Introduction
The high rates of growth and proliferation that
characterize cancer cells require a very active anabolic metabolism
to abundantly synthesize the different macromolecules constituting
structural and functional cellular components (1,2). Oncogenic
mutations, the increased usage of exogenous nutrients and the
activation of specific metabolic pathways are all factors that
reciprocally influence each other in the rewiring of cancer cell
metabolism (3).
Elevated glucose uptake and high levels of
glycolysis, despite the presence of oxygen, were established as
features of cancer cells in the middle of the last century, when
Otto Warburg described these phenomena in a seminal paper (4). Glycolytic intermediates are diverted
towards anabolic pathways for the biosynthesis of nucleotides,
amino acids and lipids, thus increasing the synthesis of
macromolecule building blocks (5).
Glutamine uptake and metabolism are also frequently boosted in
cancer cells. Glutamine catabolism provides anaplerotic
intermediates for the tricarboxylic acid cycle (6), as well as precursors for the
biosynthesis of nucleotides, all the non-essential amino acids
(NEAAs), glutathione (GSH), one of the major cellular antioxidant
defenses, and ATP energy (7,8).
Since several oncogenes control the expression of
metabolic genes, oncogenic alterations are important players in
cancer cell metabolic reprogramming (9,10).
Activated c-Myc, for example, upregulates genes involved in
glucose or glutamine uptake and catabolism (11). In addition, the constitutive growth
program driven by oncogenic c-Myc makes cells depend on
exogenous nutrients to survive, in particular glutamine, despite it
being a NEAA that cells can synthesize (12). Glutamine is synthesized from glutamate
and ammonia by the enzyme glutamine synthetase (GS; also known as
glutamate ammonia ligase, GLUL) (7).
In some cancer cell lines, c-Myc can also indirectly upregulate GS
expression, promoting the expression of a demethylase acting on its
promoter (13). GS levels can also be
post-transcriptionally regulated by glutamine levels (14).
c-Myc expression itself can be modulated by nutrient
levels, both at the transcriptional level and also during
translation (15–20). Hann et al (15) showed that cell growth at a high
density, in exhausted culture medium or in the absence of
methionine, altered c-Myc translation initiation. In these
conditions, c-Myc synthesis was induced from an in-frame CUG
translation initiation site located in the 5-untranslated region,
giving rise to a c-Myc isoform with 15 additional amino acids. To
date, contrasting results have been obtained on the possible
function of this non-canonical c-Myc isoform, known as c-Myc 1,
with contradictory data showing either a cell growth inhibitory
function of c-Myc 1 (21) or
indistinguishable properties, compared with canonical c-Myc
(22). Recently, Sato et al
(23) showed that the two c-Myc
isoforms had different transcriptional targets, with canonical
c-Myc specifically promoting the expression of genes associated
with cell growth and transformation (23).
Given the role of glucose and glutamine in cancer
cell growth, these nutrients and their metabolism are under
investigation as possible targets for cancer cell therapy (24–26).
Hence, it is important to identify factors that can counteract the
detrimental effects of nutrient shortage and thus prevent
therapeutic results. Recently, it has been shown that asparagine
can sustain cell survival in the absence of glutamine by
suppressing C/EBP homologous protein induction and apoptosis
following glutamine starvation (27).
Moreover, asparagine has been found capable of supporting
proliferation in the absence of glutamine, allowing a basal level
of protein synthesis higher than in glutamine-deprivation
conditions, particularly by increasing the synthesis of GS, and
hence endocellular glutamine levels (28).
In our laboratory, we developed a cellular system
(cen3tel) that recapitulates neoplastic transformation of human
fibroblasts (29–33). The analysis of gene and microRNA
expression of cells at different phases of the transformation
process suggested the occurrence of metabolic reprogramming during
transformation, which was particularly associated with c-Myc
overexpression (30,32). The deep cellular and molecular
characterization of these cells makes them a suitable and valuable
tool with which to study cancer cell metabolism. In our previous
study, the cellular response to glutamine or glucose deprivation in
cen3tel cells was studied and it was shown that tumorigenic cells
had become glutamine-addicted (34).
In fact, in the absence of exogenous glutamine, cells underwent a
prolonged growth arrest (72–96 h) and then began to die. By
contrast, glucose deprivation led to cell death more quickly
(within 48 h) and induced autophagy. In the present study, cen3tel
cells and other cancer cell lines were used to study the
relationship between nutrient deprivation and c-Myc expression,
along with the role of asparagine in counteracting the detrimental
effects of glutamine starvation. The results suggested an
association between c-Myc expression and the asparagine capacity to
allow cell proliferation in the absence of glutamine, possibly
through a c-Myc-mediated increase in GS levels.
Materials and methods
Cell lines and culture
In the present study, cen3tel cells at ~1,000
population doublings (PDs) after telomerase immortalization were
used. These cells are tumorigenic and metastatic in nude mice and
carry a mutation in the TP53 codon 161, which has been used
to confirm the identity of the cen3tel cells used in the present
study (30,35).
The following cells were used in the present study:
Cen3tel, MDA-MB-231 (human metastatic breast cancer), U2OS (human
osteosarcoma), A375MC2 [high metastatic melanoma cell line
(36); a generous gift from Dr
Richard Hynes, Howard Hughes Medical Institute, Center for Cancer
Research, Cambridge, MA, USA), SW480 (human colon adenocarcinoma)
and HeLa (human cervix carcinoma). Cells were cultured as
previously described (34). Cells
were counted using Burker chambers. Stock solutions of single NEAAs
(Merck KGaA) were prepared by dissolving aspartic acid and glutamic
acid in PBS (at a concentration of 25 and 50 mM, respectively), and
alanine, asparagine and proline in distilled water (at a
concentration of 150 mM). Solutions were then sterilized by
filtration. To carry out nutrient deprivation experiments, cells
were plated in complete medium [except for the experiments with
methionine sulfoximine (MSO), see below] and when they had reached
a density of ~2-3×105 cells/cm2, they were
processed as previously described (34). In the experiments in which cells were
propagated in different culture conditions, cells were plated in
complete medium and after 24 h they were incubated for an
additional 24 h in the absence of glutamine and presence of the
desired NEAAs. Then cells were collected, counted (first passage)
and replated in a defined number in a medium with the same
composition as that in which they were cultured. Cell harvesting
was repeated three additional times every 48 or 72 h (second, third
and fourth passage). For each culture condition, the number of
cells obtained at each passage was used to calculate the number of
PDs/day performed by the cells between each passage, which was
considered as a measure of cell growth in the different media.
Experiments were repeated at least three times with three
replicates, unless otherwise specified.
Treatments
The MG132 (Merck KGaA) stock solution was prepared
in DMSO at a concentration of 25 mM and kept at −20°C. Cells were
exposed to 25 µM MG132 during the last 2 or 4 h of incubation in
the different nutrient conditions.
The puromycin (Merck KGaA) stock solution was
prepared in sterile water at a concentration of 12 mM and was then
sterilized by filtration and stored at −20°C. During the last 10
min of incubation in the different nutrient conditions, cen3tel and
MDA-MB-231 cells were incubated with 90 or 2.5 µM puromycin,
respectively.
The MSO (Thermo Fisher Scientific, Inc.) stock
solution was prepared in sterile water at a concentration of 200
mM, filtered and stored at −20°C. MSO treatment was performed by
plating cells directly in the different nutrient-containing media
with or without 2 mM MSO. Cells were collected after 96 or 144 h of
incubation. The medium was refreshed 96 h after incubation.
The 10074-G5 (Cayman Chemical Company) stock
solution was prepared in DMSO at a concentration of 20 mM and
stored at −20°C. Cells were exposed to 10074-G5 during the entire
incubation period in the different nutrient conditions.
The reduced GSH (Merck KGaA) stock solution was
freshly prepared in DMEM for each experiment at a concentration of
100 mM. Twenty-four hours after plating, cells were incubated in
the different nutrient conditions with or without 5 mM GSH (Merck
KGaA) for a further 24 h. Growth was determined by calculating the
number of PDs/day performed by each cell sample in the time
interval between plating and cell harvesting.
When drugs were dissolved in DMSO, control cells
were treated with the same DMSO concentration reached in
drug-exposed samples.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted using QIAzol reagent (Qiagen
GmbH). cDNA was generated from 1 µg RNA using the QuantiTec Reverse
Transcription Kit (Qiagen GmbH). Gene expression was quantified
using SYBR®-Green chemistry with a QuantiTect SYBR Green
kit (Qiagen GmbH) using the QuantiTect Primer Assay specific for
each target gene (Hs_MYC_1_SG; Hs_GLUL_1_SG, Hs_GUSB_1_SG; Qiagen
GmbH). qPCR was performed on the Light Cycler 480 (Roche
Diagnostics), using 96-well reaction plates (Roche Diagnostics).
Raw data were normalized to the GusB housekeeping gene and
the relative expression was calculated using the ΔΔCq method
(37) and expressed as fold change
(FC)=2−∆ΔCq. qPCR analysis was carried out at least
three times using different biological replicates.
Western blotting
Whole-cell lysates for western blot analysis were
prepared using RIPA lysis buffer [1% Nonidet P-40, 50 mM Tris-HCl
(pH 8.0), 150 mM NaCl, 0.1% SDS, 0.1% DOC, 1X protease inhibitor
cocktail (Thermo Fisher Scientific, Inc.), and 1X phosphatase
inhibitor cocktail (Roche Diagnostics)]. The following antibodies
recognizing human proteins were used: Anti-c-Myc rabbit monoclonal
(cat. no. ab32072; 1:10,000; Abcam), anti-puromycin mouse
monoclonal (cat. no. MABE343; 1:10,000; Merck KGaA), anti-GS mouse
monoclonal (cat. no. 610517; 1:250; BD Transduction Laboratories),
anti-poly(ADP-ribose) polymerase 1 (PARP1) rabbit monoclonal (cat.
no. ab191217; 1:1,000; Abcam), anti-poly(ADP-ribose) (PAR) chains
mouse monoclonal (cat. no. sc-56198; 1:1,000; Santa Cruz
Biotechnology, Inc.), anti-vinculin mouse monoclonal (cat. no.
05–386; 1:5,000; Merck KGaA), anti-γ-tubulin mouse monoclonal (cat.
no. T6557; 1:10,000; Merck KGaA), anti-β-actin mouse monoclonal
(cat. no. A2066; 1:10,000; Merck KGaA). Incubations with primary
antibodies were carried out overnight at 4°C. Primary antibodies
were probed by a secondary horseradish peroxidase-conjugated
antibody (anti-mouse, cat. no. 115-035-146; anti-rabbit, cat. no.
111-035-144; 1:5,000; Jackson ImmunoResearch Laboratories, Inc.)
for 1 h at room temperature. Protein bands were visualized using an
enhanced chemiluminescent reagent (Clarity™; Bio-Rad Laboratories,
Inc.). The intensity of the band in each sample was quantified
using QuantityOne software (Bio-Rad Laboratories, Inc.), corrected
for the intensity of the corresponding band obtained with the
anti-γ-tubulin, anti-vinculin, or anti-actin antibodies, and then
normalized to the appropriate control sample.
Western blotting experiments were repeated at least
three times and with independent biological replicates.
Representative images of the results are shown in the figures.
Statistical analysis
Statistical analysis was performed using ANOVA
single factor for comparison of means of multiple groups followed
by Bonferroni post-hoc test, and two-tailed Students t-test for
comparison between two groups (Microsoft Excel, version 16.16.24
was used). P<0.05 was considered to indicate a statistically
significant difference.
Results
c-Myc expression is altered in cells
starved of glutamine and/or glucose
Given the profound effects of glutamine and
or/glucose deprivation on cell growth and viability in cen3tel
cells (34) and the association
between c-Myc and metabolism, the expression of this
oncogene was analyzed in the different starved samples.
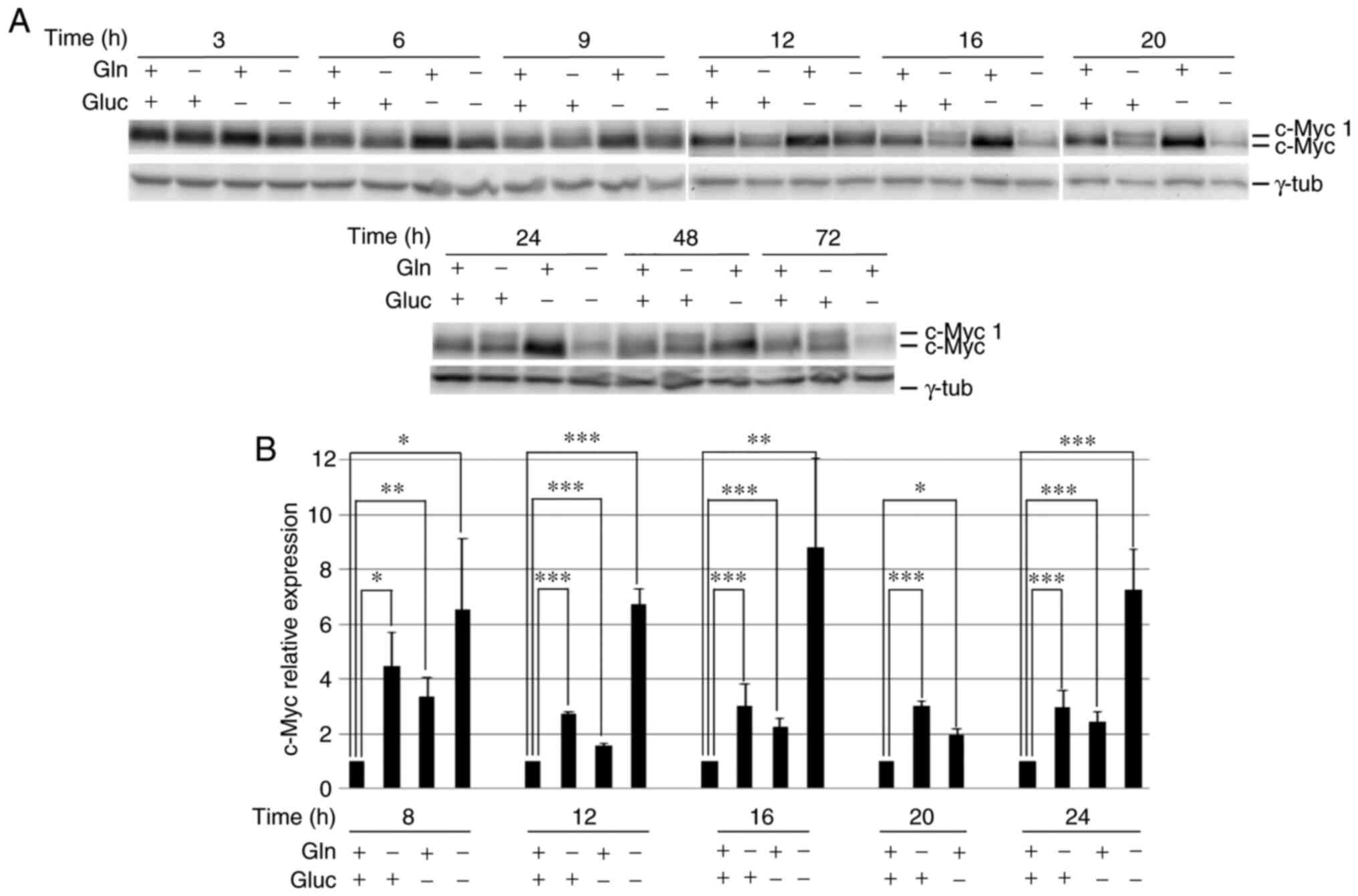
The deprivation of each nutrient caused a rapid and
specific change in c-Myc protein expression (Fig. 1A). In glucose-starved specimens, c-Myc
levels were higher than in control cells 6–48 h after starvation
and then declined (Fig. 1A), when
cells started detaching from the culture dish and died (34).
Glutamine starvation led to a decrease in the levels
of canonical c-Myc and the synthesis of the longer c-Myc isoform,
c-Myc 1 (Fig. 1A), which was
described by Hann et al (15)
in cells grown in harsh culture conditions or in the absence of
methionine. This suggested that, in the absence of glutamine, c-Myc
synthesis can start from the cryptic CUG translation initiation
site. c-Myc 1 expression began to appear 6 h after glutamine
deprivation; 12 h after starvation, the two c-Myc isoforms were
almost equally represented and together gave a c-Myc level similar
to that observed in control cells (at 12, 16 and 20 h, the levels
of the two isoforms together vs. c-Myc in control cells, was 0.8,
0.7 and 1.1, respectively; Fig.
1A).
Double-deprived cells had a mixed behavior (Fig. 1A); at 9–12 h after deprivation, c-Myc
expression was higher than in control cells, but the c-Myc 1 band
was also detectable. At subsequent incubation times, c-Myc levels
were found to be diminished. Of note, at that stage,
double-deprived cells had detached from the culture dish and
started dying (34).
c-Myc expression was analyzed at the RNA
level in samples starved of the different nutrients for 8–24 h by
RT-qPCR (Fig. 1B). In all the
conditions and at all the time points, c-Myc RNA levels were
higher in starved cells compared with control cells, regardless of
the protein level. In particular, in glutamine-starved cells, the
higher RNA expression was not accompanied with a higher c-Myc
protein level, indicating a translational impairment.
Characterization of c-Myc 1 expression
in glutamine-starved cells
The synthesis of the two c-Myc isoforms in the
absence of glutamine indicated a link between glutamine
availability and c-Myc translation from the cryptic CUG initiation
site, which, to the best of our knowledge, has not been described
so far. Thus, the present study aimed to further characterize c-Myc
1 expression in the absence of glutamine. First, it was
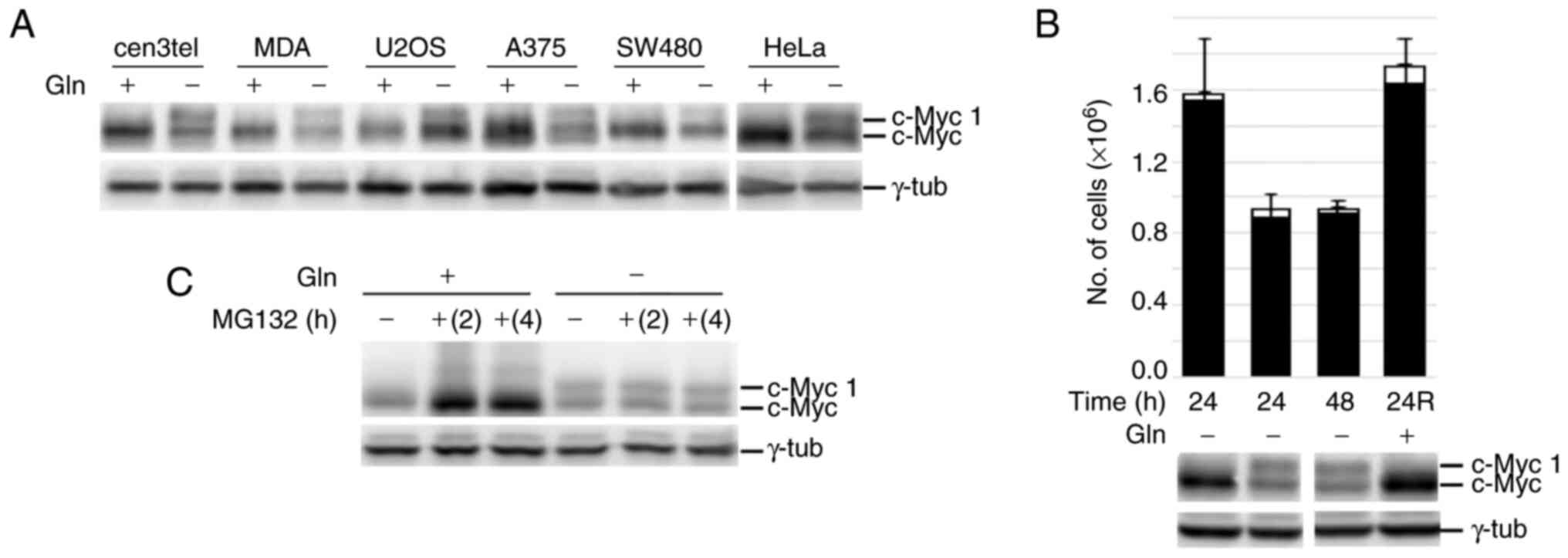
investigated whether c-Myc 1 was present in other cancer cell lines
(namely MDA-MB-231, U2OS, A375MC2, SW480 and HeLa) cultured in the
absence of glutamine. As shown in Fig.
2A, the association between glutamine deprivation and c-Myc 1
expression was not restricted to cen3tel cells. In fact, 24 h after
glutamine starvation, c-Myc 1 was expressed in a nearly 1:1 ratio
with canonical c-Myc in MDA-MB-231 and HeLa cells. In the other
cell lines, c-Myc 1 expression was induced in the absence of
glutamine, but this isoform represented a lower fraction of the
total c-Myc compared with that in cen3tel, MDA-MB-231 and HeLa
cells.
Using cen3tel cells, c-Myc 1 expression was further
characterized in the absence of glutamine. It was shown that the
increase in c-Myc 1 synthesis was reversible when glutamine levels
were restored. In fact, c-Myc 1 became undetectable when cen3tel
cells starved of glutamine for 24 h were fed with complete medium
for 24 h (Fig. 2B). Glutamine
supplementation also led to the recovery of cellular proliferation
(Fig. 2B).
Since c-Myc undergoes a rapid turnover, being
degraded by the proteasome, it was examined whether the two c-Myc
isoforms were characterized by the same turnover. Thus, 24
h-glutamine-starved cen3tel cells were exposed to the MG132
proteasomal inhibitor for 2 or 4 h, and the same results were
reported for both isoforms, that is, a compromised accumulation of
both isoforms in the presence of the proteasomal inhibitor
(Fig. 2C). This suggested that 24 h
after glutamine deprivation, the synthesis and degradation of both
c-Myc isoforms were impaired.
In glutamine-starved cells, asparagine
is necessary and sufficient to maintain c-Myc expression just as in
the presence of glutamine and allows cell survival and
proliferation
Given that glutamine is a key precursor for the
biosynthesis of NEAAs (7,8), it was analyzed whether single NEAAs,
namely alanine, asparagine, aspartic acid, glutamic acid and
proline (serine and glycine were not tested, as they are present in
the culture medium), or a pool of them, could compensate for the
absence of glutamine in the culture medium for both c-Myc synthesis
and cell growth.
As shown in Fig. 3A,
in cen3tel cells incubated with either alanine, aspartic acid,
glutamic acid or proline, the same pattern of c-Myc expression as
that in glutamine-starved cells was observed. By contrast,
following asparagine supplementation, the pattern of c-Myc
expression was the same as that in control cells, with the
expression of c-Myc 1 almost undetectable and high levels of
canonical c-Myc (~2–3 times higher than in glutamine-starved
cells). Asparagine was effective at a concentration of 0.1 mM,
which is close to the physiological concentration of this amino
acid (38). The relevance of
asparagine on c-Myc expression was confirmed by the observation
that in glutamine-starved cells incubated in medium supplemented
with the pool of the 5 NEAAs, c-Myc was synthesized as in the
presence of asparagine only, while in cells grown with a pool of
NEAAs devoid of asparagine, c-Myc was expressed as in
glutamine-starved cells. Thus, asparagine is necessary and
sufficient to maintain canonical c-Myc synthesis in the absence of
glutamine and to promote c-Myc translation from the canonical
initiation site. The same result was obtained in MDA-MB-231 cells,
which expressed high levels of c-Myc 1 in the absence of glutamine,
but not in the presence of asparagine (Fig. S1A).
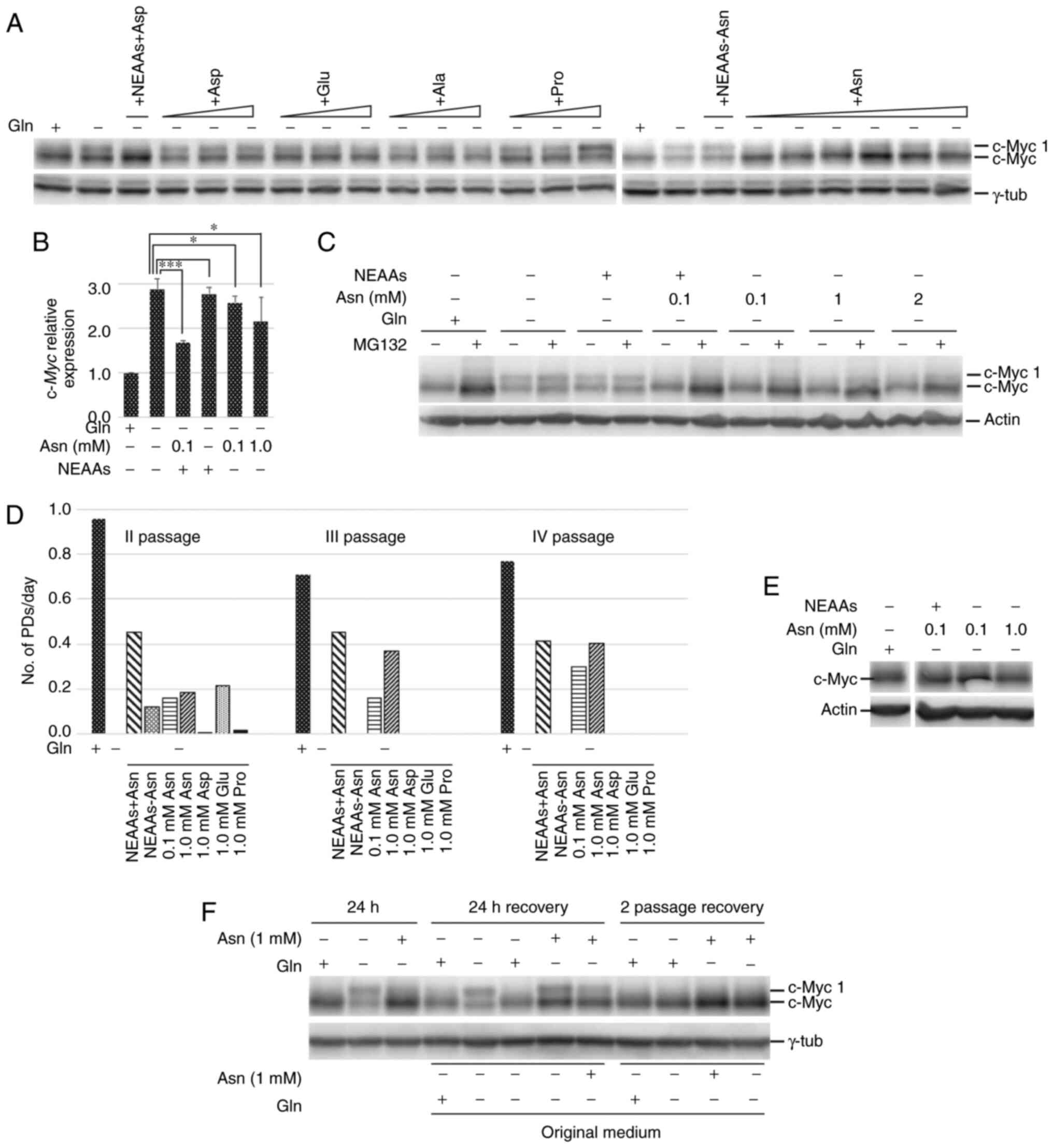 | Figure 3.Asparagine is necessary and
sufficient to maintain c-Myc expression as in the presence of
glutamine and allows cell survival and proliferation. NEAAs, a pool
of alanine, aspartic acid, glutamic acid and proline at a
concentration of 0.1 mM each, with or without 0.1 mM asparagine. In
the western blots, γ-tubulin or actin were used as the loading
control. (A) Western blot analysis of c-Myc expression in cen3tel
cells cultured for 24 h in the indicated medium; for each NEAA,
increasing concentrations were tested, for alanine, aspartic acid,
glutamic acid and proline, concentrations were 0.1, 0.5 and 1.0 mM,
for asparagine 0.1, 0.5, 1.0, 1.5 and 2.0 mM. (B) RT-qPCR of
c-Myc expression in cen3tel cells cultured for 24 h in the
indicated medium. The expression of c-Myc in cells cultured
in the different culture conditions is shown as Log2FC
relative to c-Myc expression in cells cultured in the
presence of glutamine. Error bars: Standard deviation. *P<0.05,
***P<0.005. (C) Western blot analysis of c-Myc expression in
cells cultured for 24 h in the indicated medium and incubated with
25 µM MG132 during the last 2 h of culture. (D) Cen3tel cell
propagation in different media. In the histogram, each column
represents the number of PDs/day performed by the cells in the
different culture conditions, starting from the second passage. An
exemplificative experiment is shown. (E) Western blot analysis of
c-Myc expression in cells propagated in the absence of glutamine
and in the presence of 0.1 or 1.0 mM asparagine for four passages.
(F) Western blot analysis of c-Myc expression in cen3tel cells
incubated for 24 h in the medium indicated above the lanes (first
three lanes) and then fed with different media for 24 h (lanes, 24
h recovery) or for two passages (lanes, 2 passage recovery). The
incubation medium during the recovery period is indicated above the
lanes, the original medium in which each cell sample was incubated
for the first 24 h is indicated below the lanes. Gln, glutamine;
Ala, alanine; Asn, asparagine; Asp, aspartic acid; Glu, glutamic
acid; Pro, proline; NEAAs, non-essential amino acids; RT-qPCR,
reverse transcription-quantitative PCR. |
As far as c-Myc transcription is concerned,
in cen3tel cells incubated in the presence of asparagine and in the
absence of glutamine, there were increased levels of c-Myc
RNA compared with control cells, but decreased levels compared with
glutamine-starved cells (Fig. 3B).
When cells were incubated with a pool of the 4 NEAAs (alanine,
aspartic acid, glutamic acid and proline), no changes in
c-Myc RNA levels were observed compared with
glutamine-starved cells, while, when asparagine was added to the
pool, c-Myc transcription was decreased (Fig. 3B). These results suggested that, in
the absence of glutamine but presence of asparagine, c-Myc
expression was still upregulated at the transcriptional level and,
being the RNA efficiently translated, gave rise to high levels of
canonical c-Myc.
In glutamine-starved cen3tel cells, asparagine
supported a c-Myc turnover comparable to that observed in control
cells, while a pool of alanine, aspartic acid, glutamic acid and
proline did not allow c-Myc accumulation in the absence of
glutamine when the proteasome was inhibited by MG132 (Fig. 3C).
Considering the effect of NEAAs on c-Myc expression,
it was further analyzed whether NEAAs, and in particular
asparagine, had an influence on cell growth in the absence of
glutamine. As previously mentioned, glutamine-starvation at 24–72 h
led to cen3tel cell growth arrest. When cells were incubated for
these periods of time in the absence of glutamine and in the
presence of NEAAs, either singly or in pools, no significant
differences were detected in the cell number between
glutamine-starved and NEAA-supplemented cells (Fig. S2). Longer-term experiments were
therefore performed, in which cells were propagated in the
different experimental conditions for four passages. As shown in
Fig. 3D, cells grown in the absence
of glutamine or in glutamine-free media supplemented with proline
or aspartic acid did not grow between the first and the second
passages. At the third passage, most cells were floating in the
medium. In the other culture conditions, cells showed an increase
in cell number relative to seeding, but decreased numbers compared
with the control cells. At the subsequent passages (III and IV),
only asparagine supplementation (either alone or in combination
with the other NEAAs) allowed cell growth in the absence of
glutamine, although at a lower rate compared with cells grown in
glutamine-containing medium. Therefore, asparagine is necessary and
sufficient for cell growth in the absence of glutamine.
The analysis of c-Myc expression in cells collected
at the fourth passage confirmed that glutamine-starved cells
cultured in the presence of asparagine expressed high levels of
canonical c-Myc, similar to cells propagated in complete medium
(Fig. 3E).
Finally, it was examined whether asparagine was able
to rescue canonical c-Myc expression in cells that had been starved
for glutamine. As expected, 24 h after glutamine deprivation
(Fig. 2B and 3F), the addition of glutamine allowed a
complete recovery of canonical c-Myc expression in cen3tel cells;
cells re-fed with medium without glutamine continued to exhibit an
equal expression of the two c-Myc isoforms, while cells re-fed with
asparagine still expressed c-Myc 1, but showed an increase in
canonical c-Myc levels (Fig. 3F). In
fact, the ratios between the intensities of the c-Myc and c-Myc 1
bands were 1.1 in the sample re-fed without glutamine, and 2.1 in
that re-fed with asparagine. When cells re-fed with asparagine were
maintained in culture for two passages, the pattern of c-Myc
expression became indistinguishable from that observed in cells
always maintained in medium with glutamine or asparagine, or re-fed
with glutamine following glutamine deprivation (Fig. 3F).
Thus, in glutamine-starved cells, asparagine allows
high levels of canonical c-Myc synthesis, cell survival and
proliferation, although at lower rates compared with the presence
of glutamine.
GSH prevents PARP1 activation in
glutamine-starved asparagine-supplemented cells
In our previous study, glutamine deprivation was
shown to lead to PARP1 activation, with high levels of PARP1
auto-(poly-ADP) ribosylation (PARylation) and protein PARylation
(34). In fact, compared with cells
grown in the presence of glutamine, in cells deprived of glutamine
for 24 h, the anti-PARP1 antibody revealed a reduced intensity of
the PARP1 band and an increased series of higher molecular weight
bands (Fig. 4A); this was paralleled
by an intense signal highlighted by the antibody against PAR
chains, which confirmed the activation of PARP1 in these cells
(Fig. 4A). Herein, PARP1 was also
found to be activated when glutamine-starved cen3tel cells were
supplemented with asparagine; in fact, in these cells, the signals
obtained with either the PARP1 antibody or the anti-PAR chain
antibody were comparable to those obtained in glutamine-starved
cells (Fig. 4A). This suggested that
asparagine was not sufficient to prevent cellular stress, due to
the absence of glutamine, which could contribute to the lower
proliferation rate observed in cells cultured without glutamine but
in the presence of asparagine.
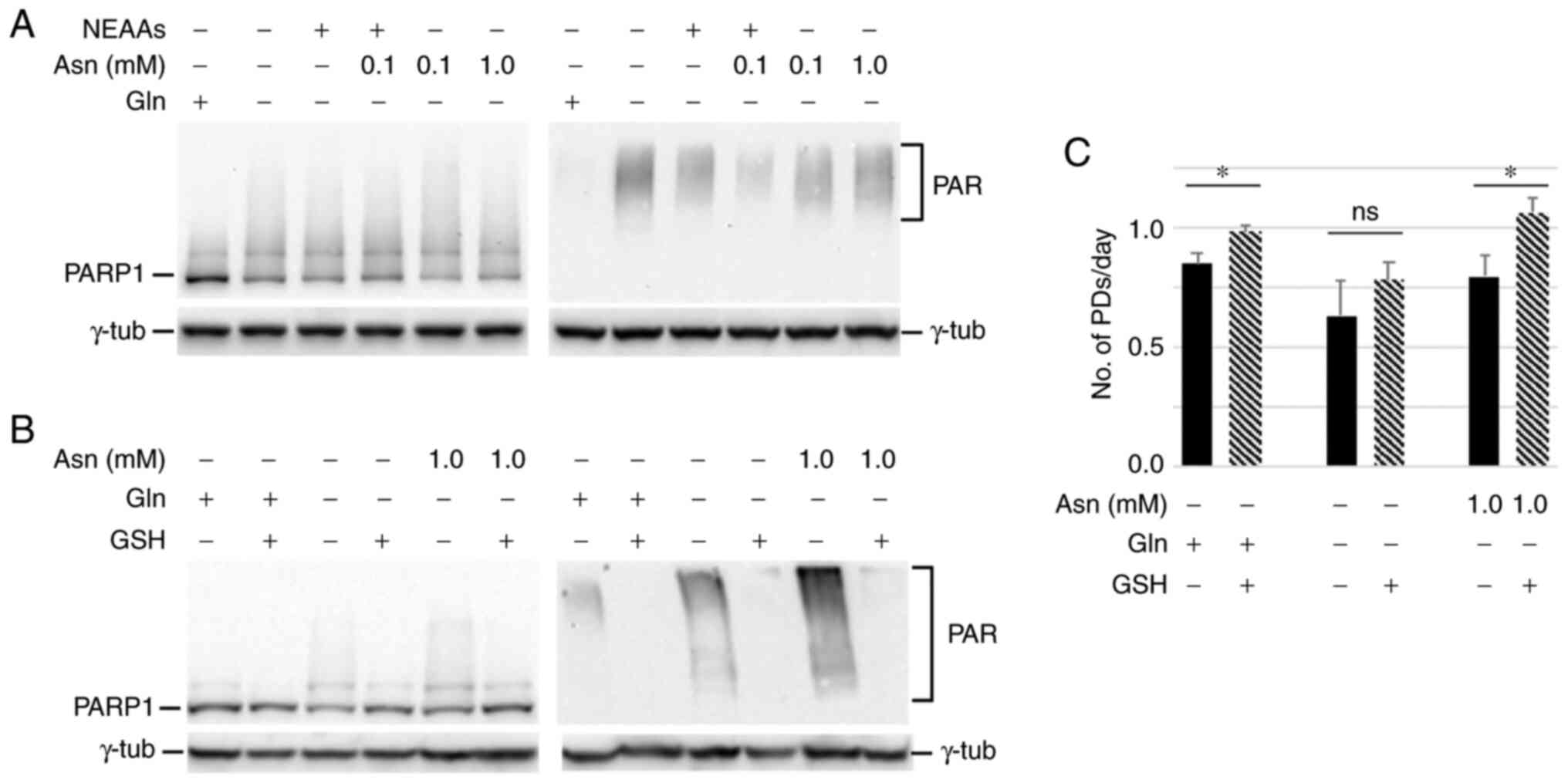 | Figure 4.GSH rescues PARP1 activation in
glutamine-starved asparagine-supplemented cells. NEAAs, a pool of
alanine, aspartic acid, glutamic acid and proline at a
concentration of 0.1 mM each. (A) Western blot analysis of PARP1
and protein PARylation with an antibody against PAR chains in cells
incubated in the indicated medium for 24 h. (B) GSH was used at 5
mM during the entire incubation period. γ-tubulin was used as the
loading control. (C) Cell growth in the indicated media plus or
minus GSH. Each column represents the number of PDs/day performed
by the cells in the different culture conditions between plating
and harvesting. Error bars, standard deviations of two independent
experiments. *P<0.05; ns, not significant. Gln, glutamine; Asn,
asparagine; GSH, glutathione; NEAAs, non-essential amino acids;
PARP1, poly(ADP-ribose) polymerase 1; PAR, poly(ADP-ribose); PDs,
population doublings. |
Given that glutamine is essential for GSH synthesis
(7,8),
it was examined whether the addition of GSH to the culture medium
could decrease PARP1 activation in the absence of glutamine.
Indeed, it was found that GSH supplementation reduced both the
PARP1 smeared signal and protein PARylation (Fig. 4B) and increased cellular proliferation
(Fig. 4C). This suggested that
oxidative stress plays a role in blocking cell growth in the
absence of glutamine, and that asparagine alone is not able to
counteract this phenomenon.
In asparagine-supplemented
glutamine-starved cells, global protein synthesis and GS levels are
higher than in cells cultured in the absence of glutamine
Since asparagine is mainly used for protein
synthesis in mammalian cells (39),
it was analyzed whether asparagine could have a positive effect on
global protein synthesis in cells starved for glutamine, which
could in turn positively affect cell survival and proliferation.
Evaluating the incorporation of puromycin, a tyrosyl-tRNA mimetic
in cen3tel cells incubated in different culture media (Fig. 5A), it was found that a 24 h glutamine
starvation period decreased protein synthesis to ~50% compared with
control cells, while asparagine in the absence of glutamine allowed
a higher level of protein synthesis; however, this was still lower
than in control cells (60–65% of control cells). Similar levels of
protein synthesis were detected both 24 h after incubation of
glutamine-starved cells in the presence of asparagine and following
propagation of the starved cells in asparagine-containing medium
for three passages. These results were consistent with the
observation that glutamine deprivation led to a modest increase in
eIF2α phosphorylation, compared with control cells, which was not
detected following asparagine supplementation (Fig. 5B). The higher level of protein
synthesis in glutamine-deprived cells fed with asparagine, compared
with those starved for glutamine, was also observed in MDA-MB-231
cells (Fig. S1B, lanes 1, 3, 5 and
7). Thus, asparagine likely allows cells to maintain a level of
protein translation in the absence of glutamine that is compatible
with cell survival and proliferation.
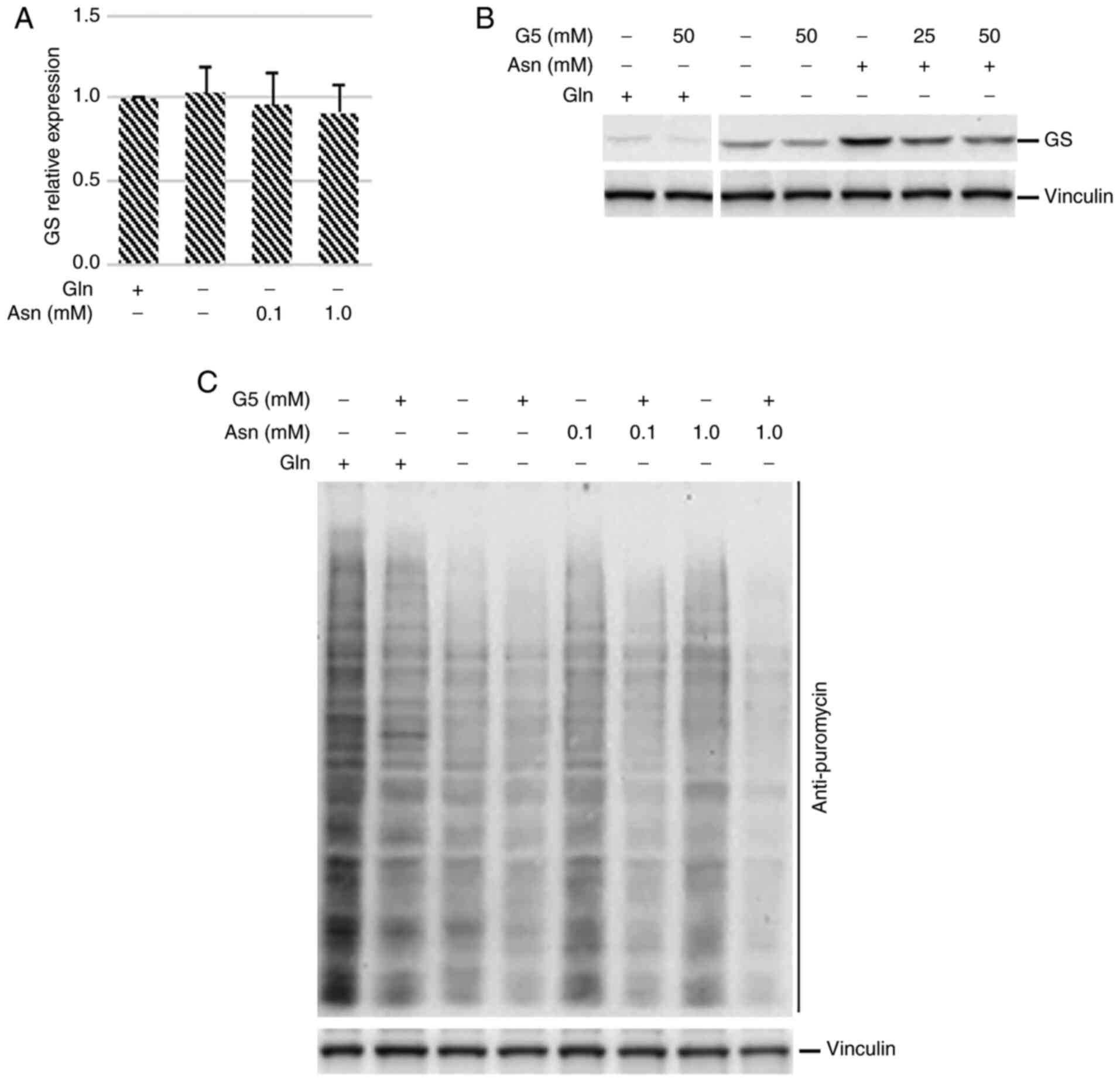
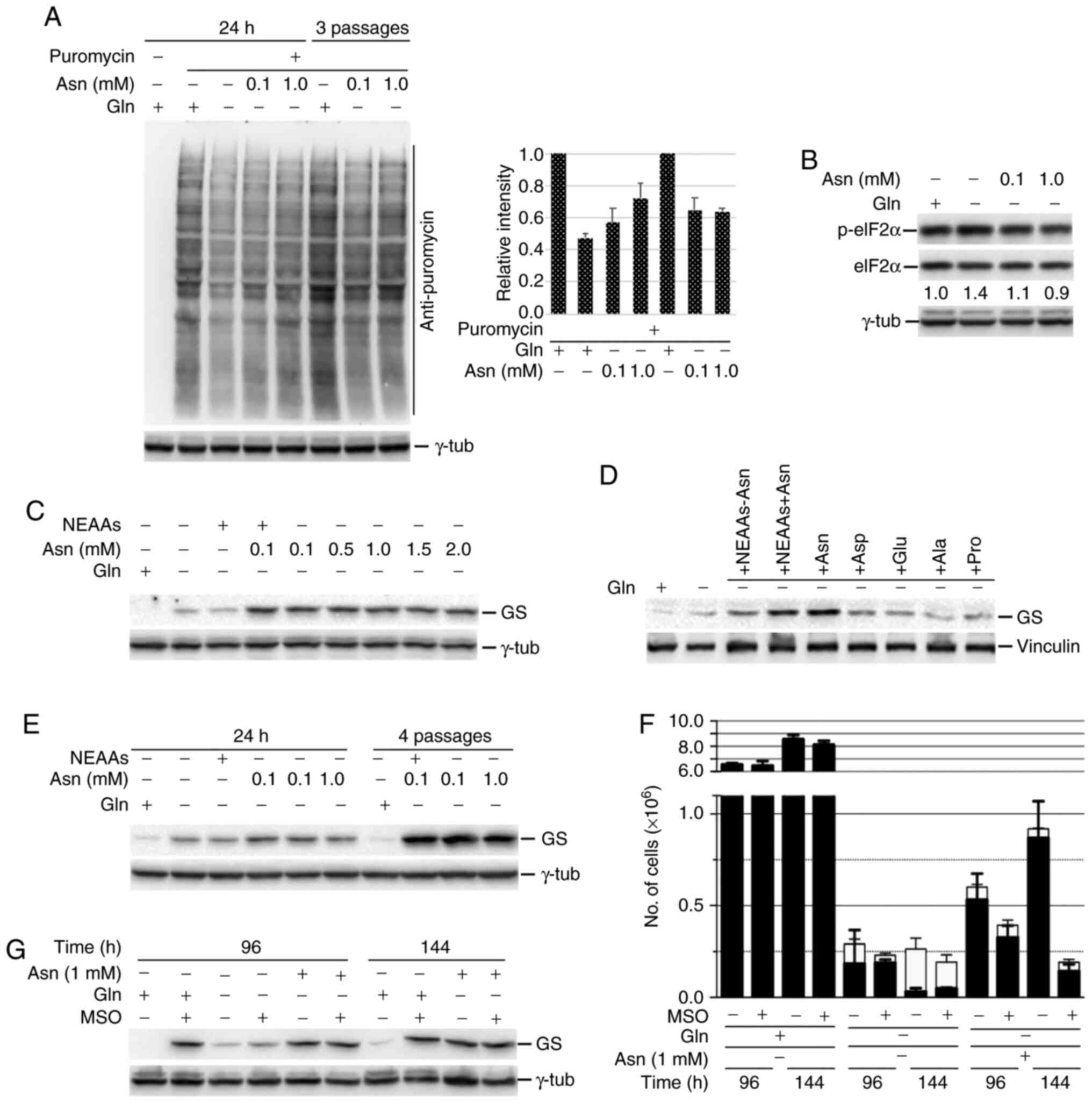 | Figure 5.Asparagine allows high levels of
global protein synthesis and GS expression in glutamine-starved
cen3tel cells. NEAAs, a pool of alanine, aspartic acid, glutamic
acid and proline at a concentration of 0.1 mM each, with or without
0.1 mM asparagine. (A) Western blot analysis of global protein
synthesis in cen3tel cells cultured for either 24 h in the medium
indicated or for 3 passages in the presence of glutamine or
asparagine. Puromycin (90 µM) was added during the last 10 min
before sample collections. Puromycin incorporation was revealed
using an anti-puromycin antibody. The plot shows the intensity of
the signal in each sample relative to cells grown in the presence
of glutamine (average values from two independent experiments;
error bars, standard deviations). (B) Western blot analysis of
eIF2α phosphorylation in cen3tel cells cultured in the indicated
medium. (C-E) Western blot analysis of GS expression in cen3tel
cells. (C and D) Cells were incubated in the presence of different
NEAAs or pool of NEAAs for 24 h or (E) cultured either for 24 h in
the medium indicated or for 4 passages in the presence of glutamine
or asparagine. (F) Cen3tel cell growth following incubation in the
medium indicated for 96 or 144 h with or without the GS inhibitor
MSO (2 mM). Error bars, standard deviation. (G) Western blot
analysis of GS expression in the samples cultured with or without
MSO. In the western blots, γ-tubulin or vinculin were used as the
loading controls. GS, glutamine synthetase; Gln, glutamine; Asn,
asparagine; NEAAs, non-essential amino acids; MSO, methionine
sulfoxamine. |
Since glutamine can be synthesized in mammalian
cells by the enzyme GS, it was analyzed whether the higher
translational ability of glutamine-deprived asparagine-fed cen3tel
cells could have an effect on the expression of GS. High levels of
this enzyme could sustain glutamine synthesis, and hence cell
survival and proliferation, when asparagine is supplied to
glutamine-starved cells. As shown in Fig.
5C-E, GS was barely detectable in cen3tel cells cultured in the
presence of glutamine. This result was not surprising, since it is
known that glutamine itself can regulate GS levels, directing the
enzyme towards ubiquitination and proteasomal degradation (14). In glutamine-starved cells, GS levels
were slightly higher than in control cells (Fig. 5C and D), but GS expression was
markedly increased when glutamine-starved cells were incubated with
asparagine, regardless of asparagine concentration (Fig. 5C). Asparagine was sufficient and
necessary to increase GS expression; in fact, no increase in GS
levels was observed when glutamine-starved cells were incubated
with a pool of alanine, aspartic acid, glutamic acid and proline,
or with each single amino acid, while an increase was observed when
asparagine was added to the pool (Fig.
5D). Of note, the propagation of glutamine-starved cells in the
presence of asparagine boosted GS expression in a
concentration-independent manner (Fig.
5E). The same type of modulation of GS expression was observed
in MDA-MB-231 cells cultured in the absence of glutamine and in the
presence of asparagine (Fig. S1C,
lanes 1, 3, 5 and 7).
To determine whether GS was required for
glutamine-starved cell survival and proliferation in the presence
of asparagine, cen3tel cells were treated with the GS inhibitor MSO
(2 mM). Cells were seeded in the presence of the inhibitor and
collected either 96 or 144 h after plating. As shown in Fig. 5F, MSO did not have any effect on cells
grown in the presence of glutamine-containing medium, while
glutamine-starved cells died independently of the presence of the
inhibitor. Glutamine-starved asparagine-supplemented cells survived
and proliferated, as expected, but died in the presence of MSO,
indicating that both GS expression and activity were essential for
cell survival and growth in the absence of glutamine.
When GS levels were analyzed in cells exposed to MSO
(Fig. 5G), it was found that the
protein accumulated in cells cultured with glutamine. It can be
speculated that this occurred because the binding between MSO and
GS (40) prevented GS ubiquitination
and degradation by the proteasome in the presence of glutamine. In
glutamine-deprived cells treated with MSO, GS levels remained low,
probably due to the fact that the impairment of translation
occurring in these conditions did not allow high levels of GS
synthesis.
Possible connection between c-Myc,
global protein synthesis and GS expression in glutamine-starved
asparagine-fed cells
The results presented so far indicated that, in the
presence of asparagine, glutamine-starved cells survive,
proliferate, express almost undetectable levels of c-Myc 1, high
levels of canonical c-Myc and GS, with the activity of this enzyme
being required for their survival. Given that c-Myc can positively
regulate GS expression through promoter demethylation
(13), GS transcripts were
analyzed by RT-qPCR in cen3tel cells incubated with different
nutrients. As shown in Fig. 6A, the
same levels of GS RNA were found in cells cultured either in
the presence or absence of glutamine, or in the absence of
glutamine but presence of asparagine. However, when cells were
exposed to 10074-G5, a compound that blocks c-Myc-Max interaction,
and hence c-Myc transcriptional activity (41), an evident decrease in GS protein
expression levels was observed in glutamine-starved cells
supplemented with asparagine, both in cen3tel and MDA-MB-231 cells
(Figs. 6B and S1C). This suggested that c-Myc could
control GS expression indirectly, at levels other than
transcription.
Since c-Myc is involved in protein synthesis
regulation at multiple levels, it was investigated whether c-Myc
inhibition with 10074-G5 had an effect on global protein
translation, which could account for the reduction in GS
expression. Indeed, 10074-G5 treatment reduced global protein
synthesis both in cen3tel and MDA-MB-231 cells incubated in
different culture conditions (Figs.
6C and S1B). This suggested that
in glutamine-starved cells fed with asparagine, c-Myc could
contribute to the maintenance of translation, which may in turn
allow the expression of GS, and hence glutamine synthesis.
Discussion
Oncogene activation and nutrient availability are
crucial factors for cancer cell hyperproliferation. c-Myc is a key
player in the metabolism of glucose and glutamine, which are avidly
used by cancer cells. Glutamine is an essential amino acid for
several tumors, particularly those bearing constitutively active
c-Myc (12). In fact,
oncogenic c-Myc causes cancer cells to become addicted to
exogenous glutamine and glutamine withdrawal leads to cell growth
arrest and cell death (6,42,43).
The expression of c-Myc in cancer cells can
be influenced by nutrient levels. Herein, glucose deprivation was
shown to lead to c-Myc overexpression in cen3tel cells,
similar to the results obtained from various cancer cell lines in
previous studies (16,17). In different cancer cell types,
glutamine deprivation was found to be associated with either
c-Myc upregulation or downregulation (16,20). In
the present study, glutamine starvation caused an alteration in
c-Myc protein expression, which, to the best of our knowledge, has
not yet been described in these culture conditions. In cen3tel
cells and various cancer cell lines cultured without glutamine, a
decrease in the expression of the canonical c-Myc isoform was
observed, together with the synthesis of the longer c-Myc isoform,
c-Myc 1. In cen3tel cells, the decline in canonical c-Myc and
synthesis of c-Myc 1 paralleled cell growth arrest. Furthermore, it
was demonstrated that asparagine was the only NEAA that, in the
absence of glutamine, rescued c-Myc expression and allowed cell
survival and proliferation, suggesting a possible link between
glutamine deprivation, aberrant c-Myc expression and cell growth
arrest.
In glutamine-starved cells, c-Myc 1 was already
detectable 6 h after deprivation and was expressed in a ~1:1 ratio
with the canonical c-Myc isoform after 24 h. This change in c-Myc
expression was specific to glutamine deprivation; in fact, it did
not occur in the absence of glucose only, but appeared shortly
after deprivation of both glucose and glutamine.
The functional role of the two c-Myc isoforms is
debated (21–23). While Blackwood et al (22) did not find variations in the
transcriptional activity of c-Myc 1 and canonical c-Myc, Hann et
al (21) showed that the
overexpression of c-Myc 1, but not that of canonical c-Myc,
significantly inhibited cell growth. In line with these results,
Sato et al (23) demonstrated
that the overexpression of the canonical c-Myc isoform in cancer
cells promoted cell proliferation and colony formation at much
higher levels than the longer isoform, and gene ontology analysis
of the genes upregulated by canonical c-Myc revealed an enrichment
in oncogenic and cell-cycle-related pathways. In the present study,
the decreased expression of canonical c-Myc and the presence of the
longer isoform were found to be associated with an arrest in cell
proliferation, supporting the hypothesis that c-Myc 1 cannot take
over the functions of canonical c-Myc, since the total level of the
c-Myc proteins was almost the same in control and glutamine-starved
cells. Indeed, it was found that asparagine was the only NEAA that
used as supplement for glutamine-starved cells allowed a high
expression of canonical c-Myc with a net decrease in c-Myc 1 levels
and, simultaneously, cell survival and proliferation. The
administration of the NEAAs alanine, aspartic acid, glutamic acid
or proline in glutamine-starved cells did not alter c-Myc
expression compared with glutamine deprivation, and did not allow
cellular proliferation, thus strengthening the idea that high
levels of canonical c-Myc are important for cellular viability and
growth. Asparagine was also able to rescue c-Myc expression in
cells that had been previously starved for glutamine.
In cells deprived of glutamine for 24 h, the
addition of the proteasomal inhibitor MG132 did not lead to the
accumulation of any c-Myc isoform, indicating that, at that stage
of starvation, both c-Myc synthesis and c-Myc degradation were
impaired. When asparagine was given to glutamine-starved cells,
c-Myc turnover appeared to occur as in the presence of glutamine,
confirming the importance of asparagine for c-Myc homeostasis.
Asparagine is synthesized by the enzyme asparagine
synthetase from aspartate and glutamine, which provides the
ammonium group (7). Thus, in the
absence of glutamine, asparagine cannot be synthesized and cells
must rely on exogenous asparagine to survive. When glutamine is
absent, asparagine behaves as an essential amino acid (27,28), with
the present results supporting this finding. Asparagine is not
catabolized in mammalian cells and is mainly used in protein
synthesis (39). Indeed, it was found
here that asparagine supplementation allowed an increase in global
translation, compared to glutamine-deprived cells and, in
particular, an increase in the levels of glutamine synthetase (GS),
the enzyme deputed to glutamine synthesis.
GS plays a fundamental role in cellular physiology;
in particular, when extracellular glutamine is scarce, it allows
the biosynthesis of endogenous glutamine, which in turn provides
the atoms required for the biosynthesis of multiple different
macromolecules. Indeed, GS has been found to be upregulated
in several cancers (13,44,45). As
also shown by Pavlova et al (28), this study confirmed that GS activity
is required for cellular viability in glutamine-deprived
asparagine-fed cells. In fact, its inhibition led to cell death in
these culture conditions. In glutamine-fed cells, MSO did not have
any effect on cellular viability, as expected. Asparagine was the
only NEAA capable of inducing an increase in GS levels in the
absence of glutamine. This increase was associated with GS ability
to sustain cellular viability and proliferation, confirming that
the high levels of this enzyme are important for cell survival when
exogenous glutamine is not available. The observation that MSO led
to the accumulation of GS in glutamine-fed cells, but not in
glutamine-deprived cells, reinforces the hypothesis that the
impairment in protein translation observed in the absence of
glutamine also prevents GS synthesis, which can instead occur
following asparagine supplementation.
However, cells maintained without glutamine but in
the presence of asparagine proliferated at a lower rate than cells
maintained in the presence of glutamine, suggesting that the high
levels of GS observed in these conditions did not provide
sufficient glutamine to support all the biosynthetic pathways in
which this NEAA is involved. Indeed, asparagine was not able to
relieve the stressful conditions induced by glutamine deprivation,
as indicated by the high levels of PARP1 auto-PARylation and
protein-PARylation, which were still present when glutamine-starved
cells were fed with asparagine. The observation that GSH
administration in asparagine-fed cells rescued cell proliferation
and PARP1 activation suggested that asparagine cannot support the
synthesis of adequate levels of GSH in the absence of glutamine,
and thus cannot prevent high levels of oxidative stress, which
could impair cell growth.
The present results revealed two main effects of
asparagine in cells deprived of glutamine: Asparagine allows a high
level of canonical c-Myc synthesis, as well as cell survival and
proliferation boosting GS levels. By inhibiting c-Myc activity
using 10074-G5, which prevents c-Myc/Max interaction (41), a decrease in GS levels was shown in
glutamine-starved asparagine-fed cells, suggesting that c-Myc can
play a role in regulating GS levels. However, this was not a
transcriptional regulation, as no differences in GS
transcript levels were observed between cells fed with glutamine,
without glutamine or with asparagine in the absence of glutamine.
Given that c-Myc is also known to indirectly control translation at
various levels, including the expression of translational factors
and ribosomal proteins (46,47), and indeed c-Myc inhibition by 10074-G5
was found to lead to a decrease in global protein synthesis, it can
be envisaged that the high levels of the canonical c-Myc isoform
found in glutamine-starved cells cultured in the presence of
asparagine could sustain translation and GS synthesis in
particular, which leads to the biosynthesis of glutamine, which can
in turn sustain cellular viability.
In conclusion, these results indicate that glutamine
deprivation can have an effect on c-Myc translation. Even though
further experiments will be required to better elucidate the roles
of c-Myc and c-Myc 1 in the response to nutrient starvation, the
observed correlation between the decrease in the expression of
canonical c-Myc, the increase in c-Myc 1 levels and growth arrest
in glutamine-deprived cells suggests that these phenomena can be
interconnected. The finding that, in the absence of glutamine,
asparagine allows both a high expression of canonical c-Myc and
cellular proliferation strengthens this hypothesis. Future
experiments testing these observations in vivo could provide
relevant information for the development of cancer therapies
targeting metabolism.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The authors did not receive support from any
organization for the submitted work.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors contributions
IC, CP and DB performed the experiments and analyzed
the data. IC and CM conceived the research concept and designed the
experiments and wrote the manuscript. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors have no relevant financial or
non-financial interests to disclose.
References
|
1
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: Metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhu J and Thompson CB: Metabolic
regulation of cell growth and proliferation. Nat Rev Mol Cell Biol.
20:436–450. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pavlova NN and Thompson CB: The emerging
hallmarks of cancer metabolism. Cell Metab. 23:27–47. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ganapathy-Kanniappan S: Molecular
intricacies of aerobic glycolysis in cancer: Current insights into
the classic metabolic phenotype. Crit Rev Biochem Mol Biol.
53:667–682. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wise DR, DeBerardinis RJ, Mancuso A, Sayed
N, Zhang XY, Pfeiffer HK, Nissim I, Daikhin E, Yudkoff M, McMahon
SB and Thompson CB: Myc regulates a transcriptional program that
stimulates mitochondrial glutaminolysis and leads to glutamine
addiction. Proc Natl Acad Sci USA. 105:18782–18787. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Altman BJ, Stine ZE and Dang CV: From
Krebs to clinic: Glutamine metabolism to cancer therapy. Nat Rev
Cancer. 16:619–634. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang J, Pavlova NN and Thompson CB:
Cancer cell metabolism: The essential role of the nonessential
amino acid, glutamine. EMBO J. 36:1302–1315. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pinweha P, Rattanapornsompong K,
Charoensawan V and Jitrapakdee S: MicroRNAs and oncogenic
transcriptional regulatory networks controlling metabolic
reprogramming in cancers. Comput Struct Biotechnol J. 14:223–233.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tarrado-Castellarnau M, de Atauri P and
Cascante M: Oncogenic regulation of tumor metabolic reprogramming.
Oncotarget. 7:62726–62753. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dejure FR and Eilers M: MYC and tumor
metabolism: Chicken and egg. EMBO J. 36:3409–3420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dang CV: A Time for MYC: Metabolism and
therapy. Cold Spring Harb Symp Quant Biol. 81:79–83. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bott AJ, Peng IC, Fan Y, Faubert B, Zhao
L, Li J, Neidler S, Sun Y, Jaber N, Krokowski D, et al: Oncogenic
Myc induces expression of glutamine synthetase through promoter
demethylation. Cell Metab. 22:1068–1077. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arad G, Freikopf A and Kulka RG:
Glutamine-stimulated modification and degradation of glutamine
synthetase in hepatoma tissue culture cells. Cell. 8:95–101. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hann SR, Sloan-Brown K and Spotts GD:
Translational activation of the non-AUG-initiated c-myc 1 protein
at high cell densities due to methionine deprivation. Genes Dev.
6:1229–1240. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun L, Song L, Wan Q, Wu G, Li X, Wang Y,
Wang J, Liu Z, Zhong X, He X, et al: cMyc-mediated activation of
serine biosynthesis pathway is critical for cancer progression
under nutrient deprivation conditions. Cell Res. 25:429–444. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu S, Yin X, Fang X, Zheng J, Li L, Liu X
and Chu L: c-MYC responds to glucose deprivation in a
cell-type-dependent manner. Cell Death Discov. 1:150572015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wong WJ, Qiu B, Nakazawa MS, Qing G and
Simon MC: MYC degradation under low O2 tension promotes survival by
evading hypoxia-induced cell death. Mol Cell Biol. 33:3494–3504.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Effenberger M, Bommert KS, Kunz V, Kruk J,
Leich E, Rudelius M, Bargou R and Bommert K: Glutaminase inhibition
in multiple myeloma induces apoptosis via MYC degradation.
Oncotarget. 8:85858–85867. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dejure FR, Royla N, Herold S, Kalb J, Walz
S, Ade CP, Mastrobuoni G, Vanselow JT, Schlosser A, Wolf E, et al:
The MYC mRNA 3′-UTR couples RNA polymerase II function to glutamine
and ribonucleotide levels. EMBO J. 36:1854–1868. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hann SR, Dixit M, Sears RC and Sealy L:
The alternatively initiated c-Myc proteins differentially regulate
transcription through a noncanonical DNA-binding site. Genes Dev.
8:2441–2452. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Blackwood EM, Lugo TG, Kretzner L, King
MW, Street AJ, Witte ON and Eisenman RN: Functional analysis of the
AUG- and CUG-initiated forms of the c-Myc protein. Mol Biol Cell.
5:597–609. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sato K, Masuda T, Hu Q, Tobo T, Gillaspie
S, Niida A, Thornton M, Kuroda Y, Eguchi H, Nakagawa T, et al:
Novel oncogene 5MP1 reprograms c-Myc translation initiation to
drive malignant phenotypes in colorectal cancer. EBioMedicine.
44:387–402. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hay N: Reprogramming glucose metabolism in
cancer: Can it be exploited for cancer therapy? Nat Rev Cancer.
16:635–649. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fung MKL and Chan GC: Drug-induced amino
acid deprivation as strategy for cancer therapy. J Hematol Oncol.
10:1442017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matés JM, Di Paola FJ, Campos-Sandoval JA,
Mazurek S and Márquez J: Therapeutic targeting of glutaminolysis as
an essential strategy to combat cancer. Semin Cell Dev Biol.
98:34–43. 2020. View Article : Google Scholar
|
|
27
|
Zhang J, Fan J, Venneti S, Cross JR,
Takagi T, Bhinder B, Djaballah H, Kanai M, Cheng EH, Judkins AR, et
al: Asparagine plays a critical role in regulating cellular
adaptation to glutamine depletion. Mol Cell. 56:205–218. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pavlova NN, Hui S, Ghergurovich JM, Fan J,
Intlekofer AM, White RM, Rabinowitz JD, Thompson CB and Zhang J: As
extracellular glutamine levels decline, asparagine becomes an
essential amino acid. Cell Metab. 27:428–438.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mondello C, Chiesa M, Rebuzzini P, Zongaro
S, Verri A, Colombo T, Giulotto E, Dincalci M, Franceschi C and
Nuzzo F: Karyotype instability and anchorage-independent growth in
telomerase-immortalized fibroblasts from two centenarian
individuals. Biochem Biophys Res Commun. 308:914–921. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zongaro S, de Stanchina E, Colombo T,
DIncalci M, Giulotto E and Mondello C: Stepwise neoplastic
transformation of a telomerase immortalized fibroblast cell line.
Cancer Res. 65:11411–11418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Belgiovine C, Frapolli R, Bonezzi K,
Chiodi I, Favero F, Mello-Grand M, Dei Tos AP, Giulotto E,
Taraboletti G, DIncalci M and Mondello C: Reduced expression of the
ROCK Inhibitor Rnd3 is associated with increased invasiveness and
metastatic potential in mesenchymal tumor cells. PLoS One.
5:e141542010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ostano P, Bione S, Belgiovine C, Chiodi I,
Ghimenti C, Scovassi AI, Chiorino G and Mondello C: Cross-analysis
of gene and miRNA genome-wide expression profiles in human
fibroblasts at different stages of transformation. OMICS. 16:24–36.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bono B, Ostano P, Peritore M, Gregnanin I,
Belgiovine C, Liguori M, Allavena P, Chiorino G, Chiodi I and
Mondello C: Cells with stemness features are generated from in
vitro transformed human fibroblasts. Sci Rep. 8:138382018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chiodi I, Picco G, Martino C and Mondello
C: Cellular response to glutamine and/or glucose deprivation in
in vitro transformed human fibroblasts. Oncol Rep.
41:3555–3564. 2019.PubMed/NCBI
|
|
35
|
Belgiovine C, Chiesa G, Chiodi I, Frapolli
R, Bonezzi K, Taraboletti G, DIncalci M and Mondello C: Snail
levels control the migration mechanism of mesenchymal tumor cells.
Oncol Lett. 12:767–771. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu L, Begum S, Hearn JD and Hynes RO:
GPR56, an atypical G protein-coupled receptor, binds tissue
transglutaminase, TG2, and inhibits melanoma tumor growth and
metastasis. Proc Natl Acad Sci USA. 103:9023–9028. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Stegink LD, Filer LJ Jr, Brummel MC, Baker
GL, Krause WL, Bell EF and Ziegler EE: Plasma amino acid
concentrations and amino acid ratios in normal adults and adults
heterozygous for phenylketonuria ingesting a hamburger and milk
shake meal. Am J Clin Nutr. 53:670–675. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ubuka T and Meister A: Studies on the
utilization of asparagine by mouse leukemia cells. J Natl Cancer
Inst. 46:291–298. 1971.PubMed/NCBI
|
|
40
|
Freikopf A and Kulka RG: Specificity of
the glutamine-binding site involved in the regulation of
glutamine-synthetase activity in hepatoma tissue-culture cells. Eur
J Biochem. 56:483–492. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yap JL, Wang H, Hu A, Chauhan J, Jung KY,
Gharavi RB, Prochownik EV and Fletcher S: Pharmacophore
identification of c-Myc inhibitor 10074-G5. Bioorg Med Chem Lett.
23:370–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yuneva M, Zamboni N, Oefner P,
Sachidanandam R and Lazebnik Y: Deficiency in glutamine but not
glucose induces MYC-dependent apoptosis in human cells. J Cell
Biol. 178:93–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qing G, Li B, Vu A, Skuli N, Walton ZE,
Liu X, Mayes PA, Wise DR, Thompson CB, Maris JM, et al: ATF4
regulates MYC-mediated neuroblastoma cell death upon glutamine
deprivation. Cancer Cell. 22:631–644. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tardito S, Oudin A, Ahmed SU, Fack F,
Keunen O, Zheng L, Miletic H, Sakariassen PØ, Weinstock A, Wagner
A, et al: Glutamine synthetase activity fuels nucleotide
biosynthesis and supports growth of glutamine-restricted
glioblastoma. Nat Cell Biol. 17:1556–1568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bott AJ, Shen J, Tonelli C, Zhan L,
Sivaram N, Jiang YP, Yu X, Bhatt V, Chiles E, Zhong H, et al:
Glutamine anabolism plays a critical role in pancreatic cancer by
coupling carbon and nitrogen metabolism. Cell Rep. 29:1287–1298.e6.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Biffo S, Manfrini N and Ricciardi S:
Crosstalks between translation and metabolism in cancer. Curr Opin
Genet Dev. 48:75–81. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Singh K, Lin J, Zhong Y, Burčul A, Mohan
P, Jiang M, Sun L, Yong-Gonzalez V, Viale A, Cross JR, et al: c-MYC
regulates mRNA translation efficiency and start-site selection in
lymphoma. J Exp Med. 216:1509–1524. 2019. View Article : Google Scholar : PubMed/NCBI
|