Introduction
Programmed cell death-ligand 1 (PD-L1) is an immune
checkpoint molecule expressed on tumor cells and tumor-infiltrating
immune cells, which is involved in the suppression of cancer
immunity (1). Anti-PD-L1 antibody
relieves T cell suppression by inhibiting the binding of PD-L1 to
programmed cell death protein 1 (PD-1) and B7.1 (also known as
CD80), which are receptors on effector T cells, and exerts
antitumor effects in various types of cancer (2). In a phase 3 OAK trial, atezolizumab
(anti-PD-L1 antibody) treatment prolonged overall survival compared
with docetaxel in previously treated patients with non-small cell
lung cancer, regardless of PD-L1 expression status (3). However, intratumor PD-L1 expression is
generally considered to enrich patients for whom anti-PD-L1/PD-1
therapy would most likely be efficacious, and tumors with the
immune-desert phenotype (low CD8-positive rate) also rarely respond
to anti-PD-L1/PD-1 therapy as a single agent (4). To expand the benefit of these
antibodies, numerous combination strategies, e.g. with bevacizumab,
chemotherapy and ipilimumab, have been extensively investigated
(5–8).
Vascular endothelial growth factor (VEGF) has been
reported to exert not only tumor angiogenesis-inducing activity,
but also immunosuppressive activity which can attenuate the
antitumor immunity elicited by anti-PD-L1/PD-1 therapy through
inhibition of dendritic cell (DC) maturation (9–12) and
accumulation of myeloid-derived suppressor cells (MDSCs) (13). It has been reported that VEGF
blockade may promote antitumor immunity by inhibiting the
accumulation of regulatory T-cells (Tregs) (14).
Therefore, the combination of anti-PD-L1/PD-1
antibody and anti-VEGF antibody has been actively investigated in
clinical studies of numerous types of cancer, such as non-small
cell lung cancer, hepatocellular carcinoma, ovarian cancer and
renal cell carcinoma (5,15,16). The
IMpower150 clinical trial conducted on non-squamous non-small cell
lung cancer demonstrated that the combination of atezolizumab plus
bevacizumab (anti-VEGF antibody) and chemotherapy markedly
prolonged the progression-free and overall survival of patients
with metastatic non-squamous non-small cell lung cancer (5). Although several possible mechanisms for
the combination of PD-L1/PD-1 and VEGF blockades have been reported
using anti-PD-L1/PD-1 blockade-sensitive models (17–19), to
the best of our knowledge, no studies have used a
PD-L1low and immune desert-like tumor model.
The present study investigated the efficacy and
mechanisms of an anti-PD-L1 and anti-VEGF combination in an
anti-PD-L1 insensitive OV2944-HM-1 (HM-1) mouse model with
PD-L1low and immune desert-like phenotypes.
Materials and methods
Cell lines and culture conditions
OV2944-HM-1 (HM-1) murine ovarian cancer cells were
purchased from RIKEN BioResource Center and maintained in MEM Alpha
(Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Bovogen
Biologicals Pty Ltd.) (20). Colon
38 murine colon cancer cells were obtained from the Japanese
Foundation for Cancer Research based on a Material Transfer
Agreement with the National Cancer Institute and were maintained in
RPMI-1640 (Merck KGaA) supplemented with 10% FBS. Both cell lines
were incubated with 5% CO2 at 37°C.
Animals
A total of 708 female 6–8-week-old B6C3F1 mice were
purchased from CLEA Japan, Inc. for the HM-1 model. A total of 80
female 7-week-old C57BL/6J mice were purchased from Charles River
Laboratories, Inc. for the Colon 38 model. All animals were housed
in a specific pathogen-free environment under controlled conditions
(temperature, 20–26°C; humidity, 35–75%; 12 h light/12 h dark
cycle), and were allowed to acclimate and recover from
shipping-related stress for 5 days or more prior to the study.
Chlorinated water and irradiated food were provided ad
libitum. The health of the mice was monitored by daily
observation. Mice at the time of tumor inoculation and at the time
of randomization were 6–11 weeks old and 8–12 weeks old,
respectively. The body weights of the B6C3F1 mice and C57BL/6J mice
at the time of randomization were 19.2-25.8 and 18.7-21.7 g,
respectively. After the experiments, all animals from which tumor
tissues were not obtained were euthanized by CO2
asphyxiation with a CO2 displacement rate of 20% of the
chamber volume per min, followed by cervical dislocation; and the
animals from which tumor tissues were obtained were euthanized by
exsanguination under 2.0-2.5% isoflurane inhalation anesthesia
using isoflurane inhalation solution (Pfizer, Inc.). Animal death
was confirmed by the loss of signs, such as response to toe pinch
and heartbeat. Finally, graying of the mucous membranes and rigor
mortis were confirmed. All animal experiments were reviewed and
approved by the Institutional Animal Care and Use Committee at
Chugai Pharmaceutical Co., Ltd. (approval nos. 15-114 and 17-059)
and were conducted between February 2017 and February 2019.
In vivo tumor growth inhibition
studies
HM-1 tumor cells (1×106 cells) in 100 µl
MEM Alpha (Thermo Fisher Scientific, Inc.) were subcutaneously
inoculated into the right flank of B6C3F1 mice. Colon 38 tumor
cells (5×106 cells) in 100 µl 50% Matrigel Growth Factor
Reduced Basement Membrane Matrix (Corning, Inc.)-RPMI-1640 (Merck
KGaA) were subcutaneously inoculated into the right flank of
C57BL/6J mice. Mice with established tumors were randomly allocated
to each treatment group (Day 1). The time intervals between tumor
inoculation and randomization were 9–16 and 14 days in HM-1 and
Colon 38 models, respectively. For treatment, anti-mouse PD-L1
monoclonal antibody (mAb; clone 6E11; provided by Genentech, Inc.,
not commercially available), which blocks the binding of both PD-L1
to PD-1 and PD-L1 to B7-1 (CD80) (21), and anti-mouse VEGF mAb (clone
B20-4.1.1; provided by Genentech, Inc., not commercially
available), were used. Optimized for recombinant production in
mammalian cells (22), B20-4.1.1 is
a variant of B20-4.1, an antibody that prevents both human VEGF and
mouse VEGF from binding VEGFR2 and VEGFR1 with high potency
(23). Anti-mouse PD-L1 mAb or mouse
IgG (SouthernBiotech) was administered intraperitoneally to the
mice at a dose of 5 mg/kg twice a week from Day 1. Anti-mouse VEGF
mAb or mouse IgG was administered intraperitoneally to the mice at
a dose of 10 mg/kg weekly from Day 1. For CD8 depletion, anti-mouse
CD8 mAb (clone 116-13.1; cat. no. BE0118; Bio X Cell) or Rat IgG
(cat. no. 55951; MP Biomedicals) was administered intraperitoneally
to the mice at a dose of 100 µg/mouse twice a week from 11 days
before randomization. For C-X-C motif chemokine receptor 3 (CXCR3)
blocking, anti-mouse CXCR3 mAb (clone CXCR3-173; cat. no. 126538;
BioLegend, Inc.) or hamster IgG (cat. no. 402020; BioLegend, Inc.)
was administered intraperitoneally to the mice at a dose of 100
µg/mouse twice a week from Day 1. The time intervals between tumor
inoculation and final tumor growth measurement were 14–35 and 36
days in HM-1 and Colon 38 models, respectively.
Tumor volume was measured twice a week. Tumor volume
was estimated using the following equation: Tumor volume =
ab2/2, where a and b are the tumor length and width
(a≥b), respectively.
Flow cytometry analysis
For the analysis of tumor-infiltrating lymphocytes,
tumor tissue was excised from control-treated mice and antitumor
agent-treated mice, and single-cell suspensions were obtained by
mincing tumors and homogenizing them by disruption and digestion
with a gentleMACS Octo Dissociator with Heaters (Wakenyaku Co.,
Ltd.) and a Tumor Dissociation Kit for mice (Miltenyi Biotec GmbH).
Single-cell suspensions were incubated with anti-mouse CD16/CD32
(Fcγ receptor) antibodies (2.4G2; Tonbo Biosciences, cat. no.
70-0161) and the fixable viability dye (FVD) eFluor 506 or FVD780
(Thermo Fisher Scientific, Inc., cat. no. 65-0866-14 or 65-0865-14,
respectively) at 4°C for 5 min, and stained with the following
anti-mouse monoclonal antibodies: PerCP-Cy5.5 anti-CD45 [30-F11;
cat. no. 550994; used for assays that did not involve anti-granzyme
B (Gzm) and anti-interferon (IFN)-γ], BV650 anti-CD8α (53–6.7; cat.
no. 563234; used for assays that did not involve anti-GzmB and
anti-IFN-γ), PE anti-CD8α (53–6.7; cat. no. 553033; used for assays
involving anti-GzmB and anti-IFN-γ), Alexa Fluor 674 anti-GzmB
(GB11; cat. no. 560212), BV711 anti-CD11c (HL3; cat. no. 563048),
PE-CF594 anti-F4/80 (T45-2342; cat. no. 565613), BV650 anti-CD4
(RM4-5; cat. no. 563747; used for assays that did not involve
anti-CXCR3), PE-Cy7 anti-CD4 (RM4-5; cat. no. 552775; used for
assays involving anti-CXCR3), BV510 anti-CD11b (M1/70; cat. no.
562950), PE-Cy7 anti-FasL (CD95; Jo2; cat. no. 557653), APC
anti-intercellular adhesion molecule-1 (ICAM-1, CD54; 3E2; cat. no.
561605) and BV605 anti-vascular cell adhesion molecule 1 (VCAM-1,
CD106; 429; cat. no. 745193) from BD Biosciences; FITC anti-major
histocompatibility complex (MHC) class I (H-2Kk; 36-7-5;
cat. no. 114905), PE-Cy7 anti-CD80 (16–10A1; cat. no. 104734),
BV605 anti-CD86 (GL1; cat. no. 105037) and PE anti-CD31 (390; cat.
no. 102408) from BioLegend, Inc.; PE-Cy5.5 anti-forkhead box P3
(Foxp3; FJK-16s; cat. no. 35-5773-82) and Alexa Fluor 700
anti-granulocyte-differentiation antigen (Ly-6G/Ly-6C (Gr-1);
RB6-8C5; cat. no. 56-5931-80) from Thermo Fisher Scientific, Inc.;
and FITC anti-CD45 (30-F11; cat. no. 35-0451; used for assays
involving anti-Gzm B and anti-IFN-γ) and APC anti-IFN-γ (XMG1.2;
cat. no. 20-7311) from Tonbo Biosciences. Appropriate conjugated
isotype-matched IgG was used as the control for each. Intracellular
cytokines were stained using a Foxp3/Transcription Factor Staining
Buffer Set (Thermo Fisher Scientific, Inc.). Cells were analyzed
using an LSRFortessa X-20 cell analyzer (BD Biosciences) and FlowJo
10 software (Tree Star, Inc.).
Immunohistochemistry
PD-L1 expression at baseline in tumor tissues was
evaluated by immunohistochemical staining using anti PD-L1 antibody
(goat anti-mouse B7-H1/PD-L1 polyclonal antibody; dilution,
1:4,000; cat. no. AF1019; R&D Systems, Inc.) as a primary
antibody. CD8α+ T cells in tumor tissues were evaluated
at baseline and on Day 8 by immunohistochemical staining using
anti-CD8 antibody [rat anti-mouse CD8 alpha monoclonal antibody
KT15; dilution, 1:500 (baseline) or 1:800 (Day 8); cat. no.
GTX76351; GeneTex, Inc.] as a primary antibody. Tumor samples were
collected on Day 1 without the drug treatment or on Day 8 with the
drug treatments. Fresh frozen blocks were prepared from the
collected tumors with optimal cutting temperature compound (O.C.T.
compound) at −78°C. Subsequently, 5-µm-thick sections from fresh
frozen tissues were fixed in 4% paraformaldehyde at 4°C for 10 min.
The endogenous peroxidase activity and endogenous non-specific
background were blocked with 0.3% hydrogen peroxide in methanol at
room temperature for 30 min. The tissue sections were incubated at
4°C overnight with anti PD-L1 antibody or anti CD8 antibody as the
primary antibody. Subsequently, the sections were incubated at room
temperature with the Universal Immuno-peroxidase Polymer reagent
(undiluted; N-Histofine® Simple Stain Mouse MAX-PO (G);
cat. no. 414351; Nichirei Bioscience, Inc.) for 15 min or the
Universal Immuno-peroxidase Polymer reagent (undiluted;
N-Histofine® Simple Stain Mouse MAX-PO(Rat); cat. no.
414311; Nichirei Bioscience, Inc.) for 30 min, respectively.
Staining was conducted at room temperature using
3,3-diaminobenzidine solution (DAB+, Liquid, 2-component system;
cat. no. K3468; Agilent Technologies, Inc.) for 5 min. All sections
were counterstained at room temperature with hematoxylin for 1–3
sec. Histological examination was performed under a light
microscope (Nikon ECLIPSE Ni; Nikon Corporation) in a blinded
manner. The evaluation was performed by an experienced pathologist.
Immunohistochemical scoring of CD8α+ T cells was carried
out using grades of 0-3: 0, none; 1, scattered cell infiltration
with 0 or 1 focal cell infiltration in a specimen; 2, scattered
cell infiltration with 2–4 focal cell infiltrations in a specimen;
and 3, diffuse cell infiltration or ≥5 focal cell infiltrations in
a specimen.
Mouse C-X-C motif chemokine ligand
(CXCL)9, CXCL10, CXCL11 and IFN-γ ELISA assay
Tumor tissues collected from mice and stored at
−80°C were homogenized with Cell Lysis buffer (Cell Signaling
Technology, Inc.) with Complete Protease Inhibitor Cocktail Tablets
and Complete Phosphatase Inhibitor Cocktail Tablets (both from
Roche Diagnostics). The homogenate was centrifuged at 9,100 × g at
4°C for 20 min. The resultant supernatant was used for the assays
as cell lysate. Protein concentration of the cell lysates was
quantified using a Direct Detect spectrometer (Merck KGaA). The
following manufacturers' kits were used for Mouse CXCL9/MIG
Quantikine ELISA Kit (cat. no. MCX900; R&D Systems, Inc.),
Mouse IP-10 (CXCL10) ELISA Kit (cat. no. BMS6018; Thermo Fisher
Scientific, Inc.), Mouse C-X-C motif chemokine 11 (CXCL11) ELISA
kit (cat. no. CSB-EL006241MO; Cusabio Technology LLC) and Mouse
IFN-γ Quantikine ELISA Kit (cat. no. MIF00; R&D Systems,
Inc.).
Immunohistochemistry and
quantification of microvessel density (MVD) in tumor tissues
MVD in tumor tissues was evaluated by
immunohistochemical staining of CD31. Tumor samples were collected
on Day 8. Fresh frozen blocks were prepared from the collected
tumors with O.C.T. compound at −78°C. Immunohistochemical staining
was conducted as described previously (24). In brief, immunohistochemical analysis
of CD31 was conducted using a Rat HRP-Polymer 1-Step (mouse
adsorbed) system (cat. no. BRR4016; Biocare Medical, LLC) according
to the manufacturer's protocols. As the primary antibody, rat
anti-mouse CD31 monoclonal antibody (clone MEC 13.3; dilution,
1:500; cat. no. 553370) was purchased from BD Biosciences. MVD (%)
was calculated from the ratio of the CD31-positive staining area to
the total observation area in the viable region. Positive staining
areas were calculated using imaging analysis software (Definiens
Tissue Studio; version 3.60; Definiens, Inc.).
Statistical analysis
The experiments were conducted twice and data from
the two experiments, which showed a similar trend, were pooled and
presented as the mean ± standard deviation. For comparisons between
two groups, data were analyzed using the Wilcoxon rank sum test.
P<0.05 was considered to indicate a statistically significant
difference. For multiple comparisons, data were analyzed with the
Wilcoxon rank sum test, and then the P-values were corrected using
the Holm-Bonferroni method. Corrected P-values <0.05 were
considered to indicate a statistically significant difference
(25). All statistical analyses were
conducted using JMP software (version 15; SAS Institute, Inc.).
Results
HM-1 model exhibits
PD-L1low and immune desert-like phenotypes
First, the HM-1 tumor was characterized at baseline
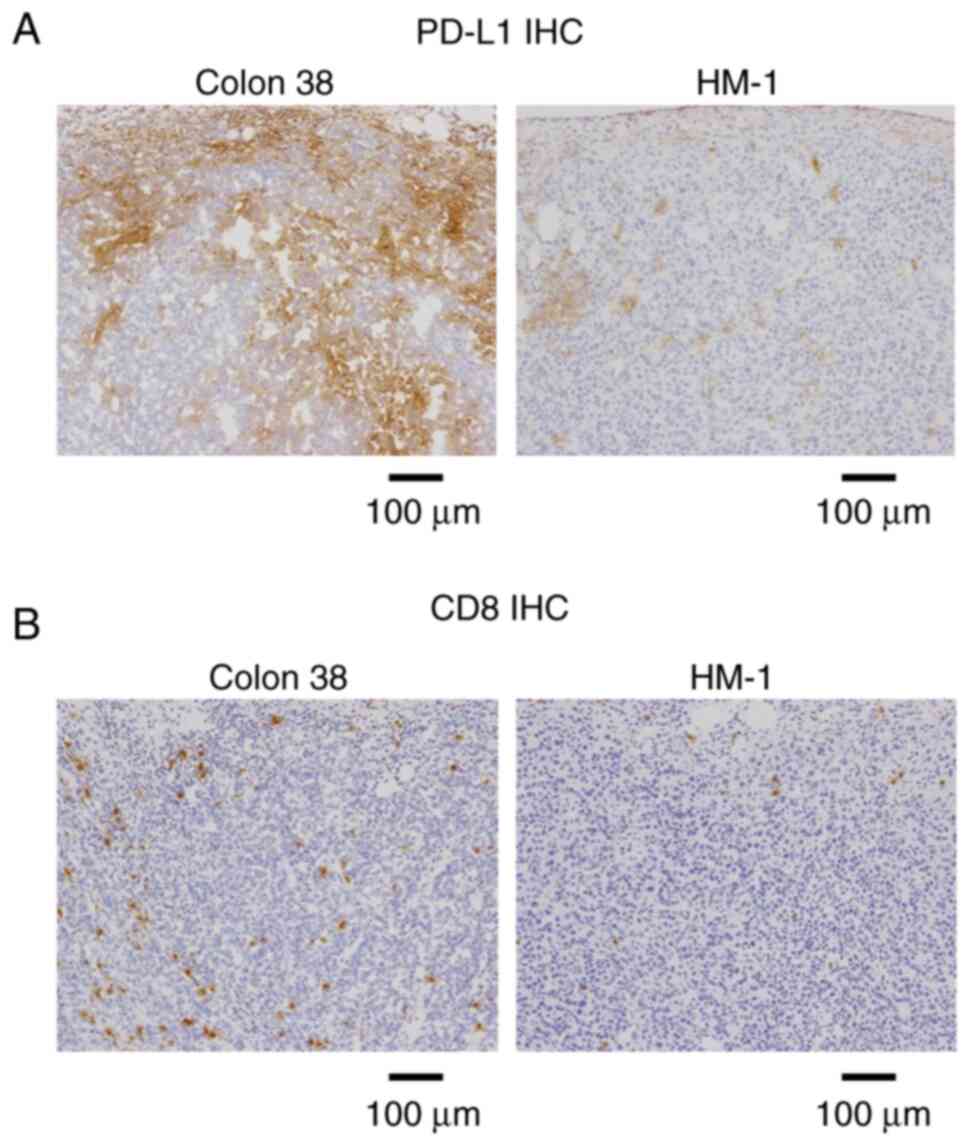
using immunohistochemistry. HM-1 tumors exhibited low PD-L1
expression, while Colon 38 tumors were PD-L1-positive (Fig. 1A). CD8+ T cells were
hardly observed in HM-1 tumors, whereas their infiltration was
prominent in Colon 38 tumors (Fig.
1B). This indicated that HM-1 tumors exhibited
PD-L1low and immune desert-like phenotypes.
Anti-PD-L1 antibody combined with
anti-VEGF antibody improves tumor control compared with anti-VEGF
antibody alone in an anti-PD-L1 insensitive HM-1 tumor model
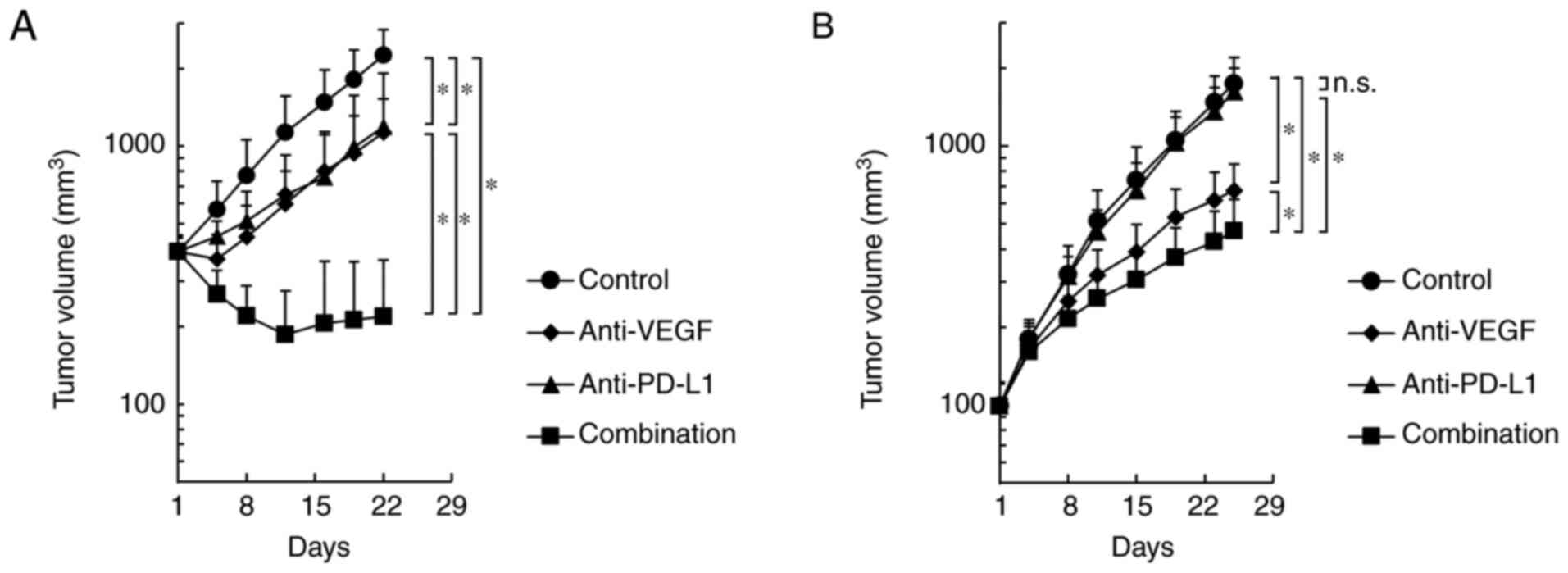
In the Colon 38 model, both anti-PD-L1 antibody
alone and anti-VEGF antibody alone significantly inhibited tumor
growth compared with the control, and combination efficacy of
anti-PD-L1 and anti-VEGF was shown (Fig.
2A). In the HM-1 model, the anti-VEGF antibody alone
significantly inhibited tumor growth compared with the control;
however, the anti-PD-L1 antibody alone did not significantly
inhibit the tumor growth compared with the control (Fig. 2B). Notably, the anti-PD-L1 antibody,
when combined with anti-VEGF, exhibited significantly stronger
antitumor efficacy compared with anti-VEGF antibody alone, even in
the anti-PD-L1 insensitive HM-1 model (Fig. 2B). Therefore, it was determined that
the combination of anti-PD-L1 antibody plus anti-VEGF antibody
exhibited more potent antitumor activity compared with single agent
treatments in not only the anti-PD-L1 sensitive model, but also in
the anti-PD-L1 insensitive model.
Higher percentage of effector
CD8+ T cells in the tumors treated with anti-PD-L1
antibody combined with anti-VEGF antibody in the HM-1 tumor
model
To investigate the effect of the anti-PD-L1 and
anti-VEGF combination treatment on immune status, the present study
analyzed the intratumoral status of CD8+ T cells on Day
8, when efficacy began to appear (Fig.
2B), using flow cytometry. No significant difference was
observed in the intratumoral percentage of CD8+ T cells
and GzmB+CD8+ T cells between the anti-PD-L1
antibody treatment and the control antibody treatment; however,
there was a significantly higher percentage of CD8+ T
cells and GzmB+CD8+ T cells in the anti-PD-L1
and anti-VEGF antibody combination treatment group compared with
the group treated with anti-VEGF antibody alone (Fig. 3A and B). Additionally,
immunohistochemical staining using an anti-CD8 antibody revealed
that the combination of anti-PD-L1 and anti-VEGF antibody induced a
significantly larger number of intratumoral CD8+ T cells
compared with the anti-VEGF antibody alone (Fig. 3C and D). On Day 4, compared with the
control treatment, anti-PD-L1 treatment both with and without
anti-VEGF treatment resulted in higher percentages of
CD8+ T cells and GzmB+CD8+ T cells
in tumor tissues in the HM-1 model (Fig. S1).
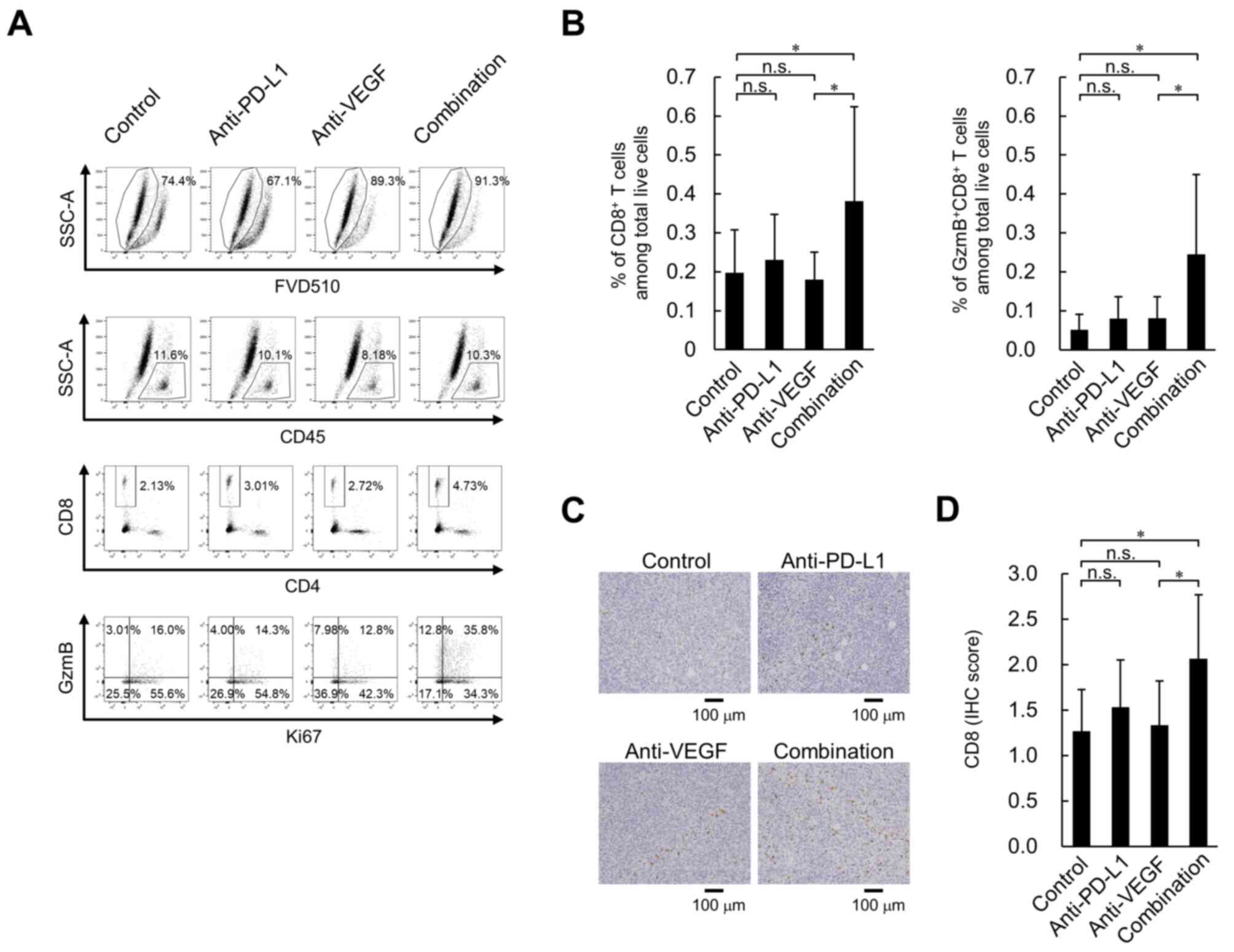 | Figure 3.Effect of anti-PD-L1 antibody in
combination with anti-VEGF antibody on percentage of
tumor-infiltrating CD8+ T cells in the OV2944-HM-1
tumors. (A) Representative flow cytometric profiles of
CD8+ T cells on Day 8. (B) Percentage of intratumoral
CD8+ T cells and GzmB+CD8+ T cells
on Day 8. These populations were determined using flow cytometry.
Data are presented as the mean + SD. CD8+ T cells;
control, n=15/group; anti-PD-L1, n=14/group; anti-VEGF, n=14/group;
combination, n=15/group. GzmB+CD8+ T cells;
n=15/group. (C) Tumor-infiltrating CD8+ T cells stained
immunohistochemically with anti-CD8 antibody on Day 8. Scale bar,
100 µm. (D) Levels of tumor-infiltrating CD8+ T cells
were indicated using IHC scores. Data are presented as the mean +
SD. n=15/group. *P<0.05 (Wilcoxon rank sum test with
Holm-Bonferroni correction). FVD510, fixable viability dye eFluor
510; GzmB, granzyme B; IHC, immunohistochemistry; n.s., no
significant difference; PD-L1, programmed death-ligand 1; SSC-A,
side scatter area; VEGF, vascular endothelial growth factor. |
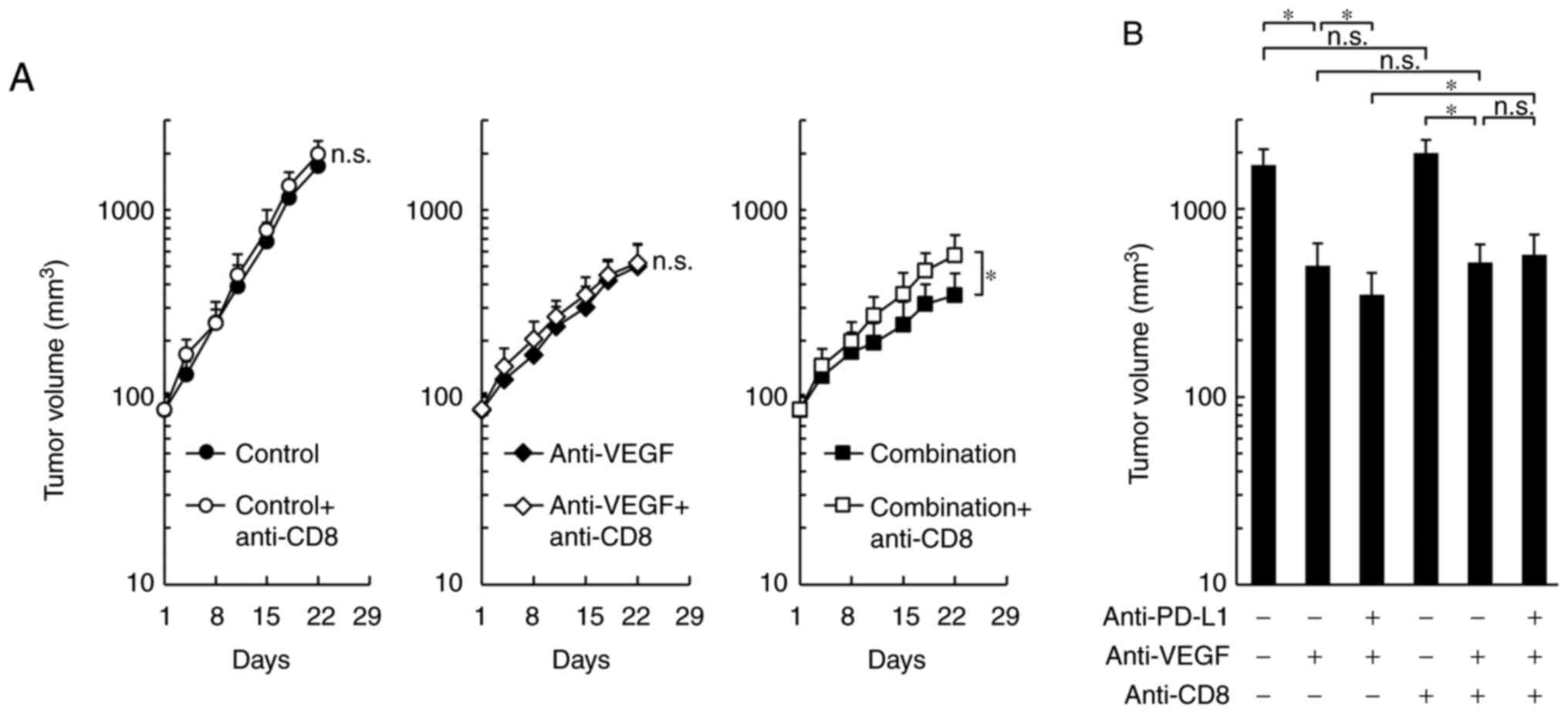
By co-administration of anti-CD8 depleting antibody,
the antitumor effects of the combination treatment were
significantly reduced to the same level as that of the anti-VEGF
antibody treatment alone, while control treatment and the anti-VEGF
antibody treatment groups were not affected (Fig. 4A and B). These results suggested that
the difference in the antitumor effect between combination
treatment and anti-VEGF treatment may be caused by CD8+
T cells.
Higher levels of intratumoral CXCR3
ligands in the combination treatment with anti-PD-L1 plus anti-VEGF
group in the HM-1 tumor model
To investigate the mechanism behind the increase in
intratumoral CD8+ T cells caused by the combination of
anti-PD-L1 and anti-VEGF, the present study focused on the CXCR3
ligands, CXCL9, CXCL10 and CXCL11, which have been reported to
recruit CD8+ T cells into tumors (26–28). On
Day 4, the protein levels of CXCL9 were significantly higher in the
anti-PD-L1 antibody alone treatment group compared with the control
treatment group (Fig. S2).
Anti-PD-L1 treatment alone also exhibited a tendency to increase
CXCL10, but this was not statistically significant. CXCL11 levels
were not affected by any of the treatments. On Day 8, no
significant difference was observed in the intratumoral protein
levels of CXCL9 between the anti-PD-L1 antibody treatment and
control treatment groups (Fig. 5A).
On the other hand, significantly higher levels were observed in the
anti-PD-L1 antibody treatment combined with the anti-VEGF antibody
group compared with the anti-VEGF antibody treatment alone group.
The combination treatment also exhibited a tendency to increase
CXCL10, but it was not statistically significant (Fig. S2). The protein expression levels of
intratumoral CXCL11 were not affected by any of the treatments.
Since CXCR3 ligands have been reported to be induced by IFN-γ
(28–30), the present study analyzed IFN-γ. On
Day 4, significantly higher levels of intratumoral IFN-γ expression
and percentages of IFN-γ+CD8+ T cells and
IFN-γ+FoxP3−CD4+ T cells in tumor
tissues were observed in the anti-PD-L1 antibody alone treatment
group compared with control treatment group (Fig. S3). On Day 8, no significant
difference was observed in these factors between the anti-PD-L1
antibody treatment and control treatment groups (Fig. S4). On the other hand, significantly
higher levels of these factors were observed in the anti-PD-L1
antibody treatment combined with the anti-VEGF antibody group
compared with the anti-VEGF antibody treatment alone group. When a
blocking antibody for CXCR3 was co-administered, the number of
intratumoral CD8+ T cells induced by the combination was
reduced to the same level as that of the control (Fig. 5B and C). This result suggested that
combination treatment promoted the trafficking of CD8+ T
cells into tumors via the CXCR3 ligands, mainly CXCL9.
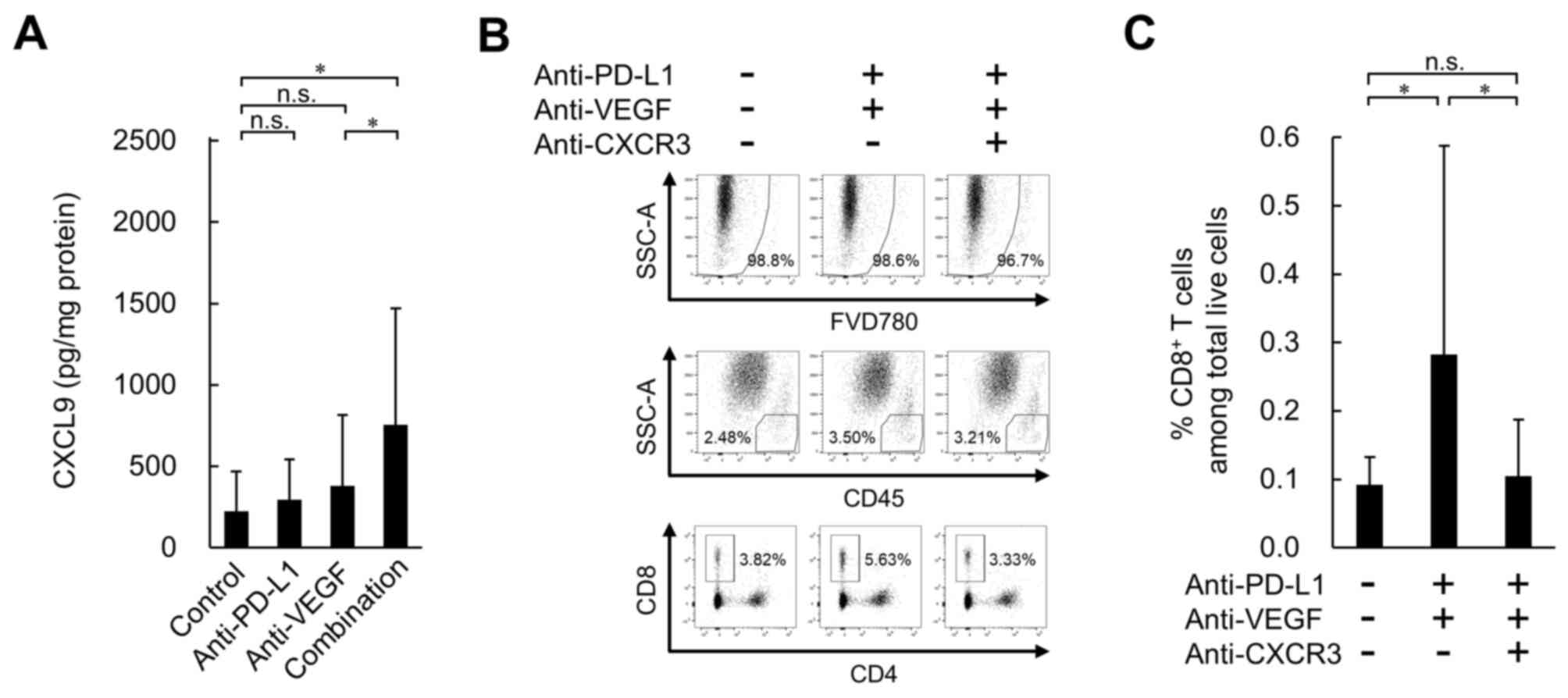 | Figure 5.Effect of anti-PD-L1 antibody in
combination with anti-VEGF antibody on expression levels of
intratumoral CXCL9, and effect of CXCR3 blocking on
tumor-infiltrating CD8+ T cells in the OV2944-HM-1
tumors. (A) Expression levels of intratumoral CXCL9 on Day 8
determined by ELISA. Data are presented as the mean + SD (control,
n=15/group; anti-PD-L1, n=15/group; anti-VEGF, n=14/group;
combination, n=15/group). (B) Representative flow cytometric
profiles of CD8+ T cells on Day 8. (C) Percentage of
intratumoral CD8+ T cells on Day 8. These populations
were determined by flow cytometry. Data are presented as the mean +
SD (n=16/group). *P<0.05 (Wilcoxon rank sum test with
Holm-Bonferroni correction). CXCL9, C-X-C motif chemokine ligand 9;
CXCR3, C-X-C motif chemokine receptor 3; FVD780, fixable viability
dye eFluor 780; n.s., no significant difference; PD-L1, programmed
death-ligand 1; SSC-A, side scatter area; VEGF, vascular
endothelial growth factor. |
Combination treatment with anti-PD-L1
plus anti-VEGF results in higher intratumoral MHC class I
expression on tumor cells compared with anti-VEGF treatment alone
in the HM-1 tumor model
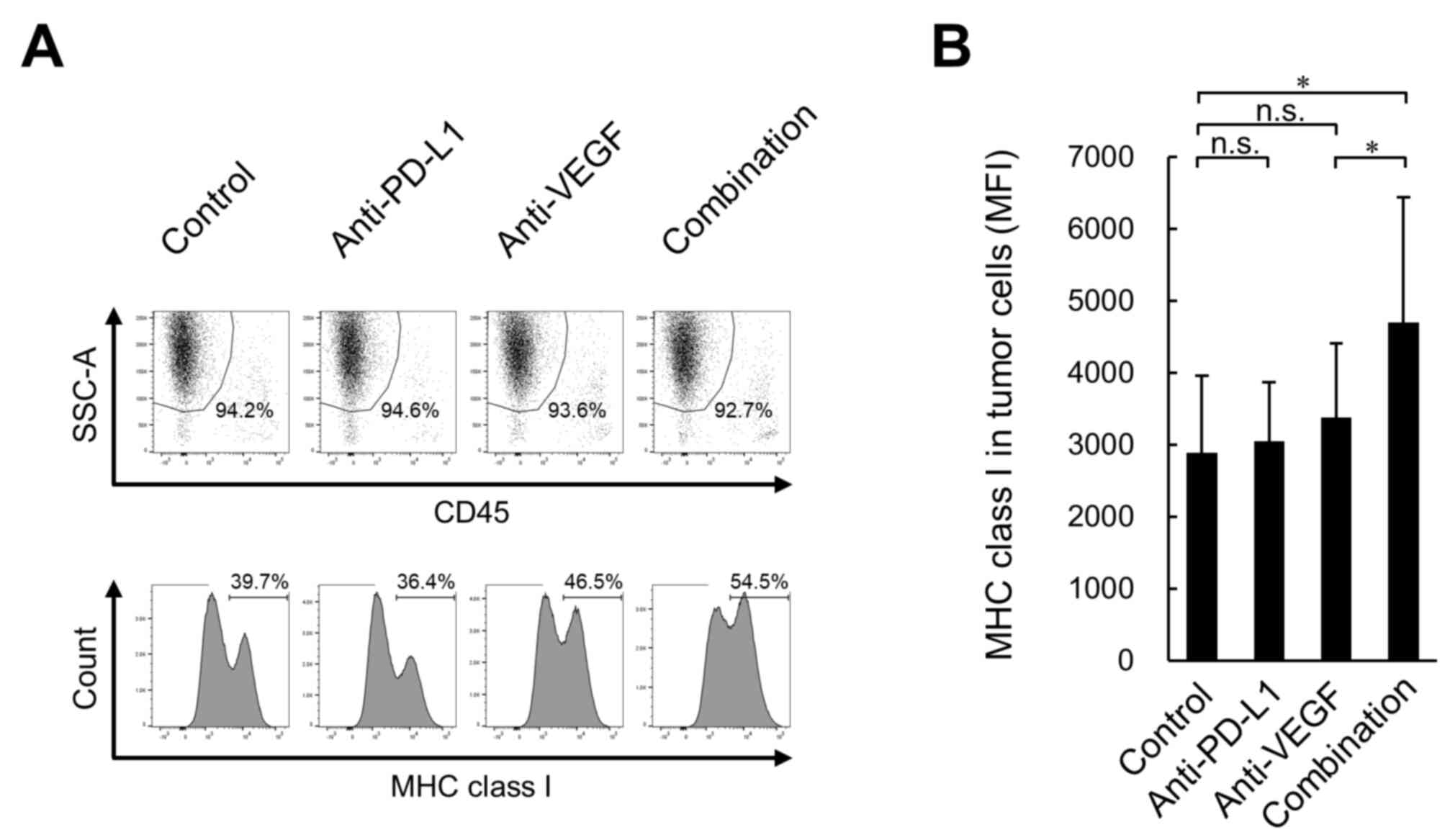
Furthermore, to investigate whether treated tumor
cells upregulate immune molecules implicated in antigen
presentation to CD8+ T cells, the present study analyzed
MHC class I (H-2Kk) protein expression in tumors.
Although no significant difference was observed in the expression
levels of H-2Kk on tumor cells (CD45−,
SSChigh) between the anti-PD-L1 antibody treatment and
the control treatment groups, significantly higher expression
levels were observed in the anti-PD-L1 antibody treatment combined
with anti-VEGF antibody group compared with the anti-VEGF antibody
treatment alone group (Fig. 6A and
B).
Combination treatment with anti-PD-L1
plus anti-VEGF does not affect DC maturation, MDSC and Treg
accumulation in tumors, or expression levels of ICAM-1, VCAM-1 and
FasL in vascular endothelial cells in the HM-1 model
To investigate the mechanisms of the anti-PD-L1 and
anti-VEGF combination treatment, the present study analyzed the
following factors: DC maturation, MDSC and Treg accumulation in
tumors, and expression levels of ICAM-1, VCAM-1 and FasL in
vascular endothelial cells. In the HM-1 model, these factors were
not affected by anti-PD-L1 or anti-VEGF either as monotreatment or
in combination, except that a significantly higher ratio of
CD8+ T cells to MDSCs was observed in the anti-PD-L1
antibody treatment combined with anti-VEGF antibody group compared
with the anti-VEGF antibody treatment alone group (Figs. S5–S7).
Anti-VEGF antibody suppresses MVD in
the HM-1 model
The present study analyzed MVD on Day 8. No
significant difference was observed in MVD in HM-1 tumor tissues on
Day 8 between the anti-PD-L1 antibody treatment and the control
treatment groups, and a significantly lower MVD was observed for
the anti-VEGF antibody treatment both with and without anti-PD-L1
treatment compared with the control treatment. No significant
difference was observed between the anti-VEGF antibody treatment
and the combination treatment groups (Fig. S8).
Discussion
Immune checkpoint inhibitors, such as
anti-PD-L1/anti-PD-1, are a standard treatment for patients with
several types of tumor; however, their remarkable efficacy may be
limited to patients with PD-L1-positive and immune-inflamed (high
CD8-positive rate) phenotypes (3,4). To
overcome this limitation, numerous clinical trials have evaluated
the efficacy of combinations with other agents, including
anti-VEGF, in numerous types of cancer, such as non-small cell lung
cancer, hepatocellular carcinoma and breast cancer (5,15,16).
Among these combinations, that of atezolizumab (anti-PD-L1
antibody) and bevacizumab (anti-VEGF antibody) plus chemotherapy
has been reported to have efficacy in the phase 3 IMpower150 trial
for non-squamous non-small cell lung cancer (5). Although several possible combination
mechanisms of anti-PD-L1/PD-1 and anti-VEGF have been reported
using preclinical models, all of these models were anti-PD-L1
sensitive (17,18). Therefore, the present study examined
the efficacy and mechanisms of the combination of anti-PD-L1
antibody and anti-VEGF antibody in the anti-PD-L1 insensitive HM-1
model. This model exhibited PD-L1low and immune
desert-like phenotypes, unlike the anti-PD-L1 sensitive Colon 38
model, which exhibited the PD-L1-positive and immune-inflamed like
phenotypes. Notably, anti-VEGF antibody triggered the efficacy of
anti-PD-L1 antibody in the anti-PD-L1 insensitive tumor model with
PD-L1low and immune desert-like phenotypes.
VEGF has been reported to exert immunosuppressive
activity through the following mechanisms: Inhibition of DC
maturation (9–12), accumulation of MDSCs in tumor
(13), and decreases of ICAM-1 and
VCAM-1 in vascular endothelial cells (31). VEGF blockade has been reported to
promote antitumor immunity through the inhibition of Treg
accumulation (14) and attenuation
of tumor endothelial FasL expression (32). In the HM-1 model with the anti-PD-L1
insensitive phenotype, these phenomena were not observed.
Additionally, VEGF has been investigated as a potential biomarker
of responses to immune checkpoint inhibitor therapies (33). In particular, it has been reported in
a pilot study that high pre-treatment serum VEGF levels in patients
with advanced melanoma may predict poor responses to ipilimumab
(anti-cytotoxic T-lymphocyte associated protein 4 antibody), while
it was not identified as a predictor of poor responses in patients
treated with pembrolizumab (anti-PD-1 antibody) alone or ipilimumab
plus nivolumab (anti-PD-1 antibody) (33). Further prospective clinical studies
with sufficient numbers of patients will be required to clarify the
utility of VEGF as a predictor in these therapies.
On Day 8, the intratumoral higher percentages of
CD8+ T cells and GzmB+CD8+ T cells
were observed only when anti-PD-L1 was combined with anti-VEGF, and
not without anti-VEGF. Since antitumor effects and MVD suppression
were observed with an anti-VEGF single agent and these antitumor
effects were not affected by CD8 depletion, it was suggested that
the anti-VEGF single agent exhibits antitumor effects through
anti-angiogenic activity in this model, not through CD8+
T cell-mediated immune enhancement. Since CD8 depletion reduced the
antitumor effect of the combination treatment to the level of the
anti-VEGF antibody treatment, it was considered that the difference
between the antitumor effect of the anti-VEGF single agent and that
of the combination treatment was due to the increase in
intratumoral CD8+ T cells caused by the combination
therapy.
The CXCR3 axis mainly positively regulates the
infiltration of CD8+ T cells into tumors (26–28). In
the HM-1 model, anti-PD-L1 treatment alone reportedly increases the
percentage of activated CD8+ T cells in lymph nodes but
not in tumors (34). This model
showed ~10-fold lower levels of intratumoral CXCR3 ligands compared
with the FM3A model, which also showed a PD-L1low and
immune desert-like phenotype but was sensitive to the anti-PD-L1
antibody (34). In a clinical study,
the untreated tumors of patients who would later respond to
atezolizumab exhibit elevated expression levels of IFN-γ and
IFN-γ-inducible genes, including CXCL9 (35). Therefore, in the HM-1 model, it is
possible that CD8+ T cells may not effectively
infiltrate the tumor from the blood because of low intratumoral
CXCR3 ligand expression, resulting in anti-PD-L1 insensitivity. In
this model, on Day 8, intratumoral CXCL9 expression was upregulated
by the anti-PD-L1 antibody combined with the anti-VEGF antibody,
but not without the anti-VEGF antibody. This upregulation was
associated with the higher percentage of intratumoral
CD8+ T cells. CXCL10 exhibited the same tendency, but
this was not statistically significant. The induction of an
intratumoral percentage of CD8+ T cells by the
combination treatment was reversed by CXCR3 blocking to the same
level as control. These results suggested that, in the HM-1 model,
the addition of the anti-VEGF antibody to the anti-PD-L1 antibody
activated the CXCR3 axis, inducing CD8+ T cell
infiltration into tumors. In addition to the CXCR3 axis, the CXCR5
axis has the potential to positively regulate the infiltration of
CD8+ T cells into tumors (36,37).
This is because CXCR5+CD8+ T cells have been
reported to strongly infiltrate colorectal and pancreatic tumors
and to exhibit strong cytotoxicity (36,37). In
a future study, it would be interesting to further investigate the
mechanistic role of this chemokine receptor and its ligands in the
CD8+ T cell infiltration of tumors as induced by the
combination of anti-PD-L1 antibody and anti-VEGF antibody.
IFN-γ is a pleiotropic molecule associated with
anti-proliferative, pro-apoptotic and antitumor mechanisms
(38). IFN-γ-induced intratumoral
expression of CXCR3 ligands has been reported to alter the local
distribution of T cells following immunotherapy (28–30).
Both higher levels of IFN-γ expression and higher percentages of
IFN-γ+CD8+ T cells and
IFN-γ+FoxP3−CD4+ T cells in tumor
tissue were observed in the combination of anti-PD-L1 antibody with
anti-VEGF antibody group; this was similar to the expression levels
of CXCR3 ligands and the percentages of CD8+ T cells.
This suggested that IFN-γ in tumor tissues induced the expression
levels of CXCR3 ligands, resulting in CD8+ T cell
infiltration in the HM-1 model; however, the mechanism by which
only the combination treatment was able to induce the higher levels
of IFN-γ and CXCR3 ligands in this model requires further
investigation. One possible explanation is that T cell
re-activation by anti-PD-L1 under anti-VEGF-induced hypoxic
conditions might contribute to increased IFN-γ-induced production
of CXCR3 ligands, which is based on a report revealing that
anti-VEGF induces hypoxia in tumors and that T cell activation
under hypoxic conditions induces IFN-γ secretion (39,40).
On Day 4, for anti-PD-L1 both with and without
anti-VEGF, higher percentages of CD8+ T cells and
GzmB+CD8+ T cells were observed in tumor
tissues in the HM-1 model. Notably, on Day 8, these higher
percentages were only observed when the anti-PD-L1 antibody was
combined with anti-VEGF antibody, but not without anti-VEGF
antibody. This indicated that, in the HM-1 model, the intratumoral
increase of activated effector T cells caused by the anti-PD-L1
single agent was transient, and was only maintainable in
combination with the anti-VEGF antibody. A transient increase with
anti-PD-L1 alone and a maintained increase with the anti-PD-L1
combined with anti-VEGF were also observed in the expression levels
of CXCR3 ligands and IFN-γ, and in the percentages of
IFN-γ+CD8+ T cells and
IFN-γ+FoxP3−CD4+ T cells in tumor
tissues. This may be why the anti-PD-L1 single agent failed to
exhibit antitumor activity, and why anti-PD-L1 exhibited extra
antitumor efficacy only when combined with anti-VEGF.
A number of cancer types acquire a phenotype of
reduced expression levels of MHC class I molecules to escape
recognition by cytotoxic T cells (41). In the present study, the combination
treatment with anti-PD-L1 antibody plus anti-VEGF antibody
upregulated the expression levels of MHC class I molecules on tumor
cells, which is another possible mechanism of immune enhancement,
and also maintained the accumulation of CD8+ T cells in
tumor tissues. This was consistent with a report revealing that
IFN-γ can upregulate MHC class I expression and enhance the
cytotoxic T cell-mediated immune response (42), as well as with results revealing
higher levels of IFN-γ expression for the combination treatment.
The combination treatment may increase the presentation of
tumor-antigens to specific cytotoxic CD8+ T cells.
In conclusion, to the best of our knowledge, this
was the first study to reveal that the addition of anti-VEGF
antibody overcomes anti-PD-L1 insensitivity in PD-L1low
and immune desert-like tumor models. This can be explained by the
increase of intratumoral CXCR3 ligands leading to the increased
infiltration of activated effector CD8+ T cells into
tumor tissues. When combined with anti-VEGF antibody, therapy with
anti-PD-L1 antibody is expected to exhibit more potent antitumor
efficacy even in anti-PD-L1 insensitive patients with
PD-L1low and immune desert-tumors.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Toshiki Iwai, Dr
Daiko Wakita and Dr Kazushige Mori for their helpful discussion and
advice, and Ms. Saki Otsuki, Ms. Masako Miyazaki, Ms. Hiromi
Sawamura and Ms. Ikuno Sugimoto for their excellent technical
assistance with the experiments (all Product Research Department,
Chugai Pharmaceutical Co., Ltd., Kamakura, Kanagawa 247-8530,
Japan).
Funding
All funding was provided by Chugai Pharmaceutical Co., Ltd.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NI, MS and OK conceived and designed the study. NI,
KY and MK acquired the data. NI, KY and MK analyzed and interpreted
the data. NI, KY, MS and OK were involved in writing, review and
revision of the manuscript. MS and OK supervised the study. NI and
MS confirmed the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were reviewed and approved by
the Institutional Animal Care and Use Committee at Chugai
Pharmaceutical Co., Ltd. (approval nos. 15-114 and 17-059;
Kamakura, Japan).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pardoll DM: The blockade of immune
checkpoints in cancer immunotherapy. Nat Rev Cancer. 12:252–264.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen DS and Mellman I: Oncology meets
immunology: The cancer-immunity cycle. Immunity. 39:1–10. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rittmeyer A, Barlesi F, Waterkamp D, Park
K, Ciardiello F, von Pawel J, Gadgeel SM, Hida T, Kowalski DM, Dols
MC, et al: Atezolizumab versus docetaxel in patients with
previously treated non-small-cell lung cancer (OAK): A phase 3,
open-label, multicentre randomised controlled trial. Lancet.
389:255–265. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen DS and Mellman I: Elements of cancer
immunity and the cancer-immune set point. Nature. 541:321–330.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Socinski MA, Jotte RM, Cappuzzo F, Orlandi
F, Stroyakovskiy D, Nogami N, Rodríguez-Abreu D, Moro-Sibilot D,
Thomas CA, Barlesi F, et al: Atezolizumab for first-line treatment
of metastatic nonsquamous NSCLC. N Engl J Med. 378:2288–2301. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
West H, McCleod M, Hussein M, Morabito A,
Rittmeyer A, Conter HJ, Kopp HG, Daniel D, McCune S, Mekhail T, et
al: Atezolizumab in combination with carboplatin plus
nab-paclitaxel chemotherapy compared with chemotherapy alone as
first-line treatment for metastatic non-squamous non-small-cell
lung cancer (IMpower130): A multicentre, randomised, open-label,
phase 3 trial. Lancet Oncol. 20:924–937. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gandhi L, Rodríguez-Abreu D, Gadgeel S,
Esteban E, Felip E, De Angelis F, Domine M, Clingan P, Hochmair MJ,
Powell SF, et al: Pembrolizumab plus chemotherapy in metastatic
non-small-cell lung cancer. N Engl J Med. 378:2078–2092. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hellmann MD, Paz-Ares L, Caro RB, Zurawski
B, Kim SW, Costa EC, Park K, Alexandru A, Lupinacci L, de la Mora
Jimenez E, et al: Nivolumab plus Ipilimumab in advanced
non-small-cell lung cancer. N Engl J Med. 381:2020–2031. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gabrilovich DI, Chen HL, Girgis KR,
Cunningham HT, Meny GM, Nadaf S, Kavanaugh D and Carbone DP:
Production of vascular endothelial growth factor by human tumors
inhibits the functional maturation of dendritic cells. Nat Med.
2:1096–1103. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gabrilovich D, Ishida T, Oyama T, Ran S,
Kravtsov V, Nadaf S and Carbone DP: Vascular endothelial growth
factor inhibits the development of dendritic cells and dramatically
affects the differentiation of multiple hematopoietic lineages in
vivo. Blood. 92:4150–4166. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oyama T, Ran S, Ishida T, Nadaf S, Kerr L,
Carbone DP and Gabrilovich DI: Vascular endothelial growth factor
affects dendritic cell maturation through the inhibition of nuclear
factor-kappa B activation in hemopoietic progenitor cells. J
Immunol. 160:1224–1232. 1998.PubMed/NCBI
|
|
12
|
Dikov MM, Ohm JE, Ray N, Tchekneva EE,
Burlison J, Moghanaki D, Nadaf S and Carbone DP: Differential roles
of vascular endothelial growth factor receptors 1 and 2 in
dendritic cell differentiation. J Immunol. 174:215–222. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Horikawa N, Abiko K, Matsumura N,
Hamanishi J, Baba T, Yamaguchi K, Yoshioka Y, Koshiyama M and
Konishi I: Expression of vascular endothelial growth factor in
ovarian cancer inhibits tumor immunity through the accumulation of
myeloid-derived suppressor cells. Clin Cancer Res. 23:587–599.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Terme M, Pernot S, Marcheteau E, Sandoval
F, Benhamouda N, Colussi O, Dubreuil O, Carpentier AF, Tartour E
and Taieb J: VEGFA-VEGFR pathway blockade inhibits tumor-induced
regulatory T-cell proliferation in colorectal cancer. Cancer Res.
73:539–549. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yi M, Jiao D, Qin S, Chu Q, Wu K and Li A:
Synergistic effect of immune checkpoint blockade and
anti-angiogenesis in cancer treatment. Mol Cancer. 18:602019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Finn RS, Qin S, Ikeda M, Galle PR, Ducreux
M, Kim TY, Kudo M, Breder V, Merle P, Kaseb AO, et al: Atezolizumab
plus bevacizumab in unresectable hepatocellular carcinoma. N Engl J
Med. 382:1894–1905. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meder L, Schuldt P, Thelen M, Schmitt A,
Dietlein F, Klein S, Borchmann S, Wennhold K, Vlasic I, Oberbeck S,
et al: Combined VEGF and PD-L1 blockade displays synergistic
treatment effects in an autochthonous mouse model of small cell
lung cancer. Cancer Res. 78:4270–4281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Chen Y, Li F, Bao L and Liu W:
Atezolizumab and bevacizumab attenuate cisplatin resistant ovarian
cancer cells progression synergistically via suppressing
epithelial-mesenchymal transition. Front Immunol. 10:8672019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kato Y, Tabata K, Kimura T,
Yachie-Kinoshita A, Ozawa Y, Yamada K, Ito J, Tachino S, Hori Y,
Matsuki M, et al: Lenvatinib plus anti-PD-1 antibody combination
treatment activates CD8+ T cells through reduction of
tumor-associated macrophage and activation of the interferon
pathway. PLoS One. 14:e02125132019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hashimoto M, Niwa O, Nitta Y, Takeichi M
and Yokoro K: Unstable expression of E-cadherin adhesion molecules
in metastatic ovarian tumor cells. Jpn J Cancer Res. 80:459–463.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oh SA, Wu DC, Cheung J, Navarro A, Xiong
H, Cubas R, Totpal K, Chiu H, Wu Y, Comps-Agrar L, et al: PD-L1
expression by dendritic cells is a key regulator of T-cell immunity
in cancer. Nat Cancer. 1:681–691. 2020. View Article : Google Scholar
|
|
22
|
Bagri A, Berry L, Gunter B, Singh M,
Kasman I, Damico LA, Xiang H, Schmidt M, Fuh G, Hollister B, et al:
Effects of anti-VEGF treatment duration on tumor growth, tumor
regrowth, and treatment efficacy. Clin Cancer Res. 16:3887–3900.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang WC, Wu X, Peale FV, Lee CV, Meng YG,
Gutierrez J, Fu L, Malik AK, Gerber HP, Ferrara N and Fuh G:
Cross-species vascular endothelial growth factor (VEGF)-blocking
antibodies completely inhibit the growth of human tumor xenografts
and measure the contribution of stromal VEGF. J Biol Chem.
281:951–961. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ishikura N, Yorozu K, Kurasawa M,
Yanagisawa M, Sugimoto M and Yamamoto K: Sustained effect of
continuous treatment with bevacizumab following bevacizumab in
combination with chemotherapy in a human ovarian clear cell
carcinoma xenograft model. Oncol Rep. 42:1057–1065. 2019.PubMed/NCBI
|
|
25
|
Holm S: A simple sequentially rejective
multiple test procedure. Scand J Statist. 6:65–70. 1979.
|
|
26
|
Tannenbaum CS, Tubbs R, Armstrong D, Finke
JH, Bukowski RM and Hamilton TA: The CXC chemokines IP-10 and mig
are necessary for IL-12-mediated regression of the mouse RENCA
tumor. J Immunol. 161:927–932. 1998.PubMed/NCBI
|
|
27
|
Hickman HD, Reynoso GV, Ngudiankama BF,
Cush SS, Gibbs J, Bennink JR and Yewdell JW: CXCR3 chemokine
receptor enables local CD8(+) T cell migration for the destruction
of virus-infected cells. Immunity. 42:524–537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tokunaga R, Zhang W, Naseem M, Puccini A,
Berger MD, Soni S, McSkane M, Baba H and Lenz HJ: CXCL9, CXCL10,
CXCL11/CXCR3 axis for immune activation - A target for novel cancer
therapy. Cancer Treat Rev. 63:40–47. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guirnalda P, Wood L, Goenka R, Crespo J
and Paterson Y: Interferon γ-induced intratumoral expression of
CXCL9 alters the local distribution of T cells following
immunotherapy with listeria monocytogenes. Oncoimmunology.
2:e257522013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gorbachev AV, Kobayashi H, Kudo D,
Tannenbaum CS, Finke JH, Shu S, Farber JM and Fairchild RL: CXC
chemokine ligand 9/monokine induced by IFN-gamma production by
tumor cells is critical for T cell-mediated suppression of
cutaneous tumors. J Immunol. 178:2278–2286. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Griffioen AW, Damen CA, Blijham GH and
Groenewegen G: Tumor angiogenesis is accompanied by a decreased
inflammatory response of tumor-associated endothelium. Blood.
88:667–673. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Motz GT, Santoro SP, Wang LP, Garrabrant
T, Lastra RR, Hagemann IS, Lal P, Feldman MD, Benencia F and Coukos
G: Tumor endothelium FasL establishes a selective immune barrier
promoting tolerance in tumors. Nat Med. 20:607–615. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Khattak MA, Abed A, Reid AL, McEvoy AC,
Millward M, Ziman M and Gray ES: Role of serum vascular endothelial
growth factor (VEGF) as a potential biomarker of response to immune
checkpoint inhibitor therapy in advanced melanoma: Results of a
pilot study. Front Oncol. 10:10412020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Iwai T, Sugimoto M, Patil NS, Bower D,
Suzuki M, Kato C, Yorozu K, Kurasawa M, Shames DS and Kondoh O:
Both T cell priming in lymph node and CXCR3-dependent migration are
the key events for predicting the response of atezolizumab. Sci
Rep. 11:139122021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Herbst RS, Soria JC, Kowanetz M, Fine GD,
Hamid O, Gordon MS, Sosman JA, McDermott DF, Powderly JD, Gettinger
SN, et al: Predictive correlates of response to the anti-PD-L1
antibody MPDL3280A in cancer patients. Nature. 515:563–567. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bai M, Zheng Y, Liu H, Su B, Zhan Y and He
H: CXCR5+ CD8+ T cells potently infiltrate
pancreatic tumors and present high functionality. Exp Cell Res.
361:39–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xing J, Zhang C, Yang X, Wang S, Wang Z,
Li X and Yu E: CXCR5+CD8+ T cells infiltrate
the colorectal tumors and nearby lymph nodes, and are associated
with enhanced IgG response in B cells. Exp Cell Res. 356:57–63.
2017.PubMed/NCBI
|
|
38
|
Castro F, Cardoso AP, Gonçalves RM, Serre
K and Oliveira MJ: Interferon-Gamma at the crossroads of tumor
immune surveillance or evasion. Front Immunol. 9:8472018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Roman J, Rangasamy T, Guo J, Sugunan S,
Meednu N, Packirisamy G, Shimoda LA, Golding A, Semenza G and
Georas SN: T-cell activation under hypoxic conditions enhances
IFN-gamma secretion. Am J Respir Cell Mol Biol. 42:123–128. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
De Almeida PE, Mak J, Hernandez G,
Jesudason R, Herault A, Javinal V, Borneo J, Kim JM and Walsh KB:
Anti-VEGF treatment enhances CD8+ T-cell antitumor
activity by amplifying hypoxia. Cancer Immunol Res. 8:8062020.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Garrido F and Algarra I: MHC antigens and
tumor escape from immune surveillance. Adv Cancer Res. 83:117–158.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rosa FM and Fellous M: Regulation of
HLA-DR gene by IFN-gamma. Transcriptional and post-transcriptional
control. J Immunol. 140:1660–1664. 1988.PubMed/NCBI
|