Introduction
Colon cancer is one of the most frequently diagnosed cancers and the third leading cause of cancer-related death (1). According to the American Cancer Society, 104,270 individuals will be diagnosed with colon cancer in the United States in 2021, with 52,980 succumbing to the disease (2). Surgical resection is the standard treatment for colon cancer. However, metastasis to other sites, such as the liver, lung and peritoneum, has frequently been reported (3). Aside from surgery, chemotherapeutic drugs, including 5-fluorouracil and oxaliplatin, are employed as the drugs of choice in colon cancer (4). Nevertheless, these drugs have adverse effects on patients, such as risk of infection, anemia, bruising, bleeding and diarrhea, resulting in a number of deaths each year (5). Therefore, there is an urgent need for potent drugs that can lessen these side effects. Biomolecules derived from food, such as flavonoids, may prove to be a promising strategy for cancer treatment with no side effects.
Naringenin (NR) is a flavanone commonly found in citrus fruits, grapefruit and tomatoes (6). It demonstrates a wide range of pharmacological activities, such as antioxidant, free radical scavenging, anti-inflammatory, immunomodulatory, antimutagenic and anticarcinogenic effects (7,8). NR has been reported to inhibit cytochrome P450 1A1, proliferating cell nuclear antigen and NF-κβ expression in the lung of benzo(a)pyrene-induced mice, resulting in anti-initiating, antiproliferative and anti-inflammatory effects (9). Moreover, it acts as a potent inducer of apoptosis in pancreatic cancer cells (10).
Diosmin (DS) is a flavone glycoside found naturally in various plant sources (11). It is mainly found in citric fruits. It exhibits anti-inflammatory, free-radical scavenging and antimutagenic properties (12). DS has been studied in various animal models and human cancer cell lines and has been suggested to be chemopreventive and antiproliferative (13). It has also been reported that DS promotes apoptosis in the lung cancer cell line HA22T and dose-dependently reduces tumor size in xenograft nude mice via protein phosphatase 2A (14).
Suppression of apoptotic machinery is a hallmark of cancer development, and thus, apoptosis induction in cancer cells is a valuable treatment method. The genetic basis of apoptosis influence cell death that can be disrupted by mutations, contributing to malignancy (15). A significant decrease in apoptosis has been observed in some transgenic mice with a mutation in the p53 gene (16). Disruption of Bax, apaf-1 and caspase9 can promote oncogenic transformation and tumor development (17). Targeting events related to apoptosis could help lead to the development of potent and specific therapeutics.
Inflammation is often linked with the development of colon cancer. It plays multiple roles in cancer progression, including initiation, promotion and invasion, by influencing immune surveillance and associated signaling pathways (18). It is, in fact, one of the hallmarks of colon cancer. Approximately 20% of patients with inflammatory bowel disease develop colon cancer in their later years (19). Colon cancer tumors are associated with a robust display of inflammatory infiltration and enhanced pro-inflammatory cytokine expression (20). A number of types of colon cancers are stimulated by NF-κB and signal transducer and activator of transcription 3 (21). These two transcription factors are an essential part of the inflammatory pathway.
Several studies have investigated the individual anticancer effect of NR and DS against different cancers and have shown promising results (22,23). In recent times, combination therapy has been acknowledged to be effective in combating drug resistance and potentiating the efficacy of chemotherapy (24). A previous study demonstrated that combining NR and tamoxifen inhibited cell proliferation more effectively in estrogen receptor (ER)-positive breast cancer cells (25). It has also been reported that the combination of melatonin and thapsigargin can induce apoptosis and cell cycle arrest in gastric cancer cells (26). Hence, combining NR and DS can further enhance the apoptotic and anti-inflammatory effects and can even reduce the required doses of these compounds. The aim of the present study was to analyze the combinatorial impact of DS and NR on colon cancer cells by targeting apoptotic and inflammatory pathways. Various endpoints, such as cell cytotoxicity, fluorescence microscopy, DNA fragmentation, cell cycle and cell apoptosis were studied to investigate the effect of DS and NR. The expression of certain apoptotic and inflammatory pathway genes were also explored in the hope that the combination of DS and NR (DiNar) may serve as a promising candidate for treating colon cancer in the future.
Materials and methods
Materials
NR and DS were procured from Sigma-Aldrich (Merck KGaA). Dulbecco's modified Eagle's medium (DMEM) powder and MTT were purchased from HiMedia Laboratories, LLC. Fetal bovine serum (FBS) and 0.25% trypsin were supplied by Gibco (Thermo Fisher Scientific, Inc.). Moreover, cDNA and SYBR-Green Master Mix were purchased from Bio-Rad Laboratories, Inc. All other chemicals and reagents of molecular grade were supplied by Thermo Fisher Scientific, Inc. NR and DS were dissolved in DMSO.
Cell lines and culture conditions
Colon cancer cell lines HCT116 and SW480 were procured from the National Cell Culture Science. The cell lines were maintained in DMEM supplemented with 10% FBS and 1% penicillin/streptomycin with 5% CO2 at 37°C in NuAire CO2 incubator (NuAire Laboratory Equipment).
Cytotoxicity assay
MTT assay was performed to assess the cytotoxic effects of NR, DS and DiNar on colon cancer cell lines. A total of 1×104 cells per well were seeded in 96-well plates containing 100 µl complete medium. Following adhesion, cells were treated with different concentrations ranging from 10 to 320 µM of NR and DS for 48 h at 37°C, after which media was removed. Subsequently, 20 µl MTT solution (5 mg/ml) and 50 µl incomplete media were added. The cells were incubated for 3 h in a CO2 incubator at 37°C. The MTT solution was removed, and 150 µl DMSO was added to each well to dissolve the formazan crystals. After 10 min, absorbance was recorded at 570 nm with a microplate reader (Bio-Rad Laboratories, Inc.) (27).
Analysis of combined drug effect
Chou and Talalay's multiple drug effect analysis, based on the median-effect principle, was used to evaluate the combined effect of DS and NR (28). The combination index (CI) and dose reduction index (DRI), presented in Table I, were calculated to determine the synergism, and additive and antagonism effect of DS and NR, as described by Chou (29). Drug combination analysis was performed by CompuSyn software version 1 (30). CI values of <1, 1 and >1 indicate synergism, additive and antagonism, respectively.
 |
Table I.
Dose reduction index values for NR and DS combination.
|
Table I.
Dose reduction index values for NR and DS combination.
| A, HCT116 cells |
|
| |
Fa |
| |
|
| Treatment groups |
0.1 |
0.2 |
0.3 |
0.4 |
0.5 |
0.6 |
0.7 |
0.8 |
0.9 |
| NR |
3.69 |
3.31 |
3.08 |
2.90 |
2.75 |
2.60 |
2.46 |
2.28 |
2.05 |
| DS |
0.92 |
1.18 |
1.40 |
1.60 |
1.82 |
2.07 |
2.37 |
2.81 |
3.61 |
|
| B, SW480 cells |
|
| |
Fa |
| |
|
| Treatment groups |
0.1 |
0.2 |
0.3 |
0.4 |
0.5 |
0.6 |
0.7 |
0.8 |
0.9 |
|
| NR |
3.58 |
3.44 |
3.36 |
3.29 |
3.23 |
3.17 |
3.10 |
3.02 |
2.91 |
| DS |
1.19 |
1.97 |
1.18 |
2.19 |
2.52 |
2.89 |
3.36 |
4.05 |
5.34 |
DAPI assay
DAPI assay was performed to observe apoptotic cells. Briefly, cells were stained with fluorescent dye DAPI to detect nuclear condensation and fragmentation (31). Cells (~1×106) were seeded in 6-well plates. All cells were treated with 50% inhibitory concentrations (IC50) of drugs, except the control. After 48 h of treatment, cells were washed with 1X phosphate-buffered saline (PBS) and fixed with 500 µl 4% paraformaldehyde for 8 min at room temperature. Following fixation, cells were washed with PBS and permeabilized with 0.1% Triton X-100 for 10 min. Subsequently, rewashing and staining were performed with DAPI (300 nm) for 30 min in the dark at room temperature. The nuclear morphology of cells was visualized under a fluorescence microscope (Nikon Corporation).
DNA fragmentation assay
The untreated cells (control) and treated cells (1×106) were harvested for the DNA fragmentation assay. Cells were washed with PBS and lysed in lysis buffer (10 mmol/l Tris, 10 mmol/l EDTA, 0.5% Triton X-100) supplemented with proteinase K (1 mg/ml; SERVA Electrophoresis GmbH). Samples were then incubated at 37°C for 1 h and heated at 70°C for 10 min. RNase A (100 µg/ml; HiMedia Laboratories, LLC) was added, followed by repeated incubation at 37°C for 1 h. The samples were treated with phenol:chloroform:isoamyl alcohol (25:24:1) and centrifuged at 7,000 × g for 15 min at 4°C. The upper layer was collected in fresh tubes, and an equal volume of isopropanol and 3 M sodium acetate was added. The samples were incubated at −20°C for 2 h, and then centrifuged at 14,000 × g for 5 min at 4°C in a refrigerated centrifuge tube (Eppendorf). The extracted DNA was washed with 70% ethanol and dissolved in 1X TE buffer. DNA was electrophoresed on 1.8% (w/v) agarose gels containing ethidium bromide. The separated DNA fragments were visualized using a UV transilluminator (254 nm, Ultra-Lum Electronic UV Transilluminator) (32).
Cell cycle analysis
In 6-well plates, 1×106 HCT116 and SW480 cells were seeded. After 24 h of adhesion, cells were treated with NR, DS and DiNar for 48 h. After 48 h, cells were harvested, fixed in 70% chilled ethanol, and incubated for 2 h at −20°C. Then, cells were washed twice with PBS and suspended in propidium iodide (PI; 50 µg/ml) and RNase A (100 µg/ml). Cells in different cell cycle phases, G0/G1, S and G2/M, were analyzed by flow cytometry (BD FACSAria III; BD Biosciences) with BD FACSDiva 8.0.2 software (BD Biosciences).
Annexin V-fluorescein isothiocyanate (FITC)/PI apoptosis assay
Early and late apoptotic cells were analyzed according to the protocol of the Annexin V-FITC apoptosis detection kit (BD Biosciences). In 6-well plates, 2×105 HCT116 and SW480 cells were seeded and treated with NR, DS and DiNar. After 48 h of treatment, cells were harvested and washed twice with PBS. Later, the cells were suspended in 100 µl binding buffer mixed with 5 µl Annexin V-FITC and 5 µl PI stain. Then, the cells were incubated for 15 min in the dark. Finally, 400 µl binding buffer was added to each sample. The stained cells were analyzed for 15 min by flow cytometry (BD FACSAria III; BD Biosciences). The data were analyzed using FACSDiva 8.0.2 software (33).
Western blotting
After 48 h of treatment, 2×106 cells were collected. The cells were lysed in radioimmune precipitation assay (RIPA) buffer containing protease inhibitor (Geno Technology, Inc.). Proteins were quantified using a BCA assay kit (Thermo Fisher Scientific, Inc.). Protein samples (20 µg) were loaded in each well, and electrophoresis was performed via SDS-PAGE on a 12% gel. The proteins were then transferred onto the PVDF membrane (Bio-Rad Laboratories, Inc.). The membrane was blocked with 5% bovine serum albumin (Bio-Rad Laboratories, Inc.) for 1 h at room temperature before being incubated overnight at 4°C with the following primary antibodies: Bcl-2 (1:1,000; cat. no. 138800; Thermo Fisher Scientific, Inc.), Bax (1:1,000; cat. no. 336400; Thermo Fisher Scientific, Inc.), caspase-3 (1:1,000; cat. no. MA511521; Thermo Fisher Scientific, Inc.), caspase-8 (1:1,000; cat. no. PA520118; Thermo Fisher Scientific, Inc.), caspase-9 (1:1,000; cat. no. PA519904; Thermo Fisher Scientific, Inc.), p53 (1:1,500; cat. no. MCA1703; Bio-Rad Laboratories, Inc.), NF-κB (1:1,000; cat. no. 513500; Thermo Fisher Scientific, Inc.), IKK-β (1:1,000; cat. no. LFMA0192; Thermo Fisher Scientific, Inc.) and IKK-α (1:1,000; cat. no. PA517803; Thermo Fisher Scientific, Inc.). Then, membranes were incubated for 2 h at room temperature with the following HRP-conjugated secondary antibodies: Anti mouse (1:10,000; cat no. 1706516; Bio-Rad Laboratories, Inc.) or anti-rabbit (1:10,000; cat. no. 1706515; Bio-Rad Laboratories, Inc.). Membranes were developed with enhanced chemiluminescence solution (Merck KGaA). β-actin was used as an internal control (34).
Reverse transcription-quantitative (RT-q)PCR
The treated and untreated cells were harvested, washed with PBS and RNA was extracted using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). A total of 1 µg RNA was used to synthesize cDNA according to the manufacturer's instruction using a cDNA synthesis kit (Bio-Rad Laboratories, Inc.). Subsequently, qPCR was performed using SYBR-Green (Bio-Rad Laboratories, Inc.) in ABI 7900HT detection system (Applied Biosystems; Thermo Fisher Scientific, Inc.). Thermocycling conditions were as follows: 95°C for 5 min; followed by 40 cycles at 95°C for 30 sec, 58°C for 30 sec and 60°C for 30 sec. GAPDH was used as an internal control for RT-qPCR. The results were analyzed using the 2−ΔΔCq method (35). The primers used are listed in Table II.
|
Table II.
List of primers used for reverse transcription- quantitative PCR.
|
Table II.
List of primers used for reverse transcription- quantitative PCR.
| Primer name |
Primer sequences (5′→3′) |
| Bax |
F: GCCCTTTTGCTTCAGGGGATG |
| |
R: CAGCTGCCACTCGGAAAAAG |
| Bcl-2 |
F: CTGCACCTGACGCCCTTCACC |
| |
R: CACATGACCCCACCGAACTCAAAGA |
| Caspase-3 |
F: ACTCCACAGCACCTGGTTATT |
| |
R: TTCTGTTGCCACCTTTCGGT |
| Caspase-8 |
F: AGAGTCTGTGCCCAAATCAAC |
| |
R: GCTGCTTCTCTCTTTGCTGAA |
| Caspase-9 |
F: TTCCCAGGTTTTGTTTCCTG |
| |
R: CCTTTCACCGAAACAGCATT |
| p53 |
F: CCAGCAGCTCCTACACCGGC |
| |
R: AAACCGTAGCTGCCCTG |
| NF-κB |
F: CTGGACCGCTTGGGTAACTC |
| |
R: CCTGCCAATGAGATGTTGTC |
| IKK-α |
F: CATGCAGGGAAAAAGGCAGA |
| |
R: TGAGAGGAGTTACCACACATGACAG |
| IKK-β |
F: GGGGCCTGGGAAATGAAAGA |
| |
R: GGTCAGCCTGATTGTGCCAT |
| GAPDH |
F: GACAGTCAGCCGCATCTTCT |
| |
R: TTAAAAGCAGCCCTGGTGAC |
Statistical analysis
Each experiment was repeated in triplicate. Statistical analysis was performed using GraphPad Prism version 8 (GraphPad Software, Inc.) and data are expressed as the mean ± standard deviation. Results were analyzed using ANOVA followed by Tukey's multiple comparison test. P<0.05 was considered to indicate a statistically significant difference.
Results
Antiproliferative efficacy of DiNar on HCT116 and SW480 cells
The cytotoxic activity of NR, DS and DiNar on colon cancer cell lines HCT116 and SW480 was evaluated by MTT assay. Cells were treated with increasing concentrations of NR, DS and DiNar for 48 h. Results showed that NR, DS and DiNar dose-dependently inhibited the growth of cells, as shown in Fig. 1A and B. The IC50 of NR and DS on HCT116 cells were 169 and 160 µM, while on SW480 cells it was 210 and 152 µM, respectively. Furthermore, the data revealed that the cytotoxic effect was significantly higher in DiNar-treated cells compared with NR or DS treatments alone. The significant difference was more evident at higher concentrations. Moreover, the IC50 value of DiNar was lower. In HCT116 cells, the combination of 70 µM NR and 70 µM DS showed 50% death of cells. Similarly, in SW480, the combination of 60 µM NR and 60 µM DS resulted in 50% death of cells (data not shown). In subsequent experiments, the IC50 value of drugs was used to treat cells.
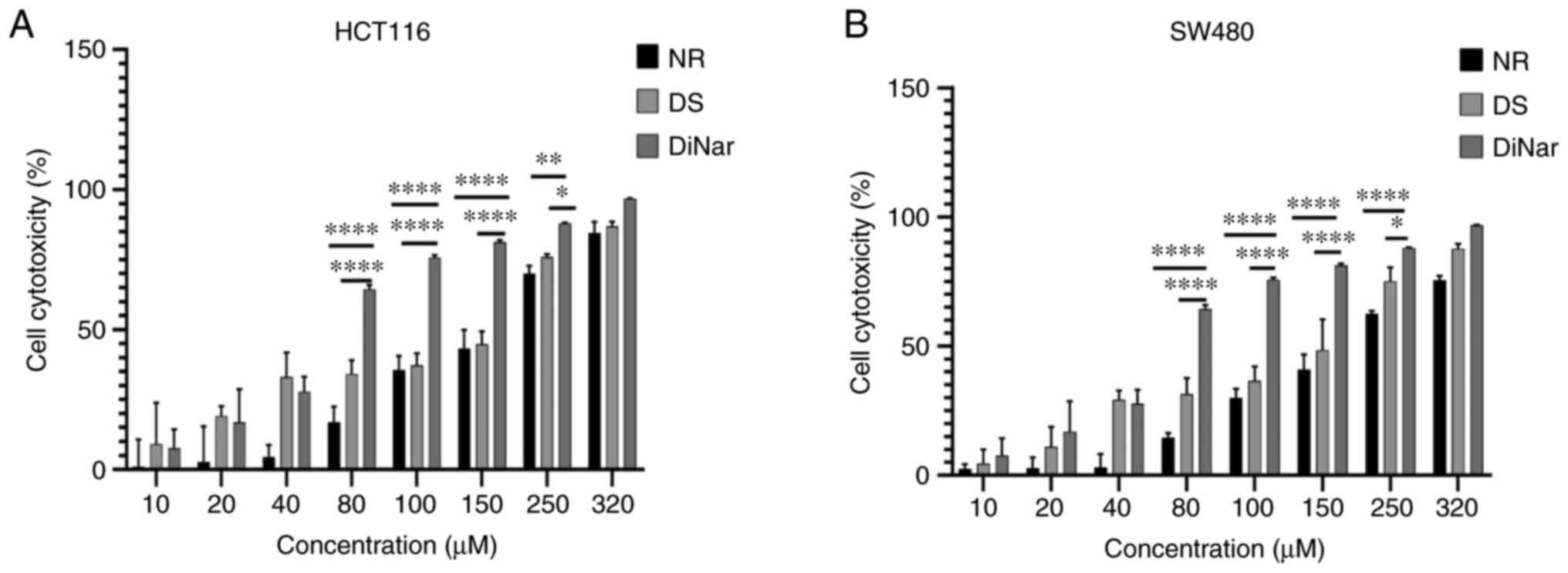 |
Figure 1.
MTT assay. (A) HCT116 and (B) SW480 cells were cultured and treated with increasing concentrations of NR (10, 20. 40, 80, 100, 150, 250 and 320 µM) and/or DS (10, 20, 40, 80, 100, 150, 250 and 320 µM) for 48 h. Cell cytotoxicity was measured by MTT assay. *P<0.05, **P<0.01 and ****P<0.0001. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
Combination of DS and NR exhibits a synergistic effect on the cytotoxicity of colon cancer cells
The combinatorial effect of DS and NR on colon cancer cell lines was evaluated by MTT assay. The data of the MTT assay were processed by CompuSyn software to measure the synergistic effect of the drugs. The CI-fa curve revealed that DiNar exerted a synergistic effect on cell growth inhibition in both HCT116 and SW480 cells, as shown in Fig. 2. In the case of HCT116 cells, CI values ranged from 0.96 to 0.76 at different effect levels from IC40 to IC90. Similarly, SW480 showed CI values ranging from 0.76 to 0.53 at various effect levels (IC40 to IC90). The CI values in both cell lines were <1, indicating the synergistic effect of DiNar. Furthermore, the values of DRI>1 in both cell lines indicated a decrease in drug dosage, resulting in less toxicity by these drugs, as mentioned in Table I.
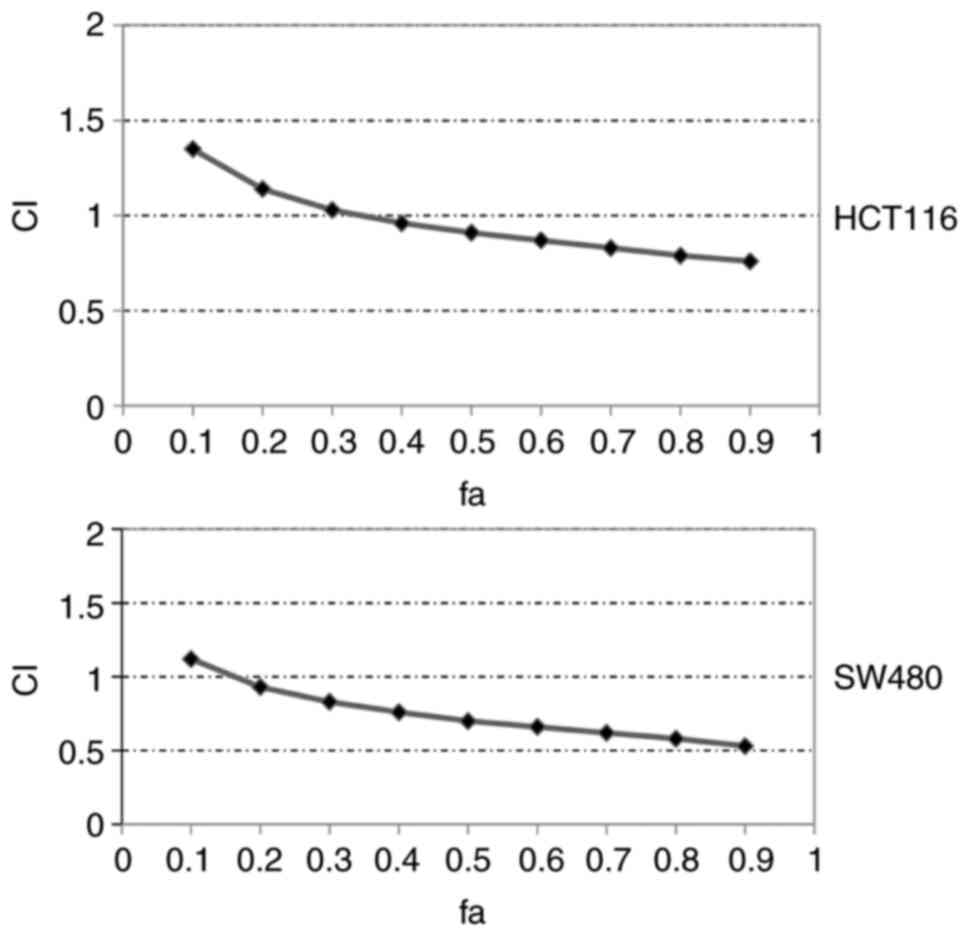 |
Figure 2.
CI of DiNar. CI of DiNar on HCT116 and SW480 cells was calculated. Fa indicates fraction affected (fa=0.4 indicates 40% of cell death). CI, combination index; DiNar, combined treatment of diosmin and naringenin; Fa, fraction affected.
|
Evaluation of DiNar-induced apoptosis using DAPI staining and DNA fragmentation
Both HCT116 and SW480 cells treated with NR, DS and DiNar were stained with DAPI (a nuclear dye that binds to DNA). Untreated HCT116 and SW480 cells were used as controls. Nuclear fragmentation, chromatin condensation and the formation of apoptotic bodies are hallmarks of apoptosis (36). NR, DS and DiNar treatments induced chromatin condensation in both HCT116 and SW480 cells, as shown in Fig. 3A and B, respectively. However, DiNar showed striking chromatin condensation compared with NR and DS alone.
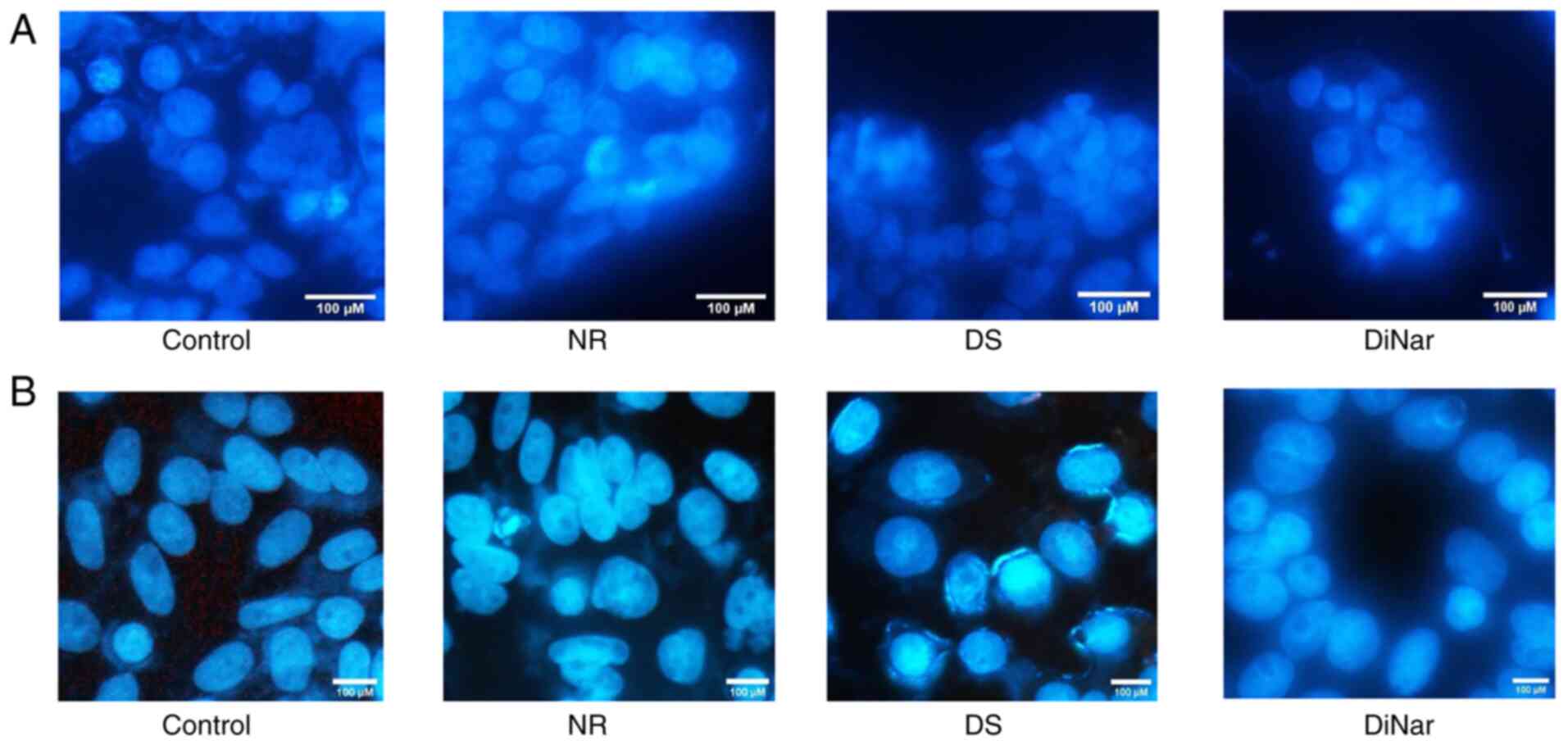 |
Figure 3.
DAPI assay. (A) HCT116 and (B) SW480 cells treated with NR, DS and DiNar for 48 h showed altered nuclear morphology. Control group showed normal morphology, whereas NR-, DS- and DiNar-treated cells showed increased nuclear condensation, cell shrinkage and staining intensity. Scale bar, 100 µM. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
Apoptosis was also assessed by analyzing DNA fragmentation. Fragments of DNA were observed in treated HCT116 and SW480 cells, while controls showed a single band. The DNA fragments were more extensively observed in the DiNar group as compared with the NR and DS groups. The results showed that the induction of apoptosis was enhanced in cells treated with DiNar (Fig. 4A and B).
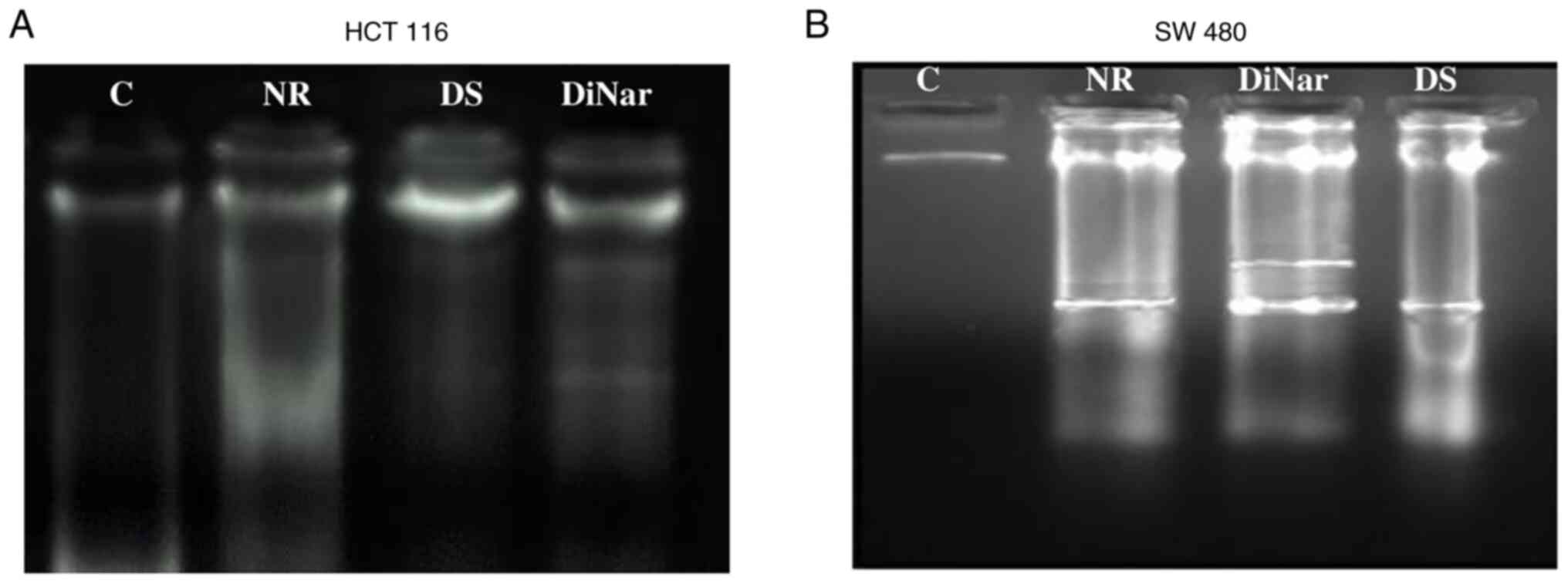 |
Figure 4.
Fragmentation of DNA. DNA fragmentation was observed in (A) HCT116 and (B) SW480 cells treated with NR, DS and DiNar. The control group showed no fragments, while the DiNar group showed more intense fragmentation as compared with NR and DS individually. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
Combined treatment of DiNar induces cell cycle arrest at the G1 phase
Cell cycle distribution in the NR, DS and DiNar groups was assessed by flow cytometry. Untreated HCT116 and SW480 cells were used as controls. In the control HCT116 cells, 28.35% of the cell population was in the G0/G1 phase, while NR-treated cells showed 55.30%, DS showed 55.75% and DiNar showed 65.35% cells in the G0/G1 phase of the cell cycle. The population of cells in G0/G1 was significantly higher in the DiNar group than NR and DS alone, as shown in Fig. 5A and B. Similar results were observed in SW480 cells as well. The control cells exhibited 71.95% cell population in the G0/G1 phase, whereas NR, DS, and DiNar showed 82.65, 84.75 and 89.25% population in the G0/G1 phase of the cell cycle, respectively. Thus, the percentage of cell population in the G0/G1 phase was greater in the DiNar group compared with the NR and DS alone, as shown in Fig. 5C and D. Thus, these findings illustrated that the treated cells were arrested in the G0/G1 phase of the cell cycle, and the effect was more pronounced in the case of DiNar.
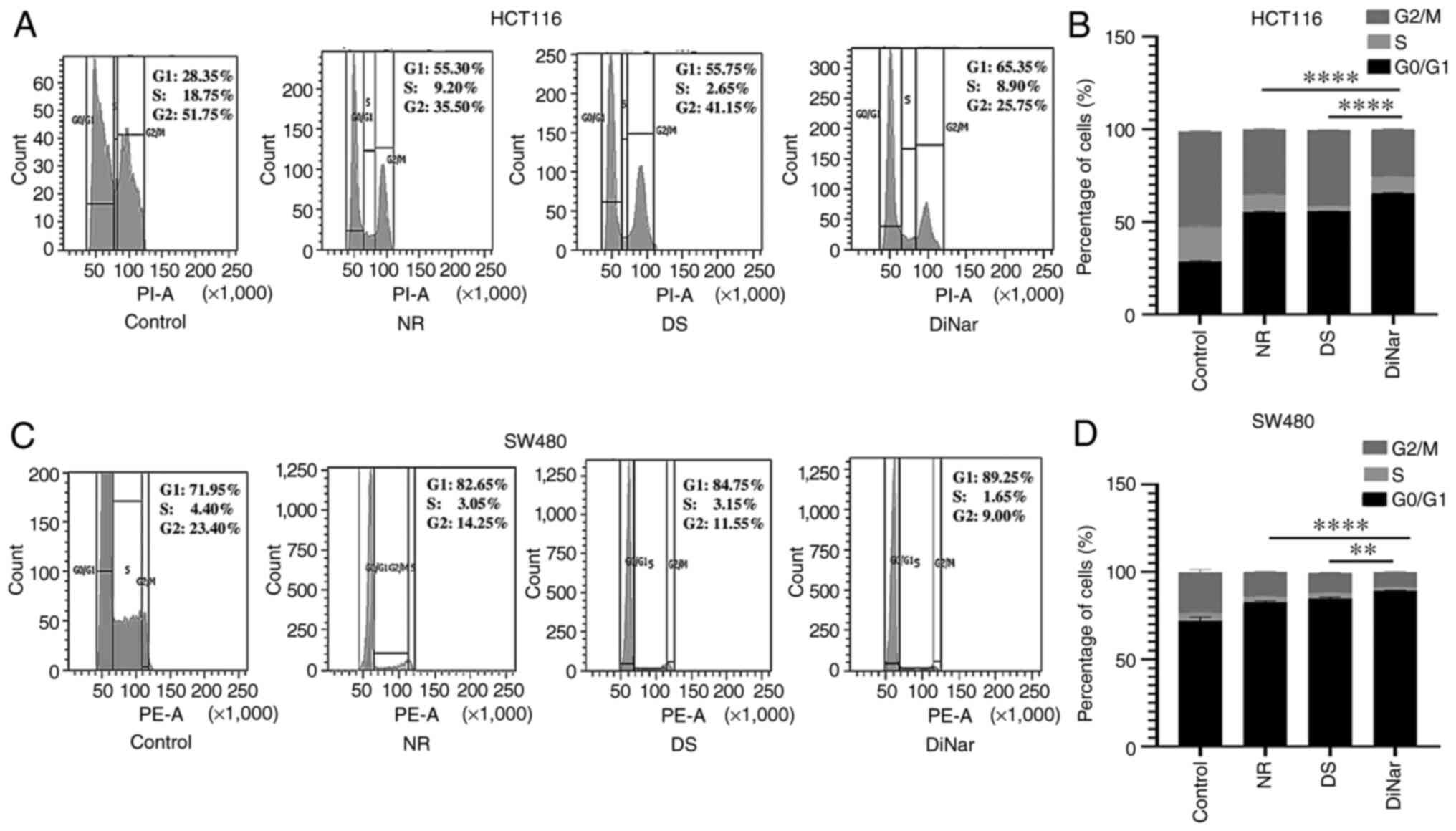 |
Figure 5.
Cell cycle analysis. NR, DS and DiNar induced cell cycle arrest in the G1 phase in (A) HCT116 and (C) SW480 cells. Cells were treated with IC50 values for 48 h. (B and D) Bar graphs represent the percentage of cells in G0/G1, S and G2/M phase of the cell cycle. The cell cycle arrest was evaluated by flow cytometry using PI staining. **P<0.01 and ****P<0.0001. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
Evaluation of DiNar-induced apoptosis via Annexin V-FITC labeling
To further investigate apoptosis, an Annexin V assay was performed. The flow cytometry analysis detected the percentage of viable cells, cells in early apoptosis, late apoptosis and necrosis stages. HCT116 cells treated with NR, DS and DiNar induced apoptosis rates of 34.5, 32.2 and 48.4%, respectively, while SW480 cells demonstrated apoptosis rates of 46.7, 36.3 and 55.0%, respectively. As shown in Fig. 6A-D, the results indicated that both NR and DS significantly increased cell apoptosis in HCT116 and SW480 cells compared with the respective controls. It was also evident that the apoptotic rate in the DiNar group was significantly higher than in the single treatment groups.
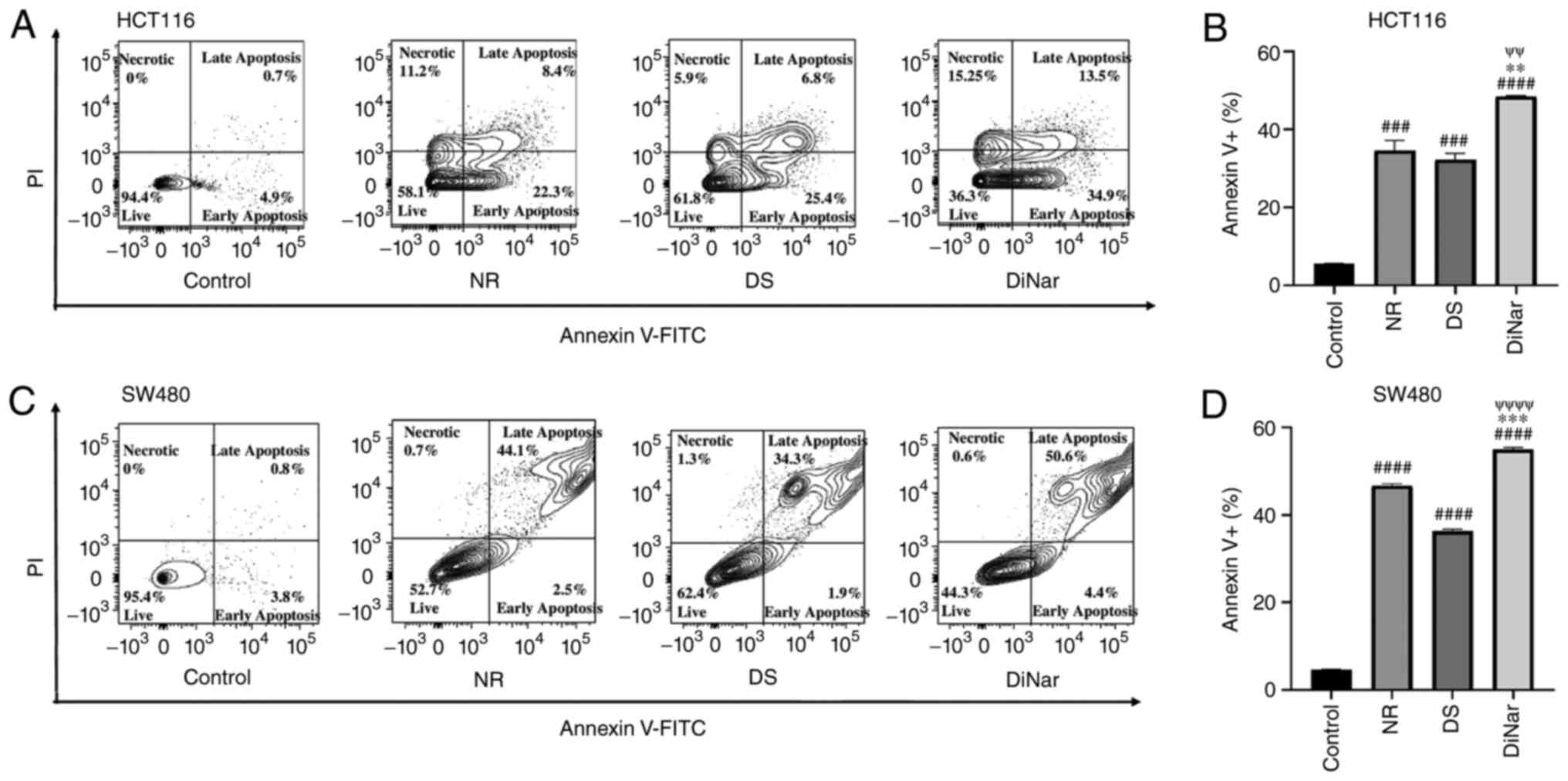 |
Figure 6.
Apoptosis assay by flow cytometry. NR, DS and DiNar induced apoptosis in HCT116 and SW480 cells as determined via Annexin V-FITC/PI staining assay. Cells were treated with IC50 concentration of NR, DS and DiNar for 48 h. Flow cytometry assay shows population of viable, early apoptotic, late apoptotic and necrotic (A) HCT116 and (B) SW480 cells. Bar graphs of (C) HCT116 and (D) SW480 cells showing increased apoptotic cells in the DiNar group. ###P<0.001 and ####P<0.0001 vs. control; **P<0.01 and ***P<0.001 vs. NR; ψψP<0.01 and ψψψψP<0.0001 vs. DS. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
Evaluation of apoptotic markers via western blotting and RT-qPCR
The expression levels of apoptotic proteins were investigated via western blotting. The results showed that the expression levels of Bax, caspase-3, caspase-8, caspase-9 and p53 were increased in both HCT116 and SW480 cells after treatment with NR and DS compared with the control cells (Fig. 7A-D). Moreover, the expression levels of these proteins were markedly increased in the DiNar group compared with the single treatment groups (Fig. 7A and C). The anti-apoptotic protein, Bcl-2, demonstrated the opposite trend. Its expression was reduced in both HCT116 and SW480 cells after NR and DS treatments compared with respective controls. The expression was further suppressed in cells treated with DiNar in both HCT116 and SW480 cells compared with the single treatment groups (Fig. 7A and C).
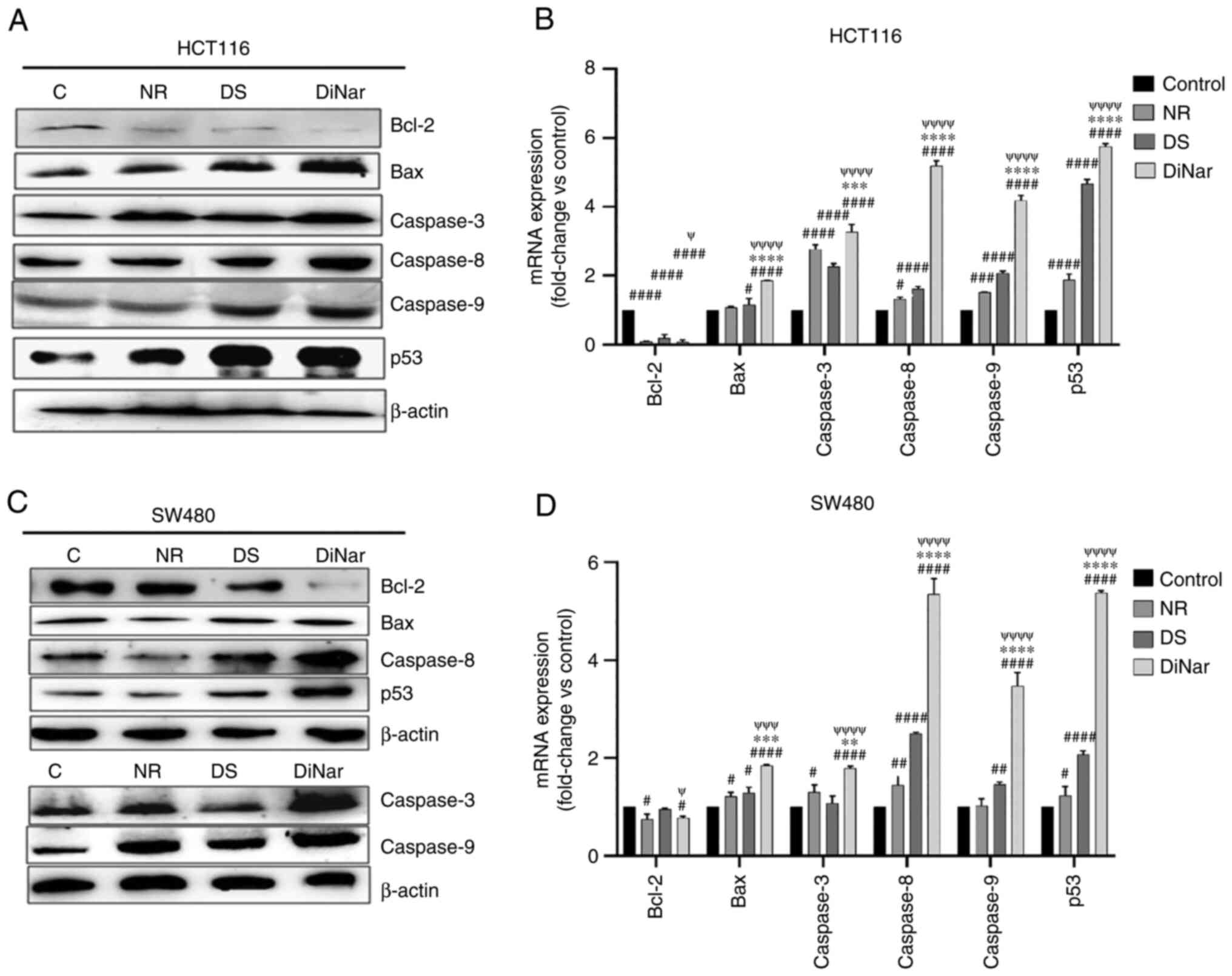 |
Figure 7.
Expression of apoptotic markers. NR-, DS- and DiNar-induced apoptosis was confirmed by detecting the upregulation of apoptotic protein Bax, caspase-3, caspase-8, caspase-9 and p53, and the downregulation of Bcl-2, in both (A) HCT116 and (C) SW480 cells, as determined via western blotting. β-actin acted as the reference protein. The fold-changes in the mRNA expression of these genes in (B) HCT116 and (D) SW480 cells were determined via reverse transcription-quantitative PCR using gene-specific primers. GAPDH was used as a control. #P<0.05, ##P<0.01, ###P<0.001 and ####P<0.0001 vs. control; **P<0.01, ***P<0.001 and ****P<0.0001 vs. NR; ψP<0.05, ψψψP<0.001 and ψψψψP<0.0001 vs. DS. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
In addition to the protein expression, the mRNA expression levels of apoptosis-related genes were analyzed via RT-qPCR. DiNar significantly reduced Bcl-2 expression in HCT116 and SW480 cells by 0.066-fold and 0.7-fold, respectively, when compared with the controls (Fig. 7B and D). On the other hand, the expression of Bax, caspase-3, caspase-8, caspase-9 and p53 mRNA levels in DiNar-treated HCT116 cells were increased by 1.85, 3.26, 5.18, 4.18 and 5.73-folds compared with the control group, respectively. Similarly, DiNar treatment enhanced the expression of Bax, caspase-3, caspase-8, caspase-9 and p53 mRNA in SW480 cells by 1.84, 1.79, 5.35, 3.47 and 5.37-folds, respectively. The expression of these mRNAs in DiNar-treated cells was significantly higher compared with the NR and DS treatments alone, as presented in Fig. 7B and D.
Evaluation of inflammatory marker expression
The expression levels of inflammatory marker proteins, NF-κB, IKK-α and IKK-β, were detected via western blotting. As shown in Fig. 8A and C, the expression levels of these inflammatory marker proteins were downregulated in NR- and DS-treated HCT116 and SW480 cells compared with the control group. However, DiNar showed remarkable suppression of these proteins in both HCT116 and SW480 cells.
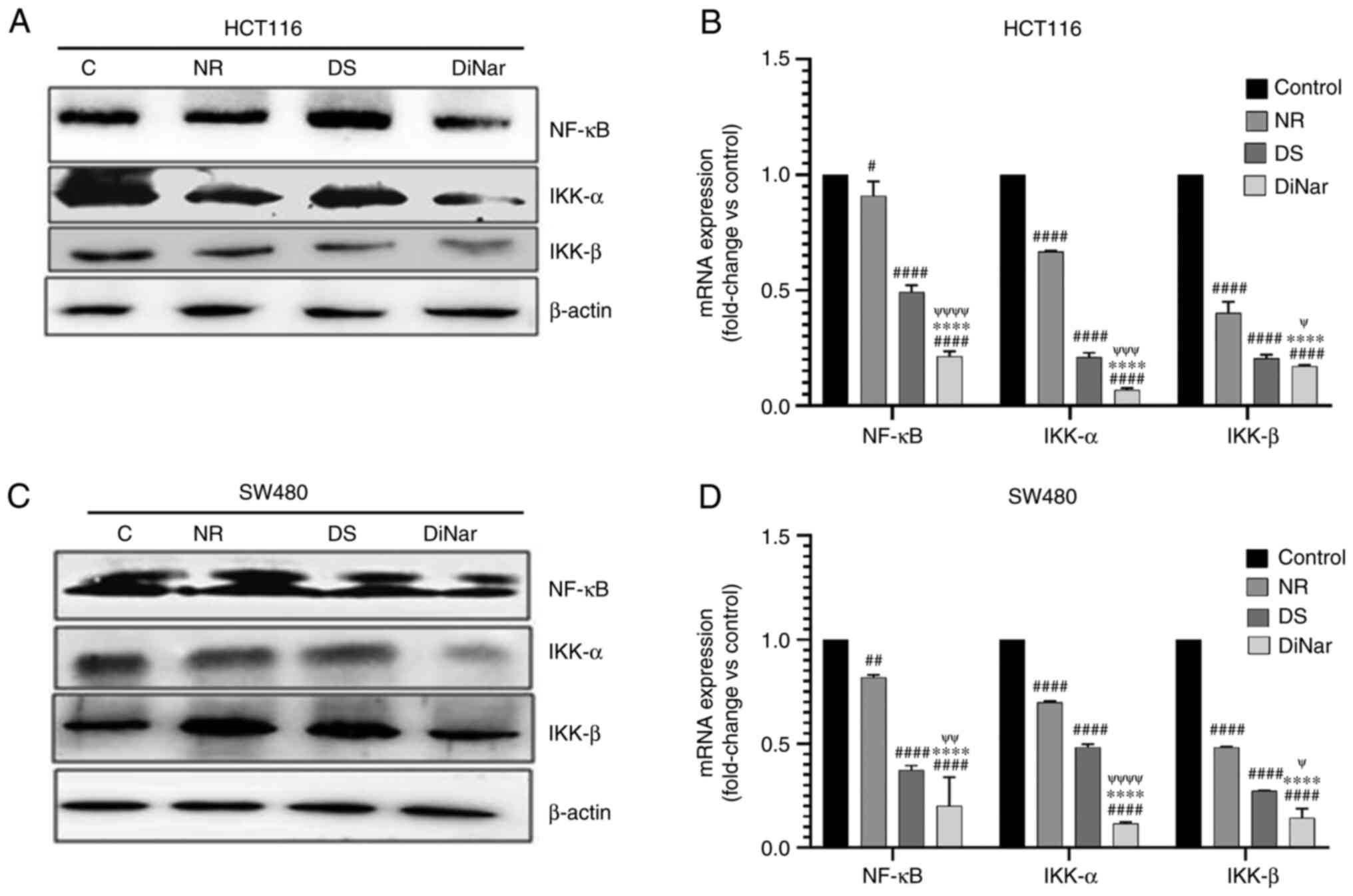 |
Figure 8.
Expression of inflammatory markers. NR-, DS- and DiNar-treated cells showed downregulation of inflammatory proteins, NF-κB, IKK-α and IKK-β, in both (A) HCT116 and (C) SW480 cells, as determined via western blotting. The lower band in NF-κB blot in part (C) represents NF-κB protein expression. β-actin acted as the reference protein. mRNA expression levels (fold-change vs. control) of these genes in (B) HCT116 and (D) SW480 cells were determined via reverse transcription-quantitative PCR using gene-specific primers. GAPDH was used as a control. #P<0.05, ##P<0.01 and ####P<0.0001 vs. control; ****P<0.0001 vs. NR; ψP<0.05, ψψP<0.01, ψψψP<0.001 and ψψψψP<0.0001 vs. DS. DS, diosmin; NR, naringenin; DiNar, combined treatment of DS and NR.
|
The mRNA expression levels of inflammatory marker genes, NF-κB, IKK-α and IKK-β, were investigated via RT-qPCR. The mRNA expression results were similar to the corresponding protein expression. The results indicated that DiNar significantly decreased the mRNA expression levels of NF-κB, IKK-α and IKK-β in HCT116 cells by 0.21, 0.068 and 0.169-folds, respectively, when compared with the control. In SW480 cells, DiNar treatment reduced mRNA expression levels of NF-κB, IKK-α and IKK-β by 0.2, 0.11 and 0.14-folds compared with the control. Moreover, DiNar treatment led to a significant decrease in mRNA expression in both HCT116 and SW480 cells compared with the NR and DS treatments alone, as shown in Fig. 8B and D.
Discussion
Flavonoids are a group of natural substances with varying phenolic structures. They are mainly found in fruits, vegetables, tea, flowers, roots and stems (37). They have a number of health benefits due to their anti-inflammatory, anticarcinogenic, antioxidant and antiproliferative properties (38). These characteristics make them potential anticancer candidates. Conventional therapies employed for treating colon cancer cause severe side effects in patients (39). There is a dire need for alternative therapeutic options with fewer side effects. Fruits and vegetables high in flavonoids are often recommended to reduce the risk of cancer (40). Some of these flavonoids are being considered as potential cancer therapeutic agents (38). In the present study, it was revealed that the flavonoids used in combination could synergistically act against the colon cancer cells and inhibit proliferation.
Multiple studies have shown that NR, a flavanone, has a significant effect on various cancer cell lines (9,41). It has an ER-dependent antiproliferative effect on cancer cells (42). Similarly, DS, a flavone glycoside, has also been reported for anti-inflammatory, antimutagenic and antioxidant properties (12). It induces apoptosis, senescence and autophagy in breast cancer cells (43). Furthermore, apoptosis induction via genotoxicity and apoptosis has been described in prostate cancer cells (44). Considering the numerous promising characteristics of flavonoids, it was hypothesized in the present study that they would have high therapeutic potential when used in combination.
The current study evaluated the combined effect of DS and NR on colon cancer cell lines HCT116 and SW480. According to these findings, DiNar had a greater inhibitory effect on both the colon cancer cell lines than NR and DS alone. The data showed that the IC50 values of NR were 169 and 210 µM for HCT116 and SW480 cells, respectively. Likewise, DS was found to have IC50 values of 160 and 152 µM on HCT116 and SW480 cell lines, respectively. The combination DiNar showed lower IC50 values when tested in different permutations and combinations. In the equimolar drug combination, IC50 values of 70 µM NR + 70 µM DS for HCT116 and 60 µM NR + 60 µM DS for SW480 were observed.
Furthermore, CI analysis was performed to investigate the synergistic effect of DS and NR on colon cancer cells. DiNar demonstrated a wide range of synergism, with CI values ranging from 0.96 to 0.76 for IC40 to IC90 in HCT116 cells and 0.93 to 0.53 for IC20 to IC90 in SW480 cells. This suggested that DiNar offered a beneficial effect on cytotoxicity in colon cancer cells. Besides, DiNar also yielded DRI values ranging from 3.6 to 5.3 in HCT116 and SW480 cells, indicating that the combination was more effective as an anticancer therapy with fewer side effects. Hence, this study demonstrated that the combination DiNar treatment inhibited cell proliferation in a synergistic manner.
The cell cycle regulates the machinery of cellular proliferation with a cascade of events that bring about cell division and duplication (45). Dysregulation of cell cycle results in abnormal and unchecked cell proliferation leading to cancer (46). A number of therapeutic agents act by inducing cell cycle arrest at a specific checkpoint. It is the most common mechanism of anticancer drugs (47). Various agents have been reported to cause cell cycle arrest at the G0/G1, S or G2/M phase (34). In the HepG2 cell line, NR was reported to inhibit cell proliferation by accumulating cells in the G0/G1 and G2/M phases of the cell cycle (48). The present study confirmed that DiNar caused greater cell population arrest in the G0/G1 phase of the cell cycle than either NR or DS. This observation was consistent in both HCT116 and SW480 cell lines.
Apoptosis, programmed cell death, is an essential physiological process for the development and homeostasis of tissues (49). Suppression of apoptosis is one of the hallmarks of cancer (50). It is associated with a series of changes in cellular morphology, such as chromatin condensation, DNA fragmentation and nuclear blebbing (51). DAPI is a fluorescent dye reported to enter the pores of the damaged cells and stains fragmented nuclear DNA (52). The intensity of the DAPI stain represents the amount of chromatin condensation. The light and uniform DAPI stain represent normal and healthy cells with no chromatin condensation, while the brighter stain indicates condensed chromatin (52). Condensed chromatin is one of the features of apoptosis (36). In the current study, these morphological changes, such as chromatin condensation, were observed in HCT116 and SW480 cells after treatment with NR, DS and DiNar. However, the morphological changes were more distinct in the cells treated with DiNar compared with NR and DS treatments alone. A DNA fragmentation assay further confirmed these results. Fragmentation of DNA is one of the features of cell apoptosis (36). This phenomenon generates fragments with varying base pairs size on agarose gel electrophoresis. In epidermoid carcinoma cells, NR has been shown to induce nuclear condensation, DNA fragmentation and cell cycle arrest in the G0/G1 phase (53). The current study observed that apoptotic DNA appeared more extensively in ladder form on an agarose gel in DiNar-treated cells compared with NR and DS treatments alone. Thus, these results further confirmed that the cytotoxic effect of NR, DS and DiNar was mediated by an apoptotic mechanism.
The present study also demonstrated the induction of nuclear apoptosis by fluorochrome-labeled Annexin V/PI using flow cytometry. Phosphatidylserine (PS) is found in the inner membrane of viable cells and is translocated to the outer membrane upon apoptosis induction. Annexin V has a high affinity for PS, and thereby flow cytometry analysis helps detect the quantity of viable and apoptotic cells (54). It is also an important tool in the evaluation of cancer drugs. In the current study, the untreated cells showed maximum viable cells, while NR, DS and DiNar treatments exhibited fewer viable cells and more apoptotic cells. The percentage of apoptotic cells was higher in DiNar treatment group than in the case of NR and DS treatments.
Various stress signals, such as DNA damage and cell cycle aberration, activate the intrinsic pathway of apoptosis. It is regulated by pro-apoptotic and anti-apoptotic genes and stimulates the cascade of caspases leading to the destruction of such cells (49). The pathway is a paramount marker and a key event that should be tested to underscore the mechanism of any anticancer drug. The present study showed an upregulation of pro-apoptotic proteins and downregulation of anti-apoptotic proteins. DiNar treatment demonstrated higher expression of pro-apoptotic protein Bax compared with the individual treatments of NR and DS. The anti-apoptotic protein Bcl-2 expression was observed to be downregulated in all of the treated cells, but the combined treatment DiNar showed the most significant downregulation in both HCT116 and SW480 cells. The expression of the tumor suppressor protein p53 was also found to be higher in DiNar treatment when compared with NR and DS. Furthermore, caspase-3, caspase-8 and caspase-9 expression were also higher in DiNar-treated HCT116 and SW480 cells.
RT-qPCR analysis of the expression of the key apoptosis-related genes was performed to strengthen the findings of the present study. Similar to other results of this study, RT-qPCR also demonstrated that the combined treatment DiNar enhanced the ability of cells to undergo apoptosis. The expression of the anti-apoptotic gene Bcl-2 was observed to be downregulated in treated cells, and DiNar showed higher downregulation of this gene. The expression levels of pro-apoptotic gene Bax, tumor suppressor gene p53, and caspase cascade genes, such as caspase-3, caspase-8 and caspase-9, were found to be significantly upregulated in the DiNar group. Thus, these results implied that apoptosis was notably higher in the cells treated with DiNar in both HCT116 and SW480 cell lines.
NF-κB plays a critical role in colon cancer as it activates most tumor-promoting cytokines. The activation of NF-κB has been found in almost half of all colon cancers (55). NF-κB activation promotes cell proliferation and instigates anti-apoptotic factors and inflammation (56). The present findings revealed that the expression levels of NF-κB, IKK-α and IKK-β were downregulated under NR, DS and DiNar treatments in both HCT116 and SW480 cell lines, but the downregulation was more pronounced in the case of DiNar. This effect was observed at both the protein and mRNA levels. Thus, the current study demonstrated that DiNar had a stronger anti-inflammatory effect and could prevent cell proliferation.
All the data taken together suggested that DiNar may inhibit cell proliferation, induce apoptosis and reduce inflammation of colon cancer cells more effectively than NR and DS. Thus, in conclusion, these findings indicted that DiNar can act as a more effective drug than the two compounds NR and DS taken individually. DiNar may also be able to serve as a safe chemotherapeutic option with fewer or no side effects when compared with the presently available drug options for colon cancer. Therefore, the combination has potential to be explored as the drug of choice against colon cancer.
Acknowledgements
The authors would like to acknowledge the Department of Science and Technology, Central Instrumentation Facility of Jamia Millia Islamia (New Delhi, India) for providing the instrumentation facility to carry out most of the high-end experiments.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
BZ, SN and MMAR conceived the present study. BZ performed the experiments and wrote the manuscript. SN and MMAR reviewed and edited the final manuscript. KUF provided technical support and contributed to data interpretation. BZ, LU and KI analyzed the data and validated the results. MMAR supervised the study. BZ and MMAR confirm the authenticity of all the raw data. All authors have read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A and Bray F: Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
American Cancer Society, . Colorectal Cancer: Facts and Figures 2021. American Cancer Society; 2021
|
|
3
|
Lee H, Choi DW, Cho YB, Yun SH, Kim HC, Lee WY, Heo JS, Choi SH, Jung KU and Chun HK: Recurrence pattern depends on the location of colon cancer in the patients with synchronous colorectal liver metastasis. Ann Surg Oncol. 21:1641–1646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Longley DB, Harkin DP and Johnston PG: 5-fluorouracil: Mechanisms of action and clinical strategies. Nat Rev Cancer. 3:330–338. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bleiberg H, Vandebroek A, Deleu I, Vergauwe P, Kalantari HR, Haens GD, Paesmans M, Peeters M, Efira A and Humblet Y: A phase II randomized study of combined infusion al leucovorin sodium and 5-FU versus the leucovorin calcium followed by 5-FU both in combination with irinotecan or oxaliplatinin patients with metastatic colorectal cancer. Acta Gastroenterol Belg. 75:14–21. 2012.PubMed/NCBI
|
|
6
|
Verhoeyen ME, Bovy A, Collins G, Muir S, Robinson S, de Vos CH and Colliver S: Increasing antioxidant levels in tomatoes through modification of the flavonoids biosynthetic pathway. J Exp Bot. 53:2099–2106. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mir IA and Tiku AB: Chemopreventive and therapeutic potential of ‘naringenin’, a flavanone present in citrus fruits. Nutr Cancer. 67:27–42. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Patel K, Singh GK and Patel DK: A review on pharmacological and analytical aspects of naringenin. Chin J Integr Med. 24:551–560. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bodduluru LN, Kasala ER, Madhana RM, Barua CC, Hussain MI, Haloi P and Borah P: Naringenin ameliorates inflammation and cell proliferation in benzo(a)pyrene induced pulmonary carcinogenesis by modulating CYP1A1, NFκB and PCNA expression. Int Immunopharmacol. 30:102–110. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park HJ, Choi YJ, Lee JH and Nam HJ: Naringenin causes ASK1-induced apoptosis via reactive oxygen species in human pancreatic cancer cells. Food Chem Toxicol. 99:1–8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Silambarasan T and Raja B: Diosmin, a bioflavonoid reverses alterations in blood pressure, nitric oxide, lipid peroxides and antioxidant status in DOCA-salt induced hypertensive rats. Eur J Pharmacol. 679:81–89. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Terao J, Piskula M and Yao Q: Protective effect of epicatechin gallate, and guercetin on lipid peroxidation in phospholipid bilayers. Arch Biochem Biophys. 308:278–284. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Naso L, Martínez VR, Lezama L, Salado C, Valcarcel M, Ferrer EG and Williams PAM: Antioxidant, anticancer activities and mechanistic studies of the flavone glycoside diosmin and its oxidovanadium(IV) complex. Interactions with bovine serum albumin. Bioorg Med Chem. 24:4108–4119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dung TD, Lin CH, Binh TV, Hsu HH, Su CC, Lin YM, Tsai CH, Tsai FJ, Kuo WW, Chen LM and Huang CY: Diosmin induces cell apoptosis through protein phosphatase 2A activation in HA22T human hepatocellular carcinoma cells and blocks tumour growth in xenografted nude mice. Food Chem. 132:2065–2073. 2012. View Article : Google Scholar
|
|
15
|
Thompson CB: Apoptosis in the pathogenesis and treatment of disease. Science. 267:1456–1462. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Attardi LD and Jacks T: The role of p53 in tumor suppression: Lessons from mouse models. Cell Mol Life Sci. 55:48–63. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lowe SW and Lin AW: Apoptosis in cancer. Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Terzic J, Grivennikov S, Karin E and Karin M: Inflammation and colon cancer. Gastroenterology. 138:2101–2114. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lakatos PL and Lakatos L: Risk for colorectal cancer in ulcerative colitis: Changes, causes and management strategies. World J Gastroenterol. 14:3937–3947. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Atreya I, Atreya R and Neurath MF: NF-kappaB in inflammatory bowel disease. J Intern Med. 263:591–596. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu H, Pardoll D and Jove R: STATs in cancer inflammation and immunity: A leading role for STAT3. Nat Rev Cancer. 9:798–809. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi X, Luo X, Chen T, Guo W, Liang C, Tang S and Mo J: Naringenin inhibits migration, invasion, induces apoptosis in human lung cancer cells and arrests tumour progression in vitro. J Cell Mol Med. 25:2563–2571. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dung TC, Lin CH, Binh TV, Hsu HH, Su CC, Lin YM, Tsai CH, Tsai FJ, Kuo WW, Chen LM and Huang CY: Diosmin induces cell apoptosis through protein phosphatase 2A activation in HA22T human hepatocellular carcinoma cells and blocks tumour growth in xenografted nude mice. Food Chemistry. 132:2065–2073. 2012. View Article : Google Scholar
|
|
24
|
Khan N, Jajeh F, Eberhard EL, Miller DD, Albrecht DM, Van Doorn R, Hruby MD, Maresh ME, Clipson L, Mukhtar H and Halberg RB: Fisetin and 5-fluorouracil: Effective combination for PIK3CA-mutant colorectal cancer. Int J Cancer. 145:3022–3032. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu Z, Huang B, Liu J, Wu X, Luo N, Wang X, Zheng X and Pan X: Combinatorial anti-proliferative effects of tamoxifen and naringenin: The role of four estrogen receptor subtypes. Toxicology. 410:231–246. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang Y, Yuan K, Tang M, Yue J, Bao L, Wu S, Zhang Y, Li Y, Wang Y, Ou X, et al: Melatonin inhibiting the survival of human gastric cancer cells under ER stress involving autophagy and Ras-Raf-MAPK signaling. J Cell Mol Med. 25:1480–1492. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Morgan DM: Tetrazolium (MTT) assay for cellular viability and activity. Methods Mol Biol. 79:179–183. 1998.PubMed/NCBI
|
|
28
|
Chou TC and Talalay P: Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 22:27–55. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chou TC: Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol Rev. 58:621–681. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang N, Wu ZM, McGowan E, Shi J, Hong ZB, Ding CW, Xia P and Di W: Arsenic trioxide and cisplatin synergism increase cytotoxicity in human ovarian cancer cells: Therapeutic potential for ovarian cancer. Cancer Sci. 100:2459–2464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nafees S, Mehdi SH, Zafaryab M, Zeya B, Sarwar T and Rizvi MM: Synergistic interaction of Rutin and Silibilin on human colon cancer cell line. Arch Med Res. 49:226–234. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Irshad M, Mehdi SJ, Al-Fatlawi AA, Zafaryab M, Ali A and Ahmad I: Phytochemical Composition of Cassia fistula fruit extracts and its anticancer activity against human cancer cell lines. J Biol Act Prod Nat. 4:158–170. 2014.
|
|
33
|
Wu M, Lao Y, Xu N, Wang X, Tan H, Fu W, Lin Z and Xu H: Guttiferone K induces autophagy and sensitizes cancer cells to nutrient stress-induced cell death. Phytomedicine. 22:902–910. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu L, Fan J, Ai G, Liu J, Luo N, Li C and Cheng Z: Berberine in combination with cisplatin induces necroptosis and apoptosis in ovarian cancer cells. Biol Res. 52:372019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Saraste A and Pulkki K: Morphologic and biochemical hallmarks of apoptosis. Cardiovasc Res. 45:528–537. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hollman PC and Katan MB: Dietary flavonoids: Intake, health effects and bioavailability. Food Chem Toxicol. 37:937–942. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ren W, Qiao Z, Wang H, Zhu L and Zhang L: Flavonoids: Promising anticancer agents. Med Res Rev. 23:519–534. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mishra J, Dromund J, Quazi SH, Karanki SS, Shaw JJ, Chen B and Kumar N: Prospective of colon cancer treatments and scope for combinatorial approach to enhanced cancer cell apoptosis. Crit Rev Oncol Hematol. 86:232–250. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Key TJ: Fruit and vegetables and cancer risk. Br J Cancer. 104:6–11. 2004. View Article : Google Scholar
|
|
41
|
Park JH, Jin CY, Lee BK, Kim GY, Choi YH and Jeong YK: Naringenin induces apoptosis through downregulation of Akt and caspase-3 activation in human leukemia THP-1 cells. Food Chem Toxicol. 46:3684–3690. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Totta P, Acconcia F, Leone S, Cardillo I and Marino M: Mechanisms of Naringenin-induced apoptotic cascade in cancer cells: Involvement of estrogen receptor ?? and ?? Signalling. IUBMB Life. 56:491–499. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lewinska A, Grochala JA, Kwasniewicz E, Deregowska A and Wnuk M: Diosmin induced senescence, apoptosis and autophagy in breast cancer cells of different p53 status and ERK activity. Toxicol Lett. 265:117–130. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lewinska A, Siwak J, Rzezutek I and Wnuk M: Diosmin induces genotoxicity and apoptosis in DU145 prostate cancer cell line. Toxicol In Vitro. 29:417–425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lewin B, Cassimeris L and Lingappa VR: Cell, Jones & Bartlett Publishers; 2015
|
|
46
|
Stewart ZA, Westfall MD and Pietenpol JA: Cell cycle dysregulation and anticancer therapy. Trends Pharmacol Sci. 24:139–145. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mills CC, Kolb EA and Sampson VB: Development of chemotherapy with cell-cycle inhibitors for adult and pediatric cancer therapy. Cancer Res. 78:320–325. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Arul D and Subramanian P: Naringenin (Citrus Flavonone) induces growth inhibition, cell cycle arrest and apoptosis in human hepatocellular carcinoma cells. Pathol Oncol Res. 19:763–770. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Khan KH, Codesido MB and Molife LR: Cancer therapeutics: Targeting the apoptotic pathway. Crit Rev Oncol Hematol. 90:200–219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hanahan D and Weinberg RA: Hallmarks of cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kerr JF, Wyllie AH and Currie AR: Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer. 26:239–257. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Jurisicova A, Varmuza S and Casper RF: Programmed cell death and human embryo fragmentation. Mol Hum Reprod. 2:93–98. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ahamad MS, Siddiqui S, Jafri A, Ahmad S and Afzal M: Induction of apoptosis and antiproliferative activity of naringenin in human epidermoid carcinoma cell through ROS generation and cell cycle arrest. PLoS One. 9:e1100032014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kagan VE, Fabisiak JP, Shvedova AA, Tyurina YY, Tyurin VA, Schor NF and Kawai K: Oxidative signaling pathway for externalization of plasma membrane phosphatidylserine during apoptosis. FEBS Lett. 477:1–7. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kojima M, Morisaki T, Sasaki N, Nakano K, Mibu R, Tanaka M and Katano M: Increased nuclear factor-kB activation in human colorectal carcinoma and its correlation with tumor progression. Anticancer Res. 24:675–681. 2004.PubMed/NCBI
|
|
56
|
Naugler WE and Karin M: NF-kappaB and cancer-identifying targets and mechanisms. Curr Opin Genet Dev. 18:19–26. 2008. View Article : Google Scholar : PubMed/NCBI
|














