Introduction
Wnt signaling pathway activation is a hallmark of
cancer stem cells. Tumor cells can lose the ability to form tumors
by modulating Wnt signaling activity. Abnormal activation of the
Wnt pathway is associated with a variety of human tumors, such as
breast cancer (1), liver cancer
(2), lung cancer (3), and promotes tumor cell proliferation
and migration.
Colorectal cancer (CRC) is one of the most common
tumors found in the digestive system (4). Continuous activation of the Wnt
signaling pathway is characteristic of colorectal tumors (5). As an extracellular antagonist of the
Wnt pathway, the secreted frizzled related proteins (SFRPs) can
competitively inhibit signaling and prevent overactivation of the
Wnt pathway (6). However, our recent
meta-analysis results indicated that hypermethylation of the SFRP
promoter is inversely related to its expression in most tumors and
contributes to canceration, at least in CRC (7). Furthermore, our previous study found
that SFRP family gene promoter methylation occurred in the early
stage of colorectal tumorigenesis, and maintained high frequency
methylation in adenomas and adenocarcinomas (8). The methylation rate of SFRP1, SFRP2,
SFRP4, and SFRP5 were found to be 93.1, 83.3, 36.1, and 52.8% in
colorectal carcinoma (8). Removal of
SFRP gene promoter methylation was found to effectively restore
gene expression, and to inhibit the abnormal activation of the Wnt
signaling pathway, indicating that SFRP gene silencing is
associated with hypermethylation of promoter region DNA, which may
be the cause of the development of CRC (8,9).
Two classes of proteins have been implicated in the
interaction between DNA methylation and histone modifications,
possibly related to the formation of tumor-specific epigenomes, the
polycomb group (PcG) protein family and the methyl-CpG-binding
domain (MBD) protein family. It is known that PcG proteins do not
function alone, but are assembled into polycomb repressive
complexes 1 and 2, which play important roles in embryonic
development, stem cell self-renewal, and cell proliferation through
epigenetic modification of target genes (10–12). As
an effector component of PRC2, enhancer of zeste homolog 2 (EZH2)
can recruit DNA-methyltransferase (DNMT)1, DNMT3A, and DNMT3B to
the promoter region of the target gene, and bind to the gene
promoter to maintain gene promoter methylation (13,14). It
has been reported that EZH2 can promote cell proliferation,
invasion and metastasis, and can also endanger DNA damage and
repair (15). At the same time,
experimental results have shown that tumor cell lines depend on
polycomb repressive complex 2 (PRC2) activity, for example,
knockout of EZH2 or other PRC2 core components, and the use of
small-molecular substances to inhibit EZH2, can reduce the
proliferation of cell lines derived from various types of cancer
(16). MBD2 plays a transcriptional
inhibitory role in tumors mainly by recruiting histone deacetylase
complexes NuRD/Mi-2 and Sin3A to form transcriptional repressors
(17,18), such as the silencing of the KAI1 gene
in prostate cancer PC3 cells (19).
We suspect that silencing of MBD2 and EZH2 can alter the
methylation status of the SFRP gene promoter and then affect its
expression to inhibit the WNT signaling pathway, which may be
another target for colorectal tumor treatment.
In the present study, our aim was to explore the
potential role of MBD2 and EZH2 proteins in CRC and their effects
on the expression of SFRP.
Materials and methods
Analysis of GEPIA and METCH
databases
GEPIA (Gene Expression Profiling Interactive
Analysis) is an online application developed by Peking University
that can be used to analyze the differential expression of genes in
cancer and normal tissues (http://gepia.cancer-pku.cn/) (20). GEPIA is a web-based tool for
analyzing normal and tumor sample RNA sequencing data based on The
Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GETx)
data (tumor samples from TCGA dataset, normal samples from two
datasets). In the present study, we observed the mRNA expression
level of SFRP genes in the GEPIA database with the settings P≤0.01
and |log2(FC)| ≥1. Under this condition, we selected 275 tumor
samples from the TCGA database and 349 normal samples from two
databases in COAD and 92 tumor samples from TCGA database and 318
normal samples from two databases in READ to analyze the expression
of SFRP in CRC. The relationship between SFRP gene expression and
promoter methylation was analyzed by the METCH database (http://methhc.mbc.nctu.edu.tw/php/index.php) (21) [Tumor=275, Normal=349 (41 from TCGA
database); READ Tumor=92, Normal=318 (10 from TCGA database)].
Cell cultures
Normal colon mucosa cell line (NCM460) and human
colorectal cancer cell lines (HCT116, SW480, HT29 and DLD1) were
donated from the team of Zhao Qiu, Department of Gastroenterology,
Zhongnan Hospital of Wuhan University (Wuhan, China). All cells
were cultured in a high glucose medium (DMEM, Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS, Hangzhou
Sijiqing Biological Engineering Materials Co., Ltd.) in a
humidified atmosphere at 37°C and 5% CO2.
RNA interference and transfection
CRC cells were seeded in a 6-well plate (50,000
cells/ml). Transfection was initiated when the cell density reached
50–60%. Cells were transfected with siRNA using a transfection
reagent (GenMute™ siRNA, cat no. SL100568; GenePharma) according to
the manufacturer's protocol. Approximately 24 h later, the cells
were harvested for analysis as described below. The small
interfering RNA (siRNA) sequences targeting human MBD2, EZH2, and
negative control (NC) were purchased from Guangzhou RiboBio Co.,
Ltd. The siRNA sequences used in the study are listed as follows:
MBD2-001 forward, 5′-GAGGCUACAAGGACUUAGUTT−3′ and reverse,
5′-ACUAAGUCCUUGUAGCCUCTT−3′; MBD2-002 forward,
5′-GAUGAUGCCUAGUAAAUUATT−3′ and reverse,
5′-UAAUUUACUAGGCAUCAUCTT−3′; MBD2-003 forward,
5′-CCUGGGAAAUACUGUUGAUTT−3′ and reverse,
5′-AUCAACAGUAUUUCCCAGGTT−3′; EZH2-2196 forward,
5′-GCAGCUUUCUGUUCAACUUTT−3′ and reverse,
5′-AAGUUGAACAGAAAGCUGCTT−3′; EZH2-488 forward,
5′-GACUCUGAAUGCAGUUGCUTT−3′ and reverse,
5′-AGCAACUGCAUUCAGAGUCTT−3′; EZH2-1952 forward,
5′-CCUGACCUCUGUCUUACUUTT−3′ and reverse,
5′-AAGUAAGACAGAGGUCAGGTT−3′. A nonspecific control siRNA was used
as a control: NC forward, 5′-UUCUCCGAACGUGUCACGUTT−3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT−3′.
RNA isolation and quantitative
real-time PCR (qPCR)
CRC cells were placed in an EP tube containing 1 ml
of TRIzol reagent (Thermo Fisher Scientific, Inc.). Total RNA
extraction was performed according to the manufacturer's
instructions. The resulting RNA was dissolved in RNase-free water
and immediately stored at −80°C. The RNA concentration was measured
using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific,
Inc.). cDNA was then synthesized using a ReverTra Ace qPCR RT kit
(Toyobo Life Science). qPCR was performed using SYBR-Green PCR
master mix in a Biorad 7500 real-time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The qPCR primers for
MBD2, EZH2, and GAPDH were purchased from QingKe. The specific
primers used are as follows: MBD2 forward,
5′-GCAAGCCTCAGTTGGCAAG-3′ and reverse, 5′-ATCGTTTCGCAGTCTCTGTTT−3′;
EZH2 forward, 5′-TCCTACATCCTTTTCATGCAACAC−3′ and reverse,
5′-GCTCCCTCCAAATGCTGGTA-3′; SFRP1 forward,
5′-TGGCCCGAGATGCTTAAGTG-3′ and reverse, 5′-GACACACCGTTGTGCCTTG-3′;
SFRP2 forward, 5′-CGACATGCTTGAGTGCGAC-3′ and reverse,
5′-CTTTGGAGCTTCCTCGGTGG-3′; SFRP4 forward,
5′-TCACCCATCCCTCGAACTCA-3′ and reverse, 5′-CATCATCCTTGAGCGCCACT-3′;
SFRP5 forward, 5′-CCTCCAGTGACCAAGATCTGC−3′ and reverse,
5′-TCCTTGATGCGCATTTTGACC−3′; GAPDH forward,
5′-AGAAGGCTGGGGCTCATTTG-3′ and reverse,
5′-GCAGGAGGCATTGCTGATGAT−3′. GAPDH was used as a control. All
reactions were run in triplicate, and the results were analyzed and
expressed relative to threshold cycle (CT) values and then
converted to fold change values (2−ΔΔCq) (22).
Western blot analysis
Cells were lysed with RIPA protein extraction
reagent (Beyotime Institute of Biotechnology) supplemented with 1%
phenylmethanesulfonyl fluoride (Seebio Science & Technology
Co., Ltd.). Protein concentrations were measured using an enhanced
BCA protein assay kit (Beyotime Institute of Biotechnology). Each
lane contained equal amounts of protein (30 µg). Proteins were
separated using 14% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and were then transferred to a
polyvinylidene fluoride membrane (Millipore, USA). The membranes
were blocked with 5% milk for 2 h at room temperature. The
membranes were then placed in a TBST solution containing anti-SFRP1
antibody (dilution 1:500; cat. no. AB_2764730; ABclonal
Biotechnology Co., Ltd.), SFRP2 (dilution 1:1,000; cat. no
AB_2766193; ABclonal Biotechnology Co., Ltd.), SFRP4 (dilution
1:1,000; cat. no. AB_2767011; ABclonal Biotechnology Co., Ltd.),
SFRP5 (dilution 1:1,000; cat. no. AB_2772201; ABclonal
Biotechnology Co., Ltd.), MBD2 (dilution 1:500; cat. no. ab38646;
Abcam), and EZH2 antibodies (dilution 1:1,000; cat. no. ab186006;
Abcam) overnight (~12 h) at 4°C. The following day, after washing
with TBST, the blots were incubated with an HRP-labelled
anti-rabbit (dilution 1:5,000, cat. no. GB233303-1, Servicebio)
secondary antibody for 2 h at room temperature. The blots were
visualized using a super ECL detection reagent (Solarbio, Beijing,
China). ImageJ 1.8.0 (NIH) was used for densitometry. Each set was
repeated at least three times.
Wound-healing assay
CRC cells were seeded in a 6-well plate (50,000
cells/ml). Twenty-four hours post-transfection, the cells had
reached 70–80% confluency. A straight scratch was created using a
sterile yellow pipette tip. PBS was added to the 6-well plate to
remove floating cells. The original culture medium was replaced
with 0.5% FBS for another 24 h as previously described (23). All tests were repeated three times.
Cell migration was observed and imaged at 0, 24, and 36 h with a
digital camera (×40 magnification; OLYMPUS U-RFL-T; Olympus
Corp.).
Cell proliferation assay
At 24 h post-transfection, SW480 and HCT116 cells
were trypsinized to provide suspensions that were then seeded in
96-well plates at densities of 2,000 cells/well. The cell
proliferation rates were calculated using the CCK-8 assay at 0, 24,
48, and 72 h. Briefly, 10 µl of CCK-8 was added to each well, and
the cells were allowed to incubate at 37°C in 5% CO2 for
2 h. The absorbance value of each well at 570 nm was recorded. Each
experiment was repeated at least three times.
Cell invasion assay
A 24-multiwell insert plate with a small chamber (BD
Biosciences) containing an 8.0-micron pore size Matrigel-coated
membrane was used for the cell invasion assays. Briefly, Matrigel
liquid was added to the chamber the day before the experiment and
placed in a 4°C refrigerator overnight to solidify. On the next
day, the cells of each group that had been transfected in a 6-well
plate were digested and centrifuged. A total of 1×105
cells were seeded into the upper chambers (coated in Matrigel) in
serum-free medium. The 24-well plates were filled with 600 µl of
DMEM containing 20% FBS as a chemo-attractant. After the plates
were incubated at 37°C for 24 h, the non-invasive cells above the
chamber were gently wiped with a wet cotton swab. The cells below
the chamber were fixed with 4% paraformaldehyde for 30 min, stained
with 0.1% crystal violet for 20 min, and then counted under an
optical microscope (40× magnification). Each set was repeated at
least three times.
Analysis of cell apoptosis and cell
cycle distribution
The cells were seeded in 6-well plates at
5×105 cells/well. After transfection with siRNA for 24
h, SW480 and HCT116 cells were harvested. Cycle and apoptosis kits
(MultiSciences, China) were used to detect apoptosis and cell cycle
distribution of the SW480 and HCT116 cells according to the
manufacturers' instructions. The cell cycle distribution and
apoptosis rate were analyzed using a flow cytometer (Beckman
CytoFLEX FCM). Specific methods were carried out according to our
previous experiments (24).
Methylation specific PCR (MSP)
DNA was extracted using TIANamp Genomic DNA Kit
(Tiangen Biotech Co.). Sodium bisulfite treatment was performed
using an EZ DNA Methylation Kit (Zymo Research). The regions of the
methylated SFRP1, SFRP2, SFRP4, and SFRP5 promoters by MSP analysis
were chromosome 8 (41309333-41309459, length: 126 bp), chromosome 4
(153788916-153789054, length: 138 bp), chromosome 7
(37916853-37916965, length: 112 bp) and chromosome 10
(97772023-97772159, length: 136 bp), respectively. The sequence
information of the SFRP primers used in the MSP experiments are
shown in Table I.
 | Table I.SFRP primer sequence information for
MSP experiments. |
Table I.
SFRP primer sequence information for
MSP experiments.
| Gene name | Primer
sequence |
|---|
|
SFRP1(U) | F:
5′-GTTTTGTAGTTTTTGGAGTTAGTGTTGTGT-3′ |
|
| R:
5′-CCTACGATCGAAAACGACGCGAACG-3′ |
|
SFRP2(U) | F:
5′-TTTTGGGTTGGAGTTTTTTGGAGTTGTGT-3′ |
|
| R:
5′-AACCCACTCTCTTCACTAAATACAACTCA-3′ |
|
SFRP4(U) | F:
5′-GGGGGTGATGTTATTGTTTTTGTATTGAT-3′ |
|
| R:
5′-CACCTCCCCTAACATAAACTCAAAACA-3′ |
|
SFRP5(U) | F:
5′-GTAAGATTTGGTGTTGGGTGGGATGTTT-3′ |
|
| R:
5′-AAAACTCCAACCCAAACCTCACCATACA-3′ |
|
SFRP1(M) | F:
5′-TGTAGTTTTCGGAGTTAGTGTCGCGC-3′ |
|
| R:
5′-CCTACGATCGAAAACGACGCGAACG-3′ |
|
SFRP2(M) | F:
5′-GGGTCGGAGTTTTTCGGAGTTGCGC-3′ |
|
| R:
5′-CCGCTCTCTTCGCTAAATACGACTCG-3′ |
|
SFRP4(M) | F:
5′-GGGTGATGTTATCGTTTTTGTATCGAC-3′ |
|
| R:
5′-CCTCCCCTAACGTAAACTCGAAACG-3′ |
|
SFRP5(M) | F:
5′-AAGATTTGGCGTTGGGCGGGACGTTC-3′ |
|
| R:
5′-ACTCCAACCCGAACCTCGCCGTACG-3′ |
Statistical analysis
The correlation between SFRP expression and its
methylation were confirmed by Pearson correlation coefficients. The
data presented in the text and figures were analyzed using one-way
analysis of variance (ANOVA) with a Bonferroni correction. Data are
presented as the mean standard ± deviation. Statistical analyses
were performed using SPSS 22.0 (IBM Corp). A P-value of <0.05
was considered as indicative of statistical significance.
Results
Expression of SFRP, MBD2, and EZH2 in
colorectal tumor tissues and cells
The TCGA database (including 275 colorectal tumor
tissues and 349 normal tissues) demonstrated that the expression of
SFRP1, SFRP2, and SFRP5 genes were lower in CRC tissues compared
(COAD, colon adenocarcinoma; READ, rectum adenocarcinoma) to that
in the corresponding paratumorous normal tissue (Fig. 1A, B and D), whereas EZH2 and SFRP4
expression levels were higher in the tumor tissue than levels in
the matching normal colorectal tissues (Fig. 1C and E). Furthermore, the expression
of SFRP1, SFRP2, SFRP4, and SFRP5 genes were all lower in the CRC
cell lines than that noted in the normal colorectal mucosal cells
(NCM460) (Fig. 1G). However, the
expression levels of MBD2 and EZH2 were much higher in the CRC
cells rather than in normal colorectal mucosal epithelial cells
(NCM460) (Fig. 1F).
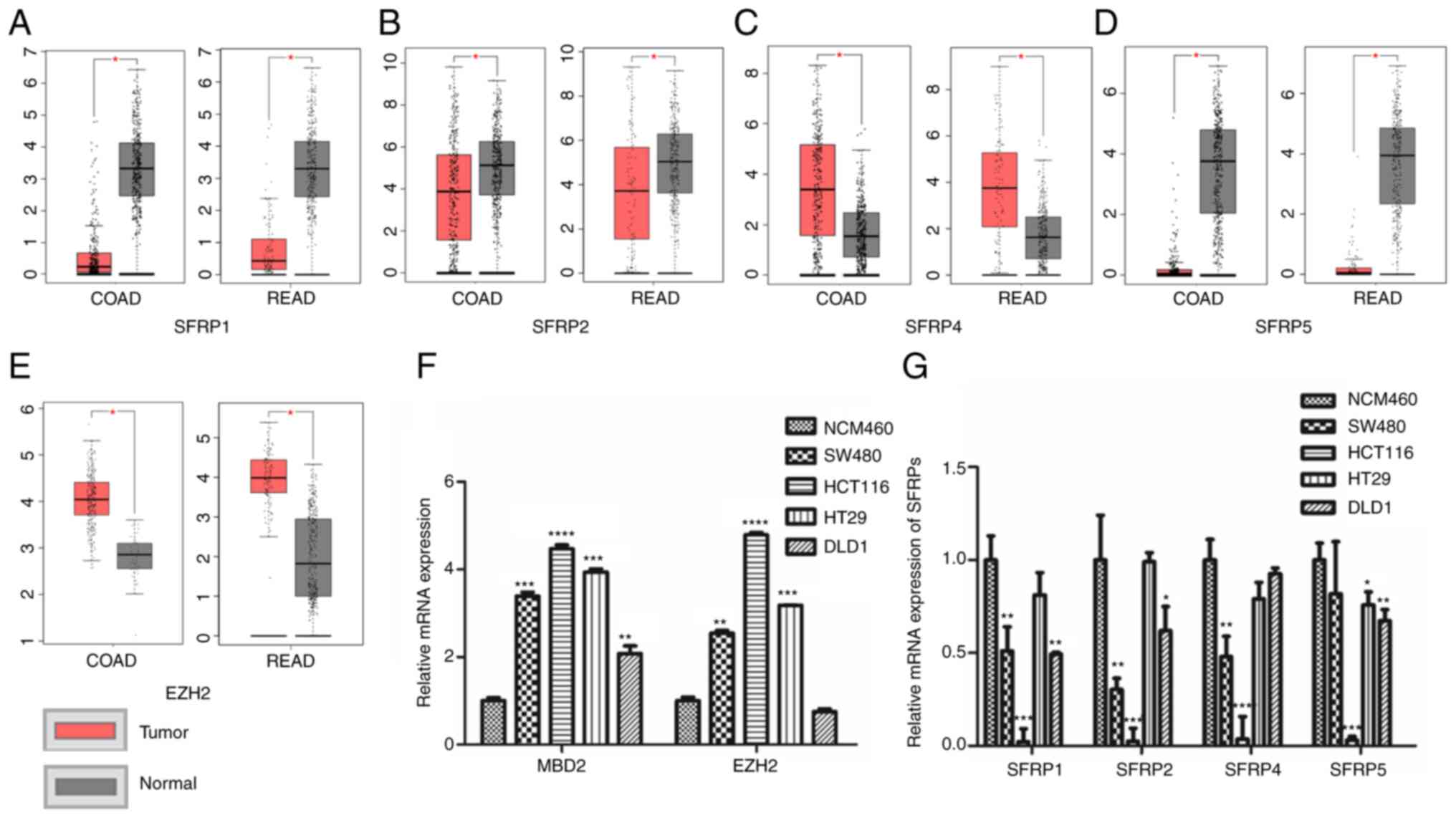 | Figure 1.Expression levels of SFRPs, MBD2 and
EZH2 in colorectal tumor tissues and cells. The expression of
SFRP1, (A) SFRP2, (B) SFRP4, (C) SFRP5 (D) and EZH2 (E) in TCGA and
GTEx database [COAD: Tumor=275, Normal=349 (41 from TCGA database);
READ Tumor=92, Normal=318 (10 from TCGA database)]. The mRNA
expression of MBD2, EZH2 (F) and SFRPs (G) in normal colorectal
epithelial cells (NCM460) and colorectal tumor cell lines (SW480,
HCT116, HT29 and DLD1) was detected by qPCR. COAD, colon
adenocarcinoma; READ, rectum adenocarcinoma; COAD num(T)=275;
num(N)=349; READ num(T)=92; num(N)=318. *P<0.05, **P<0.01,
***P<0.001, ****P<0.0001 vs. normal colonic mucosal tissues
or normal colon mucosa cell line (NCM460). SFRPs, secreted frizzled
related proteins; MBD2, methyl-CpG binding domain protein 2; EZH2,
enhancer of zeste homolog 2. |
Correlation between SFRP gene
expression and promoter methylation in colorectal tumors
MethHC online database confirmed the greater degree
of SFRP1, SFRP2, SFRP4, and SFRP5 promoter methylation in tumor
tissue compared with that noted in the paired normal samples
(Fig. 2A-D). In colorectal tumors,
the expression of SFRP1 and SFRP2 genes were inversely proportional
to the degree of promoter methylation (Fig. 2E and F, P<0.05). However, the
expression of SFRP4 and SFRP5 genes exhibited no obvious
relationship with the degree of promoter methylation (Fig. 2G and H, P>0.05).
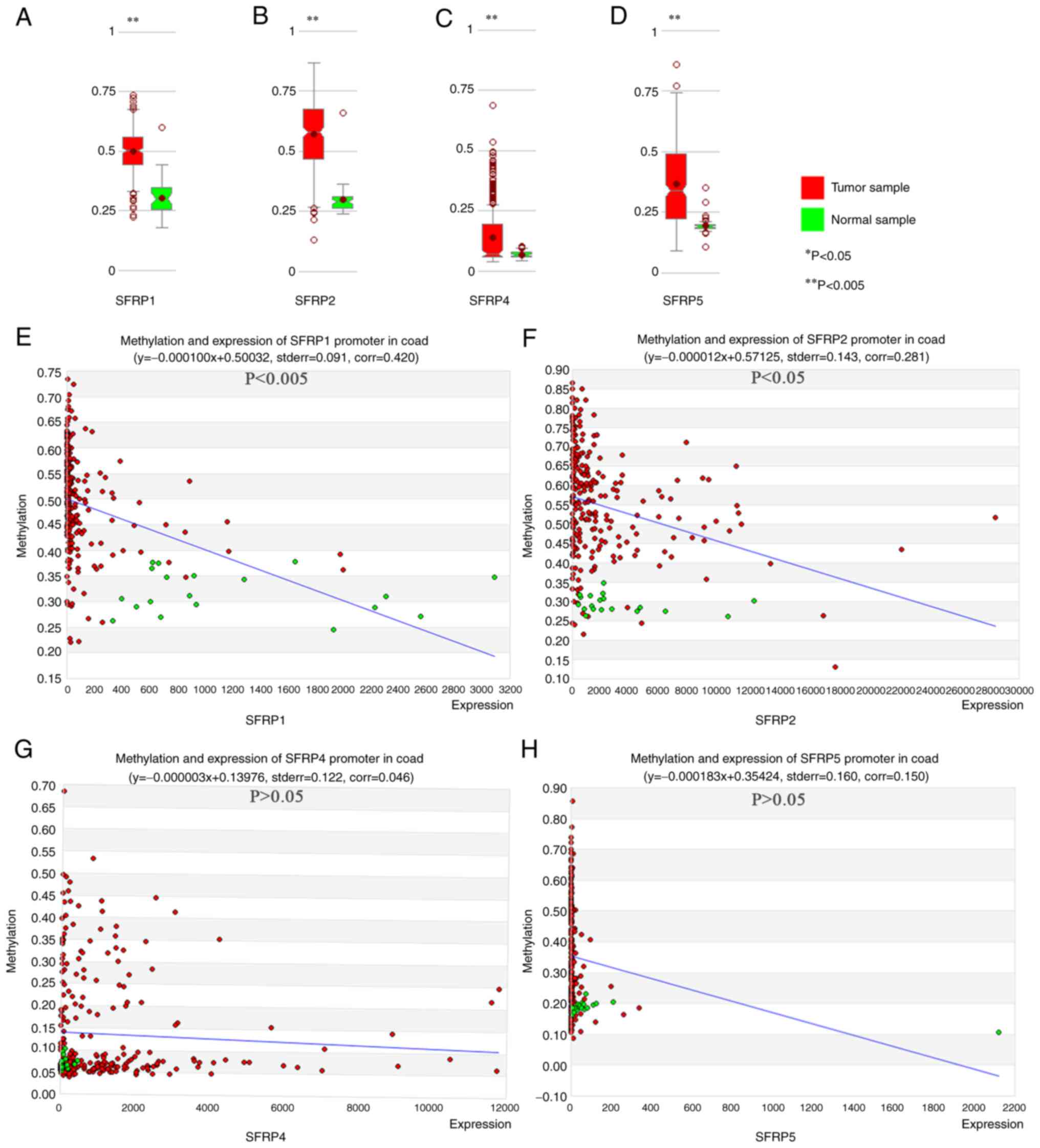 | Figure 2.MethHC online database shows the
degree of DNA methylation of SFRP promoter and its relationship
with gene expression in colorectal tumor and normal tissues. The
degree of methylation of SFRP1, (A) SFRP2 (B), SFRP4 (C) and SFRP5
(D) promoters in colorectal tumor and normal tissues. Correlations
between methylation of SFRP1 (E, corr=0.420, P<0.005), SFRP2 (F,
corr=0.281, P<0.05), SFRP4 (G, corr=0.046, P>0.05) and SFRP5
(H, corr=0.150, P>0.05) promoters and gene expression in
colorectal tumor and normal tissues. **P<0.005 vs. normal
sample. SFRP, secreted frizzled related protein. |
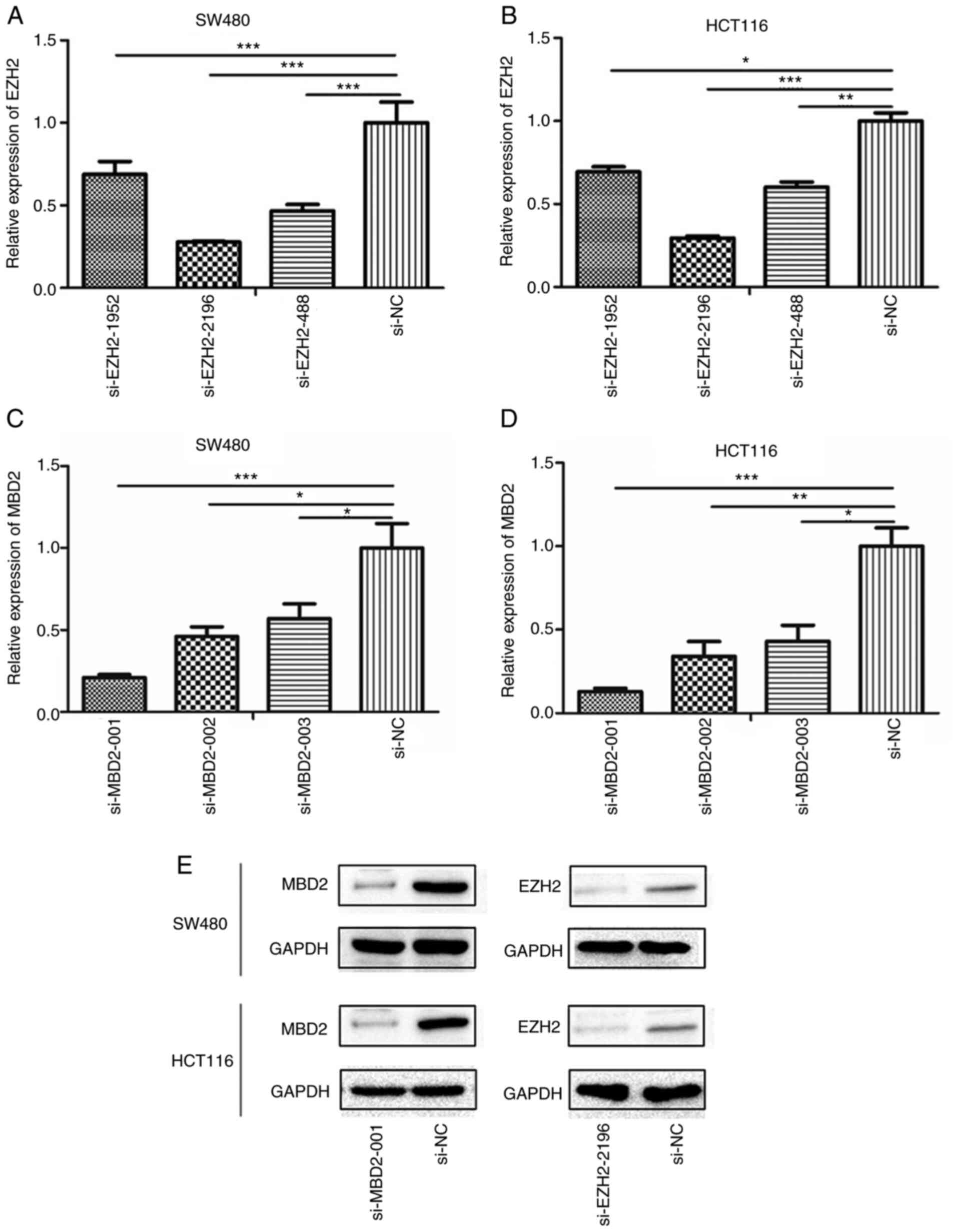
Determination of the transfection
efficiency of si-MBD2 and si-EZH2
Twenty-four hours after transfection, the
interference efficiency of siRNA-EZH2-1952, siRNA-EZH2-2196,
siRNA-EZH2-488 and siRNA-MBD2-001, siRNA-MBD2-002, siRNA-MBD2-003
was detected by qPCR. The results showed that siRNA-EZH2-2196 and
siRNA-MBD2-001 performed the best in both SW480 and HCT116 cell
lines (Fig. 3A-D). Moreover, it was
further verified that siRNA-EZH2-2196 and siRNA-MBD2-001 had the
best interference efficiency at the protein level (Fig. 3E). Therefore, we selected these
siRNAs for subsequent experiments.
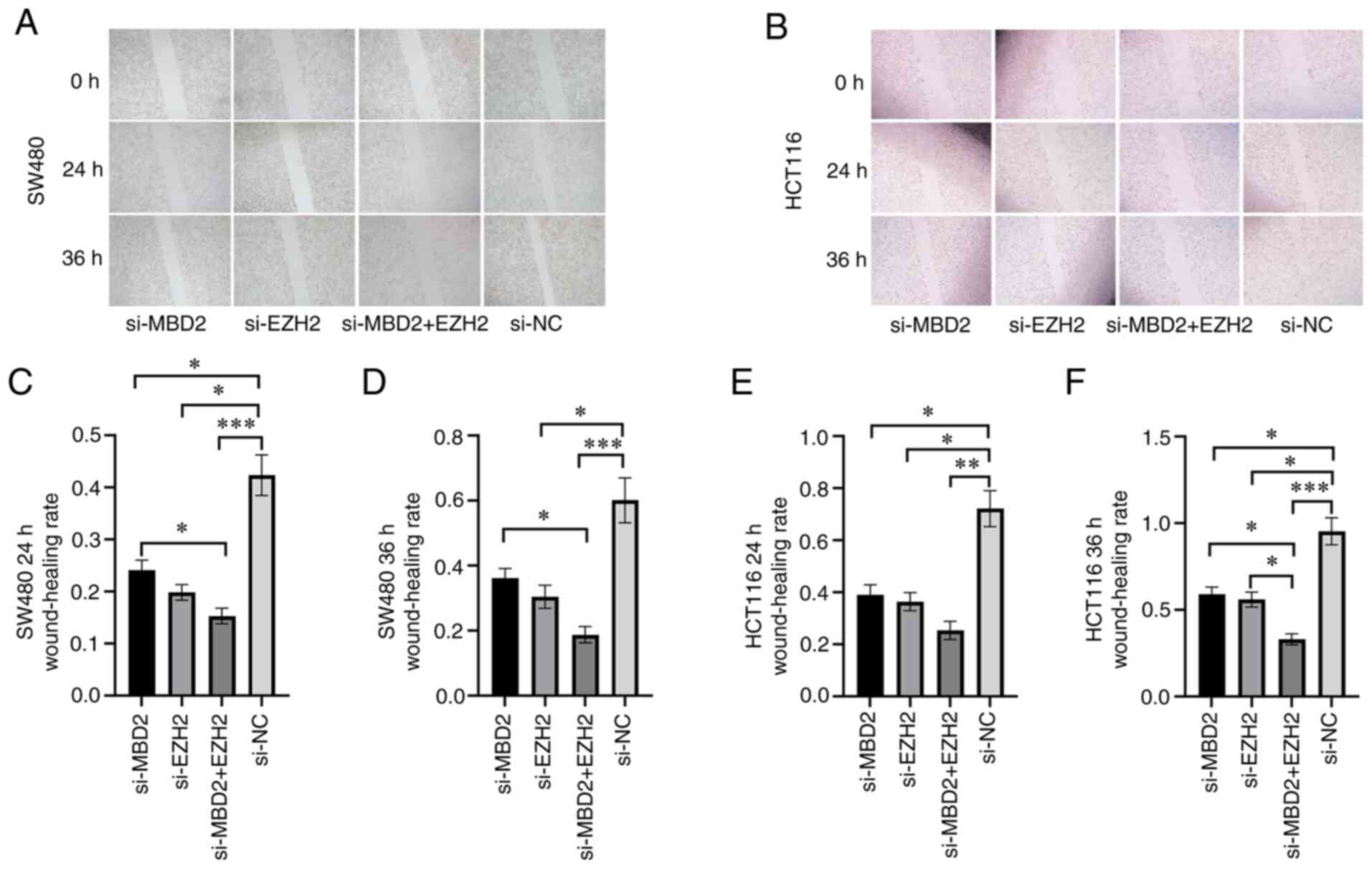
Effects of MBD2 and EZH2 knockdown on
CRC cell migration
In order to detect the effect of MBD2 and EZH2 on
the migratory ability of CRC cells, a scratch wound assay was used
to detect the wound healing of SW480 and HCT116 cells in which MBD2
and EZH2 knockdown was conducted separately or simultaneously.
Following 24 h of wounding in the SW480 cells, knockdown of either
MBD2 or EZH2 significantly inhibited cell migration when compared
with the negative control (si-NC) group (Fig. 4A and C). Furthermore, knockdown of
both MBD2 and EZH2 simultaneously inhibited cell migration more
effectively, which was statistically significant compared with the
knockdown of MBD2 and EZH2, respectively (Fig. 4A and C). Compared to the si-NC group,
knockdown of EZH2 or MBD2 decreased cell migration following 36 h
after wounding in the SW480 cells (Fig.
4A and D); while knockdown of MBD2 and EZH2 together
significantly inhibited cell migration, which was statistically
significant compared with the knockdown of MBD2 alone (Fig. 4A and D). The migration of HCT116
cells following the knockdown of MBD2 and EZH2 separately or
simultaneously was also decreased compared with the si-NC group 24
h after wounding (Fig. 4B and E).
Following 36 h after wounding in HCT116 cells, compared with the
si-NC group, knockdown of MBD2 and EZH2 separately or
simultaneously inhibited cell migration; however, the group with
simultaneous knockdown of MBD2 and EZH2 exhibited inhibition of
cell migration to a greater degree (Fig.
4B and F). Therefore, wound healing assays showed that
knockdown of MBD2 and EZH2 inhibited the migration of SW480 and
HCT116 cells compared to the control group, and depletion of MBD2
and EZH2 plays a greater role in inhibiting the migration of CRC
cells.
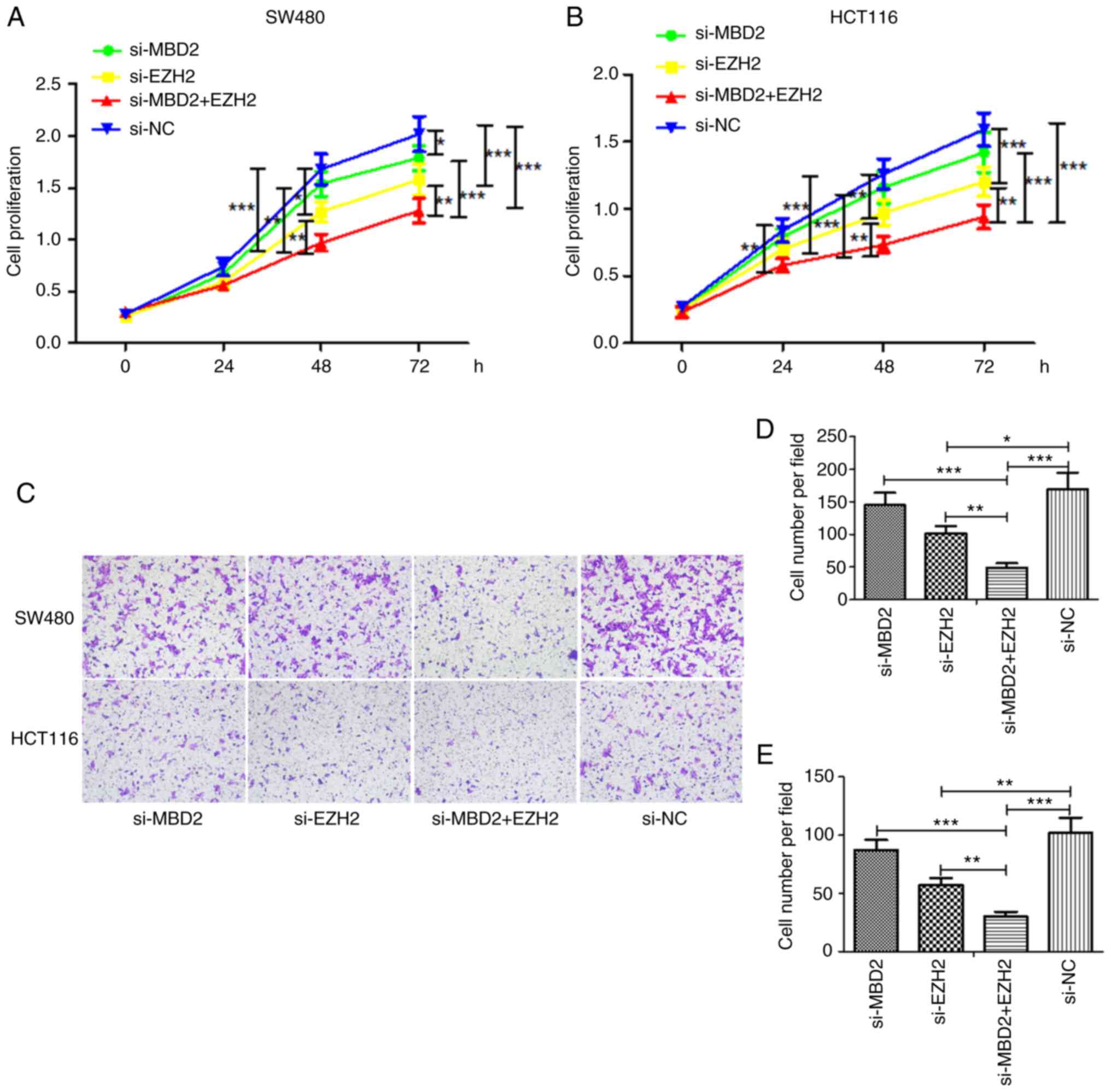
Effects of MBD2 and EZH2 on the
proliferation and invasion of CRC cell lines
CCK-8 assay was used to detect the effects of MBD2
and EZH2 on the proliferation of CRC cells. In the SW480 cell line
(Fig. 5A), the cell proliferation in
the EZH2-knockdown group was slower than that noted in the si-NC
group at 48 h; yet, knockdown of MBD2 had little effect on cell
proliferation. However, knockdown of both MBD2 and EZH2
significantly inhibited the proliferation of cells, which was
significantly different following knockdown of MBD2 or EZH2 alone.
At 72 h, the proliferation of the cells was inhibited by knockdown
of MBD2 or EZH2, and also was significantly inhibited by
interference with both MBD2 and EZH2, which was significantly
different following knockdown of MBD2 or EZH2 alone.
In HCT116 cells (Fig.
5B), knockdown of MBD2 or EZH2 at 24 h was statistically
significant at inhibiting proliferation compared with the si-NC
group, and knockdown of EZH2 at 48 and 72 h inhibited the growth of
CRC cells, while knockdown of MBD2 resulted in basically the same
result as in the si-NC group. Furthermore, knockdown of MBD2 and
EZH2 together significantly inhibited cell proliferation, which was
significantly different following knockdown of MBD2 or EZH2
alone.
The impact of MBD2 and EZH2 on CRC cell invasiveness
was investigated using Matrigel invasion chambers. As shown in
Fig. 5C-E, the same results were
obtained in both SW480 and HCT116 cells. That is, compared with the
si-NC group, knockdown of EZH2 weakened the invasiveness of CRC
cells, while knockdown of MBD2 had no significant effect on the
invasiveness of CRC cells. Yet, knockdown of MBD2 and EZH2
simultaneously inhibited the invasiveness of the CRC cells, which
was significantly different following the knockdown of MBD2 or EZH2
alone. The results were consistent with the results of the cell
proliferation.
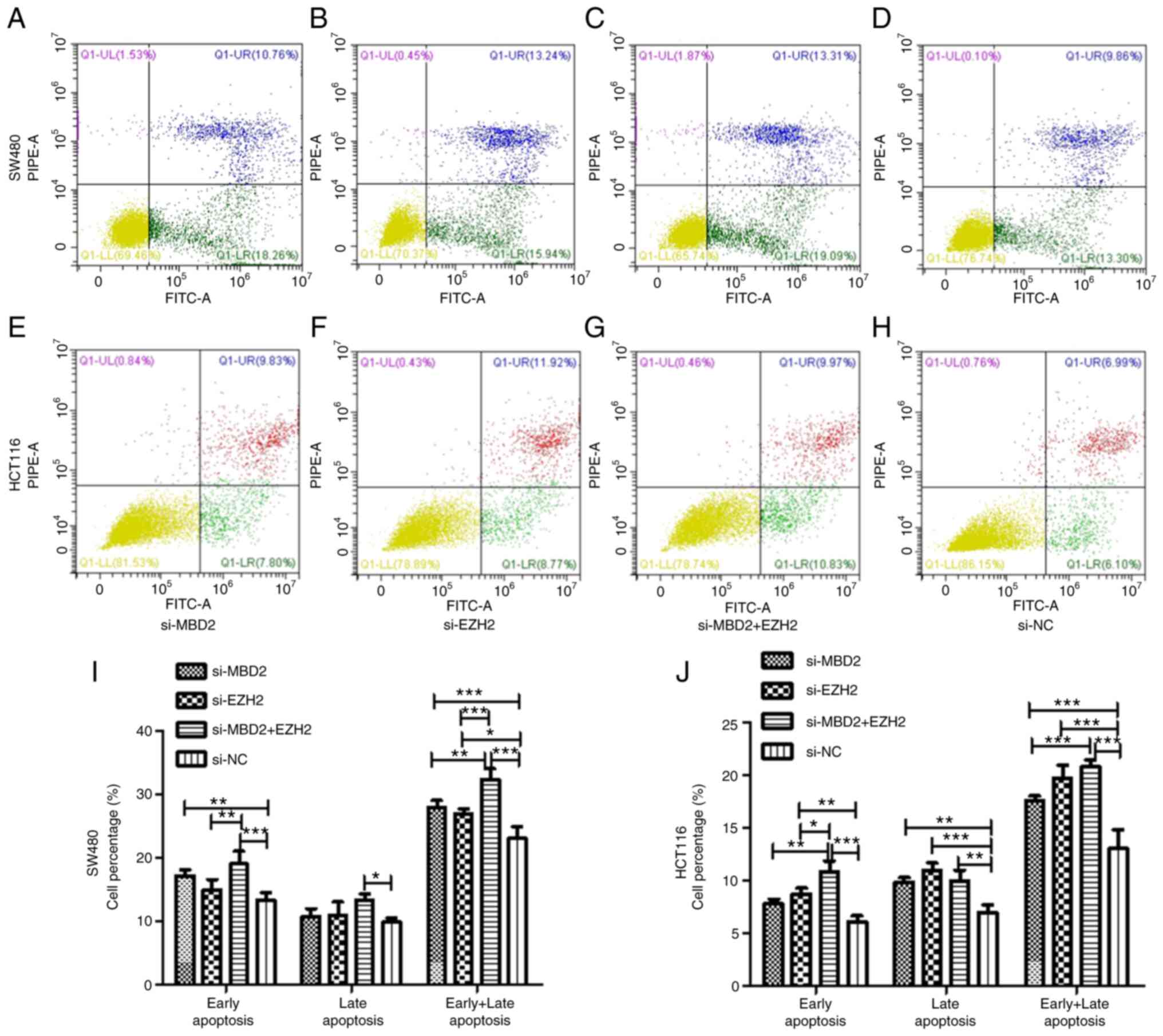
Effects of MBD2 and EZH2 silencing on
the apoptosis of CRC cells
To verify the effect of MBD2 and EZH2 on cell
apoptosis, we knocked down MBD2 and EZH2 separately or
simultaneously in SW480 and HCT116 cells and then examined
apoptosis via flow cytometry.
As is shown in Fig.
6, in SW480 cells, knockdown of MBD2 was able to increase the
percentage of early apoptosis of the cells compared with the si-NC
group, but there was no significant difference between knockdown of
EZH2 and the si-NC group. Simultaneously, knockdown of MBD2 and
EZH2 together increased the percentage of early apoptosis of the
SW480 cells more significantly. Compared with the si-NC group,
knockdown of MBD2 and EZH2 together also increased the late
apoptosis of CRC cells, but knockdown of MBD2 or EZH2 did not
affect late apoptosis. Knockdown of MBD2 and EZH2 separately and
simultaneously increased the percentage of total apoptosis of CRC
cells, and the percentage of total apoptosis of CRC cells by
knockdown of MBD2 and EZH2 together was significantly higher
following knockdown of MBD2 or EZH2 alone (Fig. 6I).
In HCT116 cells, the results indicated that
knockdown of EZH2 increased the percentage of early apoptosis of
CRC cells, while knockdown of MBD2 had no significant effect on it.
Knockdown of both MBD2 and EZH2 increased the percentage of early
apoptosis of CRC cells, and there was a statistical difference
compared with the knockdown of either MBD2 or EZH2 alone. Knockdown
of MBD2 and EZH2 separately and simultaneously increased the
percentage of late apoptosis of CRC cells compared with the si-NC
group. Moreover, knockdown of MBD2 and EZH2 simultaneously
increased the percentage of total apoptosis of the CRC cells, and
the effect was more significant than the group with knockdown of
EZH2, while there was no significant difference compared with the
MBD2-knockdown group (Fig. 6J).
Effects of MBD2 and EZH2 silencing on
the cell cycle distribution of CRC cells
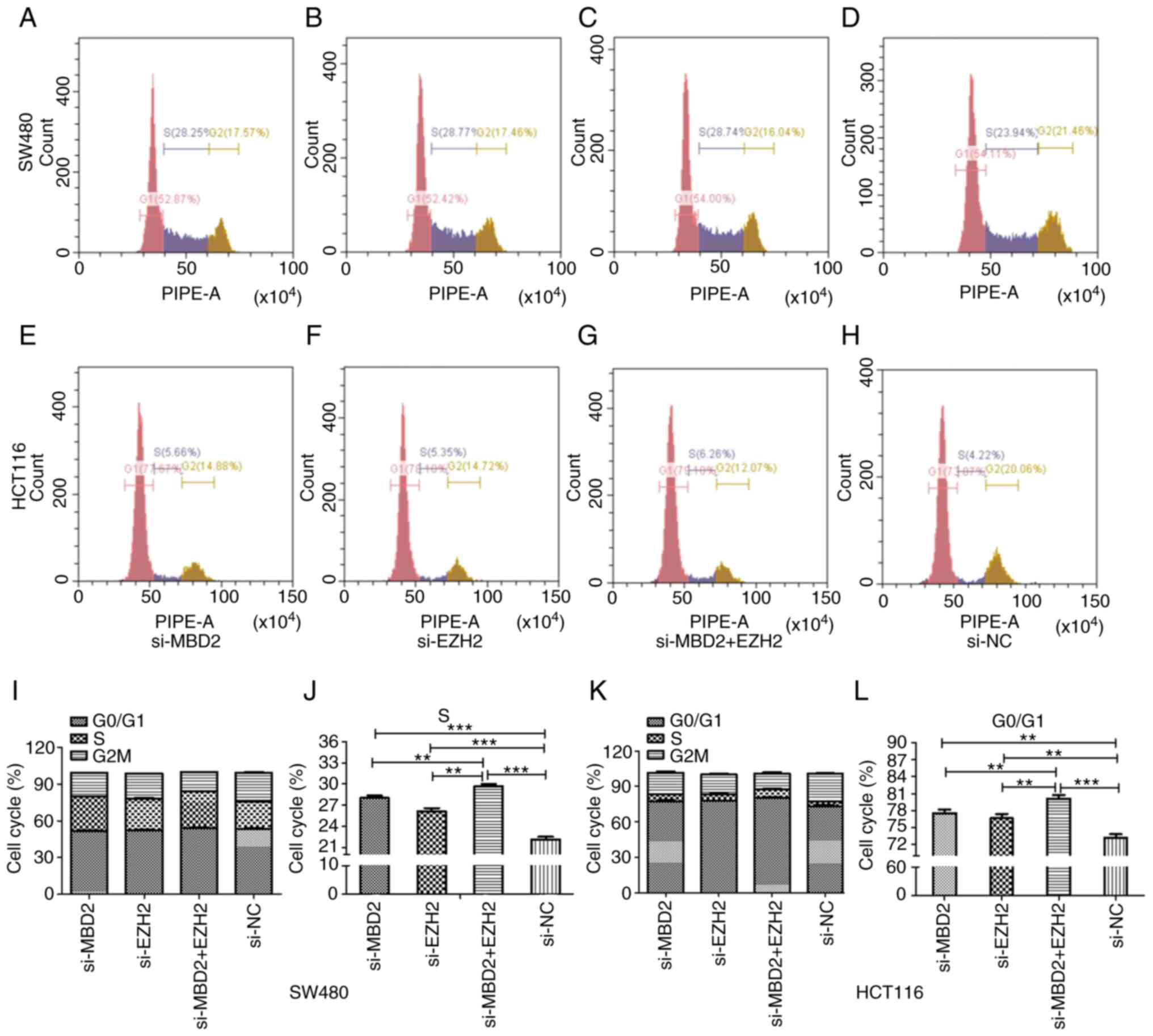
As shown in Fig. 7,
in SW480 and HCT116 cells, knockdown of MBD2 and EZH2 separately or
simultaneously affected the cell cycle distribution compared to the
si-NC group. More cells were arrested at S phase of the cell cycle
compared with the si-NC group in the SW480 cell line, and there was
a significant difference between the group with simultaneous
knockdown of MBD2 and EZH2 and the groups with knockdown of MBD2 or
EZH2 alone (Fig. 7I and J). While in
HCT116 cells, there was a higher percentage of cells arrested at
the G0/G1 phase of the cell cycle, and the percentage of cells at
G0/G1 phase in the group with simultaneous knockdown of MBD2 and
EZH2 was higher than that in the groups with knockdown of MBD2 or
EZH2 alone (Fig. 7K and L).
Effect of MBD2 and EZH2 silencing on
SFRP gene expression in CRC cells
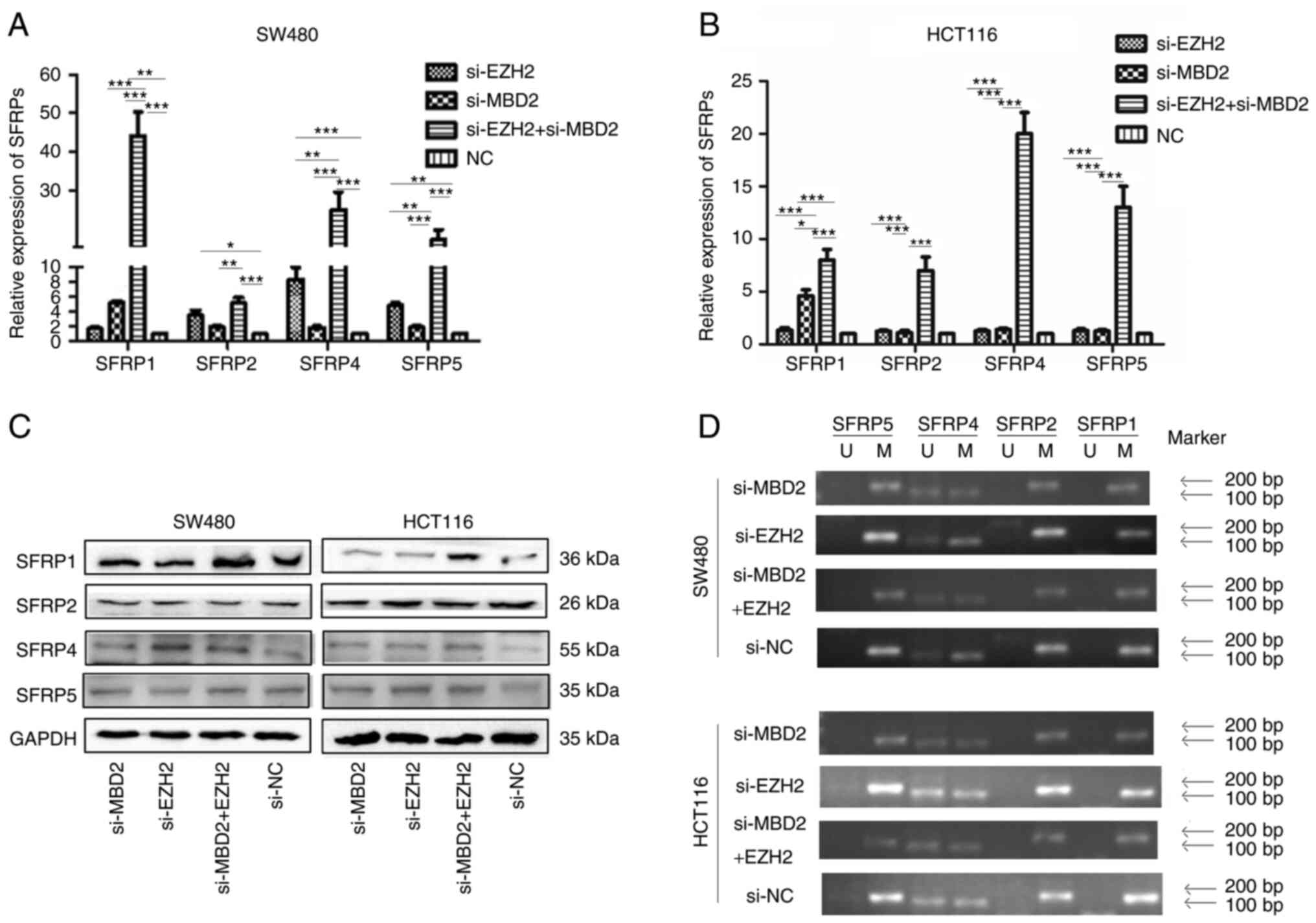
qPCR was used to detect the mRNA expression of SFRP
before and after knockdown of MBD2 and EZH2. As shown in Fig. 8A, in the SW480 cells, the expression
of SFRP1 was restored by knockdown of MBD2 (P<0.05), while the
expression levels of SFRP2, SFRP4 and SFRP5 could not be restored
by knockdown of MBD2 (P>0.05). The expression levels of SFRP2,
SFRP4 and SFRP5 could be restored by knockdown of EZH2, while the
expression of SFRP1 could not be restored by knockdown of EZH2
(P>0.05), and the expression of SFRP1, SFRP2, SFRP4 and SFRP5
could be significantly restored by simultaneous knockdown of MBD2
and EZH2 (P<0.05), which was significantly different from the
groups with knockdown of MBD2 and EZH2 separately. In HCT116 cells
(Fig. 8B), the expression levels of
SFRP1, SFRP2, SFRP4, and SFRP5 were not restored by knockdown of
EZH2. Knockdown of MBD2 restored the expression of the SFRP1 gene
(P<0.05), but had no significant effect on the expression levels
of SFRP2, SFRP4, and SFRP5 (P>0.05). However, knockdown of MBD2
and EZH2 markedly restored the expression of SFRP1, SFRP2, SFRP4,
and SFRP5 (P<0.05), which was significantly different from the
group with knockdown of MBD2 or EZH2 alone (P<0.05). The results
indicated that the expression levels of SFRP1, SFRP2, SFRP4, and
SFRP5 could be more effectively restored by knockdown of MBD2 and
EZH2 together in CRC cells.
Western blot analysis was used to investigate the
effect of MBD2 and EZH2 silencing on the expression of SFRP protein
in CRC cells. As shown in Fig. 8C,
in SW480 and HCT116, there was no significant difference in the
expression of SFRP1, SFRP2, SFRP4, and SFRP5 proteins by knockdown
of MBD2 or EZH2 alone compared to the control group, while the
protein levels of SFRP1 and SFRP4 were significantly restored by
knockdown of MBD2 and EZH2 together.
Effect of MBD2 and EZH2 on promoter methylation of
SFRP gene in colorectal cancer cells. As shown in Fig. 8D, methylation-specific PCR results
showed that following either knockdown of MBD2 and EZH2
simultaneously or separately, the SFRP1, SFRP2, and SFRP5 promoters
were all methylated except for partial methylation of the SFRP4
promoter in SW480 and HCT116 cells compared with control group.
There was no significant change in the methylation status of SFRP1,
SFRP2, SFRP4, and SFRP5 gene promoter between before and after
interfering with MBD2, EZH2, and both.
Discussion
As a negative regulator of Wnt signaling, the
secreted frizzled related proteins (SFRPs) can directly block the
transmission of the Wnt signaling pathway and are downregulated in
many types of tumors due to hypermethylation of the promoter
(25–28). In the present study, we found that
SFRP genes were downregulated in colorectal tumors by GEPIA and
MethHC online database, and this was inversely correlated with
hypermethylation of the promoter, indicating that hypermethylation
of the promoter is an important reason for downregulation of SFRP
genes.
Methyl-CpG binding domain protein 2 (MBD2) and
enhancer of zeste homolog 2 (EZH2) are important members of the
methylated DNA binding domain (MBD) and polycomb group (PcG)
protein family, respectively, and play important roles in DNA
methylation and histone modification. It was found that MBD2
recognizes methylated promoters and forms transcriptional
repressors by recruiting histone deacetylase complexes NuRD/Mi-2
and Sin3A to inhibit gene expression (17,18,29).
EZH2 as the main effector component of the PcG protein family, can
recruit DNMT1, DNMT3A and DNMT3B to the promoter region of the
target gene and binds to the gene promoter to maintain the
stability of the gene promoter methylation, causing chromatin
contraction and RNA polymerase II function pause (13). Therefore, MBD2 and EZH2 may be
important molecules regulating the expression of SFRP genes in
colorectal tumors.
Studies have found that MBD2 has been linked to
disease such as immune system function and tumorigenesis (17,30,31),
while EZH2 is closely related to tumor migration (32), proliferation (15), and invasion (33), and may be an important target for
tumor treatment (34,35). The Comet team summed up the role of
EZH2 in tumors, and found that EZH2 has the characteristics of
promoting and suppressing cancer, indicating that the relationship
between EZH2 and tumors is highly controversial (36). In our study, downregulating EZH2 was
able to inhibit tumor cell proliferation and invasion, but
downregulation of MBD2 did not affect it, and simultaneous
knockdown of MBD2 and EZH2 significantly inhibited the migration,
proliferation and cell cycle progression of colorectal tumor cells
and increased apoptosis, indicating that MBD2 can enhance the
biological function of EZH2 in CRC cells. Further studies have
shown that blocking MBD2 in colorectal tumor cells can restore
SFRP1 gene expression, but cannot restore SFRP2, SFRP4, and SFRP5
expression, indicating that MBD2 may have different regulatory
mechanisms for different member genes of the same gene family. In
addition, the expression of SFRP2, SFRP4, and SFRP5 in SW480 cells
could be restored by blocking EZH2, while the expression of SFRP2,
SFRP4 and SFRP5 in HCT116 cells could not be restored, indicating
that EZH2 has different regulatory mechanisms for SFRP gene
expression in different stages of tumor, and may have cell
specificity. Furthermore, knockdown of both MBD2 and EZH2 could
remarkably restore the expression of SFRP1, SFRP2, SFRP4, and
SFRP5, indicating that MBD2 and EZH2 have synergistic effects in
regulating SFRP gene expression. The results of
methylation-specific PCR showed that the promoter methylation
status of SFRP did not change before and after the knockdown of
MBD2 and EZH2 separately or simultaneously. Taken together, MBD2
recognizes and binds to methylated CG in the MBD domain of the SFRP
gene promoter, recruits histone deacetylase complexes NuRD/Mi-2 and
Sin3A to form transcriptional repressors, and promotes local
chromatin condensation, and to exert the function of
transcriptional inhibition (18).
Therefore, this effect can be enhanced by EZH2, which also has
chromatin remodeling. Silencing of MBD2 and EZH2 together can block
the action of histone deacetylase, alleviate the inhibition of
chromatin, and help restore gene expression. This phenomenon can
explain the synergistic effect of blocking MBD2 and EZH2 in
restoring SFRP gene expression and the biological function of
colorectal tumors.
However, our previous studies have shown that
histone deacetylation is not the main mechanism of SFRP gene
silencing. Inhibition of histone deacetylase in colorectal cancer
cell HCT116 did not restore SFRP expression (8). This mechanism cannot fully explain the
role of MBD2 in restoring SFRP expression. In combination with the
latest findings of transcription factor (37), we suspect that there is another
possibility that transcription factors can still function in some
way in the state of DNA methylation. MBD2 binds to methylated DNA
and blocks the reaction between the transcription factor and
methylated DNA. As shown in Fig.
S1, in normal colorectal cells, the SFRP gene promoter is not
methylated, the transcription factor that binds to unmethylated DNA
can bind to the promoter, and SFRP can be normally transcribed; in
colorectal tumor cells, the SFRP gene promoter is methylated,
blocking the role of transcription factors that bind to
unmethylated DNA. At the same time, MBD2, which has more advantages
in binding to methylated DNA, recognizes and binds to the
methylated region of the SFRP promoter, preventing transcription
factor binding to methylated DNA was cut off, leading to complete
inactivation of the SFRP gene in colorectal tumors; when MBD2 is
blocked in colorectal tumors, the transcription factor that bound
methylated DNA played its role at this time, and the SFRP genes can
resume transcription.
This study preliminarily demonstrated the
synergistic regulation of MBD2 and EZH2 on SFRP gene family
expression and biological function in colorectal tumor cells.
Subsequent research will further explore the transcription factors
involved in the regulation of SFRP gene expression and clarify the
specific mechanism of DNA methylation regulation of SFRP expression
in colorectal tumors, and provide a new theoretical basis for the
purpose of treating colorectal tumors.
Supplementary Material
Supporting Data
Acknowledgements
The abstract was presented at the 27th United
European Gastroenterology (UEG) Week, October 19–23, 2019 in
Barcelona, Spain.
Funding
This study was supported by the Applied Basic
Research Programs of the Wuhan Science and Technology Department
(2015061701011642). It was also funded by the Hubei Provincial
Health and Health Commission Joint Fund Key Project
(WJ2019H056).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YX designed the study, performed the experiments and
drafted the manuscript. FW, JZ, JY and YL prepared the material
used in the experiments and were involved in the literature search.
JQ evaluated all the data and revised the manuscript. ML and JZ
were involved in the statistical evaluation and in amending the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lv C, Li F, Li X, Tian Y, Zhang Y, Sheng
X, Song Y, Meng Q, Yuan S, Luan L, et al: MiR-31 promotes mammary
stem cell expansion and breast tumorigenesis by suppressing Wnt
signaling antagonists. Nat Commun. 8:10362017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Russell JO and Monga SP: Wnt/β-catenin
signaling in liver development, homeostasis, and pathobiology. Annu
Rev Pathol. 13:351–378. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tammela T, Sanchez-Rivera FJ, Cetinbas NM,
Wu K, Joshi NS, Helenius K, Park Y, Azimi R, Kerper NR, Wesselhoeft
RA, et al: A Wnt-producing niche drives proliferative potential and
progression in lung adenocarcinoma. Nature. 545:355–359. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arnold M, Sierra MS, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global patterns and trends in
colorectal cancer incidence and mortality. Gut. 66:683–691. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suzuki H, Watkins DN, Jair KW, Schuebel
KE, Markowitz SD, Chen WD, Pretlow TP, Yang B, Akiyama Y, Van
Engeland M, et al: Epigenetic inactivation of SFRP genes allows
constitutive WNT signaling in colorectal cancer. Nat Genet.
36:417–422. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rattner A, Hsieh JC, Smallwood PM, Gilbert
DJ, Copeland NG, Jenkins NA and Nathans J: A family of secreted
proteins contains homology to the cysteine-rich ligand-binding
domain of frizzled receptors. Proc Natl Acad Sci USA. 94:2859–2863.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu J, Xie Y, Li M, Zhou F, Zhong Z, Liu Y,
Wang F and Qi J: Association between SFRP promoter hypermethylation
and different types of cancer: A systematic review and
meta-analysis. Oncol Lett. 18:3481–3492. 2019.PubMed/NCBI
|
|
8
|
Qi J, Zhu YQ, Luo J and Tao WH:
Hypermethylation and expression regulation of secreted
frizzled-related protein genes in colorectal tumor. World J
Gastroenterol. 12:7113–7117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qi J and Zhu YQ: Targeting the most
upstream site of Wnt signaling pathway provides a strategic
advantage for therapy in colorectal cancer. Curr Drug Targets.
9:548–557. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Santanach A, Blanco E, Jiang H, Molloy KR,
Sansó M, LaCava J, Morey L and Di Croce L: The Polycomb group
protein CBX6 is an essential regulator of embryonic stem cell
identity. Nat Commun. 8:12352017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bracken AP and Helin K: Polycomb group
proteins: Navigators of lineage pathways led astray in cancer. Nat
Rev Cancer. 9:773–784. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kerppola TK: Polycomb group complexes-many
combinations, many functions. Trends Cell Biol. 19:692–704. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rush M, Appanah R, Lee S, Lam LL, Goyal P
and Lorincz MC: Targeting of EZH2 to a defined genomic site is
sufficient for recruitment of Dnmt3a but not de novo DNA
methylation. Epigenetics. 4:404–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Viré E, Brenner C, Deplus R, Blanchon L,
Fraga M, Didelot C, Morey L, Van Eynde A, Bernard D, Vanderwinden
JM, et al: The Polycomb group protein EZH2 directly controls DNA
methylation. Nature. 439:871–874. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang CJ, Yang JY, Xia W, Chen CT, Xie X,
Chao CH, Woodward WA, Hsu JM, Hortobagyi GN and Hung MC: EZH2
promotes expansion of breast tumor initiating cells through
activation of RAF1-β-catenin signaling. Cancer Cell. 19:86–100.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takawa M, Masuda K, Kunizaki M, Daigo Y,
Takagi K, Iwai Y, Cho HS, Toyokawa G, Yamane Y, Maejima K, et al:
Validation of the histone methyltransferase EZH2 as a therapeutic
target for various types of human cancer and as a prognostic
marker. Cancer Sci. 102:1298–1305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lai AY and Wade PA: Cancer biology and
NuRD: A multifaceted chromatin remodelling complex. Nat Rev Cancer.
11:588–596. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Desai MA, Webb HD, Sinanan LM, Scarsdale
JN, Walavalkar NM, Ginder GD and Williams DC Jr: An intrinsically
disordered region of methyl-CpG binding domain protein 2 (MBD2)
recruits the histone deacetylase core of the NuRD complex. Nucleic
Acids Res. 43:3100–3113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee J, Lee MS, Jeoung DI, Kim YM and Lee
H: Promoter CpG-site methylation of the KAI1 metastasis suppressor
gene contributes to its epigenetic repression in prostate cancer.
Prostate. 77:350–360. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang WY, Hsu SD, Huang HY, Sun YM, Chou
CH, Weng SL and Huang HD: MethHC: A database of DNA methylation and
gene expression in human cancer. Nucleic Acids Res. 43((Database
Issue)): D856–D861. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou FF, Xie W, Chen SQ, Wang XK, Liu Q,
Pan XK, Su F and Feng MH: SLC38A1 promotes proliferation and
migration of human colorectal cancer cells. J Huazhong Univ Sci
Technolog Med Sci. 37:30–36. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie Y, Yu J, Wang F, Li M, Qiu X, Liu Y
and Qi J: ERCC6L promotes cell growth and invasion in human
colorectal cancer. Oncol Lett. 18:237–246. 2019.PubMed/NCBI
|
|
25
|
Zhou Z, Wang J, Han X, Zhou J and Linder
S: Up-regulation of human secreted frizzled homolog in apoptosis
and its down-regulation in breast tumors. Int J Cancer. 78:95–99.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Suzuki H, Gabrielson E, Chen W, Anbazhagan
R, van Engeland M, Weijenberg MP, Herman JG and Baylin SB: A
genomic screen for genes upregulated by demethylation and histone
deacetylase inhibition in human colorectal cancer. Nat Genet.
31:141–149. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wong SC, Lo SF, Lee KC, Yam JW, Chan JK
and Wendy Hsiao WL: Expression of frizzled-related protein and
Wnt-signalling molecules in invasive human breast tumours. J
Pathol. 196:145–153. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vincan E and Barker N: The upstream
components of the Wnt signalling pathway in the dynamic EMT and MET
associated with colorectal cancer progression. Clin Exp Metastasis.
25:657–663. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu K, Xu C, Lei M, Yang A, Loppnau P,
Hughes TR and Min J: Structural basis for the ability of MBD
domains to bind methyl-CG and TG sites in DNA. J Biol Chem.
293:7344–7354. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Álvarez-Errico D, Vento-Tormo R, Sieweke M
and Ballestar E: Epigenetic control of myeloid cell
differentiation, identity and function. Nat Rev Immunol. 15:7–17.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wood KH, Johnson BS, Welsh SA, Lee JY, Cui
Y, Krizman E, Brodkin ES, Blendy JA, Robinson MB, Bartolomei MS and
Zhou Z: Tagging methyl-CpG-binding domain proteins reveals
different spatiotemporal expression and supports distinct
functions. Epigenomics. 8:455–473. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gunawan M, Venkatesan N, Loh JT, Wong JF,
Berger H, Neo WH, Li LY, La Win MK, Yau YH, Guo T, et al: The
methyltransferase Ezh2 controls cell adhesion and migration through
direct methylation of the extranuclear regulatory protein talin.
Nat Immunol. 16:505–516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xia L, Zhu X, Zhang L, Xu Y, Chen G and
Luo J: EZH2 enhances expression of CCL5 to promote recruitment of
macrophages and invasion in lung cancer. Biotechnol Appl Biochem.
67:1011–1019. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
First EZH2 inhibitor approved-for rare
sarcoma. Cancer Discov. 10:333–334. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mohammad F, Weissmann S, Leblanc B, Pandey
DP, Højfeldt JW, Comet I, Zheng C, Johansen JV, Rapin N, Porse BT,
et al: EZH2 is a potential therapeutic target for H3K27M-mutant
pediatric gliomas. Nat Med. 23:483–492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Comet I, Riising EM, Leblanc B and Helin
K: Maintaining cell identity: PRC2-mediated regulation of
transcription and cancer. Nat Rev Cancer. 16:803–810. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yin Y, Morgunova E, Jolma A, Kaasinen E,
Sahu B, Khund-Sayeed S, Das PK, Kivioja T, Dave K, Zhong F, et al:
Impact of cytosine methylation on DNA binding specificities of
human transcription factors. Science. 356:eaaj22392017. View Article : Google Scholar : PubMed/NCBI
|