Introduction
Pancreatic cancer is a particularly aggressive
malignant tumor with high morbidity and mortality (1). In 2018, epidemiological research
revealed that pancreatic cancer was the fourth leading cause of
cancer-related deaths in Japan and the third leading cause of
cancer-related deaths in USA, with an estimated 227,000 deaths per
year worldwide (2–4). In 2019, there were >56,000 new
cases of pancreatic cancer and 46,000 pancreatic cancer-related
deaths in the USA alone (3).
Furthermore, pancreatic cancer is predicted to be the second
leading cause of cancer-related deaths in USA by 2030 (5). Risk factors for pancreatic cancer
include smoking, a family history of chronic pancreatitis, advanced
age, occupational exposure, male sex, obesity, diabetes mellitus, a
diet high in meat and low in vegetables and folate and a non-O
blood group (6–8). The 5-year survival rate is <20%,
even after radical surgery (6).
Gemcitabine-based combination chemotherapy has been assessed for
advanced pancreatic cancer. However, even with chemotherapy and
radiotherapy, the effects are insufficient (6). Although numerous trials have been
conducted to establish improved treatments for patients with
advanced pancreatic cancer (S-1, oxaliplatin, fluorouracil and
folic acid), no standard post first-line treatment has yet been
identified (9–12). Thus, the development of new
therapeutic options is urgently required.
The high malignancy of pancreatic cancer is due in
part to constitutively activated NF-κB. NF-κB is a transcription
factor that is associated with various functions, such as cell
proliferation, inflammation and apoptosis. NF-κB is also involved
in angiogenesis and metastasis in cancer, and enhances the
production of angiogenic factors, such as VEGF and IL-8 (13,14).
In our previous study the suppression of NF-κB signaling was found
to decrease the production of both VEGF and IL-8 (15). It was confirmed that these factors
are key mediators of angiogenesis and metastasis in pancreatic
cancer (16–19). Furthermore, previous studies
revealed that these factors support angiogenesis and are necessary
for the metastasis of pancreatic cancer to the liver (20,21).
Based on these results, novel molecular therapies that target NF-κB
are currently under development.
Certain natural products possess anti-inflammatory
and anticancer effects against various cells and cancer types
(22). Hymenialdisine, a natural
product derived from the marine sponge Axinella carteri
(A. carteri), has been reported to exhibit strong
anti-inflammatory activity by suppressing NF-κB (23). Another study revealed that
10Z-Hymenialdisine inhibits the expression of IL-8 mRNA and protein
in U937 cells (24). These data
suggested that 10Z-Hymenialdisine may have potential
anti-inflammatory and anti-angiogenic effects through NF-κB and
various angiogenic factors, such as VEGF and IL-8 (24).
The anti-angiogenic effects of 10Z-Hymenialdisine in
cancer cells have not been investigated. Therefore, in the present
study, the effects of low-dose 10Z-Hymenialdisine on NF-κB
activity, angiogenesis and metastasis in pancreatic cancer were
examined. It was also investigated whether 10Z-Hymenialdisine
prevents the production of angiogenic factors, such as IL-8 and
VEGF. The anti-angiogenic effects of 10Z-Hymenialdisine were also
evaluated in vitro and in vivo.
Materials and methods
Reagents
10Z-Hymenialdisine and dimethyl sulfoxide (DMSO)
were purchased from Sigma-Aldrich; Merck KGaA. Hymenialdisine was
dissolved in DMSO to a stock concentration of 10 mM and stored at
−20°C.
Cell lines and cell culture
The three human pancreatic adenocarcinoma cell lines
MIA PaCa-2 (cat. no. CRL-1420), SW 1990 (cat. no. CRL-2172) and
BxPC-3 (cat. no. CRL-1687) and the immortalized human endothelial
cell line EA.hy926 (cat. no. CRL-2922) were purchased from the
American Type Culture Collection. MIA PaCa-2 and SW-1990 cells were
cultured in DMEM and BxPC-3 cells were cultured in RPMI medium
(both from Sigma Aldrich; Merck KGaA) in a 37°C humidified
incubator with 5% CO2. These media were supplemented
with 10% FBS, 10,000 U/ml penicillin, 25 µg/ml amphotericin B and
10 mg/ml streptomycin (all from Gibco; Thermo Fisher Scientific,
Inc.).
Colony formation assay
Colony formation assays were performed using
Diff-Quik solution (Sysmex Corporation). MIA PaCa-2, SW-1990 and
BxPC-3 cells were seeded at 5×102 cells into each well
of a six-well plate and cultured for 1 day at 37°C. The cells were
then treated with various concentrations (0, 1, 2, 5 and 10 µM) of
10Z-Hymenialdisine. After 7 days of incubation at 37°C, the plates
were gently washed with PBS and subsequently stained with Diff-Quik
solution, which included Diff-Quik fixative reagent for 5 sec,
Diff-Quik solution I (eosinophilic) for 5 sec and Diff-Quik
solution II (basophilic) for 5 sec, as designated in the
manufacturer's protocol at room temperature. Colonies were
determined to be formed when there were ≥50 cancer cells.
Cell proliferation assay
Cell proliferation assays were performed using a
Premix WST-1 Cell proliferation Assay System (Takara Bio, Inc.)
according to the manufacturer's protocol. MIA PaCa-2, SW-1990 and
BxPC-3 cells were seeded at 2×105 cells into each well
of a 96-well plate to a total volume of 100 µl and cultured for 1
day at 37°C. The cells were then treated with various
concentrations (0, 1, 2, 5, 10 and 20 µM) of 10Z-Hymenialdisine and
DMSO (equivalent to the concentration contained in 20 µM) at room
temperature. After incubation for 72 h at 37°C, absorbance was
measured at 450 nm using a Spectra Max 340 spectrophotometer
(Molecular Devices, LLC).
Immunocytochemistry for NF-κB
MIA PaCa-2, SW-1990 and BxPC-3 cells were seeded at
1×104 cells/chamber in a four-chamber glass slide and
cultured for 1 day at 37°C. The cells were treated with
10Z-Hymenialdisine (5 µM) for 2 h at room temperature, followed by
stimulation with TNF-α (1 ng/ml) for 20 min. Cells that had not
been treated were used as controls. The cells were then washed with
PBS and fixed with 4% paraformaldehyde for 20 min at room
temperature. Next, the cells were washed with PBS, permeabilized
with 0.1% Triton-X for 3 min, and incubated with 3% bovine serum
albumin (BSA; FUJIFILM Wako Pure Chemical Corporation) for 1 h at
room temperature. Subsequently, slides were treated with rabbit
antibodies against NF-κB p65 (Cell Signaling Technology, Inc.; cat.
no. 8242; 1:400) overnight at 4°C and then treated with goat
anti-rabbit IgG secondary antibodies H&L (Alexa
Fluor® 488) (cat. no. ab150077; Abcam) for 1 h at room
temperature. Primary and secondary antibodies were used at 1:400
and 1:500 dilution with 3% BSA, respectively. Cell nuclei were
subsequently stained with DAPI (50 ng/ml) at room temperature for
10 min. Images of the stained slides were captured using a BZ-X710
fluorescent microscope at ×400 magnification (Keyence
Corporation).
Protein extraction and NF-κB activity
assays
MIA PaCa-2, SW-1990 and BxPC-3 cells were seeded at
2×106 cells/dish in 100-mm dishes and cultured for 1 day
at 37°C. The cells were treated with or without 10Z-Hymenialdisine
(5 µM) for 2 h and then stimulated with or without TNF-α (1 ng/ml)
for 20 min before the end of incubation. After the indicated
treatment, nuclear protein was extracted using a Nuclear Extract
Kit (Active Motif, Inc.) according to the manufacturer's protocol.
The concentration of proteins in the nuclear extract was evaluated
using a Pierce BCA Protein Assay kit (Thermo Fisher Scientific,
Inc.) and stored at −80°C until use. The activity of NF-κB was
measured using a Trans AM NF-κB p65 Active Transcription Assay Kit
(Active Motif, Inc.) according to the manufacturer's protocol. A
total of 5 micrograms of nuclear extract protein was used in the
NF-κB activity assay.
Reverse transcription-quantitative PCR
(RT-qPCR)
MIA PaCa-2, SW-1990 and BxPC-3 cells were seeded at
4×105 cells/well in six-well plates containing medium
(MIA PaCa-2 and SW-1990 cells were cultured in DMEM and BxPC-3
cells were cultured in RPMI medium) and cultured for 1 day at 37°C.
Then, the medium was changed, and the cells were cultured for an
additional 2 h with or without 10Z-Hymenialdisine (5 µM) at 37°C.
Total RNA was extracted from cell pellets using an RNeasy Plus Mini
Kit (Qiagen KK) according to the manufacturer's protocol and
quantified using a Nano Drop 1000 spectrophotemer (Thermo Fisher
Scientific, Inc.) using 260 nm wavelength. Total RNA (1 µg) was
reverse transcribed using Super Script III First-Strand Synthesis
Super Mix for RT-qPCR kit (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. RT-qPCR was carried
out using TaqMan Fast Advanced Master Mix (cat. no. 4444964; Thermo
Fisher Scientific, Inc.) and TaqMan Gene Expression Assays for
IL-8 (Hs01553824_g1; cat. no. 4331182; Thermo Fisher
Scientific, Inc.), VEGF (Hs00900055_m1; cat. no. 4331182;
Thermo Fisher Scientific, Inc.) and GAPDH (Hs99999905_m1;
cat. no. 4331182; Thermo Fisher Scientific, Inc.) with an Applied
Biosystems 7900HT Fast Real-Time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The thermocycling conditions for
PCR were as follows: initial denaturation at 95°C for 20 sec,
followed by 40 cycles at 95°C for 1 sec and 60°C for 20 sec. The
relative expression levels of VEGF and IL-8 were
normalized to the expression of GAPDH in each sample using
the relative standard curve method (25).
ELISA analysis of IL-8 and VEGF
levels
MIA PaCa-2, SW-1990 and BxPC-3 cells were seeded at
4×105 cells/well in six-well plates containing medium
(MIA PaCa-2 and SW-1990 cells were cultured in DMEM and BxPC-3
cells were cultured in RPMI medium) and cultured for 1 day at 37°C.
The culture medium was subsequently changed, and the cells were
cultured for an additional 72 h at 37°C in the presence of
different concentrations of 10Z-Hymenialdisine (0, 1, 2 and 5 µM).
After incubation, the culture medium was collected and centrifuged
at 400 × g for 5 min at 14°C to remove particles. The supernatants
were frozen at −80°C until use. The concentrations of IL-8 and VEGF
were determined using human IL-8 (R&D Systems, Inc.; cat. no.
D8000C) and human VEGF (R&D Systems, Inc.; cat. no. DVE00)
ELISA kits according to the manufacturer's protocol.
Tube formation assays
Tube formation assays were conducted using EA.hy926
cells and Matrigel matrix (Corning Inc.) in the presence of the
supernatants of pancreatic cancer cells. Cell supernatants were
collected from the cancer cell culture medium (MIA PaCa-2 and
SW-1990 cells were cultured in DMEM and BxPC-3 cells were cultured
in RPMI medium) via centrifugation at 400 × g for 5 min at 14°C.
MIA PaCa-2, SW-1990 and BxPC-3 cells were seeded at
4×105 cells/well in six-well plates containing medium
(MIA PaCa-2 and SW-1990 cells were cultured in DMEM and BxPC-3
cells were cultured in RPMI medium) and cultured for 1 day at 37°C.
The culture media were then changed, and the cells were incubated
for an additional 48 h at 37°C in medium with or without
10Z-Hymenialdisine (5 µM), after which the supernatants were
collected and centrifuged at 400 × g for 5 min at 14°C to remove
particles. EA.hy926 cells (1.2×104 cells/well) were
seeded into each well of a 96-well plate coated with Matrigel.
Matrigel was pre-coated for 30 min at room temperature. Then, the
cells were cultured for 1 day at 37°C with 50 µl 10% FBS and 50 µl
of the aforementioned supernatant. Tube formation was observed by a
BZ-X710 fluorescent microscope at ×40 magnifications (Keyence
Corporation) and evaluated by determining the number of the
endotubes generated by EA.hy926 cells. A total of four random
fields of view were analyzed per sample.
Animal studies
Female BALB/c nu-nu 12 mice (4 weeks old; weight
range, 13–15 g) were purchased from Charles River Laboratories,
Inc. The animals were housed in standard Plexiglas cages (maximum
of 5 mice per cage) in a room maintained at constant temperature
(20–26°C) and humidity (40–60%) with a 12-h light/dark cycle. Mice
were fed regular autoclaved chow and were provided water ad
libitum. Anesthesia was administered by inhalation of
isoflurane. Induction was conducted at 4–5% concentration and
maintenance at 2–3%. Euthanasia was performed by cervical
dislocation after adequate anesthesia. All experiments were
conducted according to the Guidelines for Animal Experiments of
Nagoya City University Graduate School of Medicine Sciences and
were approved (approval no. Medical Animal 20–020) by the Animal
Care and Use Committee of the Nagoya City University Graduate
School of Medical Sciences (Nagoya, Japan).
Experimental protocol
BxPC-3 human pancreatic cells (5×106
cells in 100 µl PBS) were injected subcutaneously into the left
flank of each mouse. Tumor volume was measured once a week using
calipers. When the tumors reached a volume of ~50 mm3,
the mice were randomly divided into two groups (6 mice per
group).
Mice in group I were administered DMSO (the same
concentration as group II) diluted with PBS as a control, whereas
mice in group II received intraperitoneal injections of
10Z-Hymenialdisine (10 mg/kg/week) diluted with PBS. The tumor
volume was calculated according to the following formula: Tumor
volume (mm3)=(AxB2)/2, where A and B
represented the longest and shortest diameters in millimeters,
respectively. Mice were injected intraperitoneally with DMSO or
10Z-Hymenialdisine once a week for 5 weeks and then sacrificed 1
week later. Finally, the tumors were excised and fixed in 10%
formaldehyde at 4°C for 24 h for further analysis.
Histopathological examination
Subcutaneous xenograft tumors were fixed with 4%
paraformaldehyde at 4°C for 6 h and embedded in paraffin. The
sections (3 µm) were mounted on
3-amino-propyltriethoxysilane-coated slides. Dewaxed paraffin
sections were placed in a microwave (10 min, 600 watts) to recover
antigens before staining. The following antibodies (obtained from
Cell Signaling Technology, Inc.) were used at 4°C for 24 h: Rabbit
monoclonal anti-Ki67 antibodies (cat. no. 9027; 1:500), mouse
monoclonal anti-CD31 antibodies (cat. no. 3528; 1:1,000), rabbit
monoclonal anti-NF-κB p65 antibodies (cat. no. 8242; 1:100).
Biotin-conjugated secondary antibodies (all, Dako; Agilent
Technologies, Inc) were also used at room temperature for 45 min:
Anti-rabbit secondary antibodies (cat. no. K4003; 1:500) and
anti-mouse secondary antibodies at room temperature for 45 min
(cat. no. K4001; 1:1,500). Liqid DAB + Substrate Chromogen System
(cat. no. K3467) were used as the chromogen for detection at room
temperature for 10 min. Hematoxylin was used for nuclear
counterstaining at room temperature for 30 sec.
For the quantification of NF-κB p65 activation, the
number of nucleus-positive cells was counted. Nuclear translocation
of p65 was considered to be a marker of NF-κB activation.
Proliferation was quantified by determining the percentage of
Ki67-positive cells. Results were expressed as the mean number of
Ki67-positive cells ± SD per high-power field. A total of three
fields were examined from each tumor for each of the two treatment
groups. For the quantification of microvessel density in sections
stained for CD31, any distinct area of positive staining for CD31
was counted as a single vessel. Results were expressed as the mean
number of vessels ± the SD per high-power field. A total of three
random fields were examined and counted from each tumor for each of
the two treatment groups. Images of the stained slides were
captured using a BZ-X710 fluorescent microscope at ×200
magnification (Keyence Corporation).
Statistical analysis
All experiments, except for in vivo
procedures, were performed in triplicate. All experimental data are
presented as the mean ± SD. Multiple group comparisons were
performed using one-way ANOVA with Dunnett's post-hoc tests for
subsequent individual group comparisons. Comparisons between two
groups were performed using unpaired Student's t-tests. Comparisons
between multiple groups with >1 reference group were analysed
using Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference. All statistical
analyses were performed with EZR Version 1.54 (Saitama Medical
Center, Jichi Medical University, Saitama, Japan), which is a
modified version of R commander designed to add statistical
functions frequently used in biostatistics (26).
Results
10Z-Hymenialdisine suppresses the
proliferation of pancreatic cancer cell lines
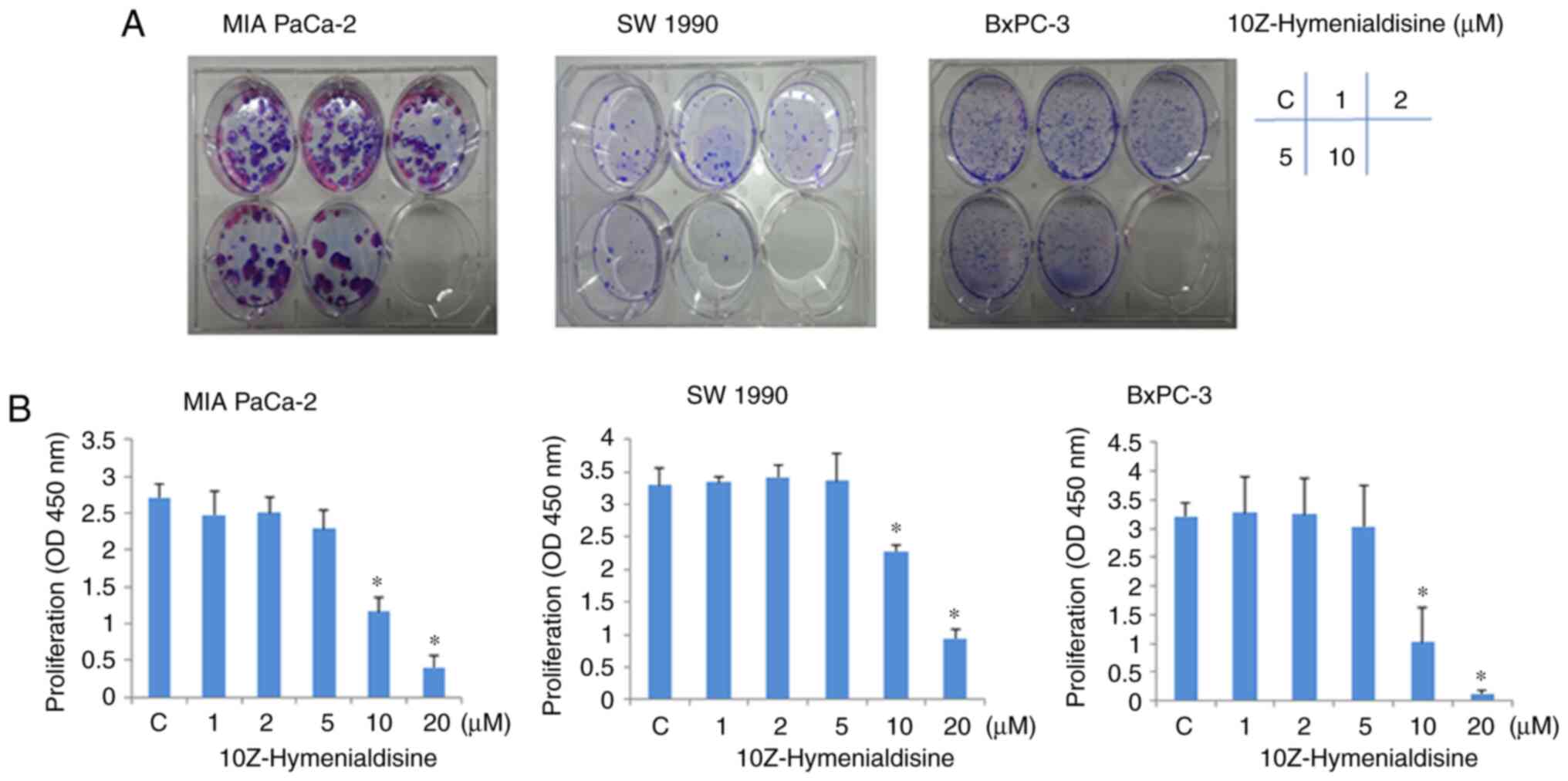
Colony formation assays revealed that the growth of
pancreatic cancer cell lines (MIA PaCa-2, SW-1990 and BxPC-3) was
suppressed by 10Z-Hymenialdisine in a concentration-dependent
manner (Fig. 1A). Consistent with
this, WST-1 assays revealed that 10Z-Hymenialdisine inhibited the
proliferation of all three pancreatic cancer cell lines in a
concentration-dependent manner, with significant inhibition
observed at 10 µM (Fig. 1B). In
subsequent experiments, to avoid cytotoxicity of
10Z-Hymenialdisine, the concentration of 10Z-Hymenialdisine was set
at 5 µM.
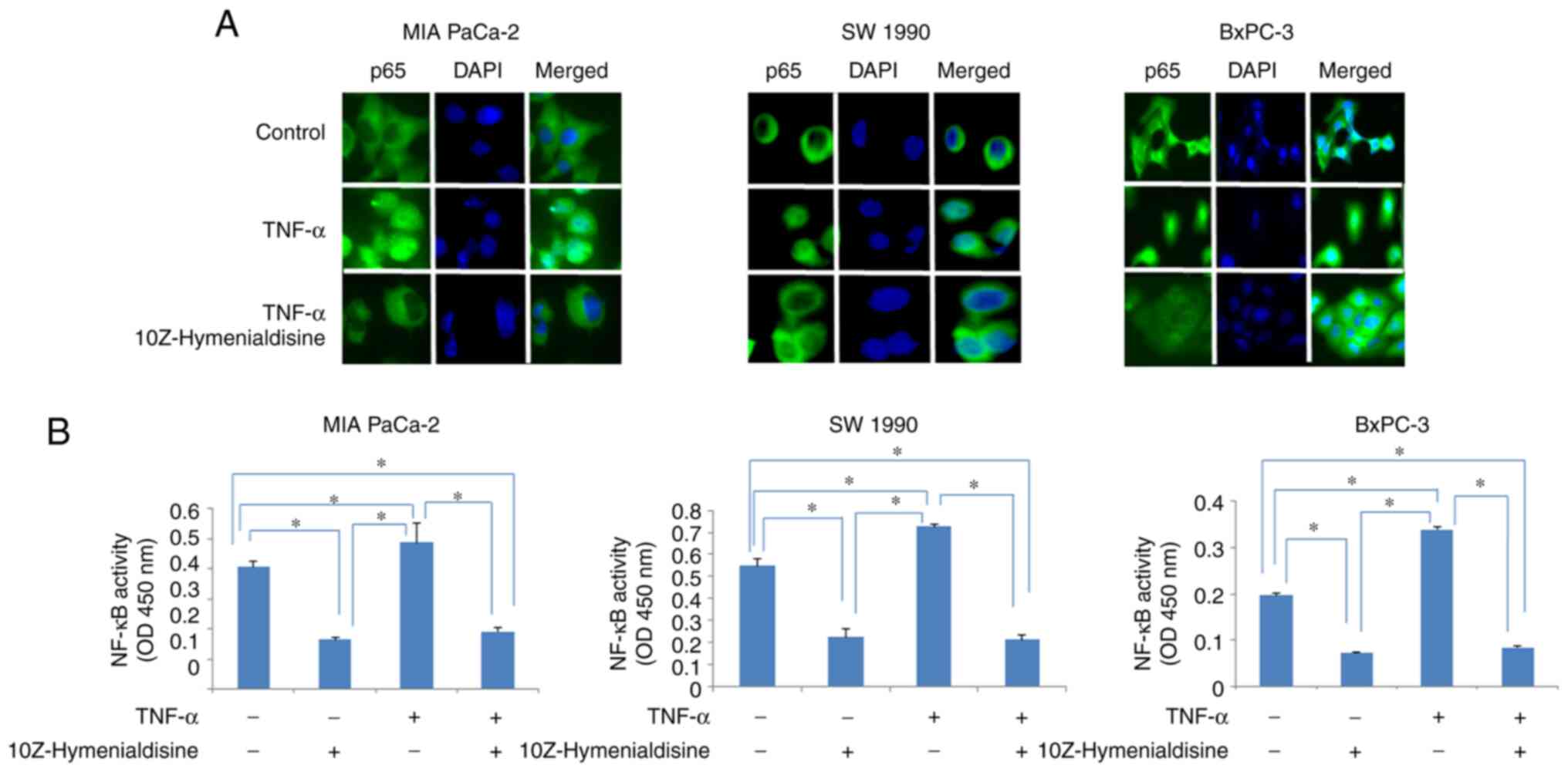
10Z-Hymenialdisine inhibits the
TNF-α-induced translocation of NF-κB
Immunocytochemical analysis revealed that p65
translocated into the nucleus of cells treated with TNF-α alone,
whereas in cells treated with TNF-α and 10Z-Hymenialdisine (5 µM),
p65 remained in the cytoplasm. The translocation of p65 into the
nucleus was then examined using ELISA (Fig. 2A and B). The results demonstrated
that 10Z-Hymenialdisine significantly decreased the translocation
of p65 into the nucleus. This effect was confirmed with TNF-α.
TNF-α alone significantly increased p65 translocation; however, the
addition of 10Z-Hymenialdisine significantly decreased this effect
(Fig. 2B).
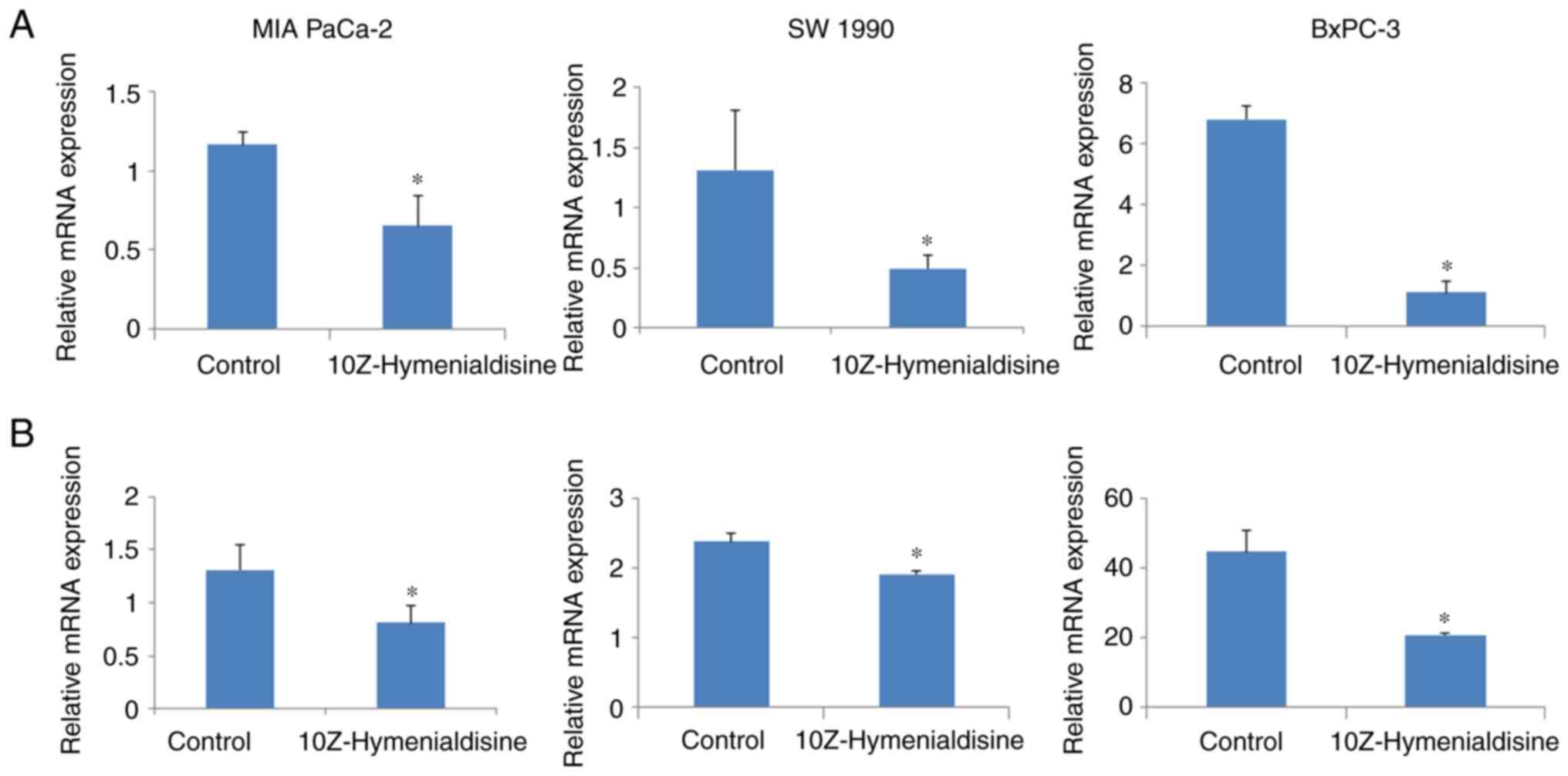
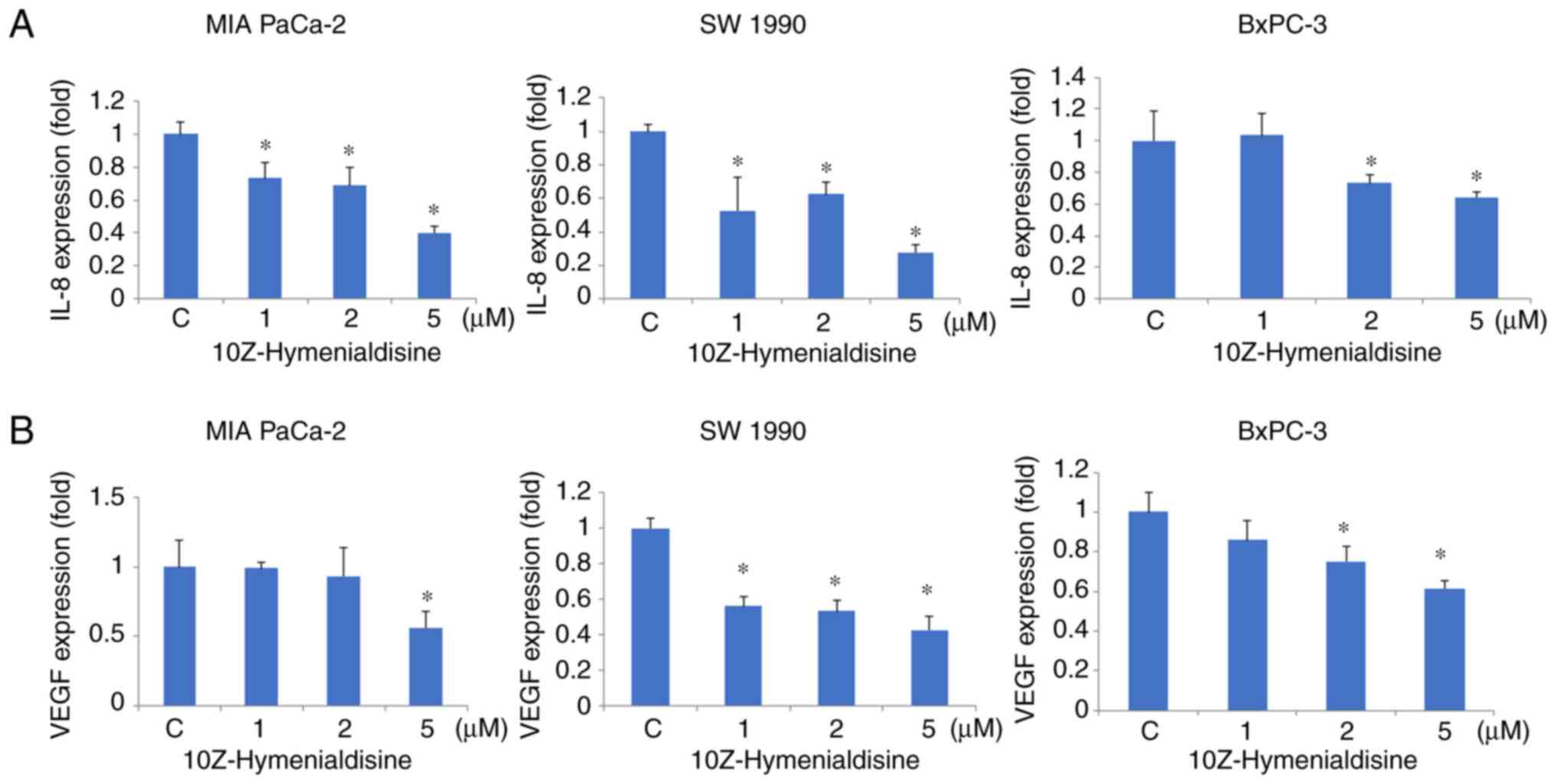
10Z-Hymenialdisine decreases mRNA
expression levels of IL-8 and VEGF
RT-qPCR revealed that 10Z-Hymenialdisine (5 µM)
significantly decreased the mRNA expression levels of IL-8 and VEGF
in pancreatic cell lines (Fig. 3A and
B). In addition, ELISA revealed that the protein secretion
levels of IL-8 and VEGF were reduced by 10Z-Hymenialdisine (1–5 µM;
Fig. 4A and B). In the present
experiment, all pancreatic cancer cell lines exhibited reduced
levels of IL-8 and VEGF proteins in the presence of different
concentrations of 10Z-Hymenialdisine (1–5 µM). For cells treated
with 5 µM 10Z-Hymenialdisine, all pancreatic cancer cell lines
demonstrated significant reductions in both IL-8 and VEGF. It was
confirmed that the concentration of 5 µM was the most appropriate
to inhibit angiogenic factors without causing cytotoxicity.
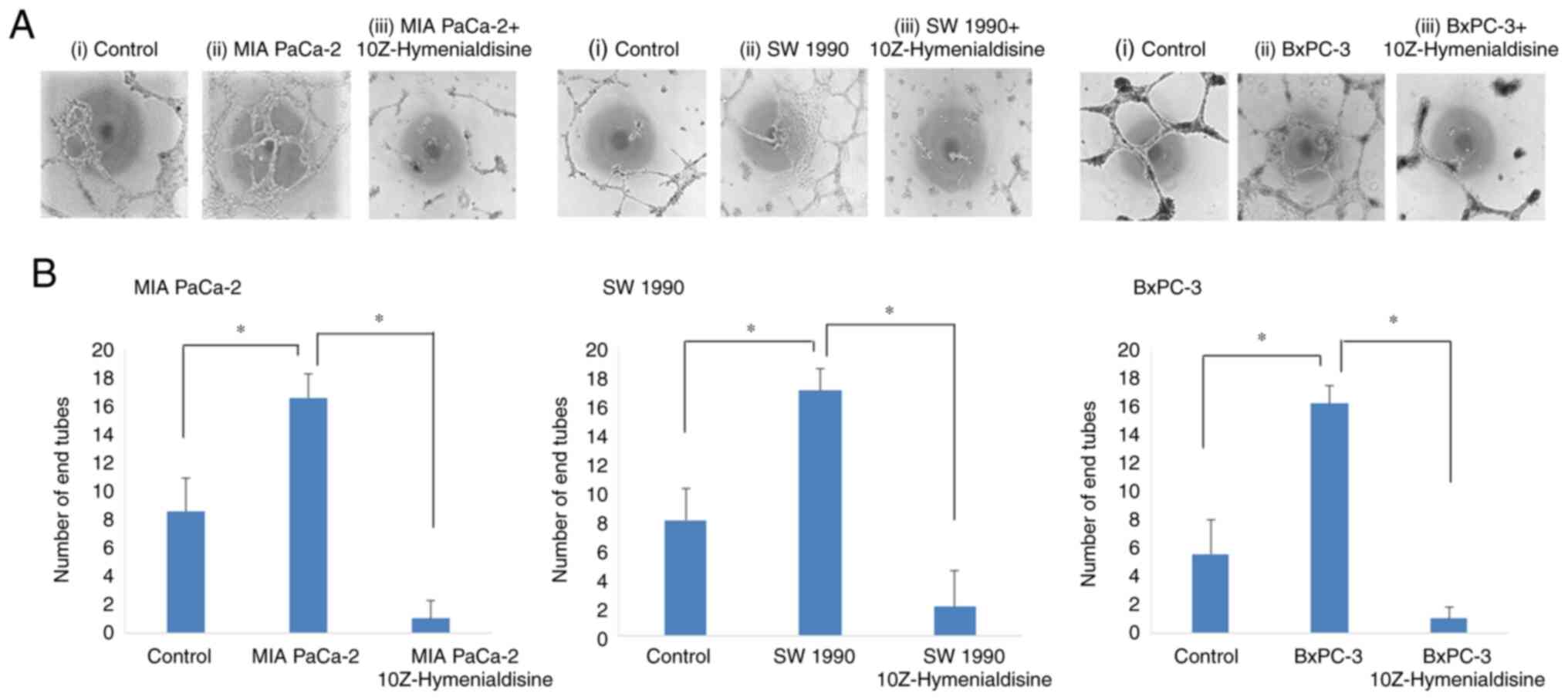
10Z-Hymenialdisine inhibits tube
formation by human endothelial (EA.hy926) cells
Subsequently, tube formation assays were performed
using human endothelial cells. Tube formation was enhanced when the
cells were incubated with supernatants from untreated pancreatic
cancer cell lines, but was significantly decreased when the cells
were incubated with supernatants from 10Z-Hymenialdisine-treated
pancreatic cancer cell lines (Fig.
5A). Tube formation was then evaluated by counting the number
of endotubes. Endotube numbers were significantly increased when
the cells were incubated with supernatants from cancer cell lines
and significantly decreased when the cells were incubated with
supernatants from 10Z-Hymenialdisine-treated pancreatic cancer cell
lines (Fig. 5B).
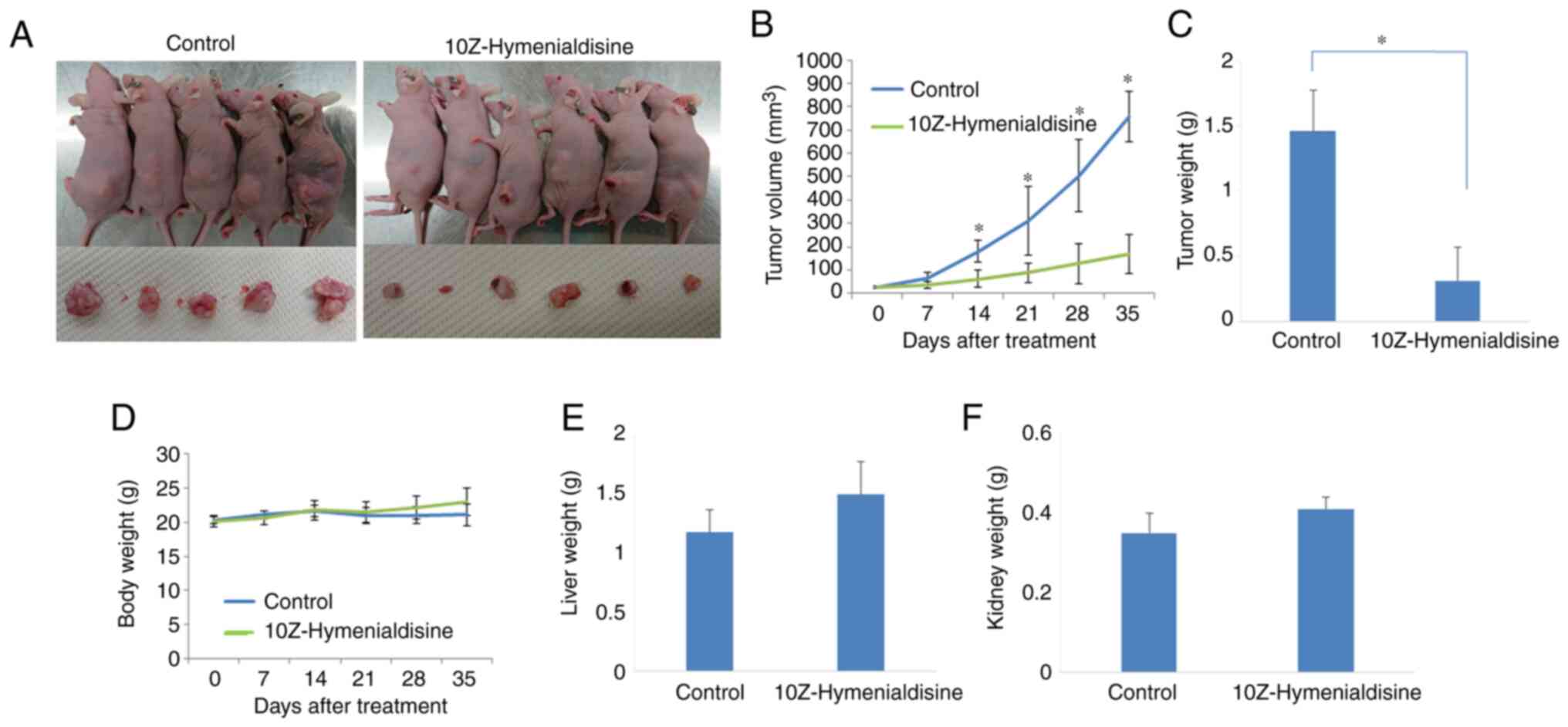
10Z-Hymenialdisine suppresses tumor
growth in a subcutaneous xenograft model
BxPC-3 cells were subcutaneously injected into mice,
after which tumors were allowed to form. When the tumors had grown,
mice were injected intraperitoneally with 10Z-Hymenialdisine (10
mg/kg) or DMSO for 5 weeks. One mouse in the control group
succumbed in the middle of the treatment course owing to the
maintenance 2–3% inhalation of isoflurane anesthesia.
10Z-Hymenialdisine significantly inhibited tumor growth 14 days
after treatment (Fig. 6A-C). Body,
liver and kidney weights were not decreased compared with those in
the control group (Fig. 6D-F).
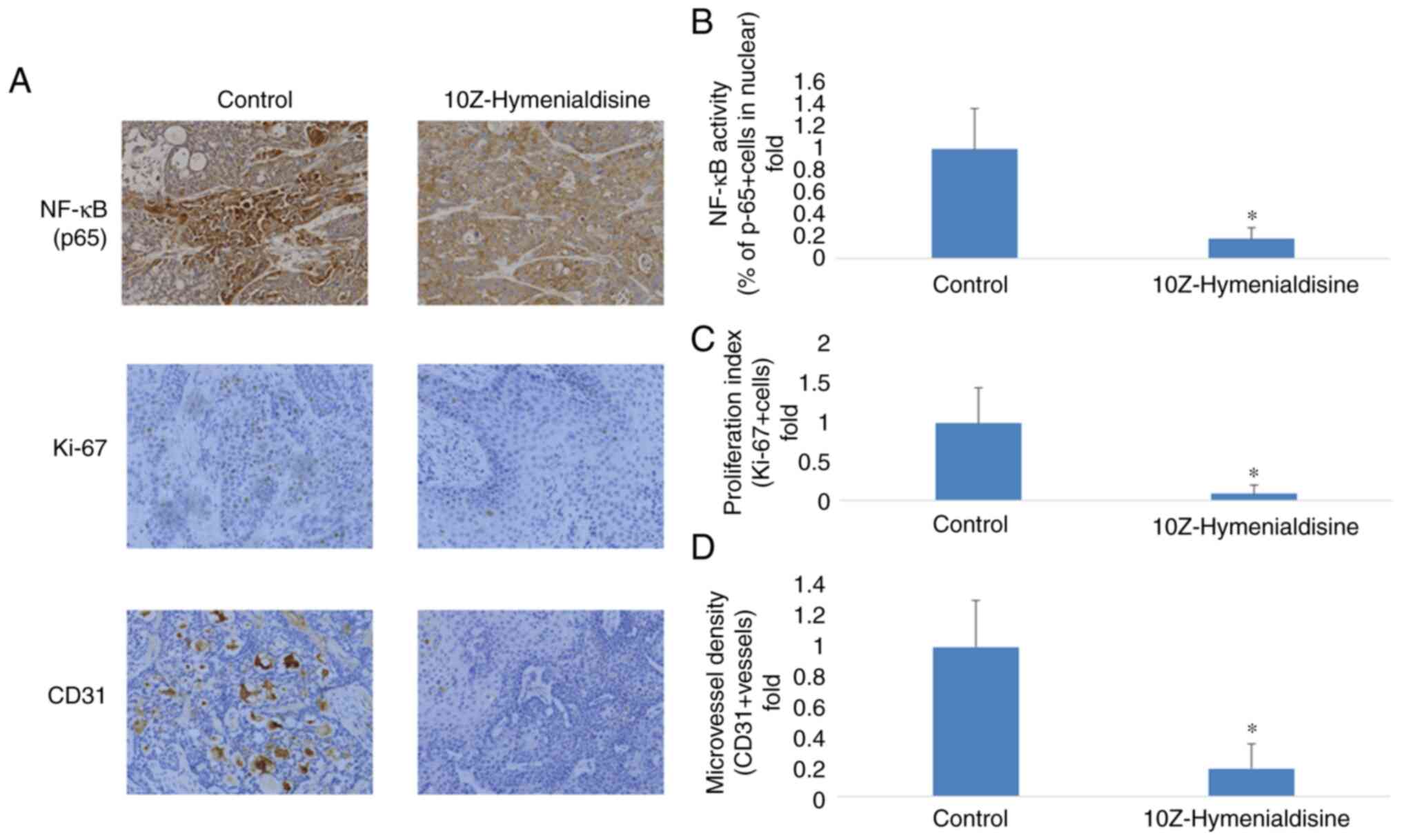
10Z-Hymenialdisine inhibits NF-κB
(p65) activation, cell proliferation (Ki-67) and angiogenesis
(CD31) in pancreatic cancer tumors
Resected pancreatic cancer tumors were evaluated
using immunocytochemical analysis. It was revealed that p65
translocation into the nucleus was decreased in groups treated with
10Z-Hymenialdisine compared with the control group. To determine
the effects of 10Z-Hymenialdisine on proliferation and
angiogenesis, the expression of Ki-67, a cell proliferation marker,
and CD31, a microvessel density marker, were examined in tumor
tissues. As demonstrated in Fig.
7, 10Z-Hymenialdisine significantly decreased the activation of
NF-κB (p65) and inhibited the expression of Ki-67 and CD31
(Fig. 7A-D).
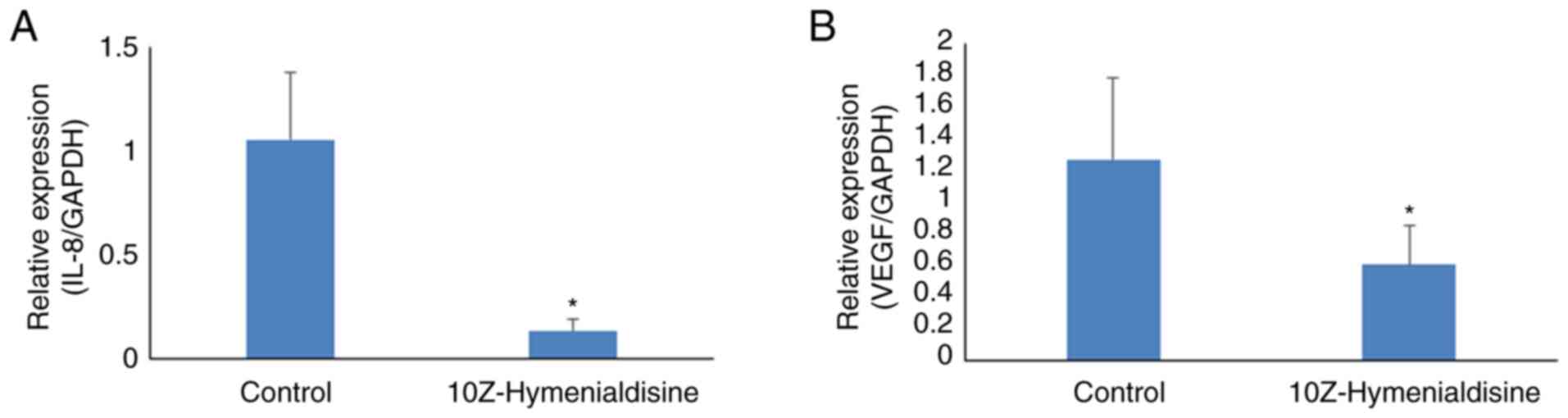
10Z-Hymenialdisine decreases mRNA
expression levels of IL-8 and VEGF in pancreatic cancer tumors
The expression levels of targeted mRNAs in resected
pancreatic cancer tumors were then evaluated using RT-qPCR. The
results revealed that 10Z-Hymenialdisine significantly decreased
the mRNA expression levels of IL-8 and VEGF in
pancreatic cancer tumors compared with that in control tumors
(Fig. 8A and B).
Discussion
10Z-Hymenialdisine is a natural product derived from
the marine sponge A. carteri. In the present study, it was
demonstrated that 10Z-Hymenialdisine inhibited the proliferation of
pancreatic cancer cells, blocked NF-κB activity and suppressed the
expression of the angiogenic factors VEGF and IL-8. In addition,
10Z-Hymenialdisine inhibited tube formation by EA.hy926 human
endothelial cells. These experiments suggested that
10Z-Hymenialdisine may have potential applications as an
anti-angiogenesis agent. Finally, using in vivo modeling, it
was identified that 10Z-Hymenialdisine inhibited tumor growth by
subcutaneously injecting BxPC-3 pancreatic cancer cells. In this
model, no decrease was observed in body, liver, or kidney weights
in the 10Z-Hymenialdisine-treated group of mice. Previous studies
revealed that NF-κB has important roles in the regulation of
apoptosis, oncogenesis and angiogenesis (27,28).
Importantly, ~70% of pancreatic cancer cells exhibit constitutive
activation of NF-κB (29).
Furthermore, in certain studies, upregulation of NF-κB explains, at
least in part, the resistance of cells to chemotherapy (30–33).
The aggressive growth and metastasis of pancreatic
cancer is partly caused by its angiogenic capacity, which has been
attributed to the release of angiogenic factors derived from tumors
and stromal cells (34). Numerous
angiogenic factors have been identified, among which VEGF and IL-8
are considered to be key mediators of angiogenesis in pancreatic
cancer (16,18). Suppression of angiogenic factors
would be a highly desirable effect of any new chemotherapeutic
agent.
Given the relationship between NF-κB and certain
angiogenic factors, such as VEGF and IL-8, suppressing the activity
of NF-κB would be beneficial in treating pancreatic cancer. A
previous study demonstrated that inhibition of NF-κB activity by
the proteasome inhibitor MG132 decreases the expression of VEGF and
IL-8 and inhibits tumor-induced angiogenesis (15). The experiments of the present study
were consistent with these previous studies. It was demonstrated
that 10Z-Hymenialdisine inhibited tube formation by EA.hy926 human
endothelial cells. Thus, the data revealed that 10Z-Hymenialdisine
suppressed the expression of VEGF and IL-8 via inhibition of NF-κB
activity in pancreatic cancer cell lines. As a result,
10Z-Hymenialdisine inhibited tube formation by human endothelial
cells.
The anti-angiogenic effects of bortezomib, an NF-κB
inhibitor, have been exploited for the treatment of patients with
multiple myeloma (35); however,
this NF-κB inhibitor has strong side effects, and adoption of such
chemotherapeutic agents is not widely accepted (35). Consequently, new NF-κB inhibitors
with low non-specific toxicity are urgently needed.
In Nagoya City University Graduate School of Medical
Sciences and Medical School, the Department of Gastroenterological
Surgery's laboratory, focus has been addressed on natural products.
Natural agents, such as escin, curcumin, sesamin, xanthohumol and
zerumbone, have been shown to have anticancer effects (22,36–40).
Previous studies have demonstrated that these natural products
inhibit NF-κB and certain angiogenic factors, such as VEGF and
IL-8, and exhibit efficacy in in vivo models (39,40).
Xanthohumol exerts anticancer effects without causing body weight
changes in mice (40).
Furthermore, curcumin, a natural product, can be used in
combination with gemcitabine to inhibit the activity of NF-κB and
exert synergistic effects on tumor suppression (41). Accordingly, 10Z-Hymenialdisine may
have applications as a combination therapy with existing anticancer
agents, such as gemcitabine, to further enhance the anticancer
effects of the treatment, similar to curcumin. Previous studies
have revealed that 10Z-Hymenialdisine inhibits NF-κB and various
cytokines, including IL-1, IL-2, IL-8 and TNF-α in certain types of
cells (24,42). Additionally, 10Z-Hymenialdisine has
been reported to have anticancer effects (43); however, this is the first report,
to the best of our knowledge, demonstrating that 10Z-Hymenialdisine
exerts inhibitory effects on pancreatic cancer cells by suppressing
NF-κB and angiogenesis.
To assess the cytotoxic effects of
10Z-Hymenialdisine, colony formation and cell proliferation assays
were conducted. Since previous studies revealed that
10Z-Hymenialdisine is active at concentrations of 1–10 µM, these
concentrations were initially examined (23,24).
In the latter experiment, 10Z-Hymenialdisine showed cytotoxic
effects at high concentrations (>10 µM). In subsequent
experiments, to confirm the anti-angiogenic effects of
10Z-Hymenialdisine, the experiments were primarily conducted using
low concentrations (≤5 µM), which did not suppress cell
proliferation. At low concentrations (≤5 µM), 10Z-Hymenialdisine
inhibited the activity of NF-κB and blocked the expression of VEGF
and IL-8. 10Z-Hymenialdisine also inhibited tube formation. Thus,
it was concluded that 10Z-Hymenialdisine may have potential
clinical applications via suppression of angiogenesis, without
causing substantial cytotoxicity.
In terms of angiogenesis, pancreatic cancer is
generally considered to be a type of ischemic tumor, and vessel
densities are low in imaging; however, angiogenesis is associated
with prognosis in patients with pancreatic cancer (44–46),
and certain studies have demonstrated that anti-angiogenic
treatments exhibit favorable efficacy in pancreatic cancer
(47,48). Similarly, to ensure that the
findings of the present study provided an adequate reflection of
pancreatic cancer, experiments were conducted using three different
pancreatic cancer cell lines [MIA PaCa-2 (undifferentiation, with
KRAS mutation), SW-1990 (high/moderate differentiation, with
KRAS mutation) and BxPC-3 (moderate differentiation, without
KRAS mutation)]. In all in vitro experiments, these
three pancreatic cancer cell lines showed similar trends with
regard to the effects of 10Z-Hymenialdisine.
In the mouse experiments of the current study, the
suppression of tumor growth by 10Z-Hymenialdisine was demonstrated.
No previous studies have described the use of 10Z-Hymenialdisine in
such in vivo models. Therefore, it remains necessary to
optimize the dosage and administration method for
10Z-Hymenialdisine. It was previously identified that another
natural product, xanthohumol, inhibits pancreatic cancer growth
(40). Xanthohumol has a molecular
weight similar to that of 10Z-Hymenialdisine, and its anticancer
effects are achieved at a similar concentration (10 mg/kg) in
vitro. Based on these findings, the appropriate concentration
and administration method for 10Z-Hymenialdisine was selected;
however, further studies are required to optimize its use.
In conclusion, the results of the present study
demonstrated that 10Z-Hymenialdisine inhibited angiogenesis by
suppressing NF-κB activity in pancreatic cancer cell lines. This
effect was achieved by blocking the expression of various
angiogenesis factors, such as IL-8 and VEGF. The anti-angiogenic
effects of 10Z-Hymenialdisine were also demonstrated using EA-hy926
human endothelial cells. Finally, it was revealed that
10Z-Hymenialdisine had anticancer effects in a mouse model. These
effects were obtained at relatively low concentrations that did not
exhibit cytotoxicity. Furthermore, there was no body weight loss or
other organ weight loss. As with the majority of studies, the
design of the current study is subject to some limitations. For
example, it was not known where and how 10Z-Hymenialdisine acts on
the NF-κB pathway. Furthermore, the animal experiment was performed
using the subcutaneous injection of pancreatic cancer cells, which
is different from actual pancreatic cancer. Therefore, the results
of the animal experiment cannot be directly applied to humans. So
further investigation is required, such as orthotopic cancer model
in vivo experiments. However, 10Z-Hymenialdisine could be a
potential therapeutic agent for pancreatic cancer.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The data generated or analyzed during this study are
included in the published article.
Authors' contributions
GU and YM contributed to the conception and design
of the study, analyzed and interpreted the data and wrote and
reviewed the manuscript. GU, YM, HM, YA, TK, KO, YH, HI, KS, KT,
MM, AM, MK and ST designed the study. GU, HM, YA, TK, OK and YH
acquired the data. KO, KS, HI, RO and HT wrote the ‘Materials and
methods’ section of the manuscript. RO and HT provided technical
support in performing RT-qPCR, immunohistochemical and angiogenesis
assays. YM, AM, MK and ST supervised the study. All authors read
and approved the final manuscript and are equally responsible for
all aspects of the study, ensuring the integrity and accuracy of
all parts of the study. GU and YM confirm the authenticity of all
the raw data.
Ethics approval and consent to
participate
All experiments were conducted according to the
Guidelines for Animal Experiments of Nagoya City University
Graduate School of Medicine Sciences and were approved (approval
no. Medical Animal 20–020) by the Animal Care and Use Committee of
the Nagoya City University Graduate School of Medical Sciences
(Nagoya, Japan).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
GBD 2017 Pancreatic Cancer Collaborators,
. The global, regional, and national burden of pancreatic cancer
and its attributable risk factors in 195 countries and territories,
1990-2017: A systematic analysis for the global burden of disease
study 2017. Lancet Gastroenterol Hepatol. 4:934–947. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klein AP, Brune KA, Petersen GM, Goggins
M, Tersmette AC, Offerhaus GJA, Griffin C, Cameron JL, Yeo CJ, Kern
S and Hruban RH: Prospective risk of pancreatic cancer in familial
pancreatic cancer kindreds. Cancer Res. 64:2634–2638. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wolpin BM, Chan AT, Hartge P, Chanock SJ,
Kraft P, Hunter DJ, Giovannucci EL and Fuchs CS: ABO blood group
and the risk of pancreatic cancer. J Natl Cancer Inst. 101:424–431.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoo C, Hwang JY, Kim JE, Kim TW, Lee JS,
Park DH, Lee SS, Seo DW, Lee SK, Kim MH, et al: A randomised phase
II study of modified FOLFIRI.3 vs modified FOLFOX as second-line
therapy in patients with gemcitabine-refractory advanced pancreatic
cancer. Br J Cancer. 101:1658–1663. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oettle H, Riess H, Stieler JM, Heil G,
Schwaner I, Seraphin J, Görner M, Mölle M, Greten TF, Lakner V, et
al: Second-line oxaliplatin, folinic acid, and fluorouracil versus
folinic acid and fluorouracil alone for gemcitabine-refractory
pancreatic cancer: Outcomes from the CONKO-003 trial. J Clin Oncol.
32:2423–2429. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Katsuda M, Miyazawa M, Ojima T, Katanuma
A, Hakamada K, Sudo K, Asahara S, Endo I, Ueno M, Hara K, et al: A
double-blind randomized comparative clinical trial to evaluate the
safety and efficacy of dendritic cell vaccine loaded with WT1
peptides (TLP0-001) in combination with S-1 in patients with
advanced pancreatic cancer refractory to standard chemotherapy.
Trials. 20:2422019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ioka T, Ueno M, Ueno H, Park JO, Chang HM,
Sasahira N, Kanai M, Chung IJ, Ikeda M, Nakamori S, et al: TAS-118
(S-1 plus leucovorin) versus S-1 in patients with
gemcitabine-refractory advanced pancreatic cancer: A randomised,
open-label, phase 3 study (GRAPE trial). Eur J Cancer. 106:78–88.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Q, Lenardo MJ and Baltimore D: 30
Years of NF-κB: A blossoming of relevance to human pathobiology.
Cell. 168:37–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Taniguchi K and Karin M: NF-κB,
inflammation, immunity and cancer: Coming of age. Nat Rev Immunol.
18:309–324. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsuo Y, Sawai H, Ochi N, Yasuda A,
Sakamoto M, Takahashi H, Funahashi H, Takeyama H and Guha S:
Proteasome inhibitor MG132 inhibits angiogenesis in pancreatic
cancer by blocking NF-kappaB activity. Dig Dis Sci. 55:1167–1176.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Itakura J, Ishiwata T, Friess H, Fujii H,
Matsumoto Y, Büchler MW and Korc M: Enhanced expression of vascular
endothelial growth factor in human pancreatic cancer correlates
with local disease progression. Clin Cancer Res. 3:1309–1316.
1997.PubMed/NCBI
|
|
17
|
Shi Q, Le X, Abbruzzese JL, Peng Z, Qian
CN, Tang H, Xiong Q, Wang B, Li XC and Xie K: Constitutive Sp1
activity is essential for differential constitutive expression of
vascular endothelial growth factor in human pancreatic
adenocarcinoma. Cancer Res. 61:4143–4154. 2001.PubMed/NCBI
|
|
18
|
Shi Q, Abbruzzese JL, Huang S, Fidler IJ,
Xiong Q and Xie K: Constitutive and inducible interleukin 8
expression by hypoxia and acidosis renders human pancreatic cancer
cells more tumorigenic and metastatic. Clin Cancer Res.
5:3711–3721. 1999.PubMed/NCBI
|
|
19
|
Matsuo Y, Ochi N, Sawai H, Yasuda A,
Takahashi H, Funahashi H, Takeyama H, Tong Z and Guha S: CXCL8/IL-8
and CXCL12/SDF-1alpha co-operatively promote invasiveness and
angiogenesis in pancreatic cancer. Int J Cancer. 124:853–861. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsuo Y, Sawai H, Funahashi H, Takahashi
H, Sakamoto M, Yamamoto M, Okada Y, Hayakawa T and Manabe T:
Enhanced angiogenesis due to inflammatory cytokines from pancreatic
cancer cell lines and relation to metastatic potential. Pancreas.
28:344–352. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matsuo Y, Sawai H, Ochi N, Yasuda A,
Takahashi H, Funahashi H, Takeyama H and Guha S: Interleukin-1alpha
secreted by pancreatic cancer cells promotes angiogenesis and its
therapeutic implications. J Surg Res. 153:274–281. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gupta SC, Kim JH, Prasad S and Aggarwal
BB: Regulation of survival, proliferation, invasion, angiogenesis,
and metastasis of tumor cells through modulation of inflammatory
pathways by nutraceuticals. Cancer Metastasis Rev. 29:405–434.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Roshak A, Jackson JR, Chabot-Fletcher M
and Marshall LA: Inhibition of NFkappaB-mediated
interleukin-1beta-stimulated prostaglandin E2 formation by the
marine natural product Hymenialdisine. J Pharmacol Exp Ther.
283:955–961. 1997.PubMed/NCBI
|
|
24
|
Breton JJ and Chabot-Fletcher MC: The
natural product Hymenialdisine inhibits interleukin-8 production in
U937 cells by inhibition of nuclear factor-kappaB. J Pharmacol Exp
Ther. 282:459–466. 1997.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kanda Y: Investigation of the freely
available easy-to-use software ‘EZR’ for medical statistics. Bone
Marrow Transplant. 48:452–458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aggarwal BB: Nuclear factor-kappaB: The
enemy within. Cancer Cell. 6:203–208. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shi Q, Le X, Wang B, Abbruzzese JL, Xiong
Q, He Y and Xie K: Regulation of vascular endothelial growth factor
expression by acidosis in human cancer cells. Oncogene.
20:3751–3756. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiong HQ, Abbruzzese JL, Lin E, Wang L,
Zheng L and Xie K: NF-kappaB activity blockade impairs the
angiogenic potential of human pancreatic cancer cells. Int J
Cancer. 108:181–188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Q, Yang G, Feng M, Zheng S, Cao Z, Qiu
J, You L, Zheng L, Hu Y, Zhang T and Zhao Y: NF-κB in pancreatic
cancer: Its key role in chemoresistance. Cancer Lett. 421:127–134.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Arlt A and Schäfer H: NFkappaB-dependent
chemoresistance in solid tumors. Int J Clin Pharmacol Ther.
40:336–347. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sebens S, Arlt A and Schäfer H: NF-kappaB
as a molecular target in the therapy of pancreatic carcinoma.
Recent Results Cancer Res. 177:151–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Holcomb B, Yip-Schneider M and Schmidt CM:
The role of nuclear factor kappaB in pancreatic cancer and the
clinical applications of targeted therapy. Pancreas. 36:225–235.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Folkman J: Angiogenesis and angiogenesis
inhibition: An overview. EXS. 79:1–8. 1997.PubMed/NCBI
|
|
35
|
Chen D, Frezza M, Schmitt S, Kanwar J and
Dou QP: Bortezomib as the first proteasome inhibitor anticancer
drug: Current status and future perspectives. Curr Cancer Drug
Targets. 11:239–253. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tsuboi K, Matsuo Y, Shamoto T, Shibata T,
Koide S, Morimoto M, Guha S, Sung B, Aggarwal BB, Takahashi H and
Takeyama H: Zerumbone inhibits tumor angiogenesis via NF-κB in
gastric cancer. Oncol Rep. 31:57–64. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aggarwal BB, Van Kuiken ME, Iyer LH,
Harikumar KB and Sung B: Molecular targets of nutraceuticals
derived from dietary spices: Potential role in suppression of
inflammation and tumorigenesis. Exp Biol Med (Maywood).
234:825–849. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Harikumar KB, Sung B, Tharakan ST, Pandey
MK, Joy B, Guha S, Krishnan S and Aggarwal BB: Sesamin manifests
chemopreventive effects through the suppression of NF-kappa
B-regulated cell survival, proliferation, invasion, and angiogenic
gene products. Mol Cancer Res. 8:751–761. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Omi K, Matsuo Y, Ueda G, Aoyama Y, Kato T,
Hayashi Y, Imafuji H, Saito K, Tsuboi K, Morimoto M, et al: Escin
inhibits angiogenesis by suppressing interleukin-8 and vascular
endothelial growth factor production by blocking nuclear factor-κB
activation in pancreatic cancer cell lines. Oncol Rep. 45:552021.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Saito K, Matsuo Y, Imafuji H, Okubo T,
Maeda Y, Sato T, Shamoto T, Tsuboi K, Morimoto M, Takahashi H, et
al: Xanthohumol inhibits angiogenesis by suppressing nuclear
factor-κB activation in pancreatic cancer. Cancer Sci. 109:132–140.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kunnumakkara AB, Guha S, Krishnan S,
Diagaradjane P, Gelovani J and Aggarwal BB: Curcumin potentiates
antitumor activity of gemcitabine in an orthotopic model of
pancreatic cancer through suppression of proliferation,
angiogenesis, and inhibition of nuclear factor-kappaB-regulated
gene products. Cancer Res. 67:3853–3861. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sharma V, Lansdell TA, Jin G and Tepe JJ:
Inhibition of cytokine production by Hymenialdisine derivatives. J
Med Chem. 47:3700–3703. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dobretsov S, Tamimi Y, Al-Kindi MA and
Burney I: Screening for anti-cancer compounds in marine organisms
in Oman. Sultan Qaboos Univ Med J. 16:e168–e174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Stipa F, Lucandri G, Limiti MR, Bartolucci
P, Cavallini M, Di Carlo V, D'Amato A, Ribotta G and Stipa S:
Angiogenesis as a prognostic indicator in pancreatic ductal
adenocarcinoma. Anticancer Res. 22:445–449. 2002.PubMed/NCBI
|
|
45
|
Benckert C, Thelen A, Cramer T, Weichert
W, Gaebelein G, Gessner R and Jonas S: Impact of microvessel
density on lymph node metastasis and survival after curative
resection of pancreatic cancer. Surg Today. 42:169–176. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Amin Z, Theis B, Russell RC, House C,
Novelli M and Lees WR: Diagnosing pancreatic cancer: The role of
percutaneous biopsy and CT. Clin Radiol. 61:996–1002. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Annese T, Tamma R, Ruggieri S and Ribatti
D: Angiogenesis in pancreatic cancer: Pre-clinical and clinical
studies. Cancers (Basel). 11:3812019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang Z, Ji S, Zhang B, Liu J, Qin Y, Xu J
and Yu X: Role of angiogenesis in pancreatic cancer biology and
therapy. Biomed Pharmacother. 108:1135–1140. 2018. View Article : Google Scholar : PubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBI
|