L, Lin S, Kang Y, Xiang Y, Xu L, Li
J, Dai X, Liang G, Huang X and Zhao C: Rhein sensitizes human
pancreatic cancer cells to EGFR inhibitors by inhibiting STAT3
pathway. J Exp Clin Cancer Res. 38:312019. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Gujar SA, Marcato P, Pan D and Lee PW:
Reovirus virotherapy overrides tumor antigen presentation evasion
and promotes protective antitumor immunity. Mol Cancer Ther.
9:2924–2933. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
He D, Li H, Yusuf N, Elmets CA, Li J,
Mountz JD and Xu H: IL-17 promotes tumor development through the
induction of tumor promoting microenvironments at tumor sites and
myeloid-derived suppressor cells. J Immunol. 184:2281–2288. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Ma Y, Kepp O, Ghiringhelli F, Apetoh L,
Aymeric L, Locher C, Tesniere A, Martins I, Ly A, Haynes NM, et al:
Chemotherapy and radiotherapy: Cryptic anticancer vaccines. Semin
Immunol. 22:113–124. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Hurwitz AA and Watkins SK: Immune
suppression in the tumor microenvironment: A role for dendritic
cell-mediated tolerization of T cells. Cancer Immunol Immunother.
61:289–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Brix K, McInnes J, Al-Hashimi A, Rehders
M, Tamhane T and Haugen MH: Proteolysis mediated by cysteine
cathepsins and legumain-recent advances and cell biological
challenges. Protoplasma. 252:755–774. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Zhou SL, Zhou ZJ, Hu ZQ, Huang XW, Wang Z,
Chen EB, Fan J, Cao Y, Dai Z and Zhou J: Tumor-Associated
neutrophils recruit macrophages and T-Regulatory cells to promote
progression of hepatocellular carcinoma and resistance to
sorafenib. Gastroenterology. 150:1646–1658.e17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Zhang R and Misra V: Effects of cyclic AMP
response element binding protein-Zhangfei (CREBZF) on the unfolded
protein response and cell growth are exerted through the tumor
suppressor p53. Cell Cycle. 13:279–292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Liu Z, Noh HS, Chen J, Kim JH, Falo LD Jr
and You Z: Potent tumor-specific protection ignited by adoptively
transferred CD4+ T cells. J Immunol. 181:4363–4370. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Krneta T, Gillgrass A and Ashkar AA: The
influence of macrophages and the tumor microenvironment on natural
killer cells. Curr Mol Med. 13:68–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
McCloskey ML, Curotto de Lafaille MA,
Carroll MC and Erlebacher A: Acquisition and presentation of
follicular dendritic cell-bound antigen by lymph node-resident
dendritic cells. J Exp Med. 208:135–148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Bruno TC: New predictors for immunotherapy
responses sharpen our view of the tumour microenvironment. Nature.
577:474–476. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Petitprez F, de Reyniès A, Keung EZ, Chen
TW, Sun CM, Calderaro J, Jeng YM, Hsiao LP, Lacroix L, Bougoüin A,
et al: B cells are associated with survival and immunotherapy
response in sarcoma. Nature. 577:556–560. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Calorini L and Bianchini F: Environmental
control of invasiveness and metastatic dissemination of tumor
cells: The role of tumor cell-host cell interactions. Cell Commun
Signal. 8:242010. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Marinkovic T and Marinkovic D: Biological
mechanisms of ectopic lymphoid structure formation and their
pathophysiological significance. Int Rev Immunol. Jun 7–2020.(Epub
ahead of print). doi: 10.1080/08830185.2020.1789620. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Somersan S, Larsson M, Fonteneau JF, Basu
S, Srivastava P and Bhardwaj N: Primary tumor tissue lysates are
enriched in heat shock proteins and induce the maturation of human
dendritic cells. J Immunol. 167:4844–4852. 2001. View Article : Google Scholar : PubMed/NCBI
|