Introduction
Prostate cancer is the most common type of malignant
tumor in males and is the leading cause of cancer-related mortality
in numerous countries (1,2). At present, in 2021 there have been
~248,530 new cases and 34,130 deaths as a result of prostate cancer
in the United States alone (3). It
has also previously been determined that the incidence rate of
prostate cancer is significantly increased in the Asian population
(4). Due to advances in medical
oncology and improvements in surgical techniques, a variety of
cancers initially respond to treatment, but tumor progression,
recurrence and resistance to chemotherapy and radiotherapy may
still occur (5). Patients with
localized prostate cancer can be initially treated with surgery and
radiotherapy, and subsequently treated using androgen deprivation
therapy. However, in certain patients the disease eventually
becomes metastatic and resistant to treatment, which is referred to
as castration-resistant prostate cancer (CRPC).
Generally, the prognosis of patients with prostate
cancer depends on the depth of tumor infiltration, as well as the
appearance of lymph nodes and long-distance metastases, which can
be assessed by pathological microscopy (6). The poor prognosis associated with
prostate cancer has prompted the research and discovery of novel
diagnostic markers. However, the molecular mechanisms governing the
development and progression of prostate cancer remain unclear. It
is therefore crucial to elucidate new molecular approaches to
improve existing prognostics and supply relevant key clinical
insights into prognosis determination in patients with prostate
cancer.
Currently, researchers are focused on targeting the
cell cycle to exploit and block the proliferative capacity of
cancer cells. Progression through the phases of the cell cycle
relies on a complex network of proteins. Previous studies have
suggested that the abnormal expression of cell cycle regulatory
proteins may contribute to the development of cancer (7,8). The
cell division cycle-associated (CDCA) protein family (CDCA1-8)
consists of a group of proteins that serve a crucial role in
numerous biological processes, including the cell cycle.
Furthermore, it has been demonstrated that CDCAs may contribute
towards prostate cancer progression. A clinical trial that was
carried out on patients with CRPC determined that CDCA1 peptide
vaccination could induce peptide-specific cytotoxic T lymphocytes
(CTLs) in patients with CRPC (9).
It has also been demonstrated that CDCA2 is upregulated in patients
with prostate cancer, which regulates cell proliferation and is a
direct target of the hypoxia signaling pathway (10). Inhibition of CDCA6 induces prostate
cancer cell death, indicating CDCA6 may potentially have use as a
drug target in CRPC (11).
Furthermore, bioinformatics analysis has revealed that CDCA8 may
facilitate the tumorigenesis and progression of prostate cancer.
CDCA8 could therefore be a novel therapeutic target and biomarker
for the diagnosis and prognosis of prostate cancer (12). CDCA3 regulates cell cycle processes
and serves a role in triggering mitosis entry. CDCA3 is frequently
upregulated in tumor tissues and is associated with the oncogenic
properties of a variety of cancers, including colorectal cancer
(CRC) (13,14), gastric cancer (GC) (15), non-small cell lung cancer (NSCLC)
(16), oral squamous cell
carcinoma (OSCC) (17) and
pancreatic cancer (PAC) (18). It
has also been reported that CDCA3 acts as a downstream target gene
of HOXB3, which promotes prostate cancer progression (19).
The aim of the present study was to determine, using
microarray analysis, whether CDCA3 served as a hub gene in prostate
cancer progression and was associated with patient prognosis. It
was also investigated whether CDCA3 was differentially expressed in
prostate cancer and paracancerous tissues and if CDCA3 was
essential for cell proliferation, apoptosis and cell cycle
arrest.
Materials and methods
Microarray analysis
Gene expression profiling datasets for prostate
cancer were analyzed using the Gene Expression Omnibus (GEO;
http://www.ncbi.nlm.nih.gov/geo/geo2r/) database. In
the GSE27616 dataset (20), four
normal prostate and nine cancerous prostate samples were analyzed.
In the GSE3325 dataset (21), six
normal prostate and 13 cancerous prostate samples were analyzed.
For microarray data analysis, differentially expressed genes (DEGs)
were defined as genes whose expression differed between normal and
cancerous samples. The overlapping DEGs in the GSE27616 and GSE3325
datasets were identified to determine the genes involved with
tumorigenesis in prostate cancer. DEGs were screened using the
P-value (P<0.01) and fold change (FC; log |FC| >1). FunRich
software (version FunRich_3.1.3) was used to identify overlapping
DEGs (22). The upregulated and
downregulated genes were analyzed.
UALCAN (http://ualcan.path.uab.edu/analysis.html) is a web
portal based on level 3 RNA-seq and clinical data from 31 cancer
types in The Cancer Genome Atlas (TCGA) database (23). It helps analyze, integrate and
discover cancer transcriptomic data, and was used to analyze TCGA
gene expression information.
Identification of hub genes and
survival analysis
Search Tool for the Retrieval of Interacting Genes
(STRING) is a biological database (https://string-db.org) for constructing a
protein-protein interaction (PPI) network, providing a system-wide
view of interactions between each member. DEGs were used to
construct a STRING database, whereby an interaction with a combined
score of >0.4 was considered to be statistically significant.
Subsequently, a PPI network was established using Cytoscape
software (Cytoscape_v3.7.2) (22),
which visually explores biomolecular interaction networks composed
of proteins, genes and other molecules. CytoHubba in Cytoscape was
applied to screen the hub genes ranked by the MCC method (24).
Disease-free survival (DFS) of patients with
prostate cancer was analyzed using the Gene Expression Profiling
Interactive Analysis (GEPIA; http://gepia.cancer-pku.cn/detail/) database. Patients
with prostate cancer were divided into a low expression and high
expression group according to their median value of gene
expression. P<0.05 was set as the significance cut-off
level.
Tissue samples
The present study was approved (approval no.
xs2020ky014) by the Ethics Committee of Xishan People's Hospital of
Wuxi City (Wuxi, China). In total, seven prostate cancer samples
were collected to investigate CDCA3 protein expression. Both
cancerous and paracancerous tissue samples were collected from each
patient. All patients (mean age, 74 years; range, 68–79 years) were
treated with radical prostatectomy between January 2021 and July
2021, and consents were obtained orally. The inclusion criteria
were tissue samples collected from patients who had not undergone
androgen deprivation therapy, chemotherapy, radiotherapy, or other
auxiliary treatment prior to surgery. The exclusion criteria were
tissue samples collected from patients who had other severe
comorbidities.
Cell culture and in vitro
transfection
The human prostate cancer DU145 (serial cat. no.
TCHu222) and PC-3 (serial cat. no. TCHu158) cell lines were
purchased from The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences. CDCA3 protein levels were highly
expressed in DU145 and PC-3 cell lines as revealed by analyzing the
Harmonizome (https://maayanlab.cloud/Harmonizome/) database, which
was the reason for selecting these two cell lines for subsequent
analysis, and the cell line expression image is presented in
Fig. S1. Cells were cultured in
RPMI-1640 medium containing 10% FBS (both from Gibco; Thermo Fisher
Scientific, Inc.) in a humidified atmosphere at 37°C with 5%
CO2. To establish prostate cancer cell lines that
persistently repressed CDCA3 expression, CDCA3 short hairpin
(sh)RNA lentiviral constructs were purchased from Shanghai GeneChem
Co. Ltd. The CDCA3 shRNA nucleotide sequence was as follows:
5′-GCACGGACACCTATGAAGA-3′, which was confirmed the validation by a
previous study (14). The negative
control was a double-stranded shRNA without sequence homology to
any known human genes, and the sequence was as follows:
5′-UUCUCCGAACGUGUCACGUTT-3′. sh-CDCA3 and sh-negative control (NC)
were transduced as previously described (25).
Cell viability assay
DU145 and PC-3 cells were seeded at 200,000
cells/well into 6-well plates and cultured in RPMI-1640 medium
containing 10% FBS at 37°C with 5% CO2 overnight for
transfection. Prostate cancer cells were transduced with sh-CDCA3
or sh-NC at a multiplicity of infection of 50 and cultured with 6
µg/ml polybrene (GeneChem Co. Ltd) for 12 h in a humidified
atmosphere at 37°C with 5% CO2. Subsequently, the cells
were seeded at 2,000 cells/well in a 96-well plate. At 24–96 h
post-transfection, cell viability was detected using a Cell
Counting Kit-8 (CCK-8) assay kit (Beyotime Institute of
Biotechnology). Cells were incubated with 10 µl CCK-8 solution/well
for 1 h at 37°C according to the manufacturer's protocol. The
absorbance was measured at a wavelength of 450 nm. Cell viability
was measured across five wells in each group. All independent
treatments were carried out in three replicates.
Colony formation assay
DU145 and PC-3 cells in the logarithmic growth phase
were collected and 1,000 cells/well were seeded into a 6-well
plate. A total of 3 replicate wells were used for each group. Cells
were cultured in RPMI-1640 medium containing 10% FBS at 37°C with
5% CO2 for 10 days and when macroscopic colonies
appeared the culture solution was discarded. After washing with
PBS, the cells were fixed in 4% paraformaldehyde for 15 min at room
temperature and stained with 0.5% crystal violet solution for 15
min at room temperature. Colonies were observed using a light
microscope and the number of colonies was counted by visual
inspection. Images were captured using a Canon EOS600D digital
camera (Canon, Inc.). The minimum number of cells per colony was
50.
Apoptosis assay and cell cycle
analysis
The apoptotic rate and cell cycle of prostate cancer
cells were investigated using flow cytometry. DU145 and PC-3 cells
were trypsinized (without EDTA). For the cell cycle analysis, cells
were washed with PBS and subsequently incubated for 30 min at 37°C
in the dark with 500 µl PI (Beyotime Institute of Biotechnology).
Cells were then scanned using a CytoFLEX flow cytometer (Beckman
Coulter, Inc.). Cells were counted and the percentages of prostate
cancer cells in the three cycle phases were compared. For the cell
apoptosis analysis, cells were washed with PBS and were
subsequently cultured for 30 min at 37°C after adding 5 µl Annexin
V-phycoerythrin and 10 µl 7-aminoactinomycin D (Hangzhou Multi
Sciences Biotech Co., Ltd.) to identify apoptotic and necroptotic
cells. Stained cells and their apoptotic rates were quantified
using the software CytExpert 2.4.0.28 (Beckman Coulter, Inc.).
Western blotting
Total protein was extracted using RIPA lysis buffer
(Beyotime Institute of Biotechnology). Protein concentration was
determined using a BCA assay (Beyotime Institute of Biotechnology).
Total protein (30 µg/lane) was separated using SDS-PAGE on a 6–12%
gel (Beyotime Institute of Biotechnology) and transferred onto a
PVDF membrane (MilliporeSigma). The membranes were blocked with 5%
skimmed milk at room temperature for 1 h and incubated overnight at
4°C with diluted primary antibodies. Subsequently, the membranes
were washed using TBS with 0.1% Tween-20 (TBST) three times and
incubated with HRP-conjugated secondary antibodies for 1 h at room
temperature. After being washed with TBST, the membranes were
visualized using electrochemiluminescence (ECL) kit (Beyotime
Institute of Biotechnology). GAPDH was used as the loading control.
The following antibodies were used: Mouse monoclonal anti-GAPDH
(1:2,000; cat. no. 33033M; BIOSS), rabbit polyclonal anti-CDCA3
(1:1,000; cat. no. YT0819), rabbit polyclonal anti-cleaved
caspase-3 (1:1,000; cat. no. YC0006), rabbit polyclonal
anti-pro-caspase-3 (1:1,000; cat. no. YT6113), and rabbit
polyclonal anti-cyclin-dependent kinase inhibitor 1 (p21; 1:1,000;
cat. no. YT3497; all from Immunoway Biotechnology Company), rabbit
polyclonal anti-cyclin D1 (1:1,000; cat. no. 0623R; BIOSS), mouse
monoclonal anti-NFκB-p65 (1:1,000; cat. no. YM311), rabbit
polyclonal anti-phosphorylated (p)-NFκB-p65 (1:1,000; cat. no.
YP0192), rabbit polyclonal anti-IKKα/β (1:1,000; cat. no. YT2302),
and rabbit polyclonal anti-NFκB-p105/p50 (1:1,000; cat. no. YT3101;
all from Immunoway Biotechnology Company), HRP-labeled goat
anti-rabbit secondary antibody (1:5,000; cat. no. 40295G-HRP;
BIOSS) and HRP-labeled goat anti-mouse secondary antibody (1:5,000;
cat. no. 0368G-HRP; BIOSS).
Statistical analysis
The softwares SPSS 17.0 (SPSS, Inc.) and ImageJ
1.8.0 (National Institutes of Health) were used to carry out
statistical analysis. Each value was acquired from at least three
independent experiments. Data are presented as the mean ± SD. A
two-tailed unpaired Student's t-test was used to analyze
statistical differences between two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Hub gene screening and survival
analysis
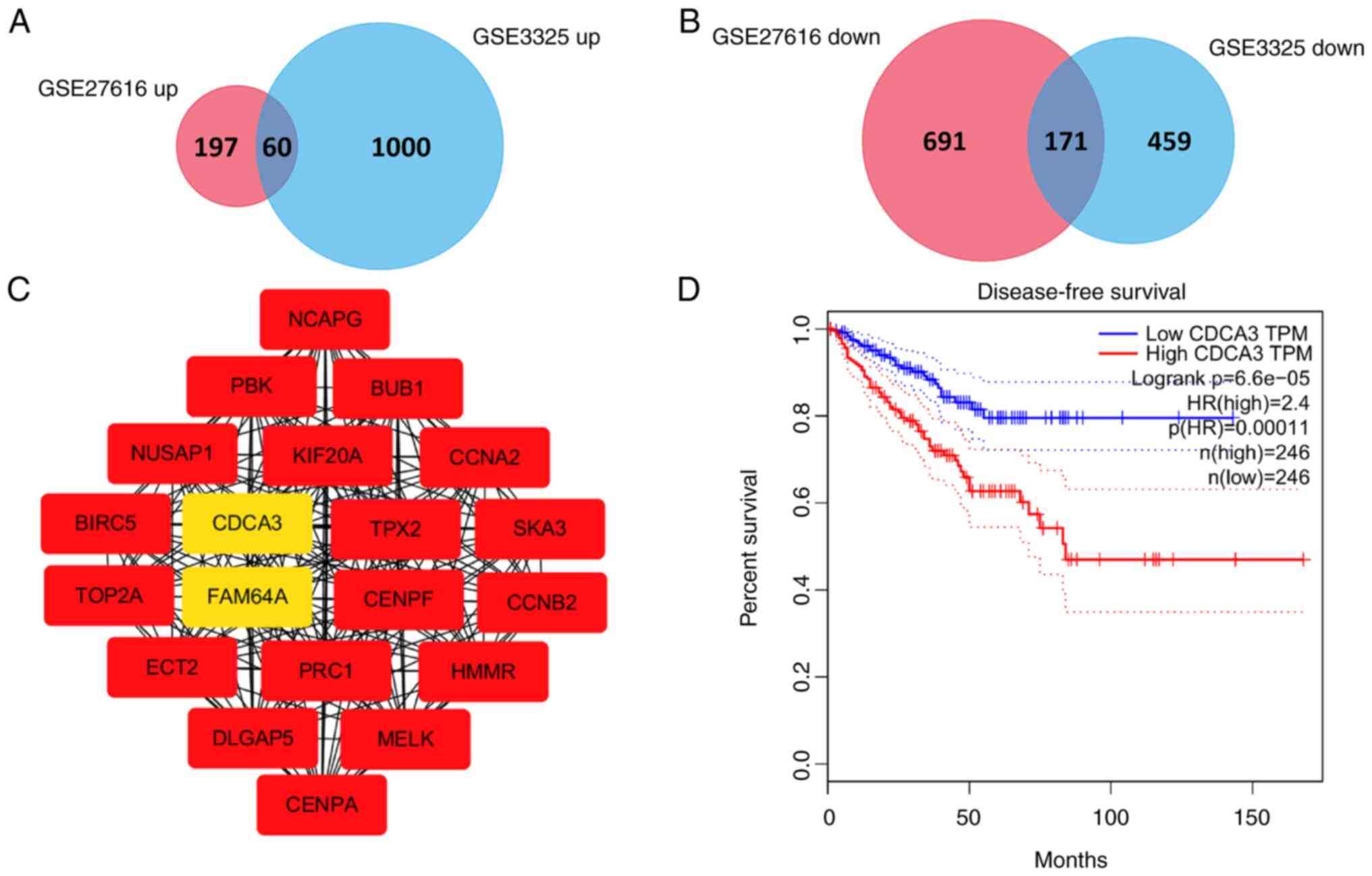
In total, 1,119 and 1,690 DEGs were identified via
the analysis of normal prostate vs. cancer tissues in the GSE27616
and GSE3325 datasets, respectively. A comparison of these two sets
of genes revealed 231 overlapping genes, including 60 upregulated
and 171 downregulated DEGs (Fig. 1A
and B). Subsequently the STRING database was used for
investigating and integrating the interactions between proteins.
The PPI network of overlapped DEGs between normal vs. cancer
tissues was constructed and data were exported for further analysis
in Cytoscape. The top 20 genes were identified as hub genes with a
score ≥1.22×1017, according to the MCC method. The PPI
network of these hub genes is presented in Fig. 1C. The identified hub genes were as
follows: Baculoviral IAP repeat containing 5 (BIRC5), DLG
associated protein 5 (DLGAP5), non-SMC condensing I complex subunit
G (NCAPG), cyclin A2 (CCNA2), cyclin B2 (CCNB2), maternal embryonic
leucine zipper kinase, targeting protein for Xklp2 (MELK), spindle
and kinetochore associated complex subunit 3 (SKA3), epithelial
cell transforming 2 (ECT2), hyaluronan mediated motility receptor,
centromere protein F (HMMR), protein regulator of cytokinesis 1
(PRC1), nucleolar and spindle-associated protein 1 (NUSAP1), DNA
topoisomerase II alpha (TOP2A), PDZ binding kinase (PBK),
centromere protein A (CENPA), kinesin family member 20A (KIF20A),
BUB1 mitotic checkpoint serine/threonine kinase (BUB1), family with
sequence similarity 64 member A (FAM64A) and CDCA3. All 20 hub
genes were expressed at elevated levels in prostate cancer compared
with normal prostate tissues and were associated with the DFS of
patients with prostate cancer, with the exception of ECT2. The DFS
curve (produced in GEPIA) for the gene of interest in the present
study, CDCA3, is presented in Fig.
1D. From these data it was indicated that prostate cancer
patients with low CDCA3 expression had a longer DFS and a better
prognosis than those with high CDCA3 expression (P<0.001).
CDCA3 overexpression in prostate
cancer tissues
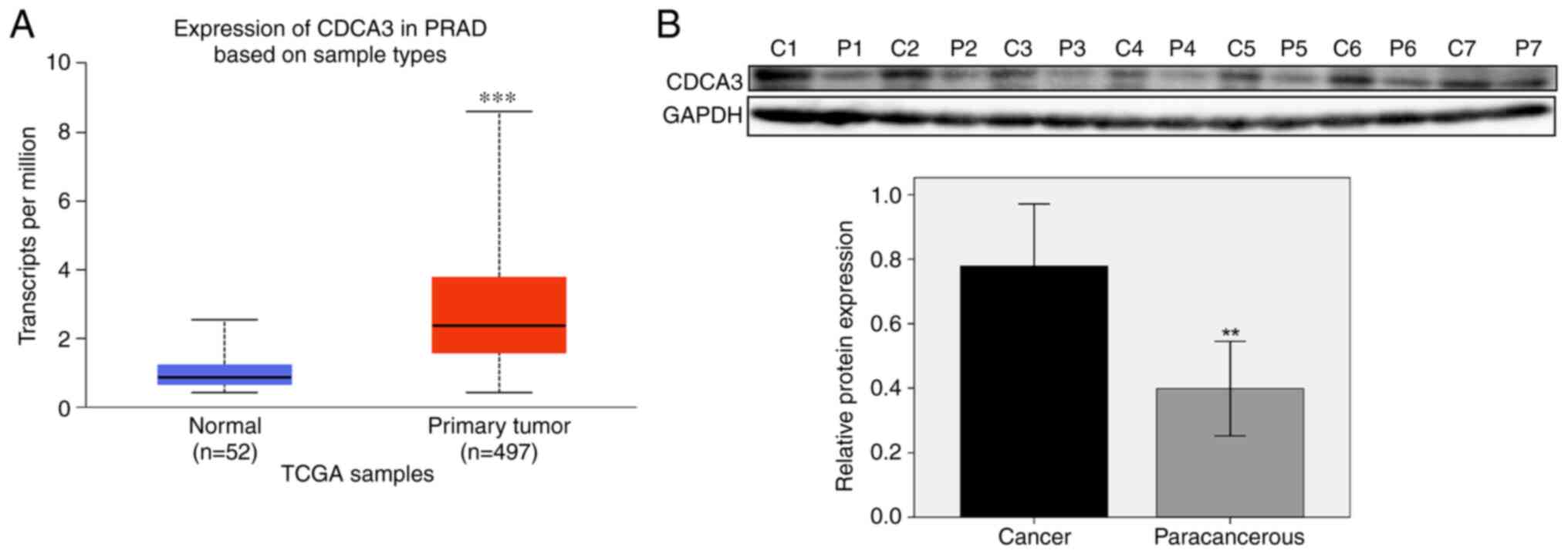
CDCA3 mRNA was demonstrated to be upregulated in
prostate cancer compared with normal prostate tissues, both in
GSE27616 and GSE3325 datasets. These results were also supported by
TCGA database. In total, 497 prostate cancer tissues and 52 normal
prostate samples were identified. CDCA3 was determined to be
significantly upregulated in prostate cancer compared with normal
tissues (median 2.372 vs. 0.856 transcripts per million;
P<0.001; Fig. 2A).
To verify the aforementioned results, prostate
cancer and paracancerous tissues were collected from seven patients
with prostate cancer at the Xishan People's Hospital of Wuxi city.
Proteins were extracted from the tissues and the CDCA3 protein
expression level was examined. The western blotting results
revealed that CDCA3 protein expression levels were significantly
higher in the prostate cancer tissue samples compared with the
paracancerous tissue samples (P<0.01; Fig. 2B).
CDCA3 knockdown inhibits cell
proliferation and induces apoptosis
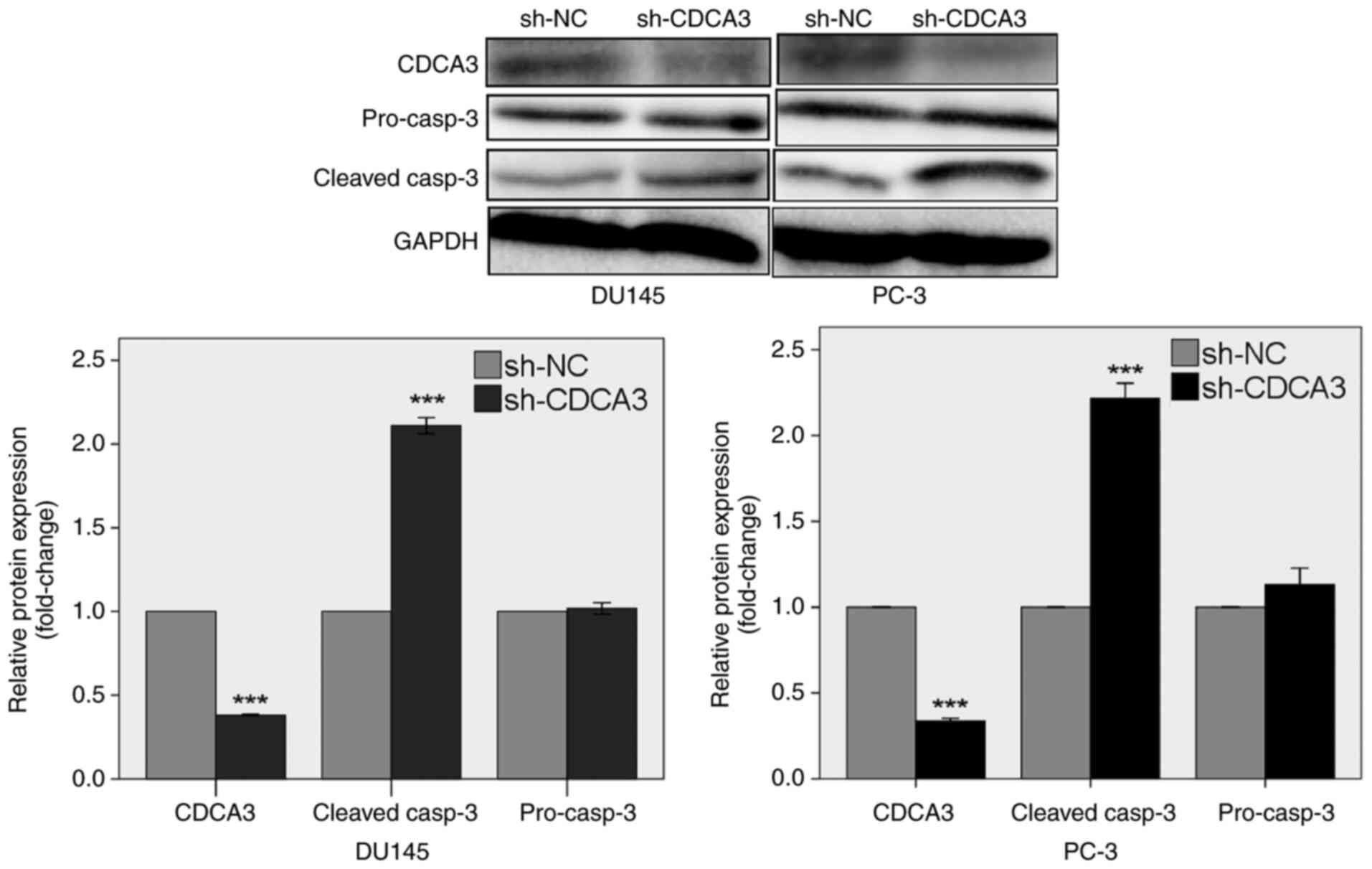
Western blotting was used to detect the inhibition
of CDCA3 protein expression following transduction with sh-CDCA3.
CDCA3 protein expression levels were suppressed by 61.9% in DU145
cells and 66.3% in PC-3 cells transduced with sh-CDCA3 compared
with the sh-NC transduced cells (P<0.001; Fig. 3). The results also demonstrated
that CDCA3 knockdown increased the protein expression levels of
cleaved caspase-3 by 2.1-fold in DU145 cells and 2.2-fold in PC-3
cells (P<0.001).
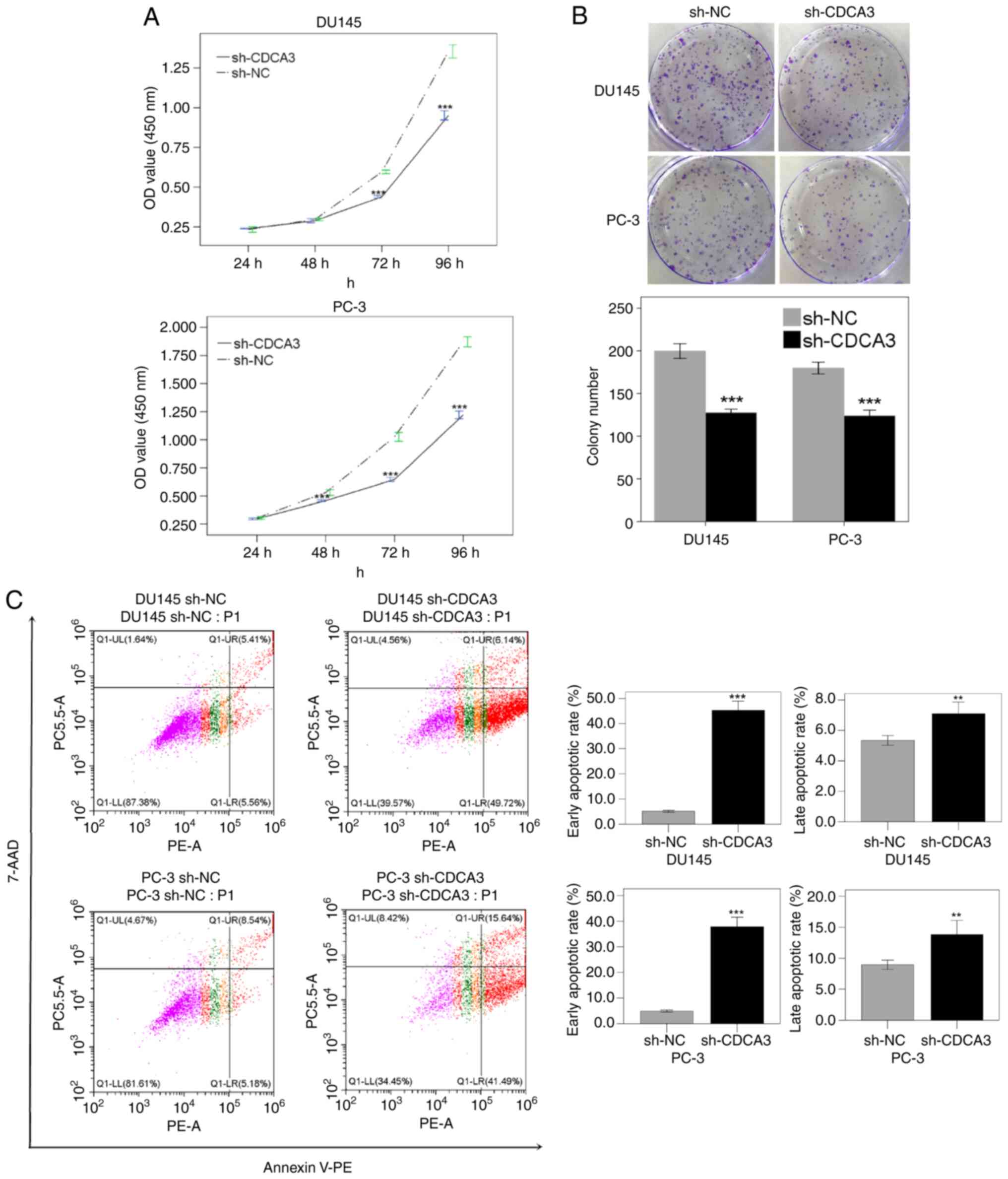
Subsequently, the effect of CDCA3 knockdown on DU145
and PC-3 cell proliferation was investigated using the CCK-8 and
colony formation assays. The CCK-8 assay results demonstrated that
knockdown of CDCA3 inhibited cell viability by 2.7% after 48 h
(P=0.28), 26.2% after 72 h (P<0.001), 29.7% after 96 h
(P<0.001) in DU145 cells, and 13.7% after 48 h (P<0.001),
37.0% after 72 h (P<0.001) and 34.7% after 96 h (P<0.001) in
PC-3 cells, compared with the sh-NC cell group (Fig. 4A). The downregulation of CDCA3 also
inhibited cell colony formation by 36.1% in DU145 cells
(P<0.005) and 31.0% in PC-3 cells (P<0.001; Fig. 4B).
The mechanism of CDCA3 in cell apoptosis regulation
was assessed using flow cytometry. The results demonstrated that
CDCA3 knockdown promoted early apoptosis both in DU145 (45.3% in
sh-CDCA3 vs. 5.1% in sh-NC; P<0.001) and PC-3 cells (37.8% in
sh-CDCA3 vs. 4.9% in sh-NC; P<0.001). The late apoptotic rate
was also increased in DU145 (7.1% in sh-CDCA3 vs. 5.3% in sh-NC;
P<0.005) and PC-3 cells (13.8% in sh-CDCA3 vs. 8.9% in sh-NC;
P<0.005; Fig. 4C).
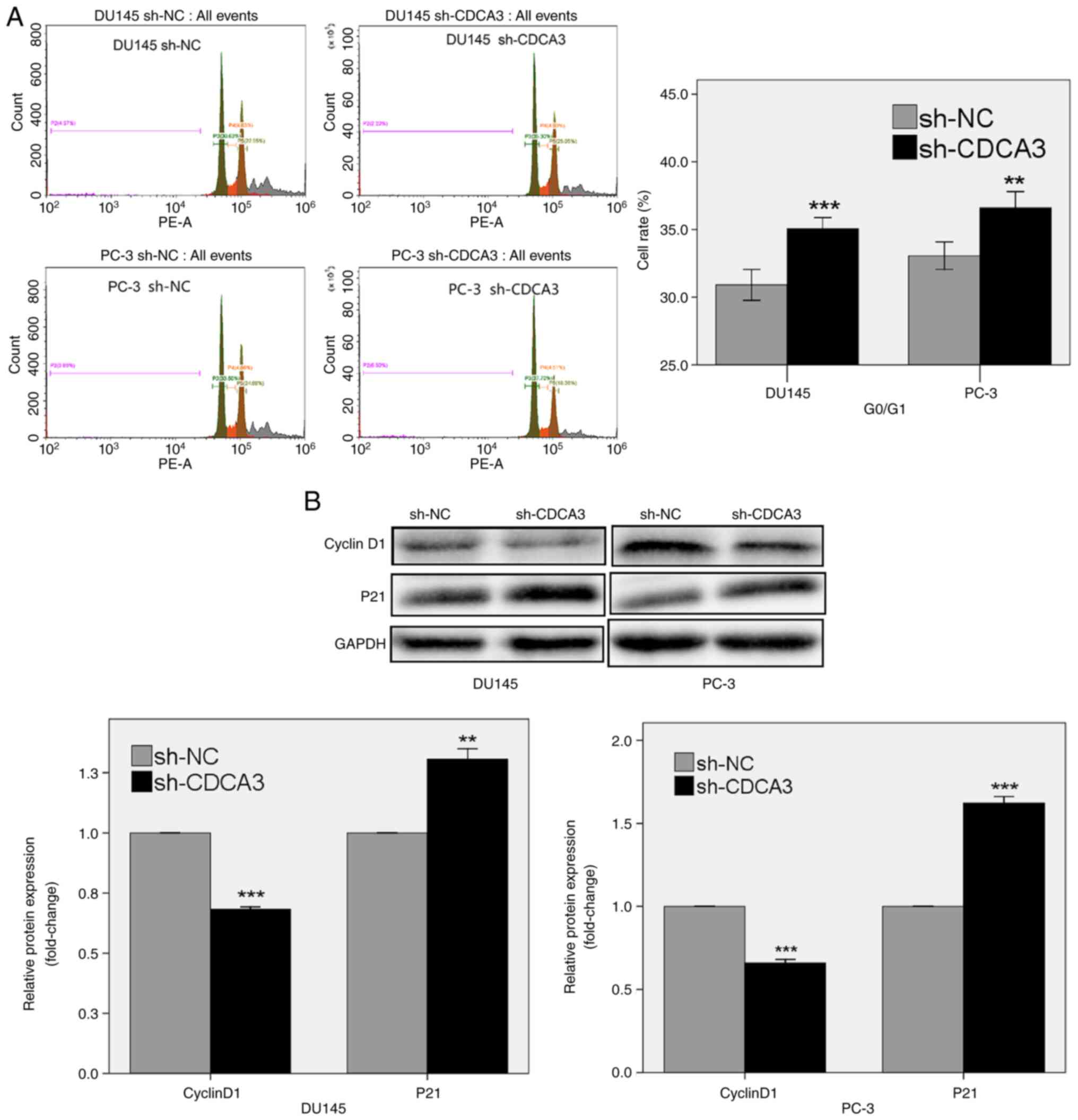
CDCA3 knockdown induces G0/G1 phase
arrest
The role of CDCA3 in modulating the prostate cancer
cell cycle was analyzed using flow cytometry. The results
demonstrated that CDCA3 knockdown induced G0/G1 phase arrest. The
percentage of cells in the G0/G1 phase in the sh-CDCA3 group was
higher compared with the sh-NC group, both in DU145 (35.06% in
sh-CDCA3 vs. 30.91% in sh-NC; P<0.001) and PC-3 cells (36.61% in
sh-CDCA3 vs. 33.06% in sh-NC; P<0.01; Fig. 5A).
Cyclin D1, a regulator of G1 phase progression,
serves a crucial role in carcinogenesis and cancer progression
(26). p21 also promotes cell
cycle arrest in response to a variety of stimuli (27). The results of the present study
demonstrated that CDCA3 knockdown in DU145 and PC-3 cells reduced
the protein expression levels of cyclin D1 and enhanced the protein
expression levels of p21 (P<0.01; Fig. 5B).
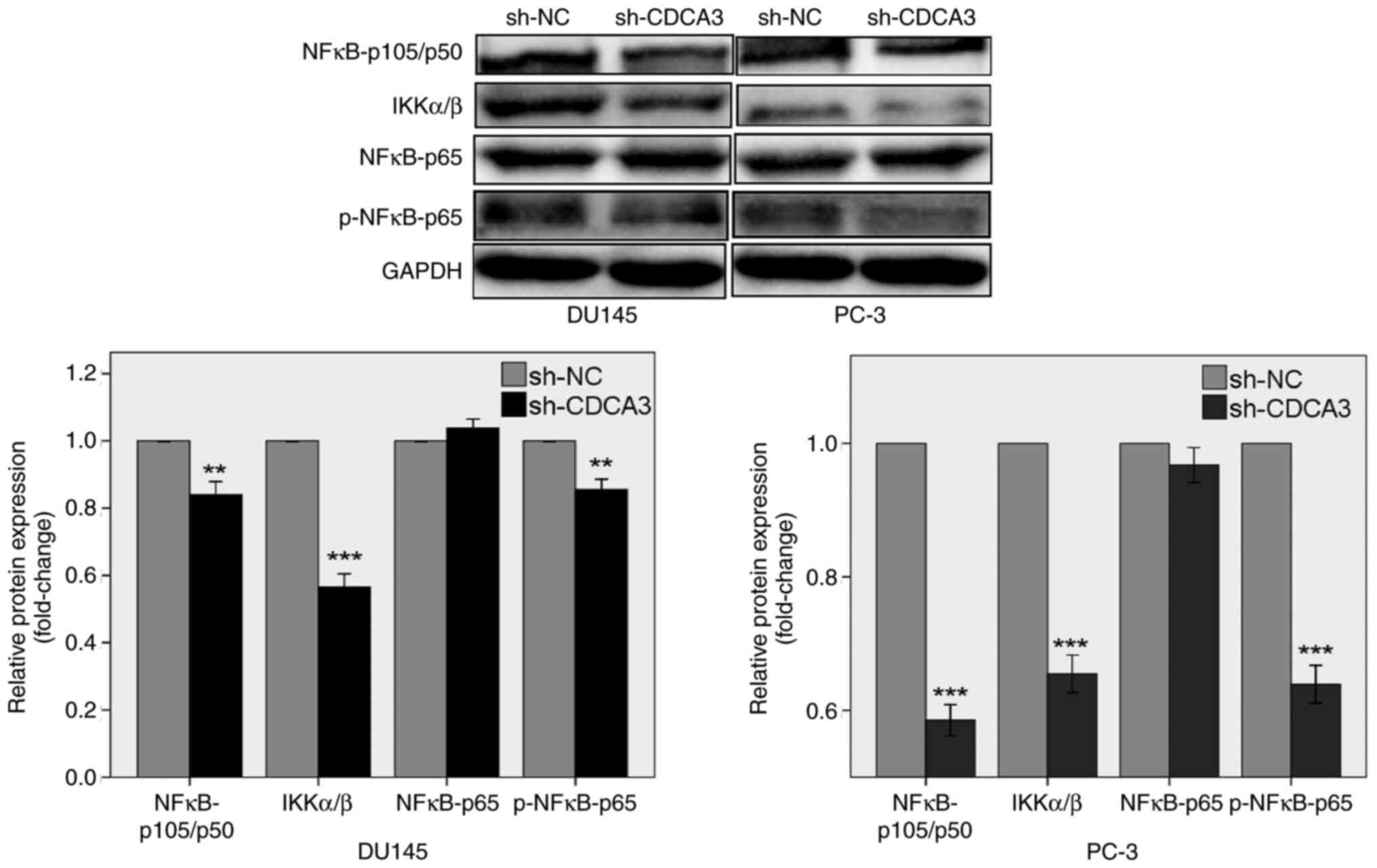
CDCA3 knockdown inhibits the NF-κB
signaling pathway
NF-κB activation is related to tumor initiation,
progression and metastasis in prostate cancer (28). A previous study reported that NF-κB
is constitutively activated in certain tumors, including in
prostate, breast, ovarian and pancreatic cancers (29). Overexpression of NF-κB in the
nucleus of prostate cancer cells is associated with a more
aggressive cancer phenotype, chemoresistance and metastasis
(28,29).
To discover the role of CDCA3 knockdown in the
regulation of the NF-κB signaling pathway, western blotting was
performed. The results demonstrated that CDCA3 knockdown suppressed
the protein expression levels of NFκB-p105/p50, IKKα/β and
p-NFκB-p65, both in DU145 and PC-3 cells (Fig. 6). CDCA3 knockdown had no effect on
the protein expression levels of NFκB-p65. These results indicated
that CDCA3 may potentially regulate the NF-κB signaling pathway in
prostate cancer DU145 and PC-3 cells.
Discussion
Abnormal cell division can lead to cancer.
Disturbance of cell cycle regulation is an important biological
feature exhibited in malignant tumors and can lead to reduced
apoptosis, unlimited proliferation and metastasis in malignant
cells (30). CDCA3 belongs to a
family of cell division-associated proteins, which function as part
of the SKP1-Cullin-RING-F-box ubiquitin ligase complex that
mediates the destruction of the mitosis-inhibitory kinase wee1
(14,16).
Previous studies have determined that CDCA3
expression levels are increased in tumor tissues and associated
with a poor patient prognosis in CRC, GC, NSCLC, OSCC and PAC
(13–18). In CRC, CDCA3 regulates E2F
transcription factor 1, thereby inhibiting the expression of p21.
Knockdown of CDCA3 results in G1/S-phase transition arrest via a
significant increase in p21 levels in SW480 cells (13). Furthermore, CDCA3 knockdown not
only induces cell cycle arrest but also promotes apoptosis via the
NF-κB signaling pathway, interacting with TNF receptor-associated
factor 2 (TRAF2) in CRC cells (14). In GC, CDCA3 knockdown inhibits cell
proliferation and induces cell cycle arrest in the G0/G1 phase via
mediating the Ras/ERK/MAPK axis (15). In NSCLC, depletion of CDCA3
expression reduces cell proliferation and causes abnormal
G2/M-phase cell cycle progression, exhibiting an upregulation in
p21 expression, independent of p53 (16). In OSCC, CDCA3 may be closely
associated with cancer progression by preventing a pause of
cell-cycle progression in the G1 phase via decreased expression of
cyclin-dependent kinase inhibitors (17). In PAC, downregulation of CDCA3
inhibits cell proliferation, promotes cell apoptosis and suppresses
in vivo tumor growth (18).
In addition, CDCA3, as a downstream target gene of numerous genes
and non-coding RNAs, promotes the progression of various tumors. In
bladder cancer (BC), kinesin family member 4A promotes the
development of BC via the transcriptional activation of CDCA3
expression (31). In prostate
cancer, HOXB3 binds to the CDCA3 promoter region and transactivates
CDCA3 expression, which upregulates CDCA3 expression and promotes
prostate cancer progression (19).
Knockdown of HOXB3 reduces the expression of CDCA3 in acute myeloid
leukemia, which decreases cell proliferation (32). In renal cell carcinoma, long
noncoding RNA, small nucleolar RNA host gene 12, promotes cell
proliferation, migration, invasion and sunitinib resistance via
CDCA3 (33).
Using the GEO database, the present study identified
20 hub genes that were expressed at elevated levels in prostate
cancer compared with normal prostate tissues. The transcriptomic
levels of CDCA3 were also significantly upregulated in prostate
cancer compared with normal tissues in TCGA database. It was
further verified by western blotting that CDCA3 protein expression
levels were upregulated in prostate cancer. Using the GEPIA
database, it was determined that high CDCA3 expression levels were
associated with a poor prognosis, as discussed for the numerous
other aforementioned tumors. The results also demonstrated that
knockdown of CDCA3 in DU145 and PC-3 cells inhibited cell
proliferation and facilitated early and late apoptosis with an
increase in the protein expression levels of cleaved caspase-3.
Regarding the molecular mechanism of CDCA3 in the
cell cycle, the NF-κB signaling pathway was further investigated.
NF-κB is a major regulator of numerous important cell processes,
such as inflammation, proliferation and apoptosis (34). Recent studies have demonstrated
that the NF-κB signaling pathway is important for prostate cancer
cell proliferation, invasion and the development of treatment
resistance (35,36). Activated NF-κB is translocated to
the nucleus and regulates gene transcription to activate a variety
of downstream targets such as cyclin D1, which is a cell cycle
regulatory protein. The activated NF-κB protein binds directly to
specific sequences in the cyclin D1 promoter, inducing the
upregulation of cyclin D1 expression levels (37). The results of the present study
have demonstrated that CDCA3 knockdown influenced cell cycle arrest
in the G0/G1 phase by decreasing cyclin D1 protein expression
levels. Furthermore, the protein expression levels of
NFκB-p105/p50, IKKα/β and p-NFκB-p65 were also reduced, while not
interacting with TRAF2 (data not shown). p21 is a crucial regulator
of cell cycle progression and serves an important role in
tumorigenesis and is regarded as a tumor suppressive protein.
Numerous studies have reported that the downregulation of p21 is
involved in a number of human cancers and is associated with cell
proliferation (38,39). p21 may also act independently or
with other cell cycle regulators, such as the CDK4/6 complex and
p53 (13). The present study
indicated that CDCA3 knockdown may upregulate p21 expression in the
cell cycle.
However, there are still many topics that remain to
be explored. Firstly, the effects of CDCA3 on the NF-κB signaling
pathway and the mechanisms involved need further investigation.
RNA-seq analysis will be performed to identify DEGs between
sh-CDCA3 and sh-NC in response to CDCA3 knockdown in DU145 and PC-3
cell lines, and the mechanisms of CDCA3 interacting with DEGs will
be investigated. Secondly, to further confirm the effects of CDCA3
in vivo, a tumor xenograft nude mouse model will be used in
a future study. Thirdly, a prostate cancer cell line with CDCA3
overexpression will be established to clarify whether CDCA3
overexpression has a consistent impact on S and G2/M phases, and
more experiments such as BrdU assay will be performed to aid in the
analysis of the cell cycle. Fourthly, more prostate cancer tissue
samples will be collected, and correlation between CDCA3 expression
and clinicopathological features in patients with prostate cancer
will be analyzed.
In conclusion, these data demonstrated that CDCA3
was upregulated in prostate cancer tissues and was associated with
a poor prognosis. The results indicated that knockdown of CDCA3
potentially suppresses prostate cancer progression via the
significant accumulation of p21 and via inhibiting the expression
of cyclin D1 by regulating the NF-κB signaling pathway.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and Technology
Development Guidance Plan (Medical and Health) Project of Wuxi
(grant no. CZ2020003), the Soft Science Research Project of Wuxi
Science and Technology Association (grant no. KX-21-C239) and the
Innovation Cultivation Fund Project of Xishan People's Hospital
(grant no. Yi202101).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DY, XH, and PG designed the study. PG and MZ carried
out the study, including data collection and data analysis. JZ
performed data analysis. PG wrote the manuscript. PG and MZ confirm
the authenticity of the raw data. All the authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved (approval no.
xs2020ky014) by the Ethics Committee of Xishan People's Hospital of
Wuxi City (Wuxi, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jin L, Zhou Y, Chen G, Dai G, Fu K, Yang D
and Zhu J: EZH2-TROAP pathway promotes prostate cancer progression
Via TWIST signals. Front Oncol. 10:5922392020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Attard G, Parker C, Eeles RA, Schröder F,
Tomlins SA, Tannock I, Drake CG and de Bono JS: Prostate cancer.
Lancet. 387:70–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD, Fuchs HE and Jemal
A: Cancer statistics, 2021. CA Cancer J Clin. 71:7–33. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wei B, Ruan J, Mi Y, Hu J, Zhang J, Wang
Z, Hu Q, Jiang H and Ding Q: Knockdown of TNF receptor-associated
factor 2 (TRAF2) modulates in vitro growth of TRAIL-treated
prostate cancer cells. Biomed Pharmacother. 93:462–469. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
De Angelis ML, Francescangeli F, La Torre
F and Zeuner A: Stem cell plasticity and dormancy in the
development of cancer therapy resistance. Front Oncol. 9:6262019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ahn JY, Hwang HS, Park YS, Kim HR, Jung
HY, Kim JH, Lee SE and Kim MA: Endoscopic and pathologic findings
associated with clinical outcomes of melanoma in the upper
gastrointestinal tract. Ann Surg Oncol. 21:2532–2539. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu ZH, Fang M and Zhou Y: Comprehensive
analysis of the expression and prognosis for CDCAs in head and neck
squamous cell carcinoma. PLoS One. 15:e02366782020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Phan NN, Wang CY, Li KL, Chen CF, Chiao
CC, Yu HG, Huang PL and Lin YC: Distinct expression of CDCA3,
CDCA5, and CDCA8 leads to shorter relapse free survival in breast
cancer patient. Oncotarget. 9:6977–6992. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Obara W, Sato F, Takeda K, Kato R, Kato Y,
Kanehira M, Takata R, Mimata H, Sugai T, Nakamura Y and Fujioka T:
Phase I clinical trial of cell division associated 1 (CDCA1)
peptide vaccination for castration resistant prostate cancer.
Cancer Sci. 108:1452–1457. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Cheng Y, Zhang Z, Bai Z, Jin H,
Guo X, Huang X, Li M, Wang M, Shu XS, et al: CDCA2 inhibits
apoptosis and promotes cell proliferation in prostate cancer and is
directly regulated by HIF-1α pathway. Front Oncol. 10:7252020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Clermont PL, Crea F, Chiang YT, Lin D,
Zhang A, Wang JZ, Parolia A, Wu R, Xue H, Wang Y, et al:
Identification of the epigenetic reader CBX2 as a potential drug
target in advanced prostate cancer. Clin Epigenetics. 8:162016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song Z, Huang Y, Zhao Y, Ruan H, Yang H,
Cao Q, Liu D, Zhang X and Chen K: The identification of potential
biomarkers and biological pathways in prostate cancer. J Cancer.
10:1398–1408. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qian W, Zhang Z, Peng W, Li J, Gu Q, Ji D,
Wang Q, Zhang Y, Ji B, Wang S, et al: CDCA3 mediates p21-dependent
proliferation by regulating E2F1 expression in colorectal cancer.
Int J Oncol. 53:2021–2033. 2018.PubMed/NCBI
|
|
14
|
Zhang W, Lu Y and Li X, Zhang J, Zheng L,
Zhang W, Lin C, Lin W and Li X: CDCA3 promotes cell proliferation
by activating the NF-κB/cyclin D1 signaling pathway in colorectal
cancer. Biochem Biophys Res Commun. 500:196–203. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Yin W, Cao W, Chen P, Bian L and
Ni Q: CDCA3 is a potential prognostic marker that promotes cell
proliferation in gastric cancer. Oncol Rep. 41:2471–2481.
2019.PubMed/NCBI
|
|
16
|
Adams MN, Burgess JT, He Y, Gately K,
Snell C, Zhang SD, Hooper JD, Richard DJ and O'Byrne KJ: Expression
of CDCA3 Is a prognostic biomarker and potential therapeutic target
in non-small cell lung cancer. J Thorac Oncol. 12:1071–1084. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Uchida F, Uzawa K, Kasamatsu A, Takatori
H, Sakamoto Y, Ogawara K, Shiiba M, Tanzawa H and Bukawa H:
Overexpression of cell cycle regulator CDCA3 promotes oral cancer
progression by enhancing cell proliferation with prevention of G1
phase arrest. BMC Cancer. 12:3212012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zou RC, Guo ZT, Wei D, Shi ZT, Ye ZC, Zhai
G, Zhong C, Tang B, Wang L and Ge JY: Downregulation of CDCA3
expression inhibits tumor formation in pancreatic cancer.
Neoplasma. 67:1223–1232. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen J, Zhu S, Jiang N, Shang Z, Quan C
and Niu Y: HoxB3 promotes prostate cancer cell progression by
transactivating CDCA3. Cancer Lett. 330:217–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu L, Guo K, Liang Z, Li F and Wang H:
Identification of candidate genes that may contribute to the
metastasis of prostate cancer by bioinformatics analysis. Oncol
Lett. 15:1220–1228. 2018.PubMed/NCBI
|
|
21
|
Guo L, Liu Y, Ding Z, Sun W and Yuan M:
Signal transduction by M3 muscarinic acetylcholine receptor in
prostate cancer. Oncol Lett. 11:385–392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gu P, Yang D, Zhu J, Zhang M and He X:
Bioinformatics analysis identified hub genes in prostate cancer
tumorigenesis and metastasis. Math Biosci Eng. 18:3180–3196. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chin CH, Chen SH, Wu HH, Ho CW, Ko MT and
Lin CY: cytoHubba: Identifying hub objects and sub-networks from
complex interactome. BMC Syst Biol. 8 (Suppl 4):S112014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou Y, Gu P, Li J, Li F, Zhu J, Gao P,
Zang Y, Wang Y, Shan Y and Yang D: Suppression of STIM1 inhibits
the migration and invasion of human prostate cancer cells and is
associated with PI3K/Akt signaling inactivation. Oncol Rep.
38:2629–2636. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qie S and Diehl JA: Cyclin D1, cancer
progression, and opportunities in cancer treatment. J Mol Med
(Berl). 94:1313–1326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma T, Chen H, Wang P, Yang N and Bao J:
Downregulation of lncRNA ZEB1-AS1 represses cell proliferation,
migration, and invasion through mediating PI3K/AKT/mTOR signaling
by miR-342-3p/CUL4B Axis in prostate cancer. Cancer Biother
Radiopharm. 35:661–672. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nguyen DP, Li J, Yadav SS and Tewari AK:
Recent insights into NF-κB signalling pathways and the link between
inflammation and prostate cancer. BJU Int. 114:168–176. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dominska K, Kowalska K, Matysiak ZE,
Pluciennik E, Ochedalski T and Piastowska-Ciesielska AW: Regulation
of mRNA gene expression of members of the NF-κB transcription
factor gene family by angiotensin II and relaxin 2 in normal and
cancer prostate cell lines. Mol Med Rep. 15:4352–4359. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ji J, Shen T, Li Y, Liu Y, Shang Z and Niu
Y: CDCA5 promotes the progression of prostate cancer by affecting
the ERK signalling pathway. Oncol Rep. 45:921–932. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zheng P, Wu K, Gao Z, Li H, Li W, Wang X,
Shi Z, Xiao F, Wang K, Li Z and Han Q: KIF4A promotes the
development of bladder cancer by transcriptionally activating the
expression of CDCA3. Int J Mol Med. 47:992021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bi L, Zhou B, Li H, He L, Wang C, Wang Z,
Zhu L, Chen M and Gao S: A novel miR-375-HOXB3-CDCA3/DNMT3B
regulatory circuitry contributes to leukemogenesis in acute myeloid
leukemia. BMC Cancer. 18:1822018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Y, Cheng G, Huang Z, Bao L, Liu J,
Wang C, Xiong Z, Zhou L, Xu T, Liu D, et al: Long noncoding RNA
SNHG12 promotes tumour progression and sunitinib resistance by
upregulating CDCA3 in renal cell carcinoma. Cell Death Dis.
11:5152020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jin M, Duan J, Liu W, Ji J, Liu B and
Zhang M: Feedback activation of NF-KB signaling leads to adaptive
resistance to EZH2 inhibitors in prostate cancer cells. Cancer Cell
Int. 21:1912021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jung AR, Kim GE, Kim MY, Ha US, Hong SH,
Lee JY, Kim SW and Park YH: HMGB1 promotes tumor progression and
invasion through HMGB1/TNFR1/NF-κB axis in castration-resistant
prostate cancer. Am J Cancer Res. 11:2215–2227. 2021.PubMed/NCBI
|
|
36
|
Staal J and Beyaert R: Inflammation and
NF-κB signaling in prostate cancer: Mechanisms and clinical
implications. Cells. 7:1222018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He S, Chen M, Lin X, Lv Z, Liang R and
Huang L: Triptolide inhibits PDGF-induced proliferation of ASMCs
through G0/G1 cell cycle arrest and suppression of the
AKT/NF-κB/cyclinD1 signaling pathway. Eur J Pharmacol.
867:1728112020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang R, Li J, Yan X, Jin K, Li W, Liu X,
Zhao J, Shang W and Zhao X: Long noncoding RNA MLK7AS1 promotes
proliferation in human colorectal cancer via downregulation of p21
expression. Mol Med Rep. 19:1210–1221. 2019.PubMed/NCBI
|
|
39
|
Cui X, Cui M, Asada R, Kanemoto S, Saito
A, Matsuhisa K, Kaneko M and Imaizumi K: The androgen-induced
protein AIbZIP facilitates proliferation of prostate cancer cells
through downregulation of p21 expression. Sci Rep. 6:373102016.
View Article : Google Scholar : PubMed/NCBIPubMed/NCBI
|