Introduction
Lung cancer is a disease that is characterized by
uncontrolled growth of abnormal cells in one or both of the lungs.
Lung cancer is a common type of cancer that has been identified as
the leading cause of cancer-related deaths worldwide. The data from
GLOBOCAN 2018, a project of the International Agency for Research
on Cancer (IARC), estimated that lung cancer caused 1.8 million
deaths, resulting in a mortality rate of 18.4%; thus, lung cancer
ranks first in mortality among all cancer types. In Thailand,
23,957 (15.1%) new lung cancer cases and 21,371 (20.1%) lung
cancer-related deaths were reported; thus, lung cancer ranks second
after liver cancer (1). The high
mortality rate results from the late diagnosis of patients already
in advanced or metastatic stages (2).
Therefore, studying and understanding the molecular mechanisms
underlying lung cancer metastasis are essential for the development
of therapeutic strategies for lung cancer.
Cancer metastasis is a complex process in which
cancer cells spread from a primary tumor site to distant sites.
Metastasis includes several processes, including cell
proliferation, adhesion, invasion, migration and angiogenesis
(3). These processes involve the
complex interactions of multiple crucial proteins and signaling
pathways that lead to cancer cell movement and survival. Cancer
metastasis is typically initiated by the attachment of cancer cells
to extracellular matrix (ECM) proteins and then by the activation
of integrins (4). Integrin activation
contributes to the formation of focal adhesions (FAs) and the
stimulation of signaling transduction pathways that regulate cell
migration and invasion. FA proteins recruit focal adhesion kinase
(FAK) to FAs, and then FAK directly binds to integrins, which
subsequently leads to FAK autophosphorylation at Tyr397. FAK is a
nonreceptor tyrosine kinase that regulates signaling related to
cell adhesion and migration in various cell lines (5,6). For cell
migration, FAK activates p85 subunit of phosphatidylinositol
3-kinase (PI3K) (7), which further
increases the phosphorylation of the protein kinase-B (AKT) and
mammalian target of rapamycin (mTOR) proteins. The involvement of
PI3K/AKT/mTOR signaling in cancer metastasis has been clearly
observed in numerous works through the stimulation of cancer cell
development, migration, invasion and motility (8). In addition, phosphorylating ribosomal
p70S6 kinase (P70S6K) and eukaryotic translation factor 4E-binding
protein 1 (4E-BP1), which are downstream effectors of mTORC1, can
also promote cancer cell migration and invasion (9) and cell cycle progression (10).
Angiogenesis is another crucial process that
promotes tumor growth and metastasis. This process is activated
through the interaction of growth factors and their specific
receptors. Vascular endothelial growth factors (VEGFs) are the main
regulators of vascular development and angiogenesis initiation.
VEGF subtype A (VEGF-A) has been widely studied and plays a major
role in angiogenesis by acting through VEGF receptor tyrosine
kinase type 2 (VEGFR2) (11,12). VEGFR2 is expressed in different types
of cells, such as neuronal cells, megakaryocytes, hematopoietic
stem cells and various cancer cells, including lung cancer cells
(13–16). The Tyr951, Tyr1175 and Tyr1214
residues of VEGFR2 are the main sites of phosphorylation that
stimulate cell migration during angiogenesis. Phosphorylation at
Tyr1175 recruits the binding of SH2 domain-containing adaptor
protein B (SHB), resulting in the activation of FAK and the
promotion of actin polymerization, lamellipodia formation and cell
migration (17,18).
For lung cancer treatment, standard treatments, such
as radiotherapy and chemotherapy, are currently accepted for
treating advanced stage lung cancer; however, there are still
several issues that need to be considered, such as treatment side
effects, drug resistance, and high treatment cost. The development
of effective anti-lung cancer agents, especially from natural
resources, is a promising approach to increase patient survival.
Terrein
(4,5-dihydroxy-3-[(E)-1′-propenyl]-2-cyclopenten-l-one,
C8H10O3) is a natural substance
produced by microorganisms. Terrein is a secondary bioactive fungal
metabolite that was first isolated from Aspergillus terreus
by Raistrick and Smith in 1935 (19).
Numerous studies have demonstrated that terrein exerts several
potential effects, including anti-inflammation, melanogenesis
inhibition and anticancer effects (20–22).
Regarding its anticancer effect, terrein was revealed to inhibit
cell proliferation through the induction of cell cycle arrest in
human hepatoma Bel7402 (23) and
human ovarian cancer cells (24).
Terrein has also been revealed to induce the apoptosis of
ABCG2-expressing breast cancer cells via the caspase-7 pathway and
inhibit AKT signaling (25).
Additionally, terrein induced apoptosis by regulating p53 and ERK
in human cervical carcinoma cells (22). Furthermore, terrein has been revealed
to suppress angiogenin production in head and neck cancer cells
(26) and androgen-dependent prostate
cancer cells (27), and both studies
suggested that terrein has an inhibitory effect on angiogenesis.
The inhibitory effect of terrein on cell migration has also been
observed in human breast cancer cells (28). However, the anticancer effect of
terrein on A549 human lung cancer cell metastasis and angiogenesis
has not yet been fully elucidated. Therefore, the present study
aimed to investigate the inhibitory effects of terrein on human
lung cancer cell metastasis and angiogenesis as well as the
important cellular mechanisms involved in both processes.
Materials and methods
Chemicals and reagents
Dulbecco's modified Eagle's medium (DMEM), Eagle's
minimum essential medium (EMEM), α-minimum essential medium
(α-MEM), fetal bovine serum (FBS), 0.25% trypsin-EDTA, and
penicillin-streptomycin were purchased from Gibco/Thermo Fisher
Scientific, Inc. 3-(4,5-Dimethylthiazol-2-yl)-2,5
diphenyltetrazolium bromide (MTT) was purchased from USB
Corporation. Matrigel was purchased from BD Biosciences. The VEGF-A
Human ELISA Kit was purchased from Abcam. The lactate dehydrogenase
(LDH) assay was purchased from G-Biosciences.
Cell culture
The A549 human NSCLC cell line (ATCC®
CCL-185™) and normal African green monkey kidney (Vero) cell line
(ATCC® CCL-81™) were purchased from ATCC. The
L6 skeletal muscle cell line and H9C2 cardiomyoblast cell line were
obtained from author Gary Sweeney, Department of Biology, York
University, Toronto, Canada (29,30). The
A549 cells and H9C2 cells were maintained in DMEM containing 10%
FBS and 1% penicillin/streptomycin. The Vero cells were maintained
in EMEM containing 10% FBS and 1% penicillin/streptomycin. The L6
cells were maintained in α-MEM containing 10% FBS and 1%
antibiotic-antimycotic. All the cell types were cultured in an
incubator at 37°C in 5% CO2 and a 95% humidified
atmosphere. The cells were provided with fresh medium 2 to 3 times
per week, and when the cells had grown to approximately 80%
confluence, they were subcultured approximately 2 or 3 times per
week.
Preparation of terrein
The fungus Aspergillus terreus CRI301 was
cultivated in Sabouraud dextrose agar under stationary conditions
at room temperature for 34 days. Then, the Aspergillus
terreus CRI301 culture was filtered to separate the cells from
the broth. The culture broth was extracted three times with an
equal volume of ethyl acetate (EtOAc), and then, the EtOAc layers
were combined and evaporated to dryness. The crude EtOAc extract
was further purified by Sephadex LH-20 column chromatography (2-cm
inner diameter and 125-cm length) and eluted with MeOH. The
structure of terrein was characterized by 1H NMR
spectroscopy (AVANCE III HD; frequency, 400 MHz; TopSpin version
3.6.2 software; Bruker).
Cell viability assay
The cytotoxic effects of terrein in A549 cells and
normal cells, including Vero cells, L6 cells and H9C2 cells, was
determined using a colorimetric MTT assay. A549, Vero, L6 and H9C2
cells were harvested with 0.25% trypsin containing 1 mM EDTA,
plated in 96-well plates at densities of 1×104,
1.8×104, 3×104, and 2×104
cells/well, respectively, and allowed to adhere overnight. Then,
the cells were treated with various concentrations of terrein: 0–1
mM for A549 cells and 0–2 mM for all the normal cell lines. The
highest concentration of dimethyl sulfoxide (DMSO; 0.1%) was used
as the vehicle control. Then, all the plates were incubated for 24
h at 37°C. Subsequently, the medium was removed, and 0.5 mg/ml MTT
solution was added to each well. The plates were further incubated
for 4 h at 37°C, and the supernatants were discarded after the
incubation. Then, the formazan crystals in each well were dissolved
in 100 µl of DMSO. The amount of purple formazan was determined by
using a multimode microplate reader (Synergy; BioTek Instruments,
Inc.) at 595 nm. All the measurements were carried out in
triplicate. The cell viability is presented as the percentage of
the control.
Cell proliferation assay
An IncuCyte proliferation assay was used to
determine the effect of terrein on human lung cancer cell
proliferation. Real-time live-cell imaging was conducted using
IncuCyte S3 (Essen BioScience; Sartorius). A549 cells were seeded
in 96-well plates at a density of 4×103 cells/well and
allowed to adhere overnight. Then, the cells were treated with
various concentrations (0–1 mM) of terrein, and the plates were
placed in the IncuCyte for imaging every 3 h for 3 days.
Proliferation curves were generated using IncuCyte proliferation
analysis with confluence as the parameter.
LDH enzyme assay
The cytotoxic effect of terrein on A549 cells was
determined using an LDH assay kit, following manufacturer's
instructions (G-Biosciences). Briefly, the cells (1×104
cells/well) were treated with various concentrations (0–1 mM) of
terrein for 24 h. Then, 25 µl of supernatant from each sample was
transferred to a new 96-well plate, and 25 µl reaction mixture was
added to each well. After incubation at 37°C for 30 min, 25 µl stop
solution was added to each well, and then, the absorbance was
measured using a multimode microplate reader (Synergy; BioTek
Instruments, Inc.) at 490 nm.
Wound healing assay
The effect of terrein on A549 cell migration was
determined by a monolayer wound healing assay. A549 cells were
harvested with 0.25% trypsin containing 1 mM EDTA, plated in 6-well
plates at a density of 5×105 cells/well in serum-free
medium and allowed to adhere overnight. After forming a confluent
monolayer, the cells were scratched with a 200-µl sterile pipette
tip from one side to the other of the wells of the 6-well culture
plate. Subsequently, the cells were washed three times with DMEM to
remove the cell debris. Then, the cells in each well were
immediately given DMEM with or without 20, 40 and 80 µM terrein.
Cell migration was monitored and imaged under a light inverted
microscope (Olympus CKX41; Olympus Corporation) using a
magnification of ×10 for 0, 6, 12 and 24 h. The wound area was
measured in three independent wound sites per group and compared
with that in the vehicle control group at 0 h. Relative cell
motility was calculated as the wound area at 6, 12 and 24 h in at
least three independent experiments. The wound areas were
quantified using ImageJ software [Java 1.8.0_112 (64_bit); National
Institutes of Health].
Transwell migration and invasion
assays
The effect of terrein on aggressive A549 cell
migration and invasion was assessed using 24-well plate Transwell
inserts with polycarbonate membranes with 8-µm pores (BD
Biosciences). Briefly, the cells were harvested, and
5×104 cells were resuspended in 200 µl serum-free medium
with or without 20, 40 and 80 µM terrein. The cells were then
plated into the Matrigel-coated upper chambers of Transwell inserts
for the invasion assay or the noncoated upper chambers of Transwell
inserts for the migration assay. In the lower chamber, 750 µl
medium containing 10% FBS was used as a chemoattractant to
stimulate cell invasion and migration. The plate was incubated at
37°C for 24 h. The Transwell insert was separated from the 24-well
plate and washed twice with cold PBS. Each sample was fixed with
100% ice-cold methanol for 20 min and washed twice with cold PBS.
Then, the samples were stained with 0.5% crystal violet at room
temperature for 15 min and washed with dH2O several
times until all the excess dye had been removed. The cells that did
not invade or migrate from the upper chamber were removed using
cotton swabs. The cells that invaded and migrated through the pores
of the insert into the lower chamber were photographed under an
light inverted microscope at a magnification of ×10 and quantified
from at least five randomly selected fields by ImageJ software
[Java 1.8.0_112 (64_bit)]. The percentage of cell invasion and
migration were compared to that in the vehicle control group, which
was set to 100%.
Adhesion assay
The effect of terrein on A549 cell adhesion was
determined. Briefly, 96-well plates were coated with 50 µl of 2.0
mg/ml Matrigel and incubated at 37°C for 2 h. Then, the Matrigel
was removed, and cells at a density of 3×104 were plated
in each well in 100 µl of serum-free medium with or without 20, 40
and 80 µM terrein. After incubation for 30 min at 37°C, the
nonadherent cells were removed by washing 4 times with 50 µl of
PBS, and 0.5 mg/ml MTT solution was added and incubated for 4 h at
37°C. The amount of adherent cells was determined by measuring the
optical density using a multimode microplate reader at 595 nm. Cell
adhesion is expressed as a percentage of that observed in the
control group.
Gelatin zymography
A gelatin zymography assay was used to examine the
effect of terrein on the activities of proteolytic enzymes released
from human lung cancer cells. Briefly, A549 cells were seeded in
6-well plates at a density of 5×105 cells/well in DMEM
containing 10% FBS and incubated overnight. The cells were treated
with or without 20, 40 and 80 µM terrein and incubated at 37°C for
24 h. After treatment, the supernatants were collected and
concentrated using Amicon Ultra-0.5 Centrifugal Filters 3K (Thermo
Fisher Scientific, Inc.). The protein concentration was measured
using a Bradford protein assay. Bovine serum albumin (BSA) was used
as the standard protein sample. After the calculation of the
protein concentration, each sample was adjusted to the same
concentration.
The protein samples were separated in 10%
SDS-polyacrylamide gels containing 0.73 mg/ml gelatin. After
electrophoresis, the gels were washed with 1X renaturing buffer
[Novex™ Zymogram Renaturing Buffer (10X) Invitrogen™; Thermo Fisher
Scientific, Inc.] for 1 h at room temperature with gentle agitation
on a benchtop rocker. The gels were washed 4 times with
ddH2O for 20 min each and then incubated with 1X
developing buffer [Novex™ Zymogram Developing Buffer (10X)
Invitrogen™; Thermo Fisher Scientific, Inc.] for 30 min at room
temperature with gentle agitation on a benchtop rocker. Then, the
gels were incubated with fresh 1X developing buffer at 37°C
overnight. Subsequently, the gels were stained with staining
solution (0.5% Coomassie Brilliant Blue R-250 in methanol:acetic
acid:water, 4:1:5, v/v/v) and then washed with destaining solution.
Finally, the bands corresponding to matrix metalloproteinase
(MMP)-2 (68 kDa) and MMP-9 (82 kDa) were clearly visible in
contrast to the blue background and were detected by a MiniBIS Pro
DNR Bio-Imaging System (BioSciences). The intensities of the bands
were quantified by ImageJ software [Java 1.8.0_112 (64_bit)]. The
percentages of the MMP-2 and MMP-9 activities are expressed
relative to those in the vehicle control group, which was set to
100%.
Reverse transcription-quantitative
(RT-q)PCR
A549 cells were seeded in a 6-well plate at a
density of 5×105 cells/well in DMEM containing 10% FBS
and incubated overnight. The cells were treated with or without 20,
40 and 80 µM terrein and incubated for 24 h at 37°C. Total RNA was
isolated using RNeasy Mini Kit following manufacturer's
instructions (Qiagen, Inc.) and RNA concentration was measured
using NanoDrop spectrophotometer (Thermo Fisher Scientific, Inc.).
First strand cDNA was synthesized from 400 ng/µl RNA using the
Thermo Scientific RevertAid Reverse Transcriptase kit (Thermo
Fisher Scientific, Inc.). Quantitative real-time PCR was conducted
using a Chromo4™ Detection system (Bio-Rad Laboratories, Inc.)
according to cycling conditions outlined by the PCR array
manufacturer. All genes were detected (listed in the Table I) and analyzed through real-time PCR.
PCR cycle was performed with iTaq™ Universal SYBR Green mixture
(Bio-Rad Laboratories, Inc.) using the following cycling
conditions: 95°C for 2 min, followed by 40 cycles at 95°C for 15
sec, 61°C for 15 sec, 72°C for 1 min, and then 72°C for 2 min. Data
were analyzed using Optical Monitor 3 software (Bio-Rad
Laboratories, Inc.) and normalized to GAPDH mRNA expression.
Relative gene expression was quantified using the 2−ΔΔCq
method (31).
 | Table I.Sequence-specific primers for
qPCR. |
Table I.
Sequence-specific primers for
qPCR.
| Gene name | Primer sequence
(5′-3′) | Product size |
|---|
| MMP-2 | F:
CTCATCGCAGATGCCTGGAA | 104 |
|
| R:
TTCAGGTAATAGGCACCCTTGAAGA |
|
| MMP-9 | F:
ATCCGGCACCTCTATGGTC | 121 |
|
| R:
CTGAGGGGTGGACAGTGG |
|
| GAPDH | F:
ATCTTCTTTTGCGTCGCCAG | 51 |
|
| R:
TTCCCCATGGTGTCTGAGC |
|
VEGF determination
To examine whether terrein could inhibit angiogenic
processes, the function of VEGF was evaluated using the VEGF human
enzyme-linked immunosorbent assay (ELISA) kit (product code
ab100662; Abcam). Briefly, the cells were treated with or without
20, 40 and 80 µM terrein for 24 h. The supernatants were collected
and concentrated using Amicon Ultra-4 Centrifugal Filters 10K
(Thermo Fisher Scientific, Inc.). The protein concentrations were
measured using a Bradford protein assay, and each sample was
adjusted to the same concentration.
For the assay, as aforementioned, an ELISA kit
(product code ab100662; Abcam), with an antibody specific for human
VEGF was used to coat the wells of the plate prior to adding the
standards and protein samples, according to the manufacturer's
instructions. Then, the plate was incubated at 4°C overnight with
gentle shaking. The solution was discarded, and the plate was
washed 4 times with 1X wash solution. Then, 100 µl of 1X
biotinylated anti-human VEGF antibody was added to each well and
incubated for 1 h at room temperature. Subsequently, the unbound
biotinylated antibody was removed by washing, and 100 µl of 1X
HRP-conjugated streptavidin was added to each well and incubated
for 45 min at room temperature. Then, 100 µl of TMB substrate
solution was added and incubated for 30 min at room temperature in
the dark. The color developed in proportion to the bound VEGF in
each sample. The stop solution was added to each well, and the
color changed from blue to yellow. The intensity of the color was
immediately measured at 450 nm.
Tube formation assay
The tube formation assay is a rapid and quantitative
method for examining cell differentiation and changes in angiogenic
processes (32). A 24-well plate was
coated with 10 mg/ml Matrigel and incubated for 1 h at 37°C. The
remaining liquid was carefully removed from the culture plate
without disrupting the layer of Matrigel matrix. Then,
5×104 A549 cells in 1 ml of serum-free medium with or
without 20, 40 and 80 µM terrein were added to each well and
incubated for 24 h at 37°C. Tube formation was photographed by
inverted microscopy (Olympus CKX41), and the tubular structures in
five randomly selected fields were quantified using the
Angiogenesis Analyzer plugin for ImageJ software [Java 1.8.0_112
(64_bit)] (33). The percentage of
tube length was compared to that in the vehicle control group.
Western blot analysis
A549 cells were seeded in 60-mm culture dishes at a
density of 5×105 cells/dish in DMEM containing 10% FBS
and incubated overnight. The cells were treated with or without 20,
40 and 80 µM terrein and incubated for 24 h at 37°C. After
treatment, the cells were lysed with RIPA buffer (5 ml of 1 M
Tris-HCl pH 7.4, 30 ml of 5 M NaCl, 5 ml of 20% NP-40, 5 ml of 10%
sodium deoxycholate, 0.5 ml of 20% SDS, 50 ml of dH2O
and protease inhibitor cocktail) on ice. Subsequently, the cell
lysates were centrifuged at 15,000 g at 4°C for 10 min. The protein
concentration was measured using a Bradford protein assay. Equal
amounts (50 µg) of the total protein extracts were separated by 10%
SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred
to PVDF membranes. The membranes were blocked with 5% BSA in TBS-T
buffer (5% w/v BSA, 1X Tris-buffered saline solution and 0.1%
Tween-20) at room temperature for 1 h and then incubated with
primary antibodies against integrin αM (1:1,000; cat. no. sc-1186;
Santa Cruz Biotechnology, Inc.), total AKT (1:1,000; product no.
9272), pAKT (Ser473) (1:1,000; product no. 9271), pAKT (Thr308)
(1:1,000; product no. 4056), total FAK (1:1,000; product no. 3285;
all from Cell Signaling Technology, Inc.), pFAK (Tyr397) (1:1,000;
cat. no. 44-624G; Invitrogen™; Thermo Fisher Scientific, Inc.),
total mTOR (1:1,000; product no. 2972), pmTOR (Ser2448) (1:1,000;
product no. 2971), total P70S6 Kinase (1:1,000; product no. 2708),
pP70S6 Kinase (Thr389) (1:1,000; product no. 9234), total PI3K p85
(1:1,000; product no. 4292; all from Cell Signaling Technology,
Inc.), pPI3K p85 (Tyr458) (1:1,000; cat. no. PA5-17387;
Invitrogen™; Thermo Fisher Scientific, Inc.), total VEGFR2
(1:1,000; product no. 9698), pVEGFR2 (Tyr1175) (1:1,000; product
no. 3770), and GAPDH (1:1,000; product no. 2118; all from Cell
Signaling Technology, Inc.) at 4°C overnight. After washing 3 times
with TBS-T, the membranes were incubated with horseradish
peroxidase-conjugated anti-mouse (1:5,000; product no. 7076) or
anti-rabbit IgG antibodies (1:5,000; product no. 7074; both from
Cell Signaling Technology, Inc.) for 1 h. The membranes were
visualized by enhanced chemiluminescence using ECL plus™ western
blotting detection reagents (Bio-Rad Laboratories) and recorded on
gel documentation (UVITE,) or X-ray film. The intensities of the
protein bands were quantified by ImageJ software [Java 1.8.0_112
(64_bit)].
Immunofluorescence microscopy
A549 cells were seeded on round micro cover glasses
(VWR®) in 24-well plates at a density of
3×103 cells/well in DMEM containing 10% FBS and
incubated overnight. The cells were treated with or without 20, 40
and 80 µM terrein for 24 h. The cells were washed three times with
PBS++ (calcium and magnesium) and fixed with 4%
paraformaldehyde for 20 min at room temperature. After washing
three times with PBS++, the cells were permeabilized
with 0.1% Triton X-100 for 5 min at room temperature, and then, the
permeabilized cells were washed three times with PBS++.
The cells were blocked with blocking buffer (3% BSA in
PBS++) for 1 h at room temperature and then incubated
with a pFAK (Tyr397) polyclonal antibody (cat. no. 44-624G;
Invitrogen™; Thermo Fisher Scientific, Inc.) (1:200) at 4°C
overnight. Then, the cells were incubated with Alexa Fluor
488-conjugated goat anti-rabbit IgG (H+L) (1:800; cat. no. A27034;
Life Technologies; Thermo Fisher Scientific, Inc.) and rhodamine
phalloidin (1:320; cat. no. R415; Life Technologies; Thermo Fisher
Scientific, Inc.) for 1 h at room temperature. The stained cells
were washed three times with PBS++, and then, the cells
on glass coverslips were mounted onto microscope slides
(VWR®) in ProLong Gold Antifade Mountant (Life
Technologies; Thermo Fisher Scientific, Inc.) and
VECTASHIELD® Mounting Medium with DAPI (Vector Lab)
(1:1). The stained cells were observed with a Nikon A1R confocal
laser scanning microscope system (Nikon Corporation) with a 60X
objective.
Statistical analysis
All the experiments were performed at least in
triplicate in each group. The data are presented as the mean ± SEM
and were analyzed by GraphPad Prism version 5.01 software (GraphPad
Software, Inc.). Statistical significance was calculated using
ANOVA with Dunnett's multiple comparison post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Purification of terrein
The fungus Aspergillus terreus CRI301 was
cultured in Sabouraud dextrose agar under stationary conditions at
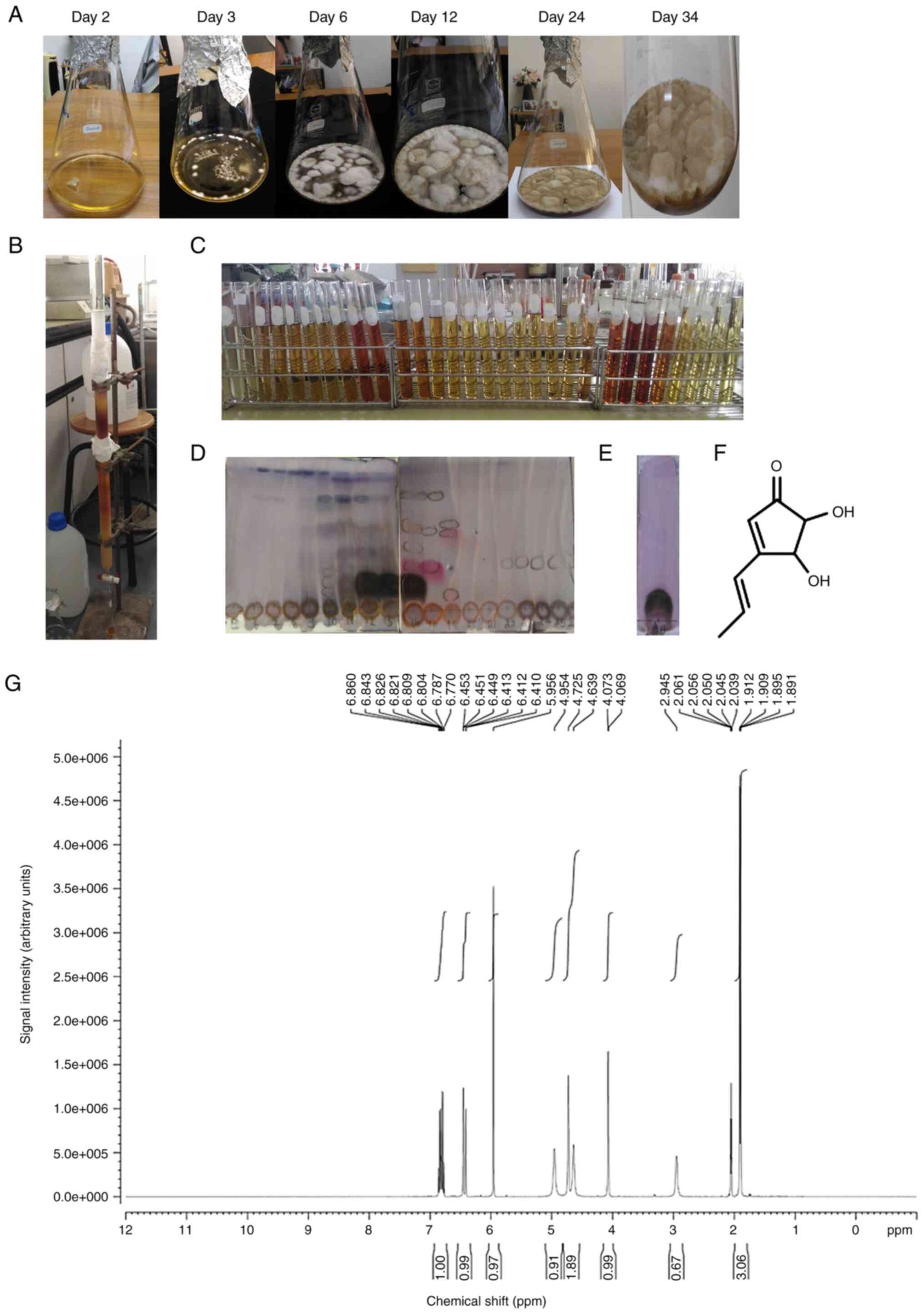
room temperature for 34 days. As revealed in Fig. 1A, the increased number of fungi was
associated with the days of culture. The culture (3.6 l) of
Aspergillus terreus CRI301 was filtered to separate the
cells from the broth. The culture broth was extracted three times
with an equal volume of EtOAc. The EtOAc layers were combined and
evaporated to dryness, yielding 2.535 g of crude extract. The crude
EtOAc extract was purified by Sephadex LH-20 column chromatography
and eluted with MeOH to yield 30 fractions (Fig. 1B). On the basis of their TLC
characteristics, similar fractions were combined to yield 10
fractions (Fig. 1C-E). Fraction 4 was
obtained as a pale yellow powder that was crystallized from
dichloromethane (CH2Cl2) and contained
terrein (258.7 mg) in the form of white needles. The structure of
terrein was characterized by 1H NMR spectroscopy and
finally confirmed by comparison with data in the literature
(Fig. 1F). Terrein was obtained in
the form of white needles. The 1H NMR spectrum of
terrein (Fig. 1G) exhibited three
methylene protons at δH 6.82 (1H,
m, H-2′), 6.43 (1H, d, J = 15.9 Hz, H-1′)
and 5.96 (1H, s, H-2), two oxygenated methine
protons at δH 4.73 (1H, s, H-4) and
4.07 (1H, s, H-5) and a methyl group at
δH 1.90 (3H, d, J = 15.9 Hz,
H-3′), as revealed in Table II.
| Table II.1H NMR (400 MHz) data of
terrein in Acetone-d6. |
Table II.
1H NMR (400 MHz) data of
terrein in Acetone-d6.
|
| Terrein |
|---|
|
|
|
|---|
| Position | δH
(ppm) |
|---|
| 1 | – |
| 2 | 5.96 (s) |
| 3 | – |
| 4 | 4.73
(1H, br s) |
| 5 | 4.07
(1H, br s) |
| 1′ | 6.43
(1H, d, J=15.9 Hz) |
| 2′ | 6.82
(1H, m) |
| 3′ | 1.90
(3H, d, J=15.9 Hz) |
Cytotoxic effects of terrein on the
viability and proliferation of lung cancer cells
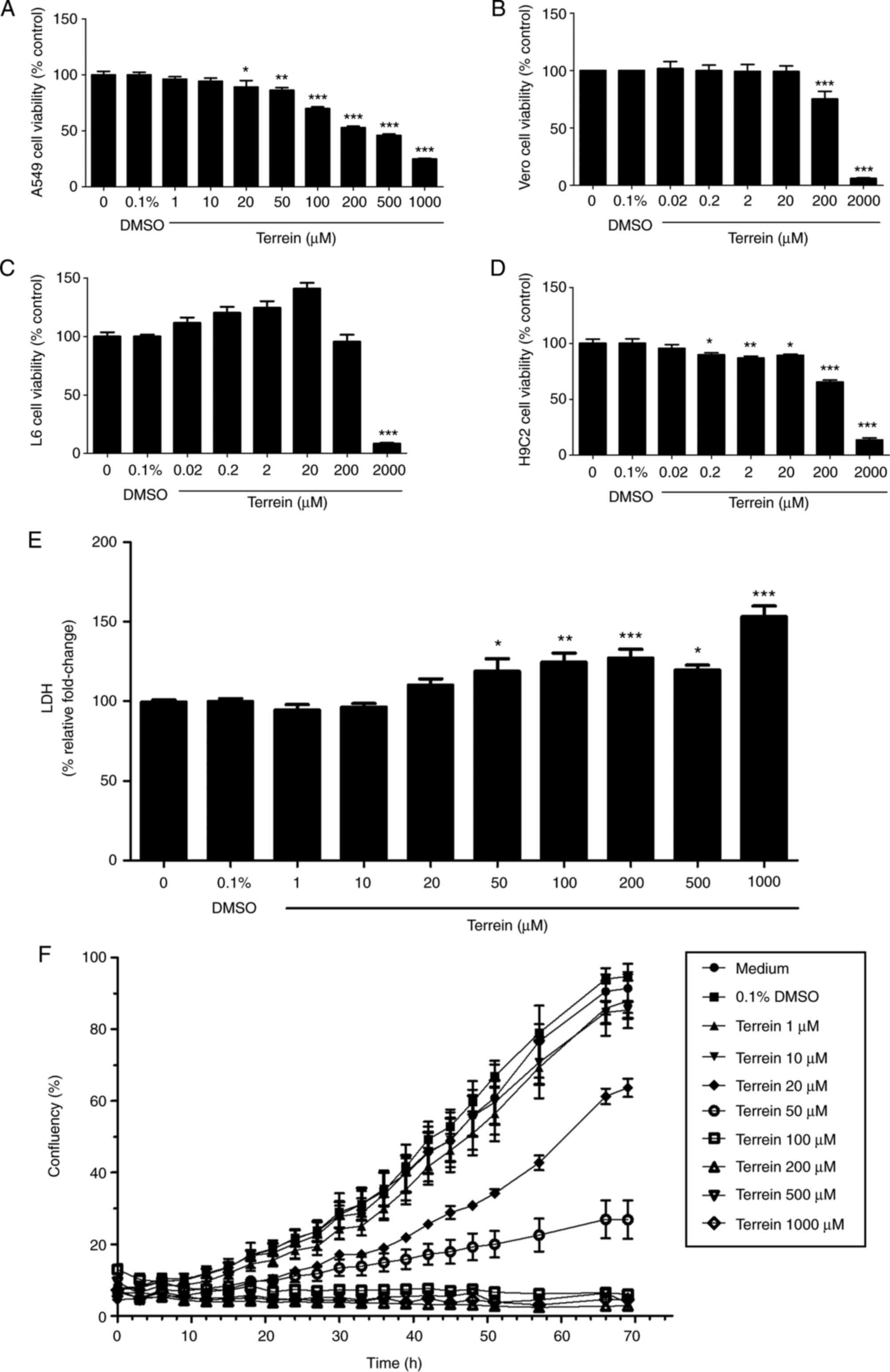
First, the cytotoxic effects of terrein on different
cell lines were examined, including A549 lung cancer cells, African
green monkey kidney (Vero) cells, L6 skeletal muscle cells and H9C2
cardiomyoblast cells, using an MTT assay. All the cell lines were
treated with various concentrations of terrein for 24 h, and the
maximum final concentration of DMSO (0.1%) was used as the vehicle
control. The results demonstrated that terrein significantly
inhibited the viability of A549 cells, Vero cells, L6 cells and
H9C2 cells with IC50 values of 229, 870, 1,240 and 579
µM, respectively (Fig. 2A-D). Terrein
exhibited more toxicity in lung cancer cells than in all the
representative normal cells. Moreover, the selective index (SI) of
terrein on A549 cells compared with that of terrein on all the
normal cell lines was calculated using the following equation:
SI=IC50 of normal cells/IC50 of A549 cells.
As a result, the SI values of terrein on A549 cells were 3.8, 5.4
and 2.5 compared to Vero cells, L6 cells and H9C2 cells,
respectively, indicating that terrein has high cytotoxic
selectivity against cancer cells compared with normal cells. In
addition, LDH assays were performed to confirm the damaging effect
of terrein. The LDH enzyme is normally used as a biomarker of
cellular cytotoxicity and cytolysis, as it is released from damaged
cells (34). As revealed in Fig. 2E, 50–1,000 µM terrein significantly
induced cytotoxicity in A549 cells.
It was also determined whether these cytotoxic
effects of terrein interfered with the process of cell
proliferation using the IncuCyte assay. Cells were treated with
different concentrations of terrein (1–1,000 µM). Cell
proliferation was monitored every 3 h for 3 days. Proliferation
curves were generated using IncuCyte proliferation analysis with
cell confluence as the parameter. The results revealed that
20–1,000 µM terrein exhibited dose-dependent inhibitory effects on
A549 cell proliferation, as revealed in Fig. 2F. The present results indicated that a
high concentration of terrein inhibited A549 cell viability and
proliferation by damaging the cells. Thus, to further investigate
the effects of terrein on metastatic processes, the concentrations
of 20, 40 and 80 µM terrein, which exhibit low levels of toxicity
in cells, were selected.
Terrein inhibits metastatic processes
of A549 lung cancer cells
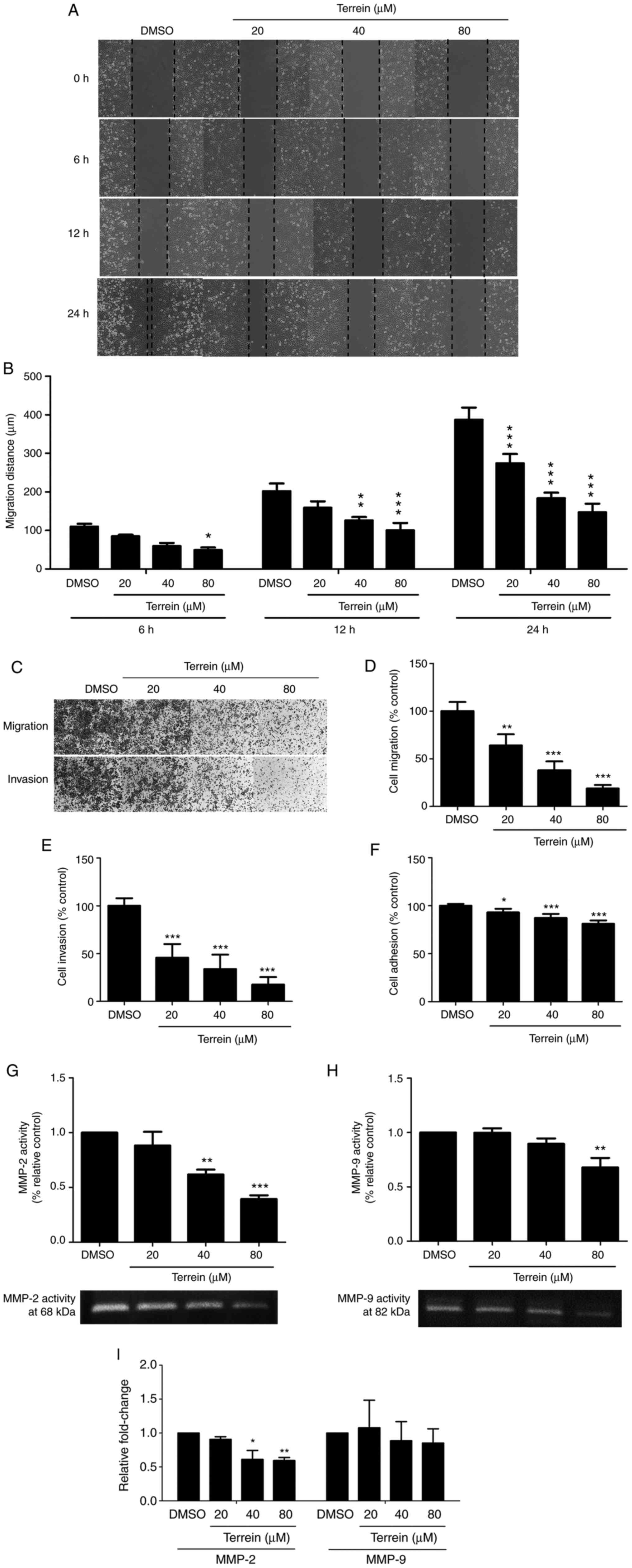
To determine the effect of terrein on the metastatic
processes of lung cancer cells, its inhibitory effect on lung
cancer cell migration was first evaluated. The migration of cells
was measured using a wound healing assay. In this method, the
concentrations of terrein that exhibited low toxicity (20, 40 and
80 µM) were used to treat A549 cells for 0, 6, 12 and 24 h. The
results revealed that terrein significantly inhibited cell
migration at 6, 12 and 24 h (Fig. 3A and
B).
The ability of terrein to suppress the migration and
invasion of A549 lung cancer cells was further examined. A549 cells
were treated with or without 20, 40 and 80 µM terrein for 24 h, and
a Transwell assay was used to observe the effect of terrein on A549
cell migration and invasion. The results revealed that terrein
significantly decreased the number of migrated and invasive cells
by as much as 78 and 82%, respectively, compared with the vehicle
control (Fig. 3C-E). The effect of
terrein on the adhesion process, since it is associated with the
early step of metastasis (35), was
also determined. As revealed in Fig.
3F, the highest concentration of terrein decreased A549 cell
adhesion by approximately 19% compared with the vehicle
control.
To assess the effect of terrein on the process of
cell invasion, the effect of terrein on the activities and
expression of MMP-2 and MMP-9 was determined using gelatin
zymography and qPCR, respectively. A549 lung cancer cells were
treated with 20, 40 and 80 µM terrein for 24 h. The results
revealed that terrein significantly suppressed the gelatinase
activities of both MMP-2 and MMP-9. Bands corresponding to MMP-2
(68 kDa) and MMP-9 (82 kDa) were clearly observed, and 40 and 80 µM
terrein significantly inhibited MMP-2 and 80 µM terrein
significantly inhibited MMP-9, as revealed in Fig. 3G and H, respectively. In addition,
terrein significantly inhibited MMP-2 expression and tended to
inhibit MMP-9 expression as revealed in the Fig. 3I. These data suggested that terrein
had the ability to inhibit the metastatic processes of lung cancer
cells.
Terrein inhibits the angiogenesis
process of A549 lung cancer cells
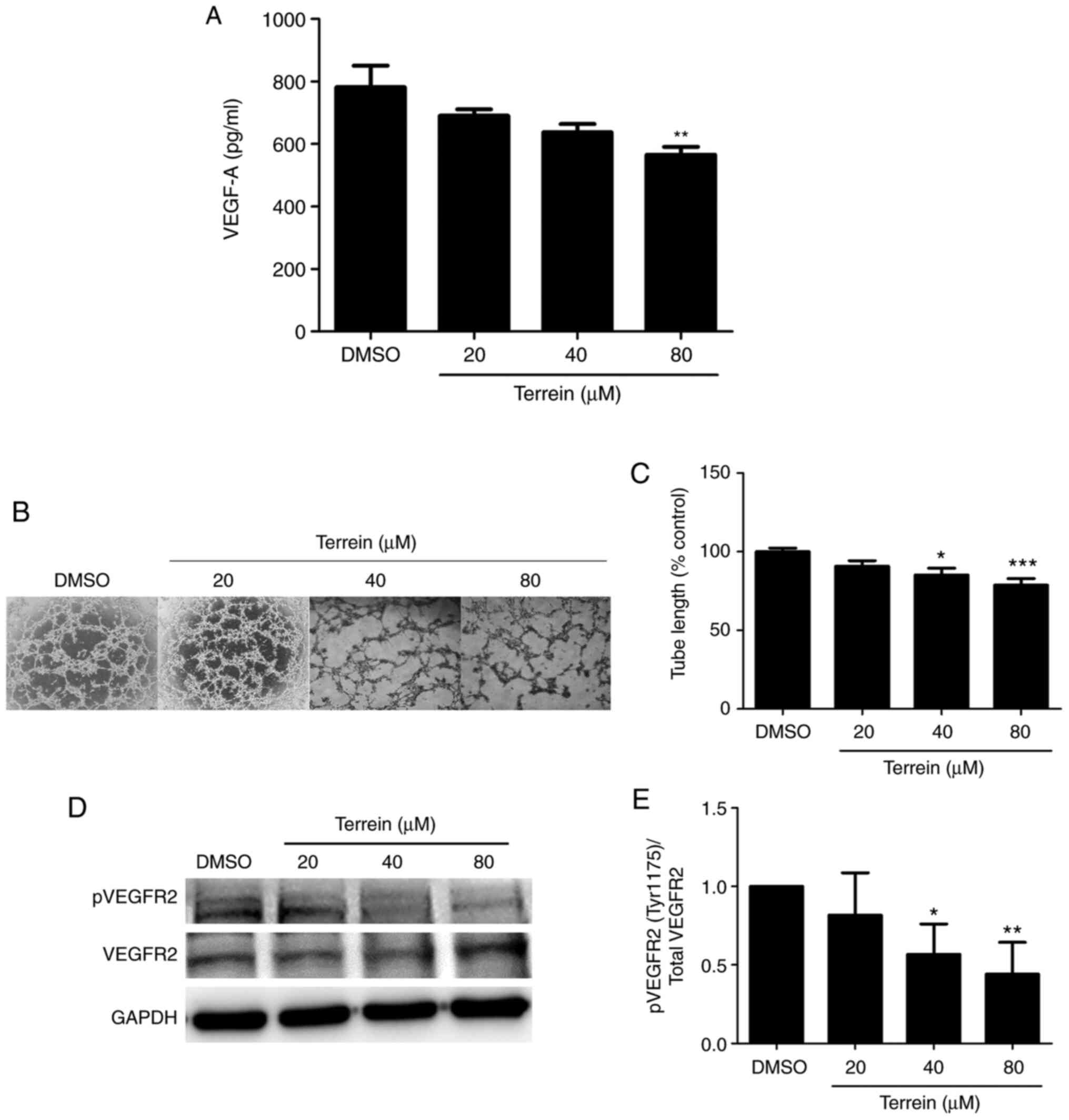
To examine whether terrein could inhibit the
angiogenesis process, A549 cells were treated with or without 20,
40 and 80 µM terrein for 24 h. The VEGF-A that was secreted by the
A549 cells into the medium was collected, concentrated and detected
by human VEGF-A ELISAs. The results revealed that terrein
significantly reduced the secretion of VEGF-A from A549 lung cancer
cells compared with the vehicle control (Fig. 4A). To further confirm this inhibitory
effect, an in vitro capillary-like tube formation assay was
performed. This is a rapid and quantitative method for examining
cell differentiation and changes associated with the angiogenesis
process. To perform the experiment, A549 cells were cultured in
Matrigel-coated plates and then treated with or without 20, 40 and
80 µM terrein for 24 h. Tube formation or the characteristics of
A549 cells that formed vessel-like channels were captured by
inverted microscopy. As revealed in Fig.
4B and C, terrein significantly suppressed the formation of
tube-like structures compared with the vehicle control. The
inhibition of VEGFR2 phosphorylation at Tyr1175, which is a crucial
site for the migration of cells during angiogenesis, was also
detected (Fig. 4D and E).
Terrein inhibits the activity of
metastasis mediators in A549 lung cancer cells
To investigate the effect of terrein on the
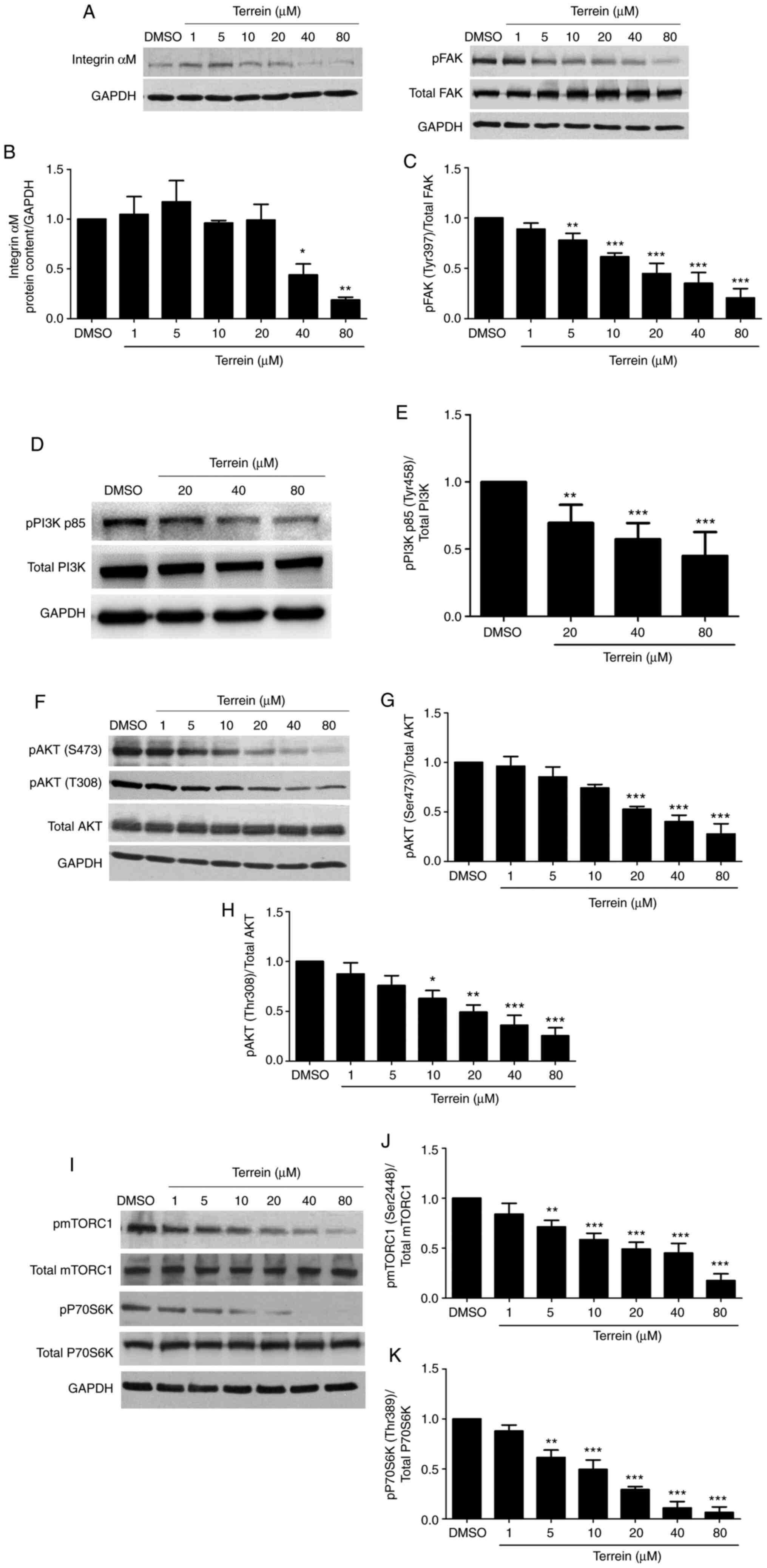
expression of metastasis mediators, such as integrin αM, FAK,
mTORC1, PI3K, AKT and P70S6K, western blotting was performed. A549
cells were treated with different concentrations of terrein for 24
h. As revealed in Fig. 5A-C, the
results indicated that terrein significantly inhibited integrin αM
expression and FAK phosphorylation at Tyr397 compared with the
vehicle control. Notably, a low concentration of terrein clearly
inhibited the phosphorylation of FAK. Active integrins recruit
various proteins, including FAK, to FAs to stimulate cell
signaling. When FAK is activated, it further stimulates the
expression and activation of downstream proteins. The signaling
proteins downstream of FAK are PI3K, AKT, mTORC1 and P70S6K. These
proteins affect cell survival, proliferation, angiogenesis,
invasion, migration and metastasis. The inhibition of FAK leads to
the suppression of these downstream signaling mediators (36). The phosphorylation of PI3K p85 at
Tyr458 (Fig. 5D and E), AKT at Ser473
(Fig. 5F and G), AKT at Thr308
(Fig. 5F and H), mTORC1 at Ser2448
(Fig. 5I and J) and pP70S6K at Thr389
(Fig. 5I and K) was decreased by
terrein in a dose-dependent manner compared with the vehicle
control. Therefore, the inhibition of these proteins by terrein
resulted in the suppression of metastatic and angiogenic processes
in lung cancer cells.
Terrein inhibits the phosphorylation
of FAK in A549 lung cancer cells
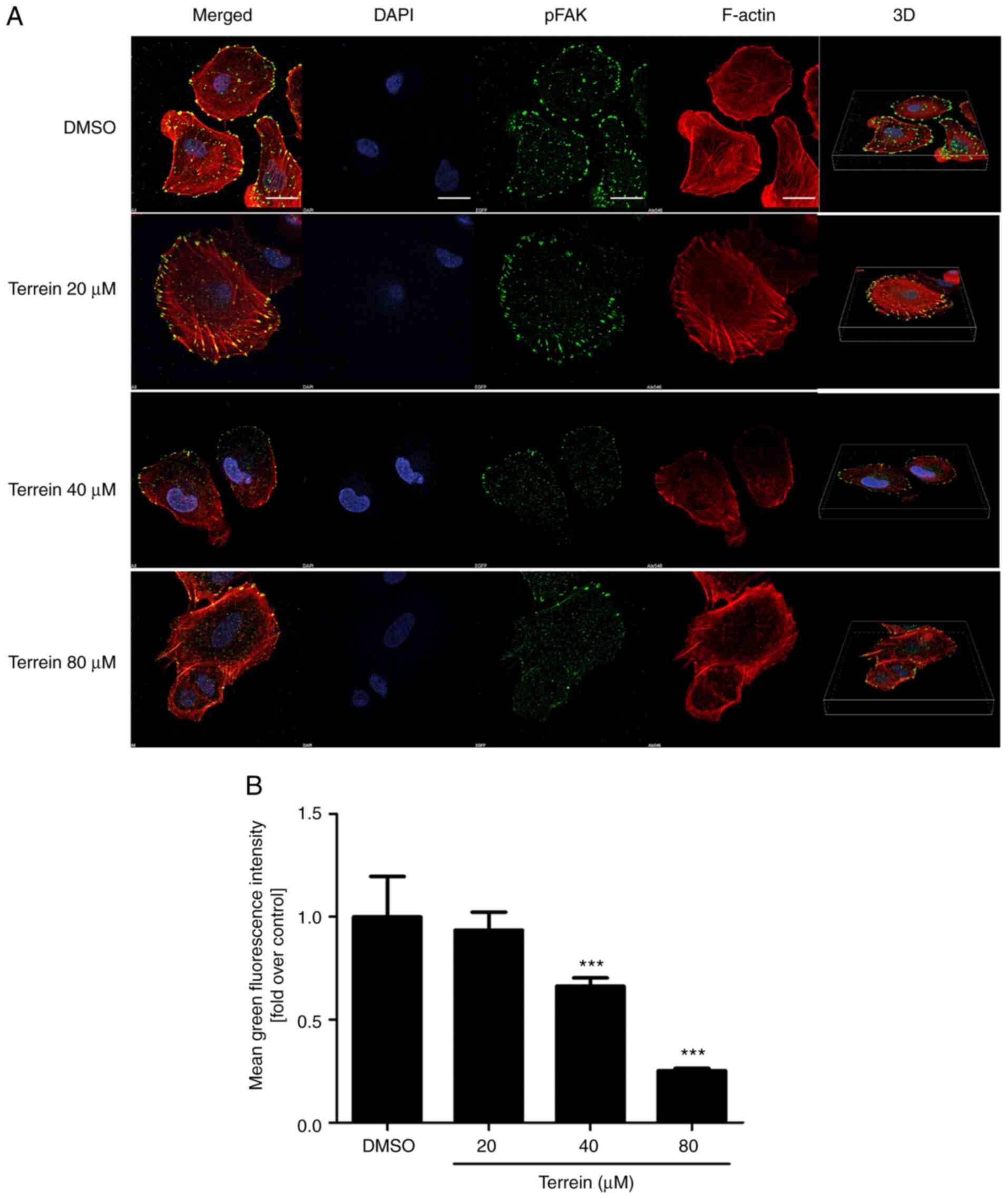
Immunofluorescence was used to further confirm the
effect of terrein on the phosphorylation of FAK at Tyr397 in A549
cells. The representative and quantitative images (Fig. 6) revealed that 40 and 80 µM terrein
significantly suppressed the phosphorylation of FAK at Tyr397
compared with the vehicle control. Combining these results with the
western blot data clearly indicated that terrein had the ability to
inhibit the phosphorylation of FAK at Tyr397, and FAK was revealed
to be an upstream regulator of numerous proteins involved in cancer
metastasis.
Discussion
Human lung cancer is the leading cause of
cancer-related deaths worldwide (1).
Most often, lung cancer is diagnosed at metastatic stages, and at
these stages, cancer cells have spread to nearby tissues or other
parts of the body (2). Therefore, the
suppression of metastasis is required to reduce the premature death
of cancer patients. Currently, the world has turned its attention
to the use natural substances for cancer treatment because they
typically have fewer side effects. Terrein is a bioactive natural
substance that has been reported to exert various biological
effects, including anti-inflammation, melanogenesis inhibition and
anticancer effects (20–22). However, to the best of our knowledge,
the effect of terrein on the molecular mechanisms that regulate
human lung cancer metastasis has not been examined.
In the present study, the effects of terrein on
metastatic processes, including cell proliferation, adhesion,
migration, invasion and angiogenesis, were investigated using A549
human lung cancer cells. Τhe present results revealed that terrein
exhibited cytotoxic effects on different cell types, including A549
cells, Vero cells, L6 cells and H9C2 cells, by inhibiting cell
viability with IC50 values of 229, 870, 1,240 and 579
µM, respectively. These results suggest that terrein has a specific
effect on A549 lung cancer cells relative to normal cells with a
relatively high selectivity index value. As revealed in other
studies, terrein exhibited a cytotoxic effect on human cervical
cancer cells (HeLa) with an IC50 value of 290 µM
(22) and on the human breast cancer
cell lines MCF-7 and MDA-MB-231 with IC50 values of
2,340 and 700 µM, respectively (28).
LDH assays were used to measure cell viability (34) and further verified this conclusion.
Overall, these data indicated that terrein was more toxic to A549
lung cancer cells than to other types of cancer cells.
Cancer metastasis includes several steps, including
cell proliferation, adhesion, migration, invasion and angiogenesis.
To determine the effects of terrein on these processes, low
concentrations of terrein (20, 40 and 80 µM) that were not toxic to
the normal cell lines tested were selected. It was first observed
that a terrein concentration as low as 20 µM started to reduce the
proliferative ability of A549 cells. The antimetastatic properties
of terrein were observed upon analysis of MMP activity. Numerous
studies have demonstrated that MMPs play important roles in tumor
progression, invasion, metastasis and angiogenesis (37,38).
Increased expression of MMPs has been revealed to be associated
with poor prognosis in several types of tumors, including breast
cancer, gastric cancer and osteosarcoma (39–41). All
invasive malignant tumors, including lung cancer cells, are known
to express high levels of MMPs, especially MMP-2 and MMP-9
(42). MMP-2 plays an essential role
in the progression of cancer because it cleaves several ECM
components and basement membranes (43); thus, the development of potential
MMP-2 inhibitors has become an important goal in lung cancer
therapy. The expression and activity of MMP-9 are increased in
NSCLC and are associated with the pathological type and clinical
stage of NSCLC (44). The expression
levels of MMP-9 were higher in advanced stage III and IV tumors
than in primary stage I and II tumors of NSCLC patients, and MMP-9
expression was higher in NSCLC patients with metastasis than in
NSCLC patients without metastasis and a reduced 5-year survival
rate (45). Both MMP-2 and MMP-9 are
classified as soluble enzymes that are secreted into the
extracellular milieu. Therefore, blocking the activities of these
enzymes has the potential to suppress lung cancer cell metastasis
and increase the possibility of NSCLC patient survival. In the
present study, it was revealed that terrein significantly
suppressed both the activities and expression of MMP-2 and MMP-9.
These findings indicated that terrein has the potential to act as
an antimetastatic agent, which, to the best of our knowledge, to
date, has never been reported.
Angiogenesis, or the creation of new blood vessels,
is another important mechanism during cancer progression. New blood
vessels support the growth of tumors by specifically feeding their
hypoxic and necrotic areas to provide essential nutrients and
oxygen (46). In the present study,
the effect of terrein on angiogenesis was investigated by
determining the expression of VEGF-A as a surrogate biomarker. VEGF
is a major chemotactic factor during angiogenesis that initiates
the migration and adhesion of cells, interactions between
endothelial cells and ECM, and formation of a tubular network
(47). In mammals, there are several
types of VEGF, including VEGF-A, VEGF-B, VEGF-C, and VEGF-D;
however, VEGF-A is widely studied and plays a major role in
angiogenesis by acting through VEGFR2 (12). It was observed that 80 µM terrein
significantly attenuated VEGF-A expression. Consistently, the ratio
of phosphorylated VEGF-A/VEGFR2 was decreased by terrein compared
to the vehicle control, thus indicating that the downstream
signaling events induced by the addition of VEGF to the cell medium
were inhibited. Furthermore, the effect of terrein on angiogenesis
was investigated by examining the in vitro capillary-like
tube formation of A549 cells. The characteristics of tube-like
structures were reduced by terrein in a dose-dependent manner.
These results suggest that terrein suppresses processes known to be
involved in angiogenesis.
Thus, our collective dataset suggests that terrein
can suppress multiple steps of cancer metastatic processes,
including proliferation, wound healing, adhesion, migration and
invasion. A more detailed mechanistic analyses was next conducted
based on the rationale that focal adhesion complexes are known to
be crucial nodes of signaling events that mediate cancer metastasis
(48). Notably, the interaction of
multiple proteins at focal adhesions is critical to promote the
protrusion of the cell membrane leading edge, resulting in the
development of invadopodia and lamellipodia (49). At focal adhesions, signal transduction
is initiated by interactions of integrins with the ECM, which
further promote the assembly of cytoplasmic scaffolds and the
recruitment of kinase proteins (50).
FAK, or focal adhesion kinase, is one of the principal integrin
signaling regulators that is recruited to the site of adhesion, and
FAK is autophosphorylated at the Tyr397 site. The
autophosphorylation of FAK contributes to the further activation of
its intrinsic kinase activity and creates docking sites for several
downstream signaling molecules (51).
FAK expression is increased in numerous highly malignant human
cancers (52). Overexpression of FAK
in cancer cells leads to resistance to the apoptotic process
(53). In addition, an increase in
FAK was revealed to contribute to the activation of the PI3K/AKT
and MEK-ERK1/2 signaling pathways, resulting in increased cancer
cell survival and proliferation (54). Therefore, to inhibit integrin
signaling, the regulation of FAK expression and phosphorylation is
a well-established goal for the development of effective
pharmaceutical antimetastatic agents. As revealed in the present
study, terrein could inhibit the expression of integrin αM and the
phosphorylation of FAK at Tyr397, as demonstrated by western blot
and immunofluorescence analyses. The present results revealed that
terrein could reduce stress fiber formation. Normally, cell
migration is associated with adhesion and actin cytoskeleton
organization, which also involves a series of lamellipodia
extension and actin polymerization. In addition, initial adhesions
are formed through the engagement of integrin receptors by the ECM
(55). The integrin-FAK signaling
pathway is known to be associated with cell adhesion and migration,
and these effects are regulated by downstream molecules. Inhibition
of FAK leads to retraction of filopodia and lamellipodia in cell
protrusions, resulting in stress fiber formation in retractile cell
bodies (6). The present results
suggested that the inhibition of FAK reduced the adhesion of cells
to the ECM, which contributed to reduced cell migration.
Next, mediators downstream of FAK, including PI3K,
AKT, mTOR and P70S6K, were further investigated. The signaling
pathway of PI3K/AKT is well recognized to associate with multiple
cellular processes such as cell proliferation, differentiation and
motility (56). In addition, mTOR is
one important downstream target of the PI3K/AKT signaling involved
with cell movement and metastasis (57). The protein mTORC1 induces protein
synthesis and cell growth by phosphorylating S6K1 and 4E-BP1
(58), while mTORC2 regulates the
organization of the actin cytoskeleton through F-actin stress
fibers, paxillin, RhoA, Rac1, Cdc42 and PKCα (59). The inhibition of the PI3K/AKT/mTOR
pathway by an mTOR inhibitor has been demonstrated to suppress
cancer cell invasion and migration and promote apoptosis in tumors
(60,61). Recent research has demonstrated that
inhibiting the FAK/PI3K/AKT/mTOR pathway and reducing MMP-2 and
MMP-9 protein expression suppressed gastric cancer cell migration
and invasion (62). Furthermore, the
PI3K/AKT/mTOR pathway plays a pivotal role in regulating
angiogenesis, increasing hypoxia-inducible factor 1α (HIF-1α)
translation and promoting VEGF production (63). The results of the present study
revealed that terrein significantly decreased the phosphorylation
of PI3K p85 at Tyr458, AKT at Ser473 and Thr308, mTORC1 at Ser2448
and pP70S6K at Thr389. Therefore, all these data indicated that
terrein has the ability to suppress the PI3K/AKT/mTOR/S6K1 pathway
and thus affects cancer cell proliferation, survival, migration,
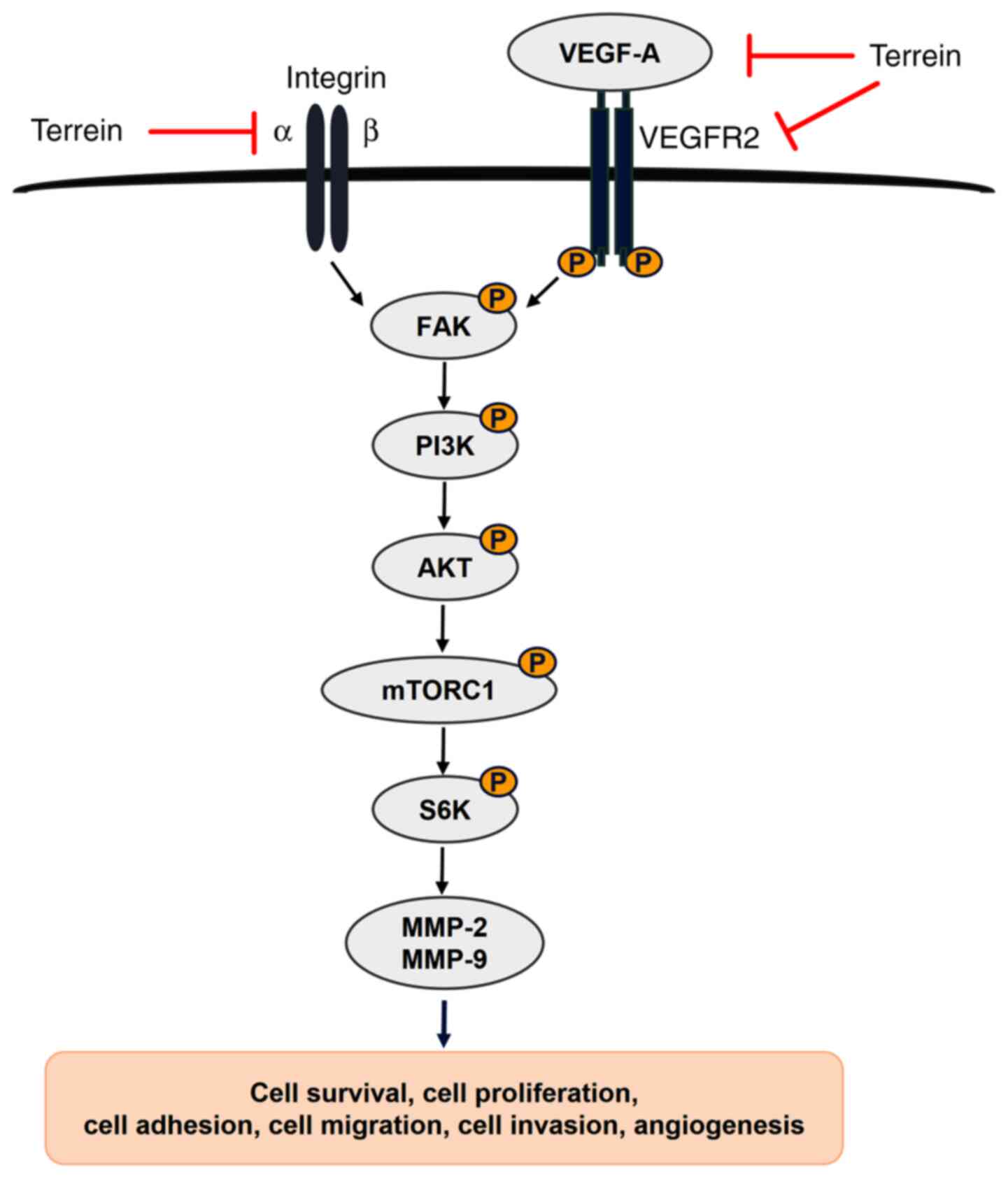
invasion, metastasis and angiogenesis, as revealed in Fig. 7.
In conclusion, the present study demonstrated the
novel finding that terrein acted as an anticancer agent in lung
cancer. This occured via numerous effects, such as the inhibition
of proliferation and metastatic processes, including adhesion,
migration and invasion. Mechanistically, terrein regulated the
integrin/FAK interaction and its downstream signaling pathway,
PI3K/AKT/mTOR/S6K1. In addition, terrein could inhibit angiogenic
processes by decreasing VEGF secretion and tube formation by
reducing the VEGF-A/VEGFR2 interaction. All these data suggested
that terrein is a potential new compound that is worthy of further
development as an anticancer agent. Further experiments to assess
the effect of terrein in vivo are further suggested.
Acknowledgments
We would like to thank Ms Vu Hong Loan Nguyen,
Department of Biology, York University, for sharing her laboratory
techniques for the IncuCyte assay.
Funding
This work was supported by research grants from the
Faculty of Medicine (grant nos. 140/2560 and 209/2562), Graduate
School and The Strategic Wisdom and Research Institute,
Srinakharinwirot University, Thailand, National Research Council of
Thailand: NRCT (2562) and Canada-ASEAN Scholarships and Educational
Exchanges for Development (2019–2020). Work in the GS lab was
funded by the National Science and Engineering Research Council
(NSERC), Canadian Institutes of Health Research (CIHR) and a Career
Investigator Award from the Heart and Stroke Foundation.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
PB performed the experiments and data analysis and
was the main author of the manuscript. WT and GS supervised the
experiments and revised the manuscript for important intellectual
content. MNA provided the materials needed for the preparation of
terrein and validated the data analysis. HKS performed the
immunofluorescence experiment and interpreted the data. CP
participated in the experimental design and provided the reagents
needed for the IncuCyte assay. All the authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Qi HW, Xin LY, Xu X, Ji XX and Fan LH:
Epithelial-to-mesenchymal transition markers to predict response of
Berberine in suppressing lung cancer invasion and metastasis. J
Transl Med. 12:222014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leber MF and Efferth T: Molecular
principles of cancer invasion and metastasis (Review). Int J Oncol.
34:881–895. 2009.PubMed/NCBI
|
|
4
|
Hamidi H and Ivaska J: Every step of the
way: Integrins in cancer progression and metastasis. Nat Rev
Cancer. 18:533–548. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maziveyi M and Alahari SK: Cell matrix
adhesions in cancer: The proteins that form the glue. Oncotarget.
8:48471–48487. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu H, Gao M, Ma Y, Wang L, Shen Y and Liu
X: Inhibition of cell migration by focal adhesion kinase:
Time-dependent difference in integrin-induced signaling between
endothelial and hepatoblastoma cells. Int J Mol Med. 41:2573–2588.
2018.PubMed/NCBI
|
|
7
|
Mitra SK, Hanson DA and Schlaepfer DD:
Focal adhesion kinase: In command and control of cell motility. Nat
Rev Mol Cell Biol. 6:56–68. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khan KH, Yap TA, Yan L and Cunningham D:
Targeting the PI3K-AKT-mTOR signaling network in cancer. Chin J
Cancer. 32:253–265. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu L, Li F, Cardelli JA, Martin KA,
Blenis J and Huang S: Rapamycin inhibits cell motility by
suppression of mTOR-mediated S6K1 and 4E-BP1 pathways. Oncogene.
25:7029–7040. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fingar DC, Richardson CJ, Tee AR, Cheatham
L, Tsou C and Blenis J: mTOR controls cell cycle progression
through its cell growth effectors S6K1 and 4E-BP1/eukaryotic
translation initiation factor 4E. Mol Cell Biol. 24:200–216. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Olsson AK, Dimberg A, Kreuger J and
Claesson-Welsh L: VEGF receptor signalling - in control of vascular
function. Nat Rev Mol Cell Biol. 7:359–371. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dimova I, Popivanov G and Djonov V:
Angiogenesis in cancer-general pathways and their therapeutic
implications. J buon. 19:15–21. 2014.PubMed/NCBI
|
|
13
|
Rydén L, Linderholm B, Nielsen NH, Emdin
S, Jönsson PE and Landberg G: Tumor specific VEGF-A and VEGFR2/KDR
protein are co-expressed in breast cancer. Breast Cancer Res Treat.
82:147–154. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tanno S, Ohsaki Y, Nakanishi K, Toyoshima
E and Kikuchi K: Human small cell lung cancer cells express
functional VEGF receptors, VEGFR-2 and VEGFR-3. Lung Cancer.
46:11–19. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hamerlik P, Lathia JD, Rasmussen R, Wu Q,
Bartkova J, Lee M, Moudry P, Bartek J Jr, Fischer W, Lukas J, et
al: Autocrine VEGF-VEGFR2-Neuropilin-1 signaling promotes glioma
stem-like cell viability and tumor growth. J Exp Med. 209:507–520.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu X and Zhou W: The emerging regulation
of VEGFR-2 in triple-negative breast cancer. Front Endocrinol
(Lausanne). 6:1592015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Le Boeuf F, Houle F and Huot J: Regulation
of vascular endothelial growth factor receptor 2-mediated
phosphorylation of focal adhesion kinase by heat shock protein 90
and Src kinase activities. J Biol Chem. 279:39175–39185. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Koch S, Tugues S, Li X, Gualandi L and
Claesson-Welsh L: Signal transduction by vascular endothelial
growth factor receptors. Biochem J. 437:169–183. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Raistrick H and Smith G: Studies in the
biochemistry of micro-organisms: The metabolic products of
Aspergillus terreus Thom. a new mould metabolic
product-terrein. Biochem J. 29:606–611. 1935. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee JC, Yu MK, Lee R, Lee YH, Jeon JG, Lee
MH, Jhee EC, Yoo ID and Yi HK: Terrein reduces pulpal inflammation
in human dental pulp cells. J Endod. 34:433–437. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park SH, Kim DS, Kim WG, Ryoo IJ, Lee DH,
Huh CH, Youn SW, Yoo ID and Park KC: Terrein: A new melanogenesis
inhibitor and its mechanism. Cell Mol Life Sci. 61:2878–2885. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Porameesanaporn Y,
Uthaisang-Tanechpongtamb W, Jarintanan F, Jongrungruangchok S and
Wongsatayanon B: Terrein induces apoptosis in HeLa human cervical
carcinoma cells through p53 and ERK regulation. Oncol Rep.
29:1600–1608. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang F, Mijiti M, Ding W, Song J, Yin Y,
Sun W and Li Z-Y: (+)-Terrein inhibits human hepatoma Bel-7402
proliferation through cell cycle arrest. Oncol Rep. 33:1191–1200.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen YF, Wang SY, Shen H, Yao XF, Zhang FL
and Lai D: The marine-derived fungal metabolite, terrein, inhibits
cell proliferation and induces cell cycle arrest in human ovarian
cancer cells. Int J Mol Med. 34:1591–1598. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liao WY, Shen CN, Lin LH, Yang YL, Han HY,
Chen JW, Kuo SC, Wu SH and Liaw CC: Asperjinone, a nor-neolignan,
and terrein, a suppressor of ABCG2-expressing breast cancer cells,
from thermophilic Aspergillus terreus. J Nat Prod.
75:630–635. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shibata A, Ibaragi S, Mandai H, Tsumura T,
Kishimoto K, Okui T, Hassan NM, Shimo T, Omori K, Hu GF, et al:
Synthetic terrein inhibits progression of head and neck cancer by
suppressing angiogenin production. Anticancer Res. 36:2161–2168.
2016.PubMed/NCBI
|
|
27
|
Arakawa M, Someno T, Kawada M and Ikeda D:
A new terrein glucoside, a novel inhibitor of angiogenin secretion
in tumor angiogenesis. J Antibiot (Tokyo). 61:442–448. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kasorn A, Loison F, Kangsamaksin T,
Jongrungruangchok S and Ponglikitmongkol M: Terrein inhibits
migration of human breast cancer cells via inhibition of the Rho
and Rac signaling pathways. Oncol Rep. 39:1378–1386.
2018.PubMed/NCBI
|
|
29
|
Jahng JWS, Alsaadi RM, Palanivel R, Song
E, Hipolito VEB, Sung HK, Botelho RJ, Russell RC and Sweeney G:
Iron overload inhibits late stage autophagic flux leading to
insulin resistance. EMBO Rep. 20:e479112019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sung HK, Song E, Jahng JWS, Pantopoulos K
and Sweeney G: Iron induces insulin resistance in cardiomyocytes
via regulation of oxidative stress. Sci Rep. 9:46682019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee H and Kang KT: Advanced tube formation
assay using human endothelial colony forming cells for in vitro
evaluation of angiogenesis. Korean J Physiol Pharmacol. 22:705–712.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carpentier G, Berndt S, Ferratge S,
Rasband W, Cuendet M, Uzan G and Albanese P: Angiogenesis analyzer
for ImageJ - A comparative morphometric analysis of ‘endothelial
tube formation assay’ and ‘fibrin bead assay’. Sci Rep.
10:115682020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kumar P, Nagarajan A and Uchil PD:
Analysis of cell viability by the lactate dehydrogenase assay. Cold
Spring Harb Protoc. 2018.2018. View Article : Google Scholar
|
|
35
|
Bendas G and Borsig L: Cancer cell
adhesion and metastasis: Selectins, integrins, and the inhibitory
potential of heparins. Int J Cell Biol. 2012:6767312012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ata R and Antonescu CN: Integrins and cell
metabolism: An intimate relationship impacting cancer. Int J Mol
Sci. 18:1892017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McCawley LJ and Matrisian LM: Matrix
metalloproteinases: Multifunctional contributors to tumor
progression. Mol Med Today. 6:149–156. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Stetler-Stevenson WG: The role of matrix
metalloproteinases in tumor invasion, metastasis, and angiogenesis.
Surg Oncol Clin N Am. 10:383–392. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li H, Zhang K, Liu LH, Ouyang Y, Bu J, Guo
HB and Xiao T: A systematic review of matrix metalloproteinase 9 as
a biomarker of survival in patients with osteosarcoma. Tumour Biol.
35:5487–5491. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Uchibori M, Nishida Y, Nagasaka T, Yamada
Y, Nakanishi K and Ishiguro N: Increased expression of
membrane-type matrix metalloproteinase-1 is correlated with poor
prognosis in patients with osteosarcoma. Int J Oncol. 28:33–42.
2006.PubMed/NCBI
|
|
41
|
Shen W, Xi H, Wei B and Chen L: The
prognostic role of matrix metalloproteinase 2 in gastric cancer: A
systematic review with meta-analysis. J Cancer Res Clin Oncol.
140:1003–1009. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guo CB, Wang S, Deng C, Zhang DL, Wang FL
and Jin XQ: Relationship between matrix metalloproteinase 2 and
lung cancer progression. Mol Diagn Ther. 11:183–192. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Houghton AM: Matrix metalloproteinases in
destructive lung disease. Matrix Biol. 44-46:167–174. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
El-Badrawy MK, Yousef AM, Shaalan D and
Elsamanoudy AZ: Matrix metalloproteinase-9 expression in lung
cancer patients and its relation to serum mmp-9 activity,
pathologic type, and prognosis. J Bronchology Interv Pulmonol.
21:327–334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zheng S, Chang Y, Hodges KB, Sun Y, Ma X,
Xue Y, Williamson SR, Lopez-Beltran A, Montironi R and Cheng L:
Expression of KISS1 and MMP-9 in non-small cell lung cancer and
their relations to metastasis and survival. Anticancer Res.
30:713–718. 2010.PubMed/NCBI
|
|
46
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Niu G and Chen X: Vascular endothelial
growth factor as an anti-angiogenic target for cancer therapy. Curr
Drug Targets. 11:1000–1017. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mirza AA, Kahle MP, Ameka M, Campbell EM
and Cuevas BD: MEKK2 regulates focal adhesion stability and
motility in invasive breast cancer cells. Biochim Biophys Acta Mol
Cell Res. 1843:945–954. 2014. View Article : Google Scholar
|
|
49
|
Machesky LM: Lamellipodia and filopodia in
metastasis and invasion. FEBS Lett. 582:2102–2111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Aoudjit F and Vuori K: Integrin signaling
in cancer cell survival and chemoresistance. Chemother Res Pract.
2012:2831812012.PubMed/NCBI
|
|
51
|
Frame MC, Patel H, Serrels B, Lietha D and
Eck MJ: The FERM domain: Organizing the structure and function of
FAK. Nat Rev Mol Cell Biol. 11:802–814. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cance WG, Harris JE, Iacocca MV, Roche E,
Yang X, Chang J, Simkins S and Xu L: Immunohistochemical analyses
of focal adhesion kinase expression in benign and malignant human
breast and colon tissues: Correlation with preinvasive and invasive
phenotypes. Clin Cancer Res. 6:2417–2423. 2000.PubMed/NCBI
|
|
53
|
Tai YL, Chen LC and Shen TL: Emerging
roles of focal adhesion kinase in cancer. Biomed Res Int.
2015:6906902015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Bouchard V, Demers M-J, Thibodeau S,
Laquerre V, Fujita N, Tsuruo T, Beaulieu J-F, Gauthier R, Vézina A,
Villeneuve L and Vachon PH: Fak/Src signaling in human intestinal
epithelial cell survival and anoikis: Differentiation
state-specific uncoupling with the PI3-K/Akt-1 and MEK/Erk
pathways. J Cell Physiol. 212:717–728. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Burridge K and Guilluy C: Focal adhesions,
stress fibers and mechanical tension. Exp Cell Res. 343:14–20.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Jiang N, Dai Q, Su X, Fu J, Feng X and
Peng J: Role of PI3K/AKT pathway in cancer: The framework of
malignant behavior. Mol Biol Rep. 47:4587–4629. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Brader S and Eccles SA: Phosphoinositide
3-kinase signalling pathways in tumor progression, invasion and
angiogenesis. Tumori. 90:2–8. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dowling RJO, Topisirovic I, Fonseca BD and
Sonenberg N: Dissecting the role of mTOR: Lessons from mTOR
inhibitors. Biochim Biophys Acta. 1804:433–439. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhou H and Huang S: Role of mTOR signaling
in tumor cell motility, invasion and metastasis. Curr Protein Pept
Sci. 12:30–42. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shih YW, Chen PS, Wu CH, Jeng YF and Wang
CJ: Alpha-chaconine-reduced metastasis involves a PI3K/Akt
signaling pathway with downregulation of NF-kappaB in human lung
adenocarcinoma A549 cells. J Agric Food Chem. 55:11035–11043. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wu YJ, Lin SH, Din ZH, Su JH and Liu CI:
Sinulariolide inhibits gastric cancer cell migration and invasion
through downregulation of the EMT process and suppression of
FAK/PI3K/AKT/mTOR and MAPKs signaling pathways. Mar Drugs.
17:6682019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Karar J and Maity A: PI3K/AKT/mTOR pathway
in angiogenesis. Front Mol Neurosci. 4:512011. View Article : Google Scholar : PubMed/NCBI
|