Introduction
Tobacco smoke, chronic alcohol use and infection
with human papillomavirus type 16 (HPV-16) are known risk factors
for laryngopharyngeal cancer (1–4); however,
there is a growing interest in identifying other risk factors.
Laryngopharyngeal reflux (LPR), a variant of
gastro-esophageal reflux disease (GERD), has also been considered a
potential risk factor in the last decade that may exert a
carcinogenic effect on the upper aerodigestive tract (5–11). Recent
findings have clarified the role of bile reflux as an independent
risk factor in hypopharyngeal carcinogenesis through preclinical
and clinical models (12–18), and a model of the molecular mechanism
of the bile-induced tumorigenic effect has been proposed (19–25). Bile
is produced in the liver and stored in the gallbladder (26) and its primary role is to assist in
lipid digestion and absorption. Regurgitation of duodenal bile
contents into the stomach and esophagus is known as bile reflux.
The present review provides the latest knowledge regarding the
association of bile reflux with hypopharyngeal cancer.
Bile as a potential carcinogen
The caustic nature of bile has long been recognized
(27). In ancient times, the medical
theory that excess, deficiency or ectopic bile in the body may
affect human health was first stated by the father of medicine,
Hippocrates (27). This medical
theory remained popular for centuries through the writings of Galen
(129–201 AD) (28) but was decisively
displaced newly published theories of cellular pathology by Virchow
and Rather in 1858 (29). In the past
century, in 1938-40, Cook (30)
proposed the possible role of bile acids in cancer. Years later
(1974–1993) several studies supported the role of bile acids as
carcinogens causing gastrointestinal cancer (31–34). In
parallel, clinical studies provided the first evidence of mixed
gastric and bile (duodenal) fluids in refluxate of patients with
GERD (35–37). During this period, Gotley et al
(38) determined an increased amount
of conjugated bile acids in 87% of aspirates using high-performance
liquid chromatography, suggesting an association between bile and
acid concentration and esophageal mucosal injury. Nehra et
al (35), Kauer et al
(39,40) and Domellof et al (41) also characterized the concentration and
composition of bile fluid in aspirates of patients with GERD. Of
note, Fein et al (42) were
the first who provided solid evidence of bile fluid as an
independent carcinogen in the gastrointestinal tract by using a rat
model and demonstrating that bile acids are able to induce
esophageal adenocarcinoma.
Until the present day, the clinical prevalence and
magnitude of bile reflux have remained to be fully determined;
however, there is growing evidence of bile contents in GERD
refluxate (31,43–45).
According to Covington et al (46), bile-containing enterogastric reflux is
much more common than previously assumed. The increased information
linking LPR and inflammatory/neoplastic disease of the upper
aerodigestive tract as well as the lack of evidence for the
carcinogenic effect of bile-containing refluxate into
laryngopharynx led to further investigation.
Considerable research efforts to explore the
carcinogenic effect of bile in the upper aerodigestive tract were
made, including clinical and experimental studies. Galli et
al (5) and Sasaki et al
(11) suggested that during LPR, bile
fluid reaches the epithelium of the upper aerodigestive tract,
which may contribute to the development of inflammatory and
neoplastic events. Furthermore, Lewin et al (47) reported a close association between LPR
and patients with premalignant lesions or early carcinomas of the
larynx. It is worth mentioning that although there have been
efforts to link the effect of other gastroesophageal refluxate
contents, such as pepsin, with pre-neoplastic events in the larynx
and pharynx, the conclusions have been divergent (48–54). A
recent study indicated a possible contributory effect of slightly
acidic or neutral pepsin to the inflammatory and neoplastic effects
of LPR (54), but there is still no
direct evidence of carcinogenesis induced by pepsin.
At the beginning of 2016, a series of in
vitro and in vivo experiments questioned the role of
bile reflux in hypopharyngeal carcinogenesis (12–14). A
study by our group from 2016 (13)
established a murine model of wild-type mice, Mus musculus
(C57BL/6J) and provided the first evidence that bile acids may
cause preneoplastic lesions in the hypopharyngeal mucosa (HM).
Using this model and long-term exposure to bile acids, the
progressive mutagenic effect of biliary refluxate causing invasive
cancer was subsequently observed (15,16). These
and other studies provided direct evidence that bile fluid is a
carcinogen, capable of inducing hypopharyngeal squamous cell
carcinoma (HSCC) (15,16). The significance of these findings was
in line with clinical findings derived from bile reflux-related
HSCC (18). Specifically, a clinical
pilot study demonstrated a characteristic bile-related molecular
phenotype that was similarly identified in bile-exposed murine HM,
which clearly differed from adjacent non-pathologic tissue
(18).
Bile refluxate composition and acidity as
critical factors of neoplastic events
Bile refluxate composition
According to Nehra et al (35) and Kauer et al (39,40), the
majority of bile acids in esophageal aspirates of patients with
GERD are conjugated with taurine or glycine and may be sulfated.
Specifically, glycine conjugates of cholic, deoxycholic and
chenodeoxycholic acids are the predominant bile acids aspirated
from the esophagus of patients with GERD (the ratio of glycine to
taurine conjugates in normal human bile is 3:1). It has also been
indicated that unconjugated secondary bile acids, such as
deoxycholic acid (DCA), may be present in the esophageal refluxate,
particularly in patients with erosive esophagitis and Barrett's
esophagus (35,41,55). It is
known that pH affects the solubility of each bile component in
various manners. At acidic pH (≤4), the conjugated bile acids are
more un-ionized and therefore capable of penetrating or interacting
with the cell membrane (taurine conjugates: pKa=1.8–1.9; glycine
conjugates: pKa=4.3–5.2) (56). At pH
<3.0, bile salts tend to precipitate, whereas between pH 5.5 and
7.0, most conjugated primary bile acids are found to be ionized and
therefore relatively inactive. However, unconjugated secondary bile
acid DCA remains unionized (pKa 5.5–6.2) and may therefore exert
its harmful effects, causing mucosal injury even at a less acidic
pH (12,16,17,55).
According to Stamp (57), duodenal
fluid, at a less acidic pH, may also contribute to gastrointestinal
tract tumorigenesis. Specifically, glycine conjugates may remain
un-charged and therefore be harmful to the epithelial cells at a
less acidic pH. Ireland et al (58) indicated that duodenal fluid
significantly contributes to the carcinogenic potential of
methyl-N-amyl nitrosamine, particularly at less acidic pH. In
addition, as bile acids are natural detergents when in high
concentration, they may interact with the cell membrane even at a
neutral pH.
Bile refluxate concentration
Although the bile composition is a crucial factor of
bile reflux-related tumorigenesis, the concentration of bile may
also potentiate its oncogenic effect. This view may be supported by
previous in vivo studies indicating that the application of
bile reflux components, chenodeoxycholic acid or DCA, at
pharmacologic concentrations, both at a neutral pH of 7.0, to
murine laryngopharyngeal mucosa was able to cause early
premalignant changes, such as hyperplasia and dysplasia, as well as
marked activation of NF-κB and its related oncogenic molecular
phenotype (13,14).
Bile refluxate acidity
It appears that acidity of LPR refluxate is a
critical factor for bile to induce a harmful effect on
laryngopharyngeal mucosa (39,59,60).
In the clinic, intraesophageal pH monitoring has been used
extensively to identify reflux episodes. Although pH varies during
gastroesophageal reflux episodes, according to Ulualp et al
(61), 24-h ambulatory pH monitoring
in the pharynx of patients with reflux laryngitis confirmed that a
drop below pH 4.0 is common and is considered diagnostic of a
reflux event, suggesting that acid may contribute to
duodenogastric-induced inflammatory and neoplastic events. Lillemoe
et al (62) demonstrated the
injurious effect of the various duodenal components on rabbit
esophageal mucosa at strongly acidic pH, supporting a synergism
between bile and HCl. A study by our group from 2019 (17) documented that the tumorigenic effect
of bile on hypopharyngeal cells is pH-dependent. Other studies from
our group (13–15) also demonstrated that a strongly acidic
pH (≤4.0) serves a critical role in the bile-induced tumorigenic
effect in murine laryngopharyngeal mucosa. It was indicated that
chronic intermittent exposure of murine HM to a mixture of bile
salts at a strongly acidic pH of 3.0 was able to progressively
induce premalignant changes, microinvasion and invasive squamous
cell carcinoma, causing increased DNA damage and oncogenic
molecular alterations (15).
Specifically, it was demonstrated that histopathologic changes
caused by acidic bile were accompanied by underlying molecular
alterations, such as increased levels of i) oxidative DNA/RNA
damage and double-strand break (DSB) markers, ii) p53 and cell
proliferation markers (Ki67, cytokeratin 14, and p63), as well as
iii) alterations of the expression of cell adhesion molecules, like
E-Cadherin and β-catenin, and iv) activation of NF-κB and other
cancer-related transcription factors, such as signal transducer and
activator of transcription 3 (STAT3) (13–15).
However, hypopharyngeal cells or mucosa exposed to the same mixture
of conjugated primary bile acids at neutral pH (7.0) produced
hyperplastic or mild dysplastic changes and significantly less
intense underlying molecular changes compared to acidic bile salts
(12,13,17). In
parallel, it was indicated that chronic exposure to acid alone or
concentrated glucose was not able to produce any histological
changes (13,15). The negative or reduced effect of acid
alone and/or bile salts at neutral pH, compared to acidic bile
salts, indicates that the latter is particularly injurious.
Primary and secondary bile acids in
refluxate
It is clear that the presence of conjugated primary
bile acids in a highly acidic refluxate exerts a tumorigenic
potency on the long-term exposed upper aerodigestive tract. This
theory may explain findings from our group (13,15)
indicating that chronic local exposure of murine HM to a mixture of
conjugated primary bile acids, at concentrations previously
measured in patients with GERD (35,40,43,35)
at a strongly acidic pH (≤4.0), is able to progressively cause
precancerous lesions and invasive cancer. Specifically, as taurine
conjugates are active at low pH (≤4.0), it appears that
taurine-conjugated bile acids may be responsible for the
tumorigenic effect of bile at lower pH (15,17). There
is also recent in vitro evidence supporting that acidity (pH
≤4.0) and bile composition may have a role in the progression of
HSCC (63).
The above observations strongly support that
controlling the pH during reflux episodes may have a protective
effect by reducing the risk of bile-induced hypopharyngeal cancer.
However, there is epidemiologic evidence that numerous patients
with refractory GERD may also experience symptoms at a weakly
acidic pH of 5.5–6.0 (60,64). Since unconjugated DCA and
glycine-conjugated bile acids may be partially active at a weakly
acidic pH, it appears that as the pH grows less acidic, approaching
pH 5.5, the partially activated primary bile acids and the
activated DCA may exert their influence (17). A recent study by our group (16) supported that DCA and
glycine-conjugated bile acids are potent activators of DNA damage
and oncogenic pathways in HM in a weakly acidic environment.
Previous findings have demonstrated a similar association between
DCA and its tumorigenic activity in the esophagus and colon
(31,65,66).
Regarding the hypopharynx, it has been documented that bile at a
weakly acidic pH (5.0–5.5) with or without DCA, similarly to a
strongly acidic pH 3.0, is able to increase the risk of
bile-related hypopharyngeal neoplasia by promoting premalignant
lesions, DNA damage and oncogenic molecular alterations, compared
to controls (16). Of note, it was
indicated that long-term exposure to a weakly acidic control (pH
5.5) was not able to induce any histological changes (16). This observation strongly supports that
the oncogenic properties of biliary esophageal reflux on
laryngopharyngeal mucosa may not be fully modified when antacid
therapy is applied.
Although further exploration with clinical evidence
is expected to strengthen these previous preclinical observations,
investigation of the mechanism by which bile refluxate exerts its
oncogenic properties is expected to contribute not only to a better
understanding of the pathophysiology of hypopharyngeal cancer but
also to alternative therapeutic strategies for patients with
refractory GERD, using specific inhibitors of relevant molecular
pathways or bile receptors.
Key role of NF-κB in bile reflux-related
hypopharyngeal carcinogenesis
Several epidemiologic studies have supported the
role of LPR in the neoplasia of the upper aerodigestive tract
(5–11,67,68).
However, the exact mechanism of LPR-related laryngopharyngeal
carcinogenesis has remained elusive and unexplored until the last
decade (69–71). Studies including that by Huo et
al (66) indicated that exposure
of esophageal cells to DCA produced elevated levels of NF-κB in
vitro, suggesting the role of NF-κB as a key molecule in
esophageal cancer (72–75). Thus, these observations supported the
hypothesis of a possible mechanistic role of NF-κB in cancer of
extraesophageal sites, such as hypopharynx.
NF-κB is a transcriptional factor complex consisting
of homo- and heterodimers of five members of the Rel family [RelA
(p65), RelB, c-Rel, NF-κB1 (p50/p105) and NF-κB2 (p52/p100)]
(76). The canonical pathway of NF-κB
activation includes phosphorylation of IκB-α, which leads to
nuclear translocation of the heterodimers p50/Rela or p50/cRel and
consequent binding to the promoters of target genes and regulation
of their expression. Constitutive activation of NF-κB has been
observed in various cancer types, linking inflammation to the
neoplastic transformation of the epithelium (77–79). In
the initiation and progression of head and neck squamous cell
carcinoma (HNSCC), several oncogenic pathways have been identified.
These commonly include epidermal growth factor receptor
(EGFR)/Ras/RAF/MAPK, PI3K/Akt1/mTOR, IΚΚ/NF-κB, STAT3 and
Wnt/β-catenin (80–89). It has been indicated that HNSCC
exhibits abundant NF-κB activation and several studies indicate
that NF-κB is upregulated in premalignant lesions and invasive
cancer (80,90–94).
The role of NF-κB in acidic bile reflux-related
laryngopharyngeal carcinogenesis was first demonstrated in the last
decade through in vitro and in vivo experimental
models (12–25). A study by our group from 2017
(21) documented the key role of
NF-κB in mediating acidic bile-induced oncogenic molecular events
in human hypopharyngeal cells (HHCs). Subsequently in 2018, a study
by our group (22) also demonstrated
that NF-κB is a crucial factor in controlling the levels of small
regulatory molecules, such as microRNA (miRNA/miR) markers, in
HHCs. A series of in vitro and in vivo studies also
suggested that NF-κB inhibition may prevent inflammatory and
neoplastic events in HHCs and HM, including STAT3 activation and
significant deregulations of several cancer-related genes and miRNA
markers (19,23–25). Other
non-specific stress factors, such as highly concentrated glucose or
acidic pepsin, were not capable of inducing activation of genes
with oncogenic function previously linked to HNSCC in vitro
(12,13,53).
Bile reflux-induced NF-κB-related mRNA
oncogenic phenotype
There is evidence that numerous types of cancer
arise from sites of chronic inflammation (95). Specifically, a wide array of chronic
inflammatory conditions predisposes susceptible cells of epithelial
origin to neoplastic transformation (carcinomas) as a multistep
process (focal proliferation of dysplastic cells with potential
progression to malignant carcinoma). An example includes reflux
esophagitis that may lead to DNA damage, development of Barrett's
esophagus and esophageal carcinoma (96). In certain cases, the progenitors of
the inflammation are known, such as bacterial infections or gastric
acids that have been associated with increased risk of
adenocarcinoma of the stomach and esophagus, respectively (97,98). It
may be assumed that bile acids interact directly with HM, as
described in a paragraph below, but may also be the progenitors of
an LPR-induced chronic inflammatory microenvironment associated
with an increased risk of hypopharyngeal cancer. It has been
indicated that a chronic inflammatory microenvironment or harmful
stimuli are able to induce a constitutive activation of NF-κB,
which may lead to a cascade of molecular alterations. Specifically,
constitutive activation of NF-κB may lead to subsequent
transcriptional activation of genes that are implicated in a
variety of signaling pathways via aberrant overexpression of
cytokines, transcription factors and growth factor receptors, such
as TNF-α, TLR and EGFR (76,77,79–89,93,94,99–103).
Preclinical studies from our group (12–25)
documented that acidic bile is able to induce activation of NF-κB
and significant overexpression of several cancer-related genes.
Specifically, acidic bile was reported to induce significant
transcriptional activation of anti-apoptotic BCL2 and other
genes previously linked to HNSCC, such as STAT3, EGFR, WNT5A,
TNF-α, ΔNp63, cREL, IL6, IL1β, AKT1 and PTGS2 (82–94,102–111)
(Fig. 1). Furthermore, a clinical
pilot study revealed that bile-related HSCC had significantly
higher levels of NF-κB and differential expression of the above
genes compared to the adjacent non-pathologic tissue or
bile-negative HSCC, providing further evidence of the central role
of NF-κB (18).
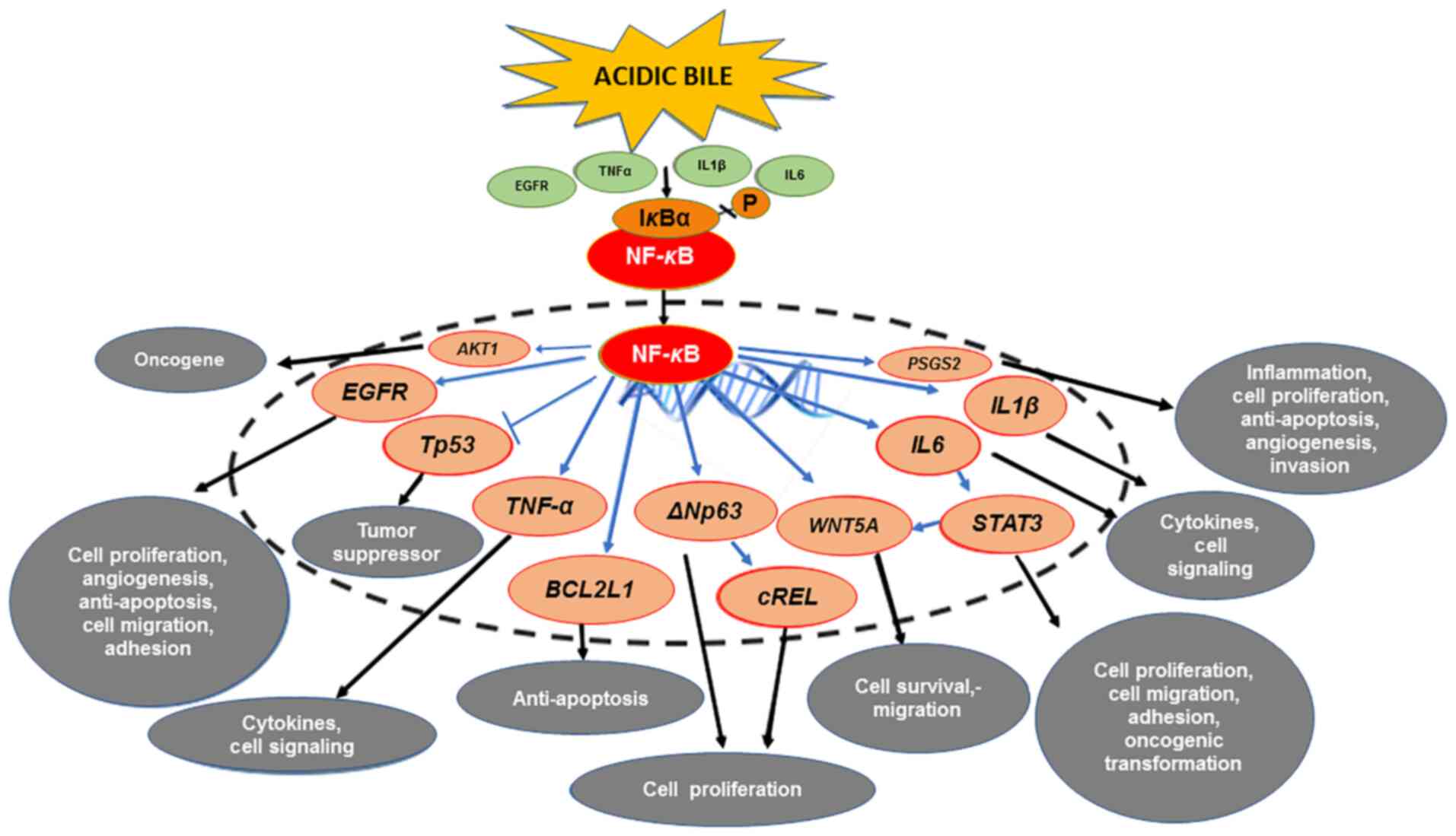 | Figure 1.The mechanistic role of NF-κB in the
mRNA oncogenic phenotype induced by bile reflux in hypopharyngeal
carcinogenesis. Acidic bile induces constitutive activation of
NF-κB via TNF-α, EGFR or TLR, which promotes the transcriptional
activation of genes with inflammatory, anti-apoptotic or oncogenic
function, such as IL6, IL1β, TNF-α, BCL2L1, EGFR, cREL, STAT3,
ΔNp63 and WNT5A. Activation of NF-κB under acidic bile
exposure also induces overexpression of AKT1, suggesting
acidic bile may contribute to the PI3K/AKT1 downstream pathway,
which is frequently activated in HNSCC. In addition, NF-κB is able
to upregulate the expression of PTGS2 (COX-2), supporting
its regulatory role in inflammatory and cancer-related downstream
signaling pathways. Finally, acidic bile-induced NF-κB activation
may prevent the upregulation of wild-type TP53
expression. |
Targeting NF-κB negatively affects the NF-κB
signaling pathway and has been indicated to be an encouraging
strategy to improve anticancer therapies (112). Several pharmacologic and dietary
inhibitors of NF-κB are considered promising thera-peutic options,
demonstrating chemo-preventive or chemo-sensitizing properties in
head and neck cancer (99,112). BAY 11-7082
[(E)-3-(4-methylphenylsulphonyl)-1-propenenitrile] is a reliable
inhibitor of the NF-κB pathway that has been widely used in
numerous studies exploring the effect of NF-κB (112,113).
It has been suggested that BAY 11-7082 offers the most rapid and
potent anti-tumor effect among other NF-κB inhibitors (112) and it may be used as a sensitizer for
anti-cancer treatment (114,115). Furthermore, curcumin is a turmeric
natural supplement with known antioxidant, anti-inflammatory and
anti-cancer properties, previously demonstrated to have potential
chemo-preventive effects in head and neck malignancies (116), by blocking NF-κB activation and
halting the proliferation of cancer cells (117) due to its pleiotropic properties
(118).
Both BAY 11-7082 and dietary curcumin have been used
in in vitro and in vivo experimental studies to
investigate the underlying mechanism of bile reflux-induced
carcinogenic effect into the hypopharynx. Studies by our group
(19,25) suggested that application of BAY
11-7082 effectively suppressed cell proliferation rates, the
activation of NF-κB and related oncogenic mRNA profiles induced by
acidic bile exposure. This oncogenic phenotype included the
significant overexpression of anti-apoptotic BCL2 and other
genes implicated in the initiation and progression of HNSCC,
including TNF-α, EGFR, STAT3, ΔNp63, cREL, IL6, IL1β, AKT1,
PTGS2 and WNT5A (82–85,87,89–94,102–111)
(Fig. 1). Parallel evidence that
acidic bile stimulus is able to activate NF-κB and its related
pathways in HHCs arose from the in vitro treatment with
curcumin, which successfully blocked the transcriptional activity
of NF-κB (20), similar to BAY
11-7082 (21,23). A study by our group from 2020
(24) documented the preventive and
therapeutic properties of curcumin on murine HM against the acidic
bile effect, thus shaping the future translational development of
effective targeted therapies using topical non-pharmacologic
inhibitors of NF-κB.
Strong evidence that NF-κB activation is able to
influence the acidic bile-induced oncogenic mRNA profile inspired a
further study on whether synchronizing its inhibition with acidic
bile exposure is significant. Thus, a study by our group from 2019
(23) reported the temporal
characteristics of NF-κB inhibition in blocking the acidic
bile-induced oncogenic molecular events in HHCs. A series of
studies also documented that topical application of BAY 11-7082 or
curcumin to HM, either before, after or simultaneous to acidic bile
exposure, successfully prevented or suppressed cell proliferation
and NF-κB-related molecular events (19,24,25).
These results revealed that the upregulation of
RELA, BCL2, STAT3, EGFR, WNT5A, TNF-α, IL6 and PTGS2
is directly promoted by acidic bile through NF-κB, shortly after
its exposure (19,24,25)
(Fig. 1), and strongly suggested that
it may be clinically feasible to topically apply NF-κB inhibitors,
without any precise synchronization with acidic bile exposure, to
prevent acidic bile-induced oncogenic molecular changes.
Interactions between NF-κB activation
and other factors
The application of NF-κB inhibitors also revealed
important information about possible interactions between acidic
bile-induced NF-κB activation and other central molecules in head
and neck cancer (Fig. 1).
The NF-κB/STAT3 crosstalk has been indicated to be
fundamental in inflammation-associated carcinogenesis in head and
neck cancer (108,119,120).
Application of NF-κB inhibitors successfully prevented the acidic
bile-induced activation and nuclear translocation of STAT3, which
is an important regulator of cell proliferation, and reduced the
transcriptional levels of IL6 and STAT3, in treated
hypopharyngeal cells (19–25) (Fig. 1).
These data strongly support the theory that the acidic bile-induced
activation of IL-6/STAT3 is NF-κB-dependent (108) (in a paragraph below, the role of
STAT3 in bile-induced carcinogenesis is discussed). In addition,
prior findings implied strong interactions between NF-κB and STAT3
in acidic bile-exposed premalignant HM (13,14),
further supporting the theory that the inflammatory response
induced by acidic bile may increase the risk of laryngopharyngeal
cancer.
EGFR is frequently overexpressed in HNSCC (121,122).
Crosstalk between NF-κB and downstream pathways of EGFR has been
observed (123,124). Although the exact role of EGFR in
bile-related hypopharyngeal carcinogenesis has remained to be
elucidated, the application of NF-κB inhibitors resulted in the
successful suppression of acidic bile-induced overexpression of
EGFR, supporting the interactions between NF-κB and EGFR
pathways during this process (Fig.
1). In addition, as both STAT3 and EGFR are important
contributors to HNSCC pathogenesis (80,121), the
above observations further emphasized the requirement to develop a
therapeutic strategy for targeting NF-κB in head and neck
malignancies and particularly in bile reflux-related HSCC.
Furthermore, NF-κB inhibition had a strong effect in
suppressing the acidic bile-induced overexpression of WNT5A
(Fig. 1), a factor related to
cancer-associated inflammation and epithelial-to-mesenchymal
transition (111), indicating that
NF-κB is able to mediate acidic bile-induced changes in
hypopharyngeal cell-cell interactions.
In addition, COX-2 is regularly highly overexpressed
during inflammatory and neoplastic processes (125) and is significantly overexpressed in
acidic bile-exposed HM (24,25). In vivo application of NF-κB
inhibitor significantly abrogated the acidic bile-induced
overexpression of PTGS2 (24,25)
(Fig. 1), further supporting the
regulatory role of NF-κB in early inflammatory and cancer-related
pathways, such as COX-2 (126).
In addition to the above, the PI3K/Akt pathway
(127) is one of the most frequently
activated pathways in head and neck cancer (128). The successful suppression of acidic
bile-induced AKT1 overexpression using topical application
of NF-κB inhibitor on murine HM suggested that NF-κB may mediate
acidic bile-induced deregulations of PI3K/Akt downstream pathways
(127) (Fig. 1).
In summary, the NF-κB pathway is a core central
pathway that interacts with multiple upstream and downstream
signaling pathways linked to the carcinogenic process. Using both a
specific NF-κB inhibitor, such as BAY 11-7082, and a more
pleiotropic NF-κB inhibitor, such as curcumin, it was documented
that the acidic bile-induced deregulations of cancer-related genes
or inflammatory factors are mediated by the NF-κB (Fig. 1). Furthermore, as curcumin is able to
prevent the bile-related anti-apoptotic effect independently of the
pH status, it may have an advantage over other NF-κB inhibitors. Of
note, curcumin specifically reduced a lower percentage of analyzed
NF-κB signaling genes compared to BAY 11-7082 (25 vs. 85%)
(20,21). Thus, curcumin may confer a clinical
advantage by preventing generalized suppression of NF-κB signaling,
which is essential to the basic metabolic function of healthy
mucosa and thereby reducing global toxicity (24,129).
Bile reflux-induced NF-κB-related
miRNA oncogenic phenotype
miRNA molecules have also been important in both
inflammation and cancer (130),
modulating the expression of genes by causing target mRNA
degradation or inhibiting their translation (131). The expression levels of certain
miRNAs, such as ‘oncomiRs’ and ‘tumor suppressor’ miRNAs, have been
indicated to be altered in tumor cells compared to normal cells
(upregulated or downregulated), and capable of contributing to
carcinogenesis, thereby demonstrating a significant regulatory role
in the multistep process of cancer initiation and progression
(132).
There is further evidence that miRNA markers, such
as ‘oncomiR’ miR-21 and ‘tumor suppressor’ miR-375, have a crucial
role in the initiation and progression of HNSCC (14,22,133,134).
Arantes et al (135) reported
the fundamental role of miR-21 as a biomarker in head and neck
carcinogenesis, while miR-375 has been proposed as a predictive
biomarker for early diagnosis in laryngeal cancer (136). In addition, interactions between
NF-κB and miRNA markers, such as miR-21, miR-34a and miR-451a, have
been importantly described by others in human cancer cells,
including HNSCC (104,137,138).
Specifically, a cluster of miRNA markers was reported to be
associated with NF-κB that may be associated with the aggressive
biological behavior of HNSCC (104).
Explorations by our group (14,15,19,14)
revealed that acidic bile caused deregulations of the expression of
oncogenic miRNA markers, previously associated with
laryngopharyngeal cancer (133–141).
Specifically, the ‘oncomiRs’ miR-21, miR-192 and miR-155, and the
‘tumor suppressors’ miR-34a, miR-375, miR-451a, miR-99a and miR-504
were indicated to be significantly altered in exposed HHCs and
murine laryngopharyngeal mucosa (14,15,19,14)
(Fig. 2). Of note, results from our
group (14,15) highlighted the role of miR-21 and
miR-375 deregulations in acidic bile-related neoplasia.
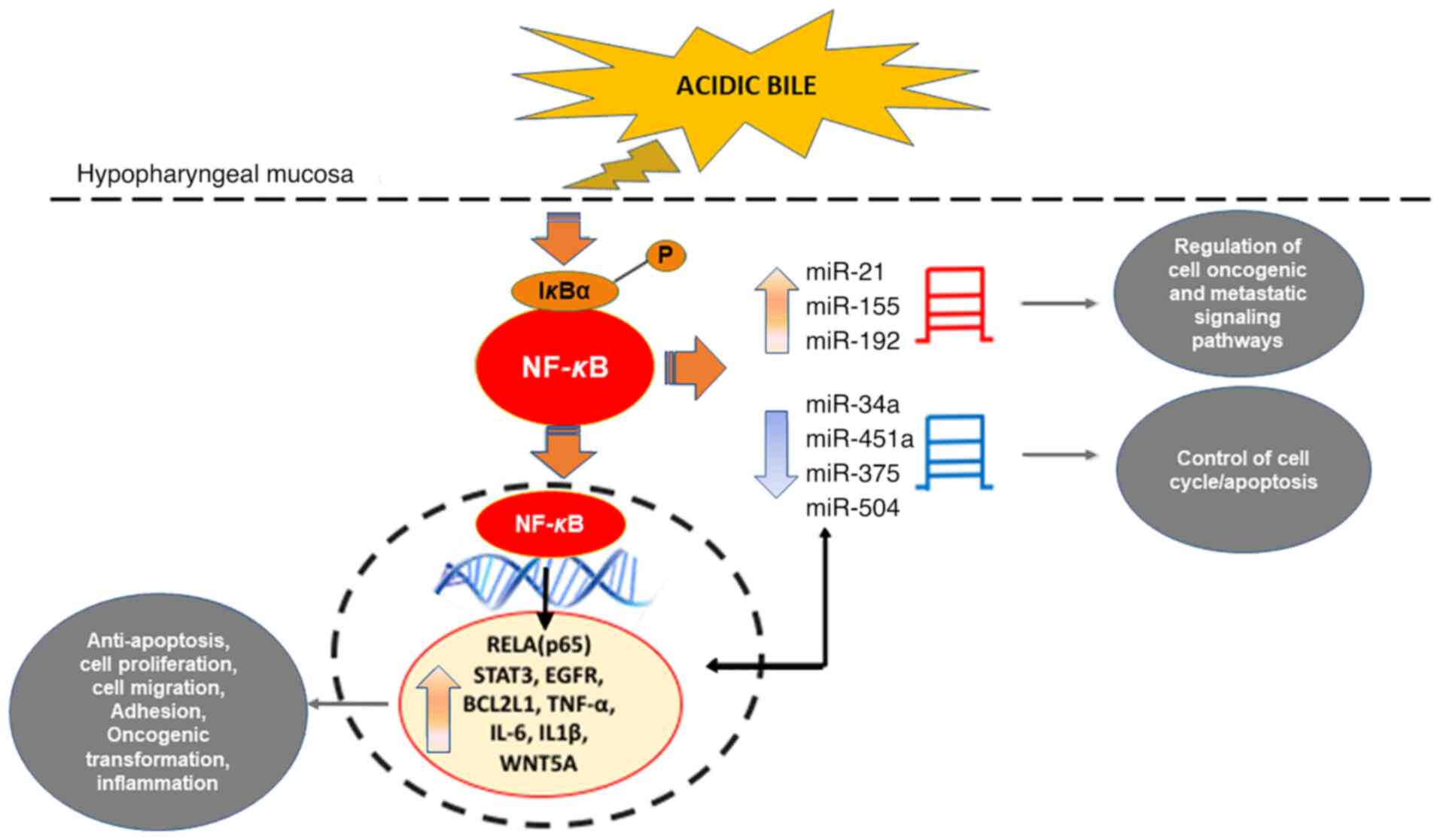
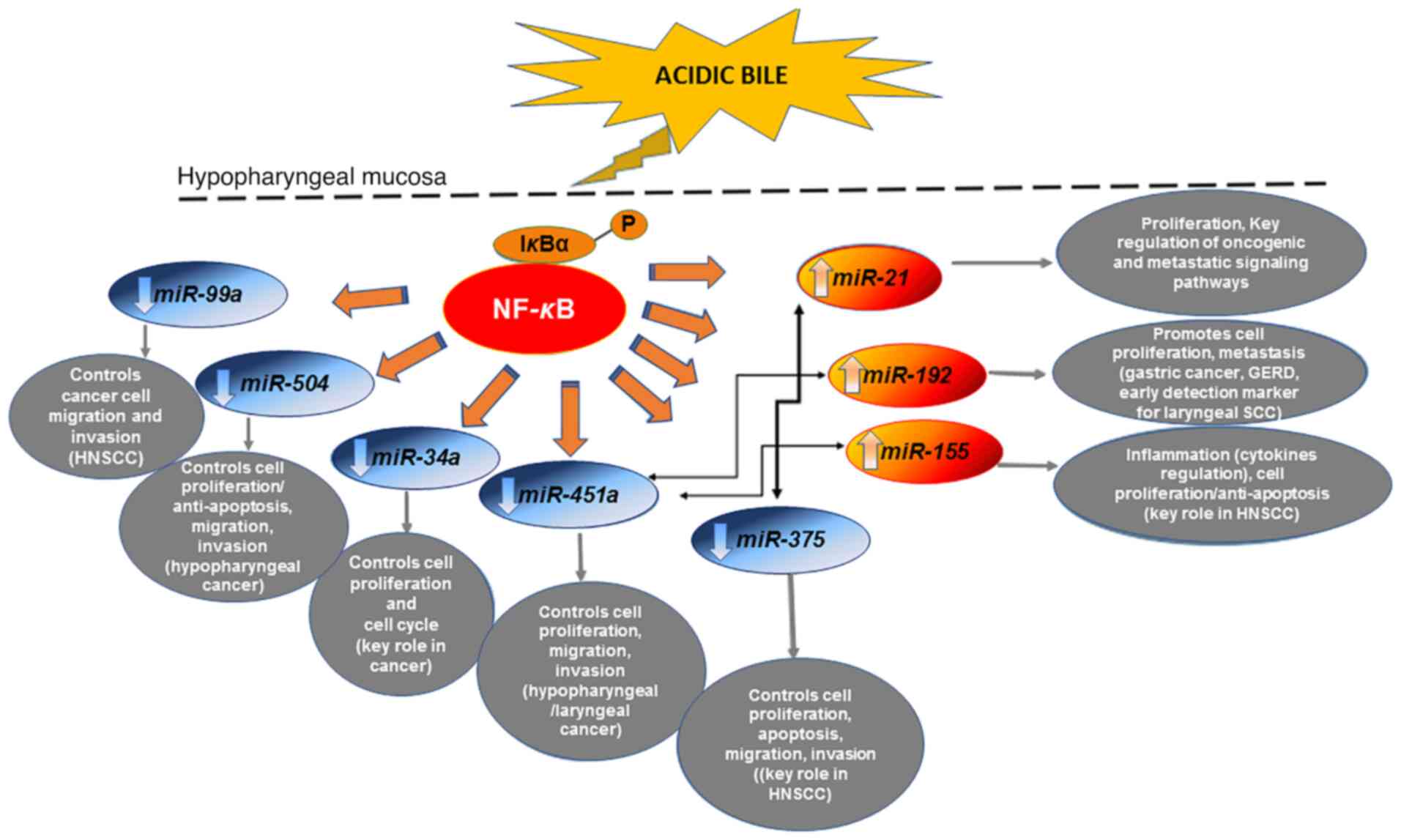 | Figure 2.The mechanistic role of NF-κB in the
miRNA oncogenic phenotype induced by bile reflux in hypopharyngeal
carcinogenesis. Chronic stimulation of laryngopharyngeal mucosa by
acidic bile induces constitutive activation of NF-κB, producing
upregulation of ‘oncomiRs’ miR-21, miR-155 and
miR-192, previously associated with oncogenic signaling
pathways in head and neck cancer and GERD. The acidic bile-induced
activation of NF-κB in treated hypopharyngeal mucosa is capable of
downregulating ‘tumor suppressor’ miR-34a, miR-451a, miR-375,
miR-99a and miR-504, are known to control the cell cycle
and are frequently affected in head and neck cancer. Acidic
bile-induced expression levels of ‘oncomiRs’ exhibited an inverse
correlation with ‘tumor suppressor’ miRNAs that appears to be
regulated by NF-κB. miRNA/miR, microRNA; GERD, gastroesophageal
reflux disease; HNSCC, head and neck squamous cell carcinoma. |
In addition, findings from HSCC tumor specimens
from patients with documented bile reflux supported its strong
association with upregulation of ‘oncomiR’ miR-21 and
downregulation of ‘tumor suppressors’ miR-34a and particularly of
miR-375, along with strong positivity for NF-κB (18) (Fig. 2).
Bile reflux-associated HSCC was also associated with a marked
reduction of ‘tumor suppressors’ miR-489, miR-504 and miR-99a
compared with their adjacent non-pathologic tissue (18), suggesting their involvement in the
onset and progression of HSCC (142,143).
Finally, bile exposure-associated HSCC exhibited differential
expression of miR-489 and miR-504, and particularly of miR-375,
compared to bile-negative HSCC, which had significantly lower NF-κB
levels. A previous view by our group (18) suggested that these miRNA markers may
have a distinct role in biliary reflux-associated hypopharyngeal
cancer.
Application of BAY 11-7082 in HHCs or murine HM was
proven to prevent miRNA deregulations caused by acidic bile,
providing insight into the interactions of transcriptionally active
NF-κB with cancer-related miRNA markers (19,22,23,25).
Specifically, it was demonstrated that BAY 11-7082 is able to
effectively reverse the acidic bile-induced downregulation of
‘tumor suppressor’ miR-451a and miR-99a, and upregulation of
‘oncomiRs’ miR-21, miR-155 and miR-192 (19,22,23,25)
(Fig. 2), which are considered
important markers for poor prognosis in head and neck cancer or
linked to gastroesophageal reflux (22,135,144–150).
Other studies have also indicated that NF-κB has a direct
regulatory effect on the expression of miR-21 and miR-155 through
their binding promoters (151,152).
Through in vitro and in vivo applications of BAY
11-7082 on hypopharyngeal cells and mucosa, respectively, a direct
effect of acidic bile on the above miRNAs was demonstrated shortly
after exposure (22,23,25). This
observation proposed the use of these miRNAs as biomarkers of early
neoplastic events in acidic bile-exposed HM, strongly supporting
the role of NF-κB as a mediator in this process. In addition,
topical in vivo application of BAY 11-7082 either before,
after or simultaneous to acidic bile significantly inhibited the
acidic bile-induced upregulation of ‘oncomiR’ miR-192 (19,25),
previously associated with GERD (148), and downregulation of ‘tumor
suppressor’ miR-504, a promising target for HSCC (142) (Fig.
2). This observation also suggested the utility of these miRNAs
as biomarkers of early neoplastic events in acidic bile-exposed
HM.
In summary, the above preclinical data provided
evidence of the role of NF-κB as a regulator of miR-192, miR-21,
miR-155, miR-451a, miR-375, miR-99a and miR-504 (Fig. 2), and proposed these miRNAs as
potential therapeutic targets of bile-related mutagenic evolution
in the HM.
Interactions between NF-κB-related
mRNA and miRNA phenotypes
Previous studies suggested that interactions
between miRNAs and mRNA molecules may be NF-κB-dependent during
carcinogenesis (141,153,154).
Rokavec et al (153) proposed
that miRNA molecules, such as miR-34a, interact with STAT3 in an
NF-κB-dependent manner. According to Tili et al (154), permanent upregulation of miR-155 may
mediate a prolonged inflammatory reaction leading to cancer.
It has been proposed that acidic bile-induced NF-κB
activation, BCL2 overexpression and significant alterations
of oncogenic EGFR, STAT3, TNF-α, IL6, IL1β and WNT5A
may directly or indirectly interact with cancer-related miRNA
markers, such as ‘oncomiRs’ miR-21 and miR-155, as well as ‘tumor
suppressors’ miR-34a, miR-375 and miR-451a (14–16,141)
(Fig. 3). These observations suggest
that inflammatory episodes caused by acidic bile may be associated
with downstream oncogenic pathways and may be effectively prevented
by NF-κB inhibition.
NF-κB as a challenging target for
cancer therapy
Recent findings documenting the crucial role of
NF-κB in bile reflux-related hypopharyngeal carcinogenesis, pose a
challenge for researchers and clinicians on how to identify
patients who are more likely to benefit from NF-κB inhibition
treatment. An NF-κB-related gene expression signature associated
with bile reflux in patients with HSCC (Figs. 1 and 2)
may provide a better prediction for inhibition selection and also
allow the development of diagnostic and prognostic biomarkers of
NF-κB inhibition response.
Although NF-κB targeted therapy has been already
applied in clinical practice with promising results in anti-cancer
therapy (129,155,156),
including HNSCC treatment (157,158),
there is an increasing effort in the pharmaceutical industry to
develop advanced NF-κB inhibitors (129,159,160).
In particular, research focuses on the identification of IKK/NF-κB
inhibitors for targeted therapy that would prevent NF-κB activation
without affecting other signaling pathways and selectively affect
malignant cells rather than normal cells. As one of the major
adverse effects of using NF-κB inhibitors as anticancer drugs is
their ability to impair innate immunity when applied in excessive
and prolonged periods (129,159–161),
research should also focus on both minimizing systemic toxicity and
prevention of long-term immunosuppression. Thus, an ideal
inhibition of NF-κB should be transient and reversible, as well as
effective when combined with other anti-cancer treatments. Prior
in vivo explorations strongly supported the effectiveness of
models using intermittent and topical treatment as opposed to
prolonged and systemic treatment. Specifically, the marked efficacy
of short-term topical treatment with NF-κB inhibitors, such as
curcumin (2 mg/kg/day) and BAY 11-7082 (6.25 mg/kg/day), in
suppressing bile reflux-induced early preneoplastic changes in the
hypopharynx (19,24,25),
strongly supports the view that non-systemic and transient NF-κB
inhibition may be clinically feasible in preventing
bile-reflux-related oncogenic effects. This area of research is
progressing rapidly; however, another adverse effect is of high
importance prior to the targeting of IKK or NF-κB in the clinic.
This refers to the enhanced production of IL-1β and related
cytokines by inhibitors of NF-κB activation during bacterial
infections (161), suggesting the
short-term use of NF-κB inhibitors in combination with
antibiotics.
Role of STAT3 in bile reflux-related
hypopharyngeal carcinogenesis
The STAT3 oncogene (162) is a transcription factor central to
head and neck cancer (119,120,122).
In addition to the significant role of NF-κB, recent preliminary
data from our team indicate the important role of STAT3 in
bile-related hypopharyngeal carcinogenesis by promoting early
oncogenic molecular events, including cancer-related inflammatory
molecules IL6, TNF-α and RELA (p65).
Specifically, using three different inhibitors with
each blocking a different step of STAT3 upstream signaling, such as
nifuroxazide, SI3-201 and STA-21 (163–165),
preliminary data from our group were obtained regarding the
mechanism of the effects of bile. In detail, acidic bile is able to
induce constitutive activation of STAT3 that may not be exclusively
dependent on JAK/STAT3 upstream signaling (165), but it may also be stimulated by
alternative signaling, such as EGFR. Although targeting STAT3,
either by its knockdown or its pharmacological inhibition, had a
minimal effect on nuclear or total phosphorylated NF-κB (p65 S536)
protein levels, it was observed to contribute, among others, to the
transcriptional activation of NF-κB. As mentioned above, previous
findings of NF-κB inhibition, using BAY 11-7082, had determined a
role of NF-κB in acidic bile-induced activation of STAT3 (19–25).
All of these observations suggest possible
molecular crosstalk between the NF-κB and STAT3 transcription
factor associated signaling pathways in bile reflux-related
inflammation and tumorigenesis in the hypopharynx, as similarly
proposed in HNSCC (108,119,120).
Bile-induced DNA damage
One of the principal questions regarding the
effects of bile on cellular physiology was how bile refluxate
induces DNA damage. Dvorak et al (59), suggested that bile at acidic pH may
potentially induce DNA damage in esophageal cells, speculating that
chronic exposure to bile acids at low pH may result in increased
genomic instability, abnormal cell signaling and resistance to
apoptosis. According to Goldman et al (166), bile in combination with acid, but
not acid alone, immediately activates all three isoforms of nitric
oxide (NO) synthase, a family of enzymes catalyzing the production
of NO, which links chronic inflammatory diseases and reactive
oxygen/nitrogen species (ROS/RNS) with cancer. They also indicated
that bile in combination with acid increased intracellular
acidification and DNA damage in esophageal cells, which may lead to
mutations and cancer progression (166). Bernstein et al (31,167)
proposed that de-conjugated secondary bile acids, such as DCA, are
capable of inducing DNA damage, giving rise to cancer due to the
accumulation of mutations. Specifically, DCA was determined to
induce increased intracellular production of ROS/RNS, resulting in
increased oxidative stress and DNA damage (64,168,169).
Recent studies by our group (15,16,19)
documented that conjugated bile acids led to the upregulation of
γ-H2AX (pS139). γ-H2AX is a consistent marker of DNA DSBs (170,171),
which was profoundly increased in bile-treated hypopharyngeal cells
or HM at acidic pH, compared to neutral pH, acid alone or neutral
control conditions (15,19). Subsequently, it was documented that
bile-treated HM at weakly acidic pH, with or without DCA, also
induced DNA damage in exposed HM (16). Specifically, premalignant and
malignant lesions caused by acidic bile demonstrated increased
levels of nuclear γ-H2AX, as well as DNA/RNA oxidative damage
(Fig. 4) (15,16).
According to previous findings, increased oxidative
damage may result in high levels of ROS or DSBs incurring direct
DNA damage (168,172,173),
which may potentially lead to tumor-initiating mutations in head
and neck cancer (174). All of these
results advocate the theory that acidic bile-induced DNA damage may
contribute to its mutated phenotype (Fig.
4). In addition, ROS is able to activate several
cancer-associated signaling pathways, including NF-κB (164), concluding that acidic bile may
contribute to evasion of apoptosis and/or proliferation of mutated
hypopharyngeal epithelial cells, resulting in malignant lesions of
HM.
In parallel, chronic exposure of murine
hypopharyngeal epithelium to acidic bile was observed to induce a
systematic release of inflammatory molecules, such as IL-6 and
TNF-α, which are considered central to head and neck carcinogenesis
(91). This systematic release of
inflammatory molecules is able to maintain the constitutive release
of other cancer-related cytokines in the microenvironment of the
exposed epithelium. It is known that the chronic inflammatory
microenvironment may lead to the production of activated
inflammatory cells that may also serve as sources of ROS and
reactive nitrogen intermediates (RNI), which are capable of
inducing DNA damage and genomic instability and so promote
mutations in neighboring epithelial cells (175,176).
In addition, inflammatory cytokines contribute to increased
intracellular ROS and RNI production in pre-malignant cells. In
conclusion, the chronic inflammatory microenvironment caused by
bile reflux may be one of the main factors in hypopharyngeal
carcinogenesis.
Possible interactions of bile refluxate with
hypopharyngeal mucosa
How acidic bile interacts with HM to exert its
harmful effect, causing DNA damage and promoting NF-κB-related
anti-apoptotic processes, leading to its malignant transformation,
has remained to be fully elucidated. According to Li and Cao
(177), bile acids may interact with
membrane receptors, such as Takeda G-protein coupled receptor
(TGR5). Their study also indicated that TGR5 is able to mediate
bile reflux-induced DNA damage in esophageal cells (177). Another cell membrane receptor that
is able to interact with bile acids is the sphingosine-1-phosphate
receptor 1, known as S1PR1 or S1P1 (178,179).
Both above-mentioned receptors were previously associated with
lower esophageal cancer related to bile reflux (180,181).
In particular, TGR5 was reported to be expressed in both
adenocarcinoma and squamous cell carcinoma of the lower esophagus
(181), while S1PR1 has been
associated with squamous cell carcinoma of the head and neck
(179).
Other studies provided evidence that bile acids are
able to activate nuclear farnesoid X receptors (FXRs) (182,183),
suggesting their contribution to pre-neoplastic changes (183) of the lower esophagus. Although
several nuclear receptors (NRs) have been identified in the head
and neck (184), an association
between NRs and bile acids has not yet been described in HNSCC.
Prior studies suggested a mutually antagonistic relationship
between FXR and NF-κB activation (185,186)
and proposed FXR receptors as useful targets for esophageal
adenocarcinoma (182). However, the
exact mechanism by which FXR affects the expression of
proinflammatory molecules, such as NF-κB in either the lower or
upper esophagus remains elusive and the role of FXR in bile
reflux-related carcinogenesis deserves further exploration. Further
investigation in the hypopharynx may identify specific receptors
activated by acidic bile and clarify the role of NRs, such as FXR,
and cell membrane receptors, such as TGR5 and S1PR1, in this
process.
Conclusion
Recent in vitro and in vivo data
provide evidence on bile reflux-associated hypopharyngeal
carcinogenesis. The composition of biliary refluxate, such as
conjugated bile acids, and acidity are pivotal factors in promoting
DNA damage, as well as histologic and molecular changes in the HM,
most likely through the constitutive activation of NF-κB. Chronic
acidic bile exposure can cause increased oxidative damage, DSBs,
overproduction of cytokines and cell-cell interaction changes,
which are critical elements of tumor initiation and progression
(187), possibly through
derangements in both pre-neoplastic/neoplastic cells and their
microenvironment. In parallel, acidic bile-induced constitutive
activation of NF-κB can promote oncogenic mRNA and miRNA
phenotypes, contributing to the proliferation of mutated cells and
thus giving rise to the malignant transformation of the exposed HM.
Primary data also support the contributing role of STAT3 in this
process. Further investigation of the proposed mechanisms mediating
bile-induced DNA damage, the tumor microenvironment and downstream
oncogenic signaling pathways in HM, as well as the identification
of specific receptors that may interact with bile, will contribute
to innovative approaches to the diagnosis and prevention of
laryngopharyngeal malignancies, as well as to the improvement of
current therapeutic approaches to LPR-related carcinogenesis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
Data sharing is not applicable.
Authors' contributions
DPV and SGD were involved in the conceptualization
of this review article. DPV, SGD, PGD and BLJ were involved in
searching the literature. DPV, PGD and SGD were involved in the
writing of the original draft. DPV, SGD, PGD and BLJ reviewed and
edited the article. All authors have read and approved the final
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HM
|
hypopharyngeal mucosa
|
|
HHCs
|
human normal hypopharyngeal cells
|
References
|
1
|
Hashibe M, Boffetta P, Zaridze D, Shangina
O, Szeszeni-Dabrowska N, Mates D, Fabiánová E, Rudnai P and Brennan
P: Contribution of tobacco and alcohol to the high rates of
squamous cell carcinoma of the supraglottis and glottis in Central
Europe. Am J Epidemiol. 165:814–820. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Curado MP and Hashibe M: Recent changes in
the epidemiology of head and neck cancer. Curr Opin Oncol.
21:194–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Talamini R, Bosetti C, La Vecchia C, Dal
Maso L, Levi F, Bidoli E, Negri E, Pasche C, Vaccarella S, Barzan L
and Franceschi S: Combined effect of tobacco and alcohol on
laryngeal cancer risk: A case-control study. Cancer Causes Control.
13:957–964. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pöschl G and Seitz HK: Alcohol and cancer.
Alcohol Alcohol. 39:155–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Galli J, Cammarota G, De Corso E, Agostino
S, Cianci R, Almadori G and Paludetti G: Biliary laryngopharyngeal
reflux: A new pathological entity. Curr Opin Otolaryngol Head Neck
Surg. 14:128–132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tutar H, Erdamar H, Köybaşioğlu A, Dinç
AE, Ceylan A and Uslu S: Can bile acids be an etiological factor
for laryngeal carcinoma? ORL J Otorhinolaryngol Relat Spec.
73:156–161. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Geterud A, Bove M and Ruth M:
Hypopharyngeal acid exposure: An independent risk factor for
laryngeal cancer? Laryngoscope. 113:2201–2205. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sereg-Bahar M, Jerin A and
Hocevar-Boltezar I: Higher levels of total pepsin and bile acids in
the saliva as a possible risk factor for early laryngeal cancer.
Radiol Oncol. 49:59–64. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Altman KW, Prufer N and Vaezi MF: A review
of clinical practice guidelines for reflux disease: toward creating
a clinical protocol for the otolaryngologist. Laryngoscope.
121:717–723. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Assimakopoulos D and Patrikakos G: The
role of gastroesophageal reflux in the pathogenesis of laryngeal
carcinoma. Am J Otolaryngol. 23:351–357. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sasaki CT, Marotta J, Hundal J, Chow J and
Eisen RN: Bile-induced laryngitis: Is there a basis in evidence?
Ann Otol Rhinol Laryngol. 114:192–197. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sasaki CT, Issaeva N and Vageli DP: In
vitro model for gastroduodenal reflux-induced nuclear factor-kappaB
activation and its role in hypopharyngeal carcinogenesis. Head
Neck. 38 (Suppl 1):E1381–E1391. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vageli DP, Prasad ML and Sasaki CT:
Gastro-duodenal fluid induced nuclear Factor-κappaB activation and
early pre-malignant alterations in murine hypopharyngeal mucosa.
Oncotarget. 7:5892–5908. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sasaki CT and Vageli DP: MiR-21, miR-155,
miR-192, and miR-375 deregulations related to NF-kappaB activation
in gastroduodenal Fluid-Induced early Preneoplastic lesions of
laryngeal mucosa in vivo. Neoplasia. 18:329–338. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sasaki CT, Doukas SG, Costa J and Vageli
DP: The progressive mutagenic effects of acidic bile refluxate in
hypopharyngeal squamous cell carcinogenesis: New insights. Cancers
(Basel). 12:10642020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sasaki CT, Doukas SG, Doukas PG and Vageli
DP: Weakly acidic bile is a risk factor for hypopharyngeal
carcinogenesis evidenced by DNA damage, antiapoptotic function, and
premalignant dysplastic lesions in vivo. Cancers (Basel).
13:8522021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Doukas SG, Cardoso B, Tower JI, Vageli DP
and Sasaki CT: Biliary tumorigenic effect on hypopharyngeal cells
is significantly enhanced by pH reduction. Cancer Med. 8:4417–4427.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sasaki CT, Doukas SG, Costa J and Vageli
DP: Biliary reflux as a causal factor in hypopharyngeal carcinoma:
New clinical evidence and implications. Cancer. 125:3554–3565.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sasaki CT, Doukas SG and Vageli DP: In
Vivo Short-Term topical application of BAY 11-7082 prevents the
acidic Bile-Induced mRNA and miRNA oncogenic phenotypes in exposed
Murine Hypopharyngeal Mucosa. Neoplasia. 20:374–386. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vageli DP, Doukas SG, Spock T and Sasaki
CT: Curcumin prevents the bile reflux-induced NF-κB-related mRNA
oncogenic phenotype, in human hypopharyngeal cells. J Cell Mol Med.
22:4209–4220. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vageli DP, Doukas SG and Sasaki CT:
Inhibition of NF-kappaB prevents the acidic bile-induced oncogenic
mRNA phenotype, in human hypopharyngeal cells. Oncotarget.
9:5876–5891. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Doukas SG, Vageli DP and Sasaki CT: NF-κB
inhibition reverses acidic bile-induced miR-21, miR-155, miR-192,
miR-34a, miR-375 and miR-451a deregulations in human hypopharyngeal
cells. J Cell Mol Med. 22:2922–2934. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Doukas PG, Vageli DP, Doukas SG and Sasaki
CT: Temporal characteristics of NF-κB inhibition in blocking
bile-induced oncogenic molecular events in hypopharyngeal cells.
Oncotarget. 10:3339–3351. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Doukas SG, Doukas PG, Sasaki CT and Vageli
D: The in vivo preventive and therapeutic properties of curcumin in
bile reflux-related oncogenesis of the hypopharynx. J Cell Mol Med.
24:10311–10321. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vageli DP, Kasle D, Doukas SG, Doukas PG
and Sasaki CT: The temporal effects of topical NF-κB inhibition, in
the in vivo prevention of bile-related oncogenic mRNA and miRNA
phenotypes in murine hypopharyngeal mucosa: A preclinical model.
Oncotarget. 11:3303–3314. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hofmann AF: Chemistry and enterohepatic
circulation of bile acids. Hepatology. 4 (Suppl 5):4S–14S. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Greek Medicine, . http://www.greekmedicine.net/b_p/Four_Humors.html
|
|
28
|
Rather LJ: Disturbance of function
(functio laesa): The legendary fifth cardinal sign of inflammation,
added by Galen to the four cardinal signs of Celsus. Bull NY Acad
Med. 47:303–322. 1971.PubMed/NCBI
|
|
29
|
Virchow R and Rather LJ: Disease, Life,
and Man: Selected Essays. Stanford University Press; Stanford, CA:
1958
|
|
30
|
Cook JW: Carcinogenic chemical agents.
Yale J Biol Med. 11:1–13. 1938.PubMed/NCBI
|
|
31
|
Bernstein H, Bernstein C, Payne CM,
Dvorakova K and Garewal H: Bile acids as carcinogens in human
gastrointestinal cancers. Mutat Res. 589:47–65. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kuwahara A, Saito T and Kobayashi M: Bile
acids promote carcinogenesis in the remnant stomach of rats. J
Cancer Res Clin Oncol. 115:423–428. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hill MJ: Bile flow and colon cancer. Mutat
Res. 238:313–320. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bayerdörffer E, Mannes GA, Ochsenkühn T,
Dirschedl P, Wiebecke B and Paumgartner G: Unconjugated secondary
bile acids in the serum of patients with colorectal adenomas. Gut.
36:268–273. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nehra D, Howell P, Williams CP, Pye JK and
Beynon J: Toxic bile acids in gastro-oesophageal reflux disease:
Influence of gastric acidity. Gut. 44:598–602. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vaezi MF and Richter JE: Double reflux:
Double trouble. Gut. 44:590–592. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vaezi MF, Singh S and Richter JE: Role of
acid and duodenogastric reflux in esophageal mucosal injury: A
review of animal and human studies. Gastroenterology.
108:1897–1907. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gotley DC, Morgan AP and Cooper MJ: Bile
acid concentrations in the refluxate of patients with reflux
oesophagitis. Br J Surg. 75:587–590. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kauer WK, Peters JH, DeMeester TR, Ireland
AP, Bremner CG and Hagen JA: Mixed reflux of gastric and duodenal
juices is more harmful to the esophagus than gastric juice alone.
The need for surgical therapy re-emphasized. Ann Surg. 222:525–531.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kauer WK, Peters JH, DeMeester TR,
Feussner H, Ireland AP, Stein HJ and Siewert RJ: Composition and
concentration of bile acid reflux into the esophagus of patients
with gastroesophageal reflux disease. Surgery. 122:874–881. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Domellof L, Reddy BS and Weisburger JH:
Microflora and deconjugation of bile acids in alkaline reflux after
partial gastrectomy. Am J Surg. 140:291–295. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fein M, Peters JH, Chandrasoma P, Ireland
AP, Oberg S, Ritter MP, Bremner CG, Hagen JA and DeMeester TR:
Duodenoesophageal reflux induces esophageal adenocarcinoma without
exogenous carcinogen. J Gastrointest Surg. 2:260–268. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
McQuaid KR, Laine L, Fennerty MB, Souza R
and Spechler SJ: Systematic review: The role of bile acids in the
pathogenesis of gastro-oesophageal reflux disease and related
neoplasia. Aliment Pharmacol Ther. 34:146–165. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Oh DS, Hagen JA, Fein M, Bremner CG, Dunst
CM, Demeester SR, Lipham J and Demeester TR: The impact of reflux
composition on mucosal injury and esophageal function. J
Gastrointest Surg. 10:787–796. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sweet MP, Patti MG, Hoopes C, Hays SR and
Golden JA: Gastro-oesophageal reflux and aspiration in patients
with advanced lung disease. Thorax. 64:167–173. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Covington MF, Krupinski E, Avery RJ and
Kuo PH: Classification schema of symptomatic enterogastric reflux
utilizing sincalide augmentation on hepatobiliary scintigraphy. J
Nucl Med Technol. 42:198–202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lewin JS, Gillenwater AM, Garrett JD,
Bishop-Leone JK, Nguyen DD, Callender DL, Ayers GD and Myers JN:
Characterization of laryngopharyngeal reflux in patients with
premalignant or early carcinomas of the larynx. Cancer.
97:1010–1014. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Johnston N, Ondrey F, Rosen R, Hurley BP,
Gould J, Allen J, DelGaudio J and Altman KW: Airway reflux. Ann N Y
Acad Sci. 1381:5–13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Adams J, Heintz P, Gross N, Andersen P,
Everts E, Wax M and Cohen J: Acid/pepsin promotion of
carcinogenesis in the hamster cheek pouch. Arch Otolaryngol Head
Neck Surg. 126:405–409. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Johnston N, Dettmar PW, Ondrey FG, Nanchal
R, Lee SH and Bock JM: Pepsin: Biomarker, mediator, and therapeutic
target for reflux and aspiration. Ann NY Acad Sci. 1434:282–289.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Johnston N, Wells CW, Samuels TL and
Blumin JH: Pepsin in nonacidic refluxate can damage hypopharyngeal
epithelial cells. Ann Otol Rhinol Laryngol. 118:677–685. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Del Negro A, Araújo MR, Tincani AJ,
Meirelles L, Martins AS and Andreollo NA: Experimental
carcinogenesis on the oropharyngeal mucosa of rats with
hydrochloric acid, sodium nitrate and pepsin. Acta Cir Bras.
23:337–342. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sasaki CT, Toman J and Vageli D: The in
vitro effect of Acidic-Pepsin on nuclear factor KappaB activation
and its related oncogenic effect on normal human hypopharyngeal
cells. PLoS One. 11:e01682692016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Doukas PG, Vageli DP, Sasaki CT and Judson
BL: Pepsin promotes activation of epidermal growth factor receptor
and downstream oncogenic pathways, at slightly acidic and neutral
pH, in exposed hypopharyngeal cells. Int J Mol Sci. 22:42752021.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Goldstein JL, Schlesinger PK, Mozwecz HL
and Layden TJ: Esophageal mucosal resistance. A factor in
esophagitis. Gastroenterol Clin North Am. 19:565–586. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Stamp D and Jenkins G: An overview of
bile-acid synthesis, chemistry and function. Bile Acids: Toxicology
and Bioactivity. Jenkins GJ and Hardie L: Royal Society of
Chemistry; Cambridge: 2008, Print: Issues in Toxicology; 4.
https://pubs.rsc.org/en/content/ebook/978-0-85404-846-5
View Article : Google Scholar
|
|
57
|
Stamp DH: Three hypotheses linking bile to
carcinogenesis in the gastrointestinal tract: certain bile salts
have properties that may be used to complement chemotherapy. Med
Hypotheses. 59:398–405. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ireland AP, Peters JH, Smyrk TC, DeMeester
TR, Clark GW, Mirvish SS and Adrian TE: Gastric juice protects
against the development of esophageal adenocarcinoma in the rat.
Ann Surg. 224:358–370. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Dvorak K, Payne CM, Chavarria M, Ramsey L,
Dvorakova B, Bernstein H, Holubec H, Sampliner RE, Guy N, Condon A,
et al: Bile acids in combination with low pH induce oxidative
stress and oxidative DNA damage: Relevance to the pathogenesis of
Barrett's oesophagus. Gut. 56:763–771. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kauer WK and Stein HJ: Role of acid and
bile in the genesis of Barrett's esophagus. Chest Surg Clin N Am.
12:39–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ulualp SO, Roland PS, Toohill RJ and
Shaker R: Prevalence of gastroesophagopharyngeal acid reflux
events: An evidence-based systematic review. Am J Otolaryngol.
26:239–244. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lillemoe KD, Gadacz TR and Harmon JW: Bile
absorption occurs during disruption of the esophageal mucosal
barrier. J Surg Res. 35:57–62. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sasaki CT, Hajek M, Doukas SG and Vageli
DP: The role of bile reflux and its related NF-κB activated pathway
in progression of hypopharyngeal squamous cell cancer. Oral Oncol.
105:1046682020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Hemmink GJ, Bredenoord AJ, Weusten BL,
Monkelbaan JF, Timmer R and Smout AJ: Esophageal pH-impedance
monitoring in patients with therapy-resistant reflux symptoms: ‘On’
or ‘off’ proton pump inhibitor? Am J Gastroenterol. 103:2446–2453.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Bernstein H, Payne CM, Bernstein C,
Schneider J, Beard SE and Crowley CL: Activation of the promoters
of genes associated with DNA damage, oxidative stress, ER stress
and protein malfolding by the bile salt, deoxycholate. Toxicol
Lett. 108:37–46. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Huo X, Juergens S, Zhang X, Rezaei D, Yu
C, Strauch ED, Wang JY, Cheng E, Meyer F, Wang DH, et al:
Deoxycholic acid causes DNA damage while inducing apoptotic
resistance through NF-κB activation in benign Barrett's epithelial
cells. Am J Physiol Gastrointest Liver Physiol. 301:G278–G286.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Langevin SM, Michaud DS, Marsit CJ, Nelson
HH, Birnbaum AE, Eliot M, Christensen BC, McClean MD and Kelsey KT:
Gastric reflux is an independent risk factor for laryngopharyngeal
carcinoma. Cancer Epidemiol Biomarkers Prev. 22:1061–1068. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Coca-Pelaz A, Rodrigo JP, Takes RP, Silver
CE, Paccagnella D, Rinaldo A, Hinni ML and Ferlito A: Relationship
between reflux and laryngeal cancer. Head Neck. 35:1814–1818. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Attwood SE, Smyrk TC, DeMeester TR,
Mirvish SS, Stein HJ and Hinder RA: Duodenoesophageal reflux and
the development of esophageal adenocarcinoma in rats. Surgery.
111:503–510. 1992.PubMed/NCBI
|
|
70
|
Fein M, Fuchs KH, Stopper H, Diem S and
Herderich M: Duodenogastric reflux and foregut carcinogenesis:
Analysis of duodenal juice in a rodent model of cancer.
Carcinogenesis. 21:2079–2084. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Miwa K, Hattori T and Miyazaki I:
Duodenogastric reflux and foregut carcinogenesis. Cancer. 75 (Suppl
6):S1426–S1432. 1995. View Article : Google Scholar
|
|
72
|
Fang Y, Chen H, Hu Y, Djukic Z, Tevebaugh
W, Shaheen NJ, Orlando RC, Hu J and Chen X: Gastroesophageal reflux
activates the NF-κB pathway and impairs esophageal barrier function
in mice. Am J Physiol Gastrointest Liver Physiol. 305:G58–G65.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
McAdam E, Haboubi HN, Griffiths AP, Baxter
JN, Spencer-Harty S, Davies C and Jenkins GJ: Reflux composition
influences the level of NF-κB activation and upstream kinase
preference in oesophageal adenocarcinoma cells. Int J Cancer.
136:527–535. 2015.PubMed/NCBI
|
|
74
|
Bus P, Siersema PD and van Baal JW: Cell
culture models for studying the development of Barrett's esophagus:
A systematic review. Cell Oncol (Dordr). 35:149–161. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hormi-Carver K, Zhang X, Zhang HY,
Whitehead RH, Terada LS, Spechler SJ and Souza RF: Unlike
esophageal squamous cells, Barrett's epithelial cells resist
apoptosis by activating the nuclear factor-κB pathway. Cancer Res.
69:672–677. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang H and Cho CH: Effect of NF-kappaB
signaling on apoptosis in chronic inflammation-associated
carcinogenesis. Curr Cancer Drug Targets. 10:593–599. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
DiDonato JA, Mercurio F and Karin M:
NF-kappaB and the link between inflammation and cancer. Immunol
Rev. 246:379–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hoesel B and Schmid JA: The complexity of
NF-kappaB signaling in inflammation and cancer. Mol Cancer.
12:862013. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Nottingham LK, Yan CH, Yang X, Si H,
Coupar J, Bian Y, Cheng TF, Allen C, Arun P, Gius D, et al:
Aberrant IKKα and IKKβ cooperatively activate NF-κB and induce
EGFR/AP1 signaling to promote survival and migration of head and
neck cancer. Oncogene. 33:1135–1147. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Stadler ME, Patel MR, Couch ME and Hayes
DN: Molecular biology of head and neck cancer: Risks and pathways.
Hematol Oncol Clin North Am. 22:1099–1124. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Molinolo AA, Amornphimoltham P, Squarize
CH, Castilho RM, Patel V and Gutkind JS: Dysregulated molecular
networks in head and neck carcinogenesis. Oral Oncol. 45:324–334.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
King KE, Ponnamperuma RM, Allen C, Lu H,
Duggal P, Chen Z, Van Waes C and Weinberg WC: The p53 homologue
DeltaNp63alpha interacts with the nuclear factor-kappaB pathway to
modulate epithelial cell growth. Cancer Res. 68:5122–5131. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Jackson-Bernitsas DG, Ichikawa H, Takada
Y, Myers JN, Lin XL, Darnay BG, Chaturvedi MM and Aggarwal BB:
Evidence that TNF-TNFR1-TRADD-TRAF2-RIP-TAK1-IKK pathway mediates
constitutive NF-kappaB activation and proliferation in human head
and neck squamous cell carcinoma. Oncogene. 26:1385–1397. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dong J, Jimi E, Zeiss C, Hayden MS and
Ghosh S: Constitutively active NF-kappaB triggers systemic
TNFalpha-dependent inflammation and localized TNFalpha-independent
inflammatory disease. Genes Dev. 24:1709–1717. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Foxwell BM, Bondeson J, Brennan F and
Feldmann M: Adenoviral transgene delivery provides an approach to
identifying important molecular processes in inflammation: Evidence
for heterogenecity in the requirement for NF-kappaB in tumour
necrosis factor production. Ann Rheum Dis. 59 (Suppl 1):i54–i59.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Guyer RA and Macara IG: Loss of the
polarity protein PAR3 activates STAT3 signaling via an atypical
protein kinase C (aPKC)/NF-κB/interleukin-6 (IL-6) axis in mouse
mammary cells. J Biol Chem. 290:8457–8468. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhao Y, Zhang C, Huang Y, Yu Y, Li R, Li
M, Liu N, Liu P and Qiao J: Up-regulated expression of WNT5a
increases inflammation and oxidative stress via PI3K/AKT/NF-κB
signaling in the granulosa cells of PCOS patients. J Clin
Endocrinol Metab. 100:201–211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Bo H, Zhang S, Gao L, Chen Y, Zhang J,
Chang X and Zhu M: Upregulation of Wnt5a promotes
epithelial-to-mesenchymal transition and metastasis of pancreatic
cancer cells. BMC Cancer. 13:4962013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Klein JD and Grandis JR: The molecular
pathogenesis of head and neck cancer. Cancer Biol Ther. 9:1–7.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Allen CT, Ricker JL, Chen Z and Van Waes
C: Role of activated nuclear factor-kappaB in the pathogenesis and
therapy of squamous cell carcinoma of the head and neck. Head Neck.
29:959–971. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Loercher A, Lee TL, Ricker JL, Howard A,
Geoghegen J, Chen Z, Sunwoo JB, Sitcheran R, Chuang EY, Mitchell
JB, et al: Nuclear factor-kappaB is an important modulator of the
altered gene expression profile and malignant phenotype in squamous
cell carcinoma. Cancer Res. 64:6511–6523. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Chung CH, Parker JS, Ely K, Carter J, Yi
Y, Murphy BA, Ang KK, El-Naggar AK, Zanation AM, Cmelak AJ, et al:
Gene expression profiles identify epithelial-to-mesenchymal
transition and activation of nuclear factor-kappaB signaling as
characteristics of a high-risk head and neck squamous cell
carcinoma. Cancer Res. 66:8210–8218. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Lee TL, Yang XP, Yan B, Friedman J, Duggal
P, Bagain L, Dong G, Yeh NT, Wang J, Zhou J, et al: A novel nuclear
factor-kappaB gene signature is differentially expressed in head
and neck squamous cell carcinomas in association with TP53 status.
Clin Cancer Res. 13:5680–5691. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Coussens LM and Werb Z: Inflammation and
cancer. Nature. 420:860–867. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Souza RF: From reflux esophagitis to
esophageal adenocarcinoma. Dig Dis. 34:483–490. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Wroblewski LE, Peek RM Jr and Wilson KT:
Helicobacter pylori and gastric cancer: Factors that modulate
disease risk. Clin Microbiol Rev. 23:713–739. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Shaheen N and Ransohoff DF:
Gastroesophageal reflux, Barrett esophagus, and esophageal cancer:
Scientific review. JAMA. 287:1972–1981. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Vander Broek R, Snow GE, Chen Z and Van
Waes C: Chemoprevention of head and neck squamous cell carcinoma
through inhibition of NF-kappaB signaling. Oral Oncol. 50:930–941.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Baldwin AS Jr: The NF-kappa B and I kappa
B proteins: New discoveries and insights. Annu Rev Immunol.
14:649–683. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Lee TL, Yeh J, Friedman J, Yan B, Yang X,
Yeh NT, Van Waes C and Chen Z: A signal network involving
coactivated NF-kappaB and STAT3 and altered p53 modulates
BAX/BCL-XL expression and promotes cell survival of head and neck
squamous cell carcinomas. Int J Cancer. 122:1987–1998. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Wheeler SE, Suzuki S, Thomas SM, Sen M,
Leeman-Neill RJ, Chiosea SI, Kuan CT, Bigner DD, Gooding WE, Lai SY
and Grandis JR: Epidermal growth factor receptor variant III
mediates head and neck cancer cell invasion via STAT3 activation.
Oncogene. 29:5135–5145. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yan B, Yang X, Lee TL, Friedman J, Tang J,
Van Waes C and Chen Z: Genome-wide identification of novel
expression signatures reveal distinct patterns and prevalence of
binding motifs for p53, nuclear factor-kappaB and other signal
transcription factors in head and neck squamous cell carcinoma.
Genome Biol. 8:R782007. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Yan B, Li H, Yang X, Shao J, Jang M, Guan
D, Zou S, Van Waes C, Chen Z and Zhan M: Unraveling regulatory
programs for NF-kappaB, p53 and microRNAs in head and neck squamous
cell carcinoma. PLoS One. 8:e736562013. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Sriuranpong V, Park JI, Amornphimoltham P,
Patel V, Nelkin BD and Gutkind JS: Epidermal growth factor
receptor-independent constitutive activation of STAT3 in head and
neck squamous cell carcinoma is mediated by the autocrine/paracrine
stimulation of the interleukin 6/gp130 cytokine system. Cancer Res.
63:2948–2956. 2003.PubMed/NCBI
|
|
106
|
Yang X, Lu H, Yan B, Romano RA, Bian Y,
Friedman J, Duggal P, Allen C, Chuang R, Ehsanian R, et al: ΔNp63
versatility regulates a Broad NF-κB gene program and promotes
squamous epithelial proliferation, migration, and inflammation.
Cancer Res. 71:3688–3700. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Du J, Romano RA, Si H, Mattox A, Bian Y,
Yang X, Sinha S, Van Waes C and Chen Z: Epidermal overexpression of
transgenic ΔNp63 promotes type 2 immune and myeloid inflammatory
responses and hyperplasia via NF-κB activation. J Pathol.
232:356–368. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Squarize CH, Castilho RM, Sriuranpong V,
Pinto DS Jr and Gutkind JS: Molecular cross-talk between the
NFkappaB and STAT3 signaling pathways in head and neck squamous
cell carcinoma. Neoplasia. 8:733–746. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Yap LF, Ahmad M, Zabidi MM, Chu TL, Chai
SJ, Lee HM, Lim PV, Wei W, Dawson C, Teo SH, et al: Oncogenic
effects of WNT5A in Epstein-Barr virus associated nasopharyngeal
carcinoma. Int J Oncol. 44:1774–1780. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Sherwood V, Chaurasiya SK, Ekström EJ,
Guilmain W, Liu Q, Koeck T, Brown K, Hansson K, Agnarsdóttir M,
Bergqvist M, et al: WNT5A-mediated β-catenin-independent signalling
is a novel regulator of cancer cell metabolism. Carcinogenesis.
35:784–794. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Asem MS, Buechler S, Wates RB, Miller DL
and Stack MS: Wnt5a Signaling in Cancer. Cancers (Basel). 8:792016.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Nakanishi C and Toi M: Nuclear
factor-kappaB inhibitors as sensitizers to anticancer drugs. Nat
Rev Cancer. 5:297–309. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Meng Z, Lou S, Tan J, Xu K, Jia Q and
Zheng W: Nuclear factor-kappa B inhibition can enhance apoptosis of
differentiated thyroid cancer cells induced by 131I. PLoS One.
7:e335972012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Lin Y, Bai L, Chen W and Xu S: The NF-κB
activation pathways, emerging molecular targets for cancer
prevention and therapy. Expert Opin Ther Targets. 14:45–55. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Van Waes C: Nuclear factor-kappaB in
development, prevention, and therapy of cancer. Clin Cancer Res.
13:1076–1082. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Wilken R, Veena MS, Wang MB and Srivatsan
ES: Curcumin: A review of anti-cancer properties and therapeutic
activity in head and neck squamous cell carcinoma. Mol Cancer.
10:122011. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
LoTempio MM, Veena MS, Steele HL,
Ramamurthy B, Ramalingam TS, Cohen AN, Chakrabarti R, Srivatsan ES
and Wang MB: Curcumin suppresses growth of head and neck squamous
cell carcinoma. Clin Cancer Res. 11:6994–7002. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Naksuriya O, Okonogi S, Schiffelers RM and
Hennink WE: Curcumin nanoformulations: A review of pharmaceutical
properties and preclinical studies and clinical data related to
cancer treatment. Biomaterials. 35:3365–3383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Fan Y, Mao R and Yang J: NF-κB and STAT3
signaling pathways collaboratively link inflammation to cancer.
Protein Cell. 4:176–185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Gaykalova DA, Manola JB, Ozawa H, Zizkova
V, Morton K, Bishop JA, Sharma R, Zhang C, Michailidi C, Considine
M, et al: NF-κB and stat3 transcription factor signatures
differentiate HPV-positive and HPV-negative head and neck squamous
cell carcinoma. Int J Cancer. 137:1879–1889. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Khaznadar SS, Khan M, Schmid E, Gebhart S,
Becker ET, Krahn T and von Ahsen O: EGFR overexpression is not
common in patients with head and neck cancer. Cell lines are not
representative for the clinical situation in this indication.
Oncotarget. 9:28965–28975. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Johnson DE, Burtness B, Leemans CR, Lui
VW, Bauman JE and Grandis JR: Head and neck squamous cell
carcinoma. Nat Rev Dis Primers. 6:922020. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Vander Broek R, Mohan S, Eytan DF, Chen Z
and Van Waes C: The PI3K/Akt/mTOR axis in head and neck cancer:
Functions, aberrations, cross-talk, and therapies. Oral Dis.
21:815–825. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Li Z, Yang Z, Passaniti A, Lapidus RG, Liu
X, Cullen KJ and Dan HC: A positive feedback loop involving
EGFR/Akt/mTORC1 and IKK/NF-kB regulates head and neck squamous cell
carcinoma proliferation. Oncotarget. 7:31892–31906. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Harris RE: Cyclooxygenase-2 (cox-2) and
the inflammogenesis of cancer. Subcell Biochem. 42:93–126. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Lim JW, Kim H and Kim KH: Nuclear
factor-kappaB regulates cyclooxygenase-2 expression and cell
proliferation in human gastric cancer cells. Lab Invest.
81:349–360. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Meng F, Liu L, Chin PC and D'Mello SR: Akt
is a downstream target of NF-kappa B. J Biol Chem. 277:29674–29680.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Ahmad A, Biersack B, Li Y, Kong D, Bao B,
Schobert R, Padhye SB and Sarkar FH: Targeted regulation of
PI3K/Akt/mTOR/NF-κB signaling by indole compounds and their
derivatives: Mechanistic details and biological implications for
cancer therapy. Anticancer Agents Med Chem. 13:1002–1013. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Baud V and Karin M: Is NF-κB a good target
for cancer therapy? Hopes and pitfalls. Nat Rev Drug Discov.
8:33–40. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Tili E, Michaille JJ and Croce CM:
MicroRNAs play a central role in molecular dysfunctions linking
inflammation with cancer. Immunol Rev. 253:167–184. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Svoronos AA, Engelman DM and Slack FJ:
OncomiR or tumor suppressor? The duplicity of microRNAs in cancer.
Cancer Res. 76:3666–3670. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Hu A, Huang JJ, Xu WH, Jin XJ, Li JP, Tang
YJ, Huang XF, Cui HJ and Sun GB: MiR-21 and miR-375 microRNAs as
candidate diagnostic biomarkers in squamous cell carcinoma of the
larynx: Association with patient survival. Am J Transl Res.
6:604–613. 2014.PubMed/NCBI
|
|
134
|
Hu A, Huang JJ, Xu WH, Jin XJ, Li JP, Tang
YJ, Huang XF, Cui HJ, Sun GB, Li RL and Duan JL: MiR-21/miR-375
ratio is an independent prognostic factor in patients with
laryngeal squamous cell carcinoma. Am J Cancer Res. 5:1775–1785.
2015.PubMed/NCBI
|
|
135
|
Arantes LM, Laus AC, Melendez ME, de
Carvalho AC, Sorroche BP, De Marchi PR, Evangelista AF,
Scapulatempo-Neto C, de Souza Viana L and Carvalho AL: MiR-21 as
prognostic biomarker in head and neck squamous cell carcinoma
patients undergoing an organ preservation protocol. Oncotarget.
8:9911–9921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Wu Y, Yu J, Ma Y, Wang F and Liu H:
MiR-148a and miR-375 may serve as predictive biomarkers for early
diagnosis of laryngeal carcinoma. Oncol Lett. 12:871–878. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Li J, Wang K, Chen X, Meng H, Song M, Wang
Y, Xu X and Bai Y: Transcriptional activation of microRNA-34a by
NF-kappa B in human esophageal cancer cells. BMC Mol Biol.
13:42012. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Li HP, Zeng XC, Zhang B, Long JT, Zhou B,
Tan GS, Zeng WX, Chen W and Yang JY: MiR-451 inhibits cell
proliferation in human hepatocellular carcinoma through direct
suppression of IKK-β. Carcinogenesis. 34:2443–2451. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Zhou P, Zeng F, Liu J, Lv D and Liu S:
Correlation betweenmir-21 expression and laryngeal carcinoma risks:
A meta-analysis. J Evid Based Med. 9:32–37. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Shen Z, Zhan G, Ye D, Ren Y, Cheng L, Wu Z
and Guo J: MicroRNA-34a affects the occurrence of laryngeal
squamous cell carcinoma by targeting the antiapoptotic gene
survivin. Med Oncol. 29:2473–2480. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Chen Y, Xian PF, Yang L and Wang SX:
MicroRNA-21 promotes proliferation of fibroblast-like synoviocytes
through mediation of NF-κB nuclear translocation in a rat model of
collagen-induced rheumatoid arthritis. Biomed Res Int.
2016:92790782016.PubMed/NCBI
|
|
142
|
Kikkawa N, Kinoshita T, Nohata N, Hanazawa
T, Yamamoto N, Fukumoto I, Chiyomaru T, Enokida H, Nakagawa M,
Okamoto Y and Seki N: MicroRNA-504 inhibits cancer cell
proliferation via targeting CDK6 in hypopharyngeal squamous cell
carcinoma. Int J Oncol. 44:2085–2092. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Kikkawa N, Hanazawa T, Fujimura L, Nohata
N, Suzuki H, Chazono H, Sakurai D, Horiguchi S, Okamoto Y and Seki
N: MiR-489 is a tumour-suppressive miRNA target PTPN11 in
hypopharyngeal squamous cell carcinoma (HSCC). Br J Cancer.
103:877–884. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Fukumoto I, Kinoshita T, Hanazawa T,
Kikkawa N, Chiyomaru T, Enokida H, Yamamoto N, Goto Y, Nishikawa R,
Nakagawa M, et al: Identification of tumour suppressive
microRNA-451a in hypopharyngeal squamous cell carcinoma based on
microRNA expression signature. Br J Cancer. 111:386–394. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Hui AB, Lenarduzzi M, Krushel T, Waldron
L, Pintilie M, Shi W, Perez-Ordonez B, Jurisica I, O'Sullivan B,
Waldron J, et al: Comprehensive microRNA profiling for head and
neck squamous cell carcinomas. Clin Cancer Res. 16:1129–1139. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Tai J, Xiao X, Huang ZG, Yu ZK, Chen XH,
Zhou WG, Chen XJ, Rao YS, Fang JG and Ni X: MicroRNAs regulate
epithelial-mesenchymal transition of supraglottic laryngeal cancer.
Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. 48:499–503. 2013.(In
Chinese). PubMed/NCBI
|
|
147
|
Zhao XD, Zhang W, Liang HJ and Ji WY:
Overexpression of miR-155 promotes proliferation and invasion of
human laryngeal squamous cell carcinoma via targeting SOCS1 and
STAT3. PLoS One. 8:e563952013. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Luzna P, Gregar J, Uberall I, Radova L,
Prochazka V and Ehrmann J Jr: Changes of microRNAs-192, 196a and
203 correlate with Barrett's esophagus diagnosis and its
progression compared to normal healthy individuals. Diagn Pathol.
6:1142011. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Bus P, Siersema PD, Verbeek RE and van
Baal JW: Upregulation of miRNA-143, −145, −192, and −194 in
esophageal epithelial cells upon acidic bile salt stimulation. Dis
Esophagus. 27:591–600. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Chen D, Cabay RJ, Jin Y, Wang A, Lu Y,
Shah-Khan M and Zhou X: MicroRNA deregulations in head and neck
squamous cell carcinomas. J Oral Maxillofac Res. 4:e22013.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Yang Z, Fang S, Di Y, Ying W, Tan Y and Gu
W: Modulation of NF-κB/miR-21/PTEN pathway sensitizes non-small
cell lung cancer to cisplatin. PLoS One. 10:e01215472015.
View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Xiao T, Ling M, Xu H, Luo F, Xue J, Chen
C, Bai J, Zhang Q, Wang Y, Bian Q and Liu Q: NF-κB-regulation of
miR-155, via SOCS1/STAT3, is involved in the
PM2.5-accelerated cell cycle and proliferation of human
bronchial epithelial cells. Toxicol Appl Pharmacol. 377:1146162019.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Rokavec M, Öner MG, Li H, Jackstadt R,
Jiang L, Lodygin D, Kaller M, Horst D, Ziegler PK, Schwitalla S, et
al: IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated
colorectal cancer invasion and metastasis. J Clin Invest.
124:1853–1867. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Tili E, Croce CM and Michaille JJ:
MiR-155: On the crosstalk between inflammation and cancer. Int Rev
Immunol. 28:264–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Bharti AC, Donato N, Singh S and Aggarwal
BB: Curcumin (diferuloylmethane) down-regulates the constitutive
activation of nuclear factor-kappa B and IkappaBalpha kinase in
human multiple myeloma cells, leading to suppression of
proliferation and induction of apoptosis. Blood. 101:1053–1062.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Dai Y, Pei XY, Rahmani M, Conrad DH, Dent
P and Grant S: Interruption of the NF-kappaB pathway by Bay 11-7082
promotes UCN-01-mediated mitochondrial dysfunction and apoptosis in
human multiple myeloma cells. Blood. 103:2761–2770. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Monisha J, Roy NK, Bordoloi D, Kumar A,
Golla R, Kotoky J, Padmavathi G and Kunnumakkara AB: Nuclear factor
kappa B: A potential target to persecute head and neck cancer. Curr
Drug Targets. 18:232–253. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Karin M, Yamamoto Y and Wang QM: The IKK
NF-kappa B system: A treasure trove for drug development. Nat Rev
Drug Discov. 3:17–26. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Ramadass V, Vaiyapuri T and Tergaonkar V:
Small molecule NF-κB pathway inhibitors in clinic. Int J Mol Sci.
21:51642020. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Greten FR, Arkan MC, Bollrath J, Hsu LC,
Goode J, Miething C, Göktuna SI, Neuenhahn M, Fierer J, Paxian S,
et al: NF-kappaB is a negative regulator of IL-1beta secretion as
revealed by genetic and pharmacological inhibition of IKKbeta.
Cell. 130:918–931. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Bromberg JF, Wrzeszczynska MH, Devgan G,
Zhao Y, Pestell RG, Albanese C and Darnell JE Jr: Stat3 as an
oncogene. Cell. 98:295–303. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Geiger JL, Grandis JR and Bauman JE: The
STAT3 pathway as a therapeutic target in head and neck cancer:
Barriers and innovations. Oral Oncol. 56:84–92. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Nelson EA, Walker SR, Kepich A, Gashin LB,
Hideshima T, Ikeda H, Chauhan D, Anderson KC and Frank DA:
Nifuroxazide inhibits survival of multiple myeloma cells by
directly inhibiting STAT3. Blood. 112:5095–5102. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Siddiquee KA, Gunning PT, Glenn M, Katt
WP, Zhang S, Schrock C, Sebti SM, Jove R, Hamilton AD and Turkson
J: An oxazole-based small-molecule Stat3 inhibitor modulates Stat3
stability and processing and induces antitumor cell effects. ACS
Chem Biol. 2:787–798. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Johnson DE, O'Keefe RA and Grandis JR:
Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev
Clin Oncol. 15:234–248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Goldman A, Shahidullah M, Goldman D,
Khailova L, Watts G, Delamere N and Dvorak K: A novel mechanism of
acid and bile acid-induced DNA damage involving Na+/H+ exchanger:
Implication for Barrett's oesophagus. Gut. 59:1606–1616. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Bernstein H, Bernstein C, Payne CM and
Dvorak K: Bile acids as endogenous etiologic agents in
gastrointestinal cancer. World J Gastroenterol. 15:3329–3340. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Lechner S, Müller-Ladner U, Schlottmann K,
Jung B, McClelland M, Rüschoff J, Welsh J, Schölmerich J and
Kullmann F: Bile acids mimic oxidative stress induced upregulation
of thioredoxin reductase in colon cancer cell lines.
Carcinogenesis. 23:1281–1288. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Kinner A, Wu W, Staudt C and Iliakis G:
Gamma-H2AX in recognition and signaling of DNA double-strand breaks
in the context of chromatin. Nucleic Acids Res. 36:5678–5694. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Rothkamm K and Löbrich M: Evidence for a
lack of DNA double-strand break repair in human cells exposed to
very low x-ray doses. Proc Natl Acad Sci USA. 100:5057–5062. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Nikitaki Z, Hellweg CE, Georgakilas AG and
Ravanat JL: Stress-induced DNA damage biomarkers: Applications and
limitations. Front Chem. 3:352015. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Tsantoulis PK, Kotsinas A, Sfikakis PP,
Evangelou K, Sideridou M, Levy B, Mo L, Kittas C, Wu XR,
Papavassiliou AG and Gorgoulis VG: Oncogene-induced replication
stress preferentially targets common fragile sites in preneoplastic
lesions. A genome-wide study. Oncogene. 27:3256–3264. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Choudhari SK, Chaudhary M, Gadbail AR,
Sharma A and Tekade S: Oxidative and antioxidative mechanisms in
oral cancer and precancer: A review. Oral Oncol. 50:10–18. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Janssens S and Tschopp J: Signals from
within: The DNA-damage-induced NF-kappaB response. Cell Death
Differ. 13:773–784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Kawanishi S, Ohnishi S, Ma N, Hiraku Y and
Murata M: Crosstalk between DNA damage and inflammation in the
multiple steps of carcinogenesis. Int J Mol Sci. 18:18082017.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Tafani M, Sansone L, Limana F, Arcangeli
T, De Santis E, Polese M, Fini M and Russo MA: The interplay of
reactive oxygen species, hypoxia, inflammation, and sirtuins in
cancer initiation and progression. Oxid Med Cell Longev.
2016:39071472016. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Li D and Cao W: Bile acid receptor TGR5,
NADPH Oxidase NOX5-S and CREB mediate bile acid-induced DNA damage
in Barrett's esophageal adenocarcinoma cells. Sci Rep. 6:315382016.
View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Nema R, Vishwakarma S, Agarwal R, Panday
RK and Kumar A: Emerging role of sphingosine-1-phosphate signaling
in head and neck squamous cell carcinoma. Onco Targets Ther.
9:3269–3280. 2016.PubMed/NCBI
|
|
179
|
Tamashiro PM, Furuya H, Shimizu Y, Iino K
and Kawamori T: The impact of sphingosine kinase-1 in head and neck
cancer. Biomolecules. 3:481–513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Liu R, Li X, Hylemon PB and Zhou H:
Conjugated bile acids promote invasive growth of esophageal
adenocarcinoma cells and cancer stem cell expansion via sphingosine
1-phosphate receptor 2-mediated yes-associated protein activation.
Am J Pathol. 188:2042–2058. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Pang C, LaLonde A, Godfrey TE, Que J, Sun
J, Wu TT and Zhou Z: Bile salt receptor TGR5 is highly expressed in
esophageal adenocarcinoma and precancerous lesions with
significantly worse overall survival and gender differences. Clin
Exp Gastroenterol. 10:29–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Guan B, Li H, Yang Z, Hoque A and Xu X:
Inhibition of farnesoid X receptor controls esophageal cancer cell
growth in vitro and in nude mouse xenografts. Cancer.
119:1321–1329. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Yu JH, Zheng JB, Qi J, Yang K, Wu YH, Wang
K, Wang CB and Sun XJ: Bile acids promote gastric intestinal
metaplasia by upregulating CDX2 and MUC2 expression via the
FXR/NF-κB signaling pathway. Int J Oncol. 54:879–892.
2019.PubMed/NCBI
|
|
184
|
Schweitzer A, Knauer SK and Stauber RH:
Nuclear receptors in head and neck cancer: Current knowledge and
perspectives. Int J Cancer. 126:801–809. 2010.PubMed/NCBI
|
|
185
|
Gadaleta RM, Oldenburg B, Willemsen EC,
Spit M, Murzilli S, Salvatore L, Klomp LW, Siersema PD, van Erpecum
KJ and van Mil SW: Activation of bile salt nuclear receptor FXR is
repressed by pro-inflammatory cytokines activating NF-κB signaling
in the intestine. Biochim Biophys Acta. 1812:851–858. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Wang YD, Chen WD, Wang M, Yu D, Forman BM
and Huang W: Farnesoid X receptor antagonizes nuclear factor kappaB
in hepatic inflammatory response. Hepatology. 48:1632–1643. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Wang Z, Li Y and Sarkar FH: Signaling
mechanism(s) of reactive oxygen species in epithelial-mesenchymal
transition reminiscent of cancer stem cells in tumor progression.
Curr Stem Cell Res Ther. 5:74–80. 2010. View Article : Google Scholar : PubMed/NCBI
|