Introduction
Cancer is one of the major issues affecting public
health and is considered the leading cause of mortality worldwide
(1,2). There were 1,806,590 new cancer cases
and 606,520 cancer-related deaths in the United States in 2020
(2). The major treatment
strategies for cancer include surgery, radiotherapy, chemotherapy,
biotherapy and immunotherapy. Moreover, traditional Chinese
medicine (TCM) has gradually become a hot topic of cancer research
and therapy in recent years due to its therapeutic efficacy
(particularly in combination with Western medicine), mild
side-effects, its cost-effectiveness and widely available sources.
However, its scope of clinical application in cancer therapy is
limited by complex components and unclear therapeutic targets
(3,4). Therefore, the use of modern science
and technology to examine the effects of active components from TCM
and the associated molecular mechanisms in cancer cells and animal
models of human cancer is a crucial and feasible strategy for the
discovery and development of novel anticancer drugs and cancer
therapeutics.
Apoptosis is a type of programmed cell death
accompanied by cell membrane contraction, nucleus fragmentation,
chromatin condensation and chromosome DNA breakage (5). The study of apoptosis has been an
emerging field in cancer research for several years (5). Apoptosis can be initiated either
through the extrinsic (death receptor-mediated) or the intrinsic
(mitochondria-mediated) pathway, which activates the initiator
caspases for the further activation of the caspase cascade and
caspase-3 to induce cellular apoptosis (6,7).
Autophagy is a highly self-regulated catabolic process in
eukaryotic organisms, and similar to apoptosis, it plays a critical
role in the growth and development of organisms (8,9).
Autophagy can potentially function as the dual nature mechanisms of
pro-survival and pro-death in the initiation and progression of
cancer (10). The pro-survival
effects of autophagy on cancer are mainly due to a mechanism that
permits increased tolerance to stress, obtaining ATP and metabolic
intermediates (11). However, an
increase in the autophagic flux may induce cell death as a
tumor-suppressive mechanism (12).
There are two well-known signaling pathways involved in the
autophagy progress, namely the PI3K/Akt/mTOR/p70S6K and
Ras/Raf/MEK1/2/ERK1/2 signaling pathways (13,14).
It has been reported that autophagy can be negatively controlled by
mTOR, while AMP-activated protein kinase (AMPK) can inhibit mTOR to
govern autophagy (15).
Furthermore, a connection between autophagy and apoptosis exists;
on the one hand, autophagy occurs before apoptosis and persistent
autophagy can induce cell death and increase apoptosis; on the
other hand, autophagy serves as a protector to prevent cells from
undergoing apoptosis by inhibiting the release of apoptotic factors
(16,17).
TCM has long been widely used for the prevention and
treatment of various diseases, including cancer in China and other
Asian countries. TCM has attracted increasing attention and has
become an emerging field for anticancer drug discoveries and
development due to its wide availability in nature and the fact
that it can be easily obtained; moreover, some active components
isolated and extracted from TCM have exhibited significant
antitumor activity with less side-effects compared to commonly used
chemotherapeutic agents in cancer therapy (3,18,19).
Alpinia katsumadai Hayata (AKH) has long been
widely used as a TCM in China and Korea. AKH was recorded in
Chinese Pharmacopoeia as being pungent in flavor, ‘warm’ in nature,
and attributive to the stomach and spleen meridians (20). AKH is believed to promote qi,
disperse cold, relieve pain, dry dampness and stop vomiting
(20). AKH is commonly used to
treat internal obstruction, abdominal distension, belching and
retching counterflow, as well as to increase appetite (20). Modern pharmacological studies have
demonstrated that AKH has anti-bacterial, antioxidant,
anti-inflammatory and anti-asthmatic activities (6,21).
Previous studies have also proven that the active components of AKH
can induce the apoptosis and autophagy of cancer cells (22,23).
However, studies on the effects of AKH on the proliferation,
apoptosis and autophagy of cancer cells, as well as on the
associated mechanisms are limited.
The present study investigated the in vitro
effects of an ethanol (EtOH) extract of AKH on cell proliferation
using a Cell Counting Kit-8 (CCK-8) assay, apoptosis using Hoechst
33342/propidium iodide (PI) staining and Annexin-V-fluorescein
isothiocyanate (FITC)/PI double staining and autophagy using
Ad-GFP-LC3B transfection in cancer cells, respectively. In
addition, the association between AKH-induced apoptosis and
autophagy was examined using the autophagy inhibitors,
3-methyladenine (3MA, M9281) and Baflomycin A1 (Baf-A1). The
expression levels of cleavage poly(ADP-ribose) polymerase (PARP),
caspase-8, caspase-3 and caspase-9 were examined by western blot
analysis. Furthermore, the in vivo antitumor effects of AKH
were evaluated in the nude mice with A549 lung tumor xenografts.
The components of AKH were detected by liquid chromatography mass
spectrometry (LCMS)-ion trap (IT)-time-of-flight (TOF) mass
spectrometry.
Materials and methods
Preparation of AKH
The seeds of AKH (cat. no. T000500063) were
purchased from Sichuan Hongpu Pharmaceutical Co., Ltd. A total of
10 kg dehydrated powdered seeds of AKH were extracted with 95% EtOH
(Chengdu Chron Chemicals Co., Ltd.) for 48 h at room temperature.
The EtOH extraction was concentrated with a rotary evaporator under
50°C under reduced pressure as previously described (24).
Chemicals and reagents
3MA was purchased from MilliporeSigma. RPMI-1640
medium, fetal bovine serum (FBS) and dimethyl sulfoxide (DMSO) were
purchased from Gibco; Thermo Fisher Scientific, Inc. The Hoechst
33342/PI kit and Baf-A1 were purchased from Beijing Solarbio
Science & Technology Co., Ltd. The Cell Counting Kit (CCK)-8
kit, Ad-GFP-LC3B and actin mouse monoclonal antibody (cat. no.
AF0003) were purchased from the Beyotime Institute of
Biotechnology. The Annexin V-FITC apoptosis detection kit was
purchased from BD Biosciences. The pro-caspase-3 (cat. no.
ab32150), cleaved caspase-3 (cat. no. ab32042), caspase-8 (cat. no.
ab32397), caspase-9 (cat. no. ab32068), PARP (cat. no, ab32138) and
cleaved PARP (cat. no. ab32561) antibodies were purchased from
Abcam. LC3B (cat. no. 2775S), Beclin-1 (cat. no. 3738S), AMPK (cat.
no. 2532S), phosphorylated (p-)AMPK at Ser 485 (cat. no. 4184S),
Akt (cat. no. 9272S), p-Akt at Ser473 (cat. no. 9271S), mTOR (cat.
no. 2972S), p-mTOR at Ser2448 (cat. no. 2971S), p70S6K (cat. no.
9202S) and p-p70S6K at Thr389 (cat. no. 9205S) antibodies were
purchased from Cell Signaling Technology, Inc.
Cells and cell culture
The human lung cancer A549 (cat. no. CCL-185),
breast cancer MDA-MB-468 (cat. no. HTB-132), cervical cancer HeLa
(cat. no. CCL-2), glioblastoma of unknown origin U87 (cat. no.
HTB-14) and colon cancer HCT-116 (cat. no. CCL-247) cell lines were
purchased from the American Type Culture Collection (ATCC). The
human melanoma A875 (cat. no. CL-0255) and normal hepatic stellate
LX-2 (cat. no. CL-0560) cell lines were purchased from Procell Life
Science & Technology. The human pancreatic cancer Panc-28 cell
line was kindly provided by Dr Joshua Liao (University of
Minnesota, Austin, MN, USA). All cancer cells were cultured in
RPMI-1640 medium, whereas the LX-2 cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS
and 1% penicillin/streptomycin and maintained in an atmosphere of
5% carbon dioxide (CO2) at 37°C and renewed with new
medium every 3–5 days. The LX-2 cells were analyzed by short tandem
repeat profiling, cell morphology and karyotyping assay. All cell
lines were verified on the Cellosaurus database and it was
confirmed that they were not considered problematic for use.
Cell viability assay
The effect of AKH on the viability of various cancer
cells and normal hepatic stellate LX-2 cells was evaluated by CCK-8
assay. Briefly, cells (>90% confluency, 5×103
cells/well) were seeded in 96-well culture plates. Following
overnight incubation, the cells were treated with the solvent
solution (vehicle control) or various concentrations of AKH (50,
100, 150, 200, 300 and 400 µg/ml) for 48 h, respectively. For the
study of time-dependent effects of AKH on cell growth inhibition,
the Panc-28 and A549 cells (the two most sensitive cells to AKH
treatment (Table I and Fig. 1A) were seeded (5×103
cells/well) and treated with the vehicle control or AKH at 100,
150, 200, 250, 300 and 400 µg/ml for 24, 48 and 72 h, respectively.
Subsequently, 10% CCK-8 solution (10 µl/well) were added to each
well and the cells were incubated for an additional 1 h at 37°C.
The products of CCK-8 formazan were estimated by measuring the
optical density (OD) at a 450 nm absorbance using a microplate
reader (Model 680, Bio-Rad Laboratories, Inc.). Cell viability was
calculated as the OD of the drug-treated sample/OD of the control
sample ×100%. The values of half inhibitory concentration
(IC50) were calculated by regression analysis using SPSS
20.0 statistical software (IBM Corporation). All experiments were
performed for at least three times in triplicate.
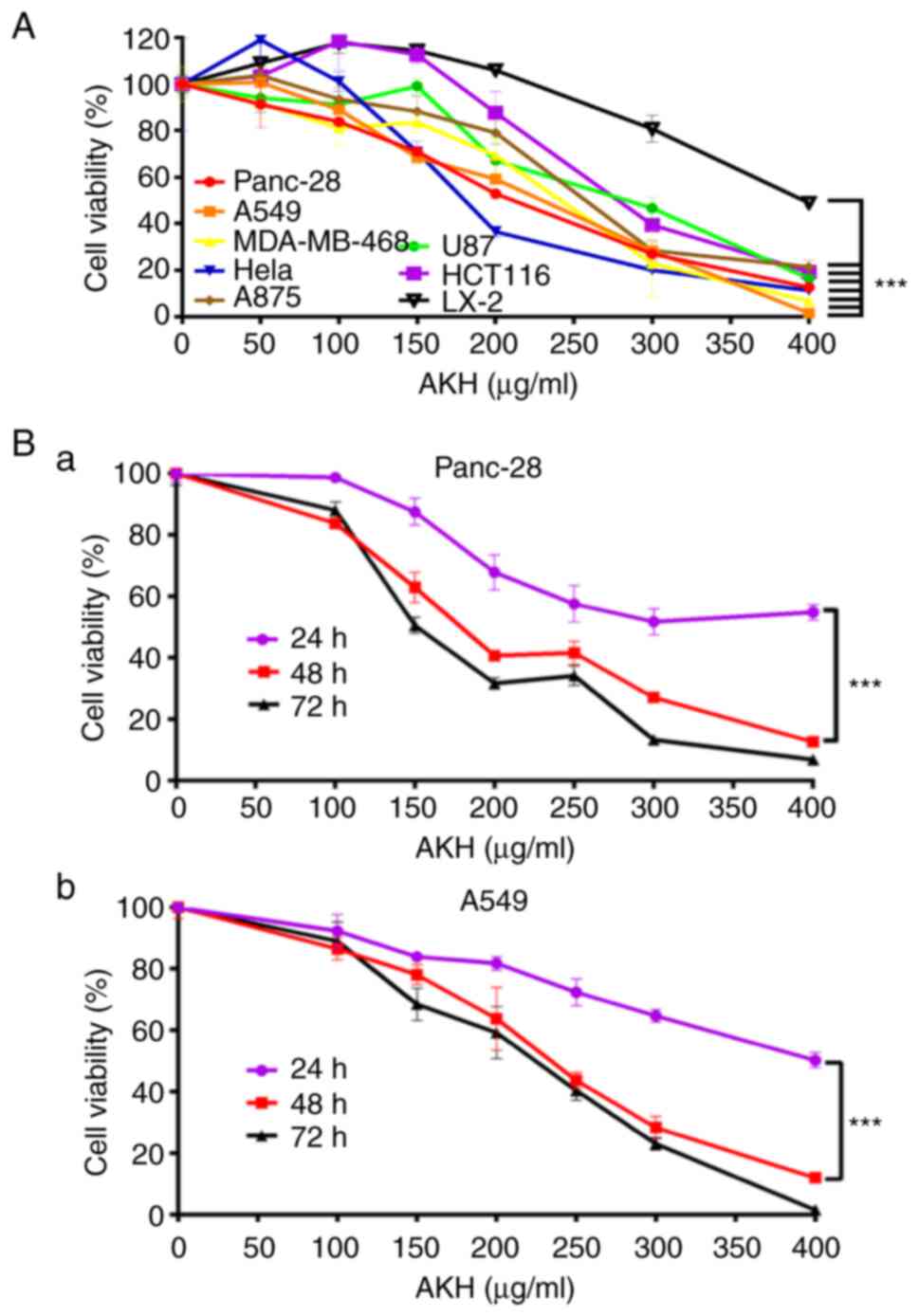 | Figure 1.AKH inhibits the proliferation of
various cancer cells, as demonstrated using CCK-8 assay. (A)
Viability of Panc-28, A549, MDA-MB-468, Hela, A875, U87, HCT-116
and LX-2 cells following treatment with the vehicle control or
various concentrations of AKH (50, 100, 150, 200, 300 and 400
µg/ml) for 48 h. (B) Time-effects of AKH on (a) Panc-28 and (b)
A549 cells. The cells were treated with the vehicle control or AKH
at 100, 150, 200, 250, 300 and 400 µg/ml for 24, 48, or 72 h,
respectively. The data are representative of three independent
experiments (n=3) run in triplicate and are expressed as the mean ±
SD. ***P<0.001 vs. LX-2 cells in A and vs. 24 h in B (determined
using one-way ANOVA). AKH, Alpinia katsumadai Hayata. |
 | Table I.IC50 values of AKH in
various cell lines determined by CCK-8 assay. |
Table I.
IC50 values of AKH in
various cell lines determined by CCK-8 assay.
| Cell line | Cell type | IC50
(µg/ml) |
|---|
| Panc-28 | Human pancreatic
cancer | 202.7±6.4 |
| A549 | Human lung
cancer | 219.0±9.9 |
| MDA-MB-468 | Human breast
cancer | 234.7±8.4 |
| Hela | Human cervical
cancer | 236.2±8.8 |
| A875 | Human melanoma
cancer | 259.6±11.3 |
| U87 | Human glioblastoma
of unknown origin | 262.0±15.9 |
| HCT-116 | Human colon
cancer | 284.0±9.1 |
| LX-2 | Human hepatic
stellate cell | 395.4±2.21 |
Hoechst 33342/PI double staining
assay
Apoptotic cancer cells were initially detected using
the Hoechst 33342/PI double stain apoptosis detection kit (Beijing
Solarbio Science & Technology Co., Ltd.) following the
manufacturer's instructions. Briefly, the Panc-28 or A549 cells
(>90% confluency) were seeded in six-well plates at a density of
3×105 cells/well. The cells were treated with the
vehicle control or AKH at 150, 200 and 250 µg/ml for 48 h following
24 h of culture. The medium was then removed, and the cells were
washed with PBS for three times in 15 min (5 min each). Finally,
the cells were stained with Hoechst 33342/PI kit in the dark for 30
min at room temperature and apoptotic cells were observed under a
Leica DM6 B fluorescence microscope (Leica Microsystems GmbH). All
experiments were performed for at least three times in
triplicate.
Annexin V/PI staining assay
The detection of the apoptosis of Panc-28 and A549
cells was also performed using flow cytometric analysis with the
Annexin V-FITC apoptosis detection kit according to the
manufacturer's protocol as described in a previous study by the
authors (25). Briefly,
~5×105 cells (>90% confluency) were seeded in each
well of six-well culture plates and cultured overnight at 37°C in
an incubator. Various concentrations (150, 200 and 250 µg/ml) of
AKH were added to the plates and incubated at 37°C for 48 h. The
cells were harvested by trypsinization, washed with PBS twice, and
resuspended in 1X binding buffer at a concentration of
1×106 cells. Subsequently, 100 µl (1×106)
cells were mixed with 5 µl of Annexin V-FITC and 5 µl PI and
incubated at room temperature in the dark for 15 min. The samples
(500 µl) were analyzed for apoptotic cells using a flow cytometer
(Becton, Dickinson and Company). All experiments were performed for
at least three times in triplicate.
Ad-GFP-LC3B transfection
To detect the formation of autophagosomes, the
Panc-28 and A549 cells were transfected with Ad-GFP-LC3B (Beyotime
Institute of Biotechnology) following the manufacturer's
instructions. Briefly, the cells (>90% confluency) were seeded
in a 24-well plate at a density of 5×104 cells/well and
transfected with Ad-GFP-LC3B at a multiplicity of 40 with
2×106 plaque forming units (pfu) adenovirus for 24 h at
room temperature. The cells were then treated with medium (control)
or AKH (250 µg/ml) for 24 h, respectively. They were then fixed
with 4% polyoxymethylene and observed under a Leica DM6 B
fluorescence microscope at ×40 magnification (Leica Microsystems
GmbH). All experiments were performed for at least three times in
triplicate.
Western blot analysis
The Panc-28 and A549 cells (>90% confluency) were
seeded in six-well plates at a density of 5×105
cells/well. Following 24 h of incubation, the cells were treated
with the solvent solution or AKH at 150, 200 and 250 µg/ml for 48
h. The cells were then collected by centrifugation at 1,500 × g at
room temperature for 5 min and lysed in ice-cold RIPA lysate buffer
as described in a previous study by the authors (25). The protein concentrations were
determined using the BCA method. Subsequently, equal amounts of
proteins (30–40 µg) were separated by 10% sodium dodecyl sulphate
(SDS)-polyacrylamide gel electrophoresis (PAGE) and transferred
onto polyvinylidene fluoride (PVDF) membranes (EMD Millipore).
Non-specific binds were blocked with 5% non-fat dry milk in 1X TBST
buffer (1X TBS and 0.1% Tween-20) for 1 h at room temperature and
subsequently incubated with the corresponding primary antibodies
(1:1,000) of caspase-8, caspase-3, caspase-9, PARP, LC3B, Beclin-1,
Akt, p-Akt, mTOR, p-mTOR, AMPK, p-AMPK, p70S6K, p-p70S6K or actin
overnight at 4°C. The membranes were washed four times (10 min
each) with 1X TBST buffer and incubated with horseradish peroxidase
(HRP)-conjugated goat anti-mouse (cat. no. A0216) or anti-rabbit
secondary antibodies (cat. no. A0208, Beyotime Institute of
Biotechnology) at 1:1,000 for 1 h at room temperature on a shaker
and washed four times with 1X TBST buffer for 40 min. Actin was
used as a loading control. Finally, the membranes were developed
and visualized using the ECL Western Blotting detection kit (cat.
no. 32109, Thermo Fisher Scientific Inc.) after washing with 1X
TBST four times. The OD values of the blots were measured using
ImageJ software version 1.52a (National Institutes of Health). All
samples were performed for three times in triplicate.
Antitumor activity of AKH against A549
tumor xenografts in nude mice in vivo
Female athymic Balb/C nude mice (6–8 weeks old; body
weight, 18–20 g) were purchased from Chengdu Dasuo Laboratory
animal Co., Ltd. and housed in a specific pathogen-free (SPF)
facility with a constant laboratory condition of a 12-h light/dark
cycle and provided with food and water ad libitum. A549
tumor xenografts were initially established in nude mice by a
subcutaneous (s.c.) injection of cultured A549 cells
(1×107 cells/ml in the logarithmical stage with >90%
confluency) in 150 µl of serum-free RPMI-1640 medium into the right
flanks of the mice. The tumor xenografts were subsequently passed
several generations by transplantation of ~50 mg non-necrotic tumor
tissues into the right flanks of mice with the help of a trocar
prior to treatment, which was initiated 2 weeks later when the
tumors reached ~100 mm3 (mg) as previously described
(26,27). The mice were randomly divided into
four groups with 5 mice in each group and treated as follows: i)
Normal saline (negative vehicle control); ii) cisplatin (Jiangsu
Hansoh Pharmaceutical Co., Ltd.) 5 mg/kg (positive control); iii)
AKH at 100 mg/kg; and iv) AKH at 400 mg/kg. The powder of AKH EtOH
extract was first dissolved in normal saline and the solution was
administered to the mice so there was no converted into an EtOH
extract in the body. AKH and normal saline were orally administered
by gavage once a day for 12 consecutive days and cisplatin by
intraperitoneal (i.p.) injection every 3 days (on days 0, 3, 6, 9
and 12, for a total of five doses) with an ~0.2 ml volume per 20 g
mouse weight (28). The body
weights of the mice and tumor volumes were monitored every 3 days
during treatment. The tumors were measured by the longest axis (L)
and shortest axis (W) with the help of a Vernier caliper and the
tumor volume (mm3) or weight (mg) was calculated using
the following formula: 1/2 (L × W2) (mm3)
(26,28). At the end of the experiment (24 h
after the final treatment) for a duration of 4 weeks, all mice were
euthanized by rapid cervical dislocation, and the tumors were
removed and weighed to obtain images and calculate the rate of
tumor inhibition. The specific criteria used to determine
euthanasia timepoints for the mice was when the tumor reached
~2,000 mg (mm3) or 20% of body weight loss. Death was
verified by monitoring breathing, heartbeat, flexor reflex and
corneal contact response. The tumor growth inhibition (TGI) was
calculated by the mean tumor volume (weight) of the treatment group
(TG)/relative to the control group (CG) using the following
formula: (MTWTG-MTWCG) ÷ MTWCG ×100% (26,27).
All animal experiments were approved (permit no.
201802-112) by the Institutional Animal Care and Use Committee of
Southwest Medical University (Luzhou, China) and strictly followed
the guidelines for the investigation of experimental pain in
conscious animals for improving animal welfare to minimize animal
suffering (29).
Analysis of the composition of AKH by
LCMS-IT-TOF assay
LC-MS analysis of the composition of AKH was
performed using a Shimadzu LCMS-IT-TOF mass spectrometer (Shimadzu
Corp.). The samples of AKH were separated on an Agilent Eclipse
plus C18 column (100×2.1 mm i.d. 1.8 µM particle size; Agilent
Technologies, Inc.). The separation process was followed the
gradient elution procedure. Chromatographic analysis was used the
mobile phase A, which was composed of acetonitrile modified with
0.5% formic acid, while the mobile phase B was composed of water
modified with 0.5% formic acid. The linear gradient was as
A:B=40–90% for 24 min. The flow rate was 0.2 ml/min and the column
temperature was 30°C. The injection volume was 5 µl. For mass
detection, the following parameters were used for analytical MS:
Nozzle voltage, 4.5 KV (+)/-3.5 KV (−) in the detection modes of
positive ion and negative ion; electrospray ionization (ESI)
voltage, 1.65 KV; nebulizing gas (N2) flow rate, 1.5 l/min; and
drying gas flow rate, 10.0 l/min. The desolation line was heated to
250°C and the heat block was heated to 450°C. Collision-induced
dissociation gas pressure was set to 230 kPa. The MS data were
acquired from m/z 100 to 2,000. The data were analyzed using Lab
Solutions software (version 5.75, Shimadzu Corp.).
Statistical analysis
Data were analyzed using SPSS 20.0 statistical
software (IBM Corporation) and presented as the mean ± standard
deviation (SD). One-way analysis of variance (ANOVA) and the
univariate analysis of general linear model were used to determine
statistical significance. Tukey's test was applied as the post hoc
test. The data of western blot analysis were quantified using Image
J software version 1.52a (National Institute of Health). A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
AKH inhibits the proliferation of
various cancer cells
The present study first investigated the inhibitory
effects of AKH at multiple concentrations (0, 50, 100, 150, 200,
300 and 400 µg/ml) on the proliferation of various cancer cells and
compared to normal hepatic stellate LX-2 cells by CCK-8 assay. The
results revealed that AKH significantly (P<0.001) inhibited the
growth of various cancer cells in a concentration-dependent manner;
however, it exerted a weak inhibitory effect on the LX-2 cells
(Fig. 1A). The IC50
values of AKH were 202.7±6.4, 219.7±9.9, 234.7±8.4, 236.2±8.8,
259.6±11.3, 262.0±15.9, 284.0±9.1 and 395.4±2, 21 µg/ml for the
Panc-28, A549, MDA-MB-468, HeLa, A875, U87, HCT-116 and LX-2 cells,
respectively (Table I). These
results indicated that AKH exerted the most potent cytotoxic effect
on the Panc-28 and A549cells. Subsequently, the time-dependent
effects of AKH on the growth inhibition in Panc-28 and A549 cells
were examined by treating the cells with AKH at 0 (control), 100,
150, 200, 250, 300 and 400 µg/ml for 24, 48 and 72 h, respectively.
The results revealed that the inhibitory effects of AKH on the
Panc-28 and A549 cells were time-dependent; AKH was more effective
at 48 and 72 h than at 24 h (P<0.001); no statistically
significant difference was observed between the 48 and 72 h time
points (P>0.05), although the inhibitory effect of AKH was the
most prominent at 72 h (Fig. 1Ba and
Bb).
AKH induces the apoptosis of Panc-28
and A549 cells
Apoptosis is closely related to the fate of cancer
and is one of the key targets for novel anticancer drug discovery
and development (19). Therefore,
the present study examined the effects of AKH on the apoptosis of
Panc-28 and A549 cells using Hoechst 33342/PI staining assay. The
results demonstrated that treatment with AKH at 150, 200 and 250
µg/ml for 48 h significantly induced cellular apoptosis in a
concentration-dependent manner; the cells displayed typical
morphological features of apoptosis, including chromatin
condensation, nuclear shrinkage, and the formation of apoptotic
bodies (Fig. 2A and B).
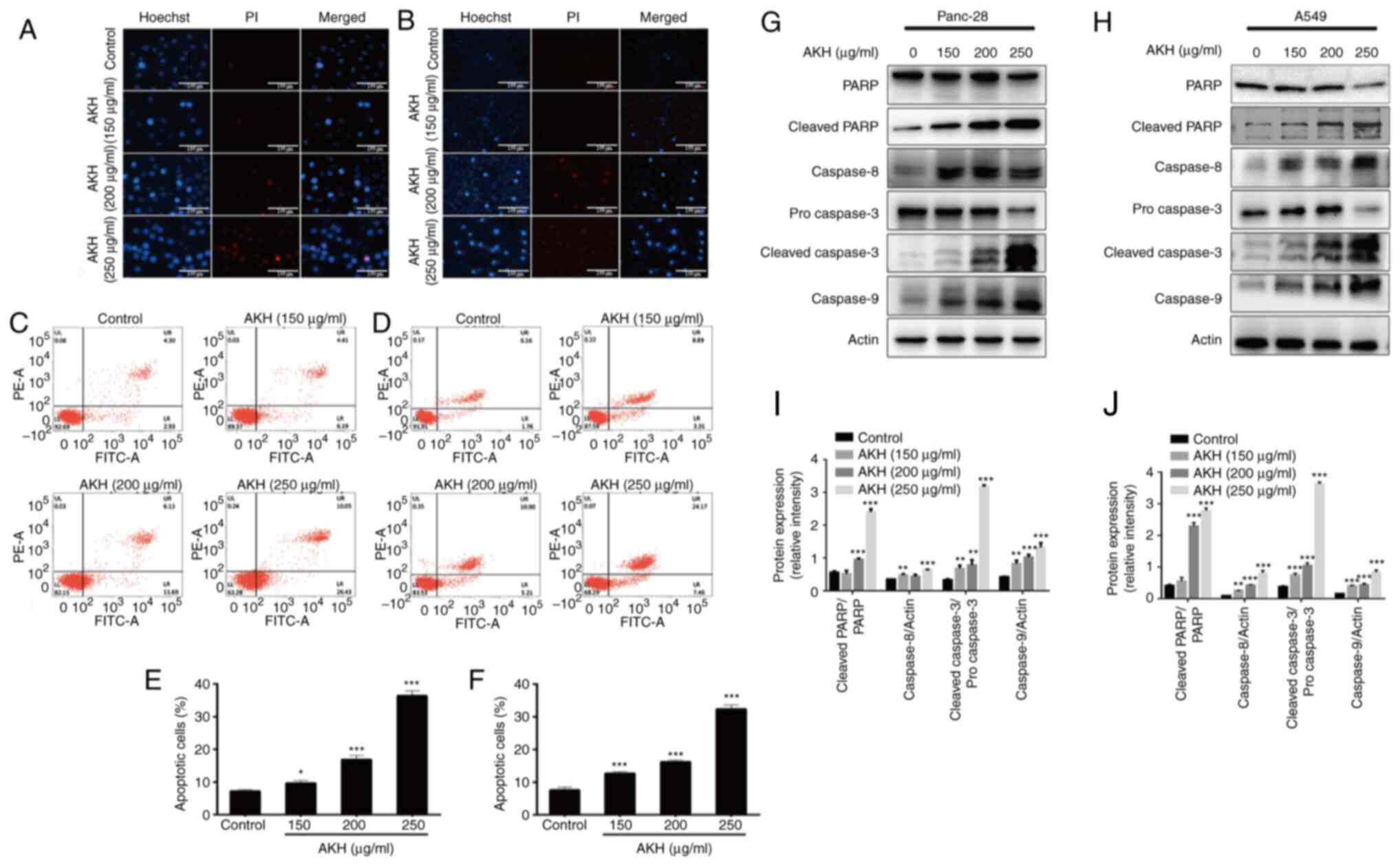 | Figure 2.Effects of AKH on apoptosis and the
expression of apoptosis-related proteins in Panc-28 and A549 cells.
(A) The morphological features of apoptosis induced by AKH in
Panc-28 by Hoechst 33342/PI staining assay. (B) The morphological
features of apoptosis induced by AKH in A549 cells by Hoechst
33342/PI staining assay. (C) Analysis of apoptosis of Panc-28 cells
examined by flow cytometry with Annexin-V-FITC/PI double staining
assay. (D) Analysis of apoptosis of A549 cells by flow cytometry
with Annexin-V-FITC/PI double staining assay. (E) Percentage of
apoptotic Panc-28 cells. (F) Percentage of apoptotic A549 cells.
(G) The expression of apoptosis-related proteins PARP,
cleaved-PARP, caspase-8, pro-caspase-3, cleaved-caspase-3 and
caspase-9 in Panc-28 cells by western blot analysis. (H) The
expression of apoptosis-related proteins PARP, cleaved-PARP,
caspase-8, pro-caspase-3, cleaved-caspase-3 and caspase-9 in A549
cells by western blot analysis. (I) The protein expression of
cleaved-PARP/PARP, caspase-8, cleaved-caspase-3/pro-caspase 3 and
caspase-9 in Panc-28 cells. (J) The protein expression of
cleaved-PARP/PARP, caspase-8, cleaved-caspase-3/pro-caspase-3 and
caspase-9 in A549 cells. Actin was used as a loading control. The
cells were treated with the vehicle control or AKH at 150, 200 and
250 µg/ml for 48 h. The data are representative of three
independent experiments (n=3) run in triplicate and are expressed
as the mean ± SD. *P<0.05, **P<0.01 and ***P<0.001 vs.
vehicle control (determined using one-way ANOVA). AKH, Alpinia
katsumadai Hayata; PARP, poly(ADP-ribose)polymerase. |
For the quantitative analysis of the effects of AKH
on apoptosis induction, flow cytometric analysis was then performed
using Annexin-V-FITC/PI double staining assay in the Panc-28 and
A549 cells. The data demonstrated that treatment with AKH for 48 h
markedly induced apoptosis in a concentration-dependent manner; the
apoptotic rates were 7.23±0.45 and 7.58±0.93%, 9.68±0.73 and
12.68±0.53%, 16.89±1.22 and 16.23±0.52%, and 36.45±1.65 and
32.35±.26% for the Panc-28 and A549 cells treated with the vehicle
(control), or 150, 200 and 250 µg/ml of AKH, respectively (Fig. 2C-F).
To further investigate the underlying mechanisms of
the AKH-induced apoptosis of the cancer cells, the expression of
cleaved PARP, caspase-8, caspase-3 and caspase-9 was examined using
western blot analysis of the Panc-28 and A549 cells. The results
revealed that the relative expression levels of the apoptotic
proteins, cleaved PARP, caspase-8, cleaved caspase-3 and caspase-9,
were significantly increased in a concentration-dependent manner
following treatment with AKH (Fig. 2G
and H). These data indicate that AKH markedly induced apoptosis
and that apoptosis plays a crucial role in the inhibitory effects
of AKH on the proliferation of cancer cells.
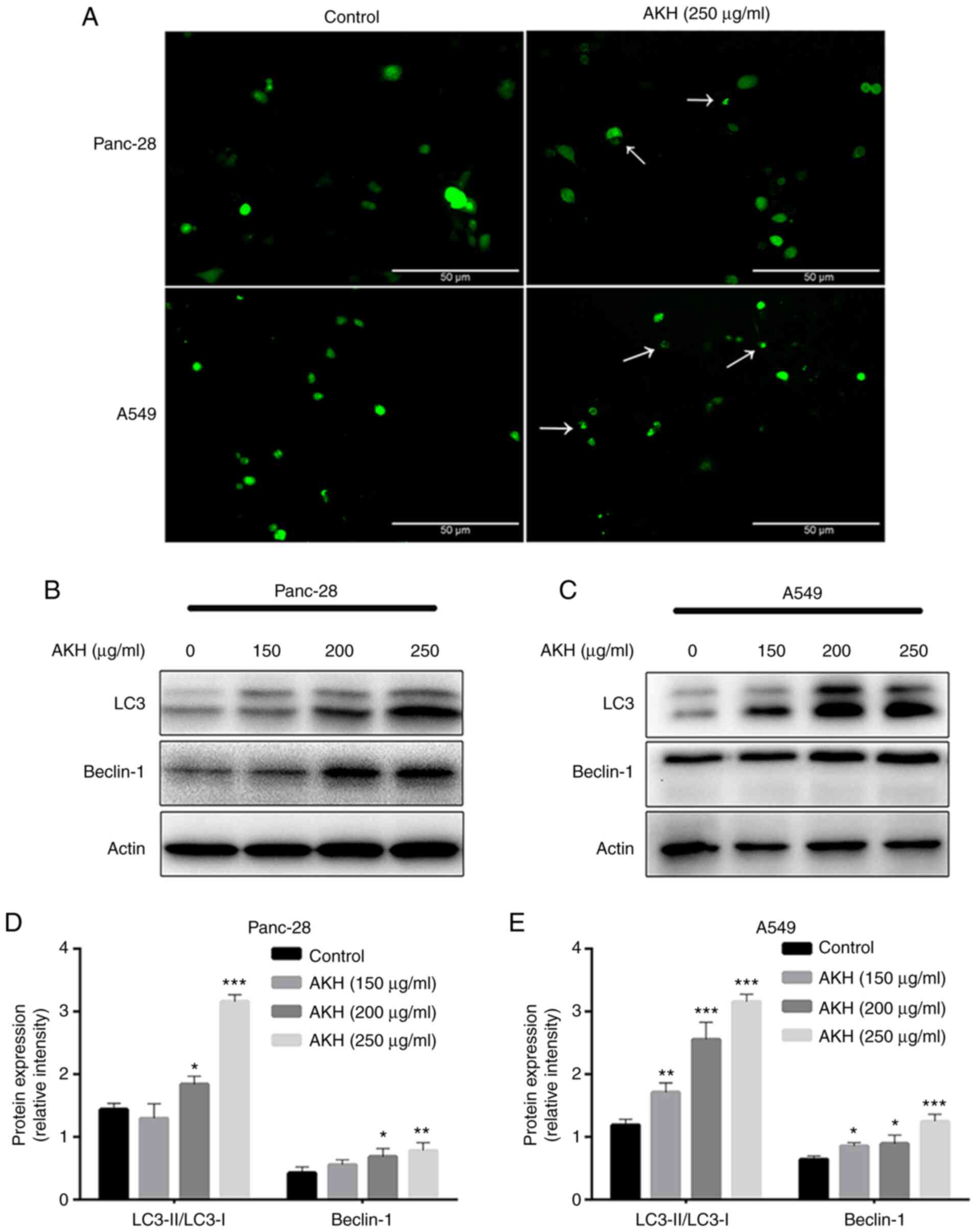
AKH induces autophagy and autophagy
plays a role in the regulation of the apoptosis of Panc-28 and A549
cells
Autophagy plays a dual role in cancer initiation and
progression, autophagy and apoptosis are highly interconnected in
determining the fate of cancer cells (30,31).
Therefore, the present study evaluated the effects of AKH on
autophagy induction and the role of AKH-induced autophagy in the
regulation of apoptosis. To examine the effects of AKH on
autophagy, Panc-28 and A549 cells were transfected with Ad-GFP-LC3B
and treated with the vehicle solution (control) and AKH (250 µg/ml)
for 24 h. The results revealed that there were diffuse green spots
in the control cells and evident green autophagy spots in the
AKH-treated cells (Fig. 3A),
indicating that AKH induced autophagy. The present study then
further detected LC3-II formation after the transfected cells were
treated with various concentrations of AKH (150, 200 and 250 µg/ml)
for 48 h. The results revealed that the conversion of LC3-II and
the expression of Beclin-1 were markedly enhanced in a
concentration-dependent manner in the Panc-28 and A549 cells
(Fig. 3B-E).
To further investigate the association between
AKH-induced autophagy and apoptosis, the Panc-28 and A549 cells
were pre-treated with the autophagy inhibitors, 3MA (5 mM) or
Baf-A1 (10 nM), for 24 h followed by treatment with AKH (250 µg/ml)
for 48 h. The cells were then examined by flow cytometry and
western blot analysis. The results of CCK-8 assay revealed that
both the autophagy inhibitors, 3MA and Baf-A1, significantly
decreased the inhibitory effects of AKH on the viability of the
Panc-28 (P<0.001; Fig. 4A) and
A549 (P<0.05 or P<0.01; Fig.
4B) cells. The flow cytometry data revealed that both 3MA and
Baf-A1 significantly inhibited the apoptosis induced by AKH in the
Panc-28 (P<0.01 or P<0.001; Fig.
4C) and A549 (P<0.05; Fig.
4D) cells. Moreover, the data of western blot analysis also
demonstrated that 3MA and Baf-A1 markedly reduced the expression
level of apoptosis- and autophagy-related proteins, such as cleaved
PARP, pro- and cleaved caspase-3 and Beclin-1 in the Panc-28 and
A549 cells (Fig. 4E and F).
However, 3MA decreased the expression of LC3 due to inhibiting
autophagosome formation; however, Baf-A1 increased the expression
of LC3 by preventing lysosome degradation (Fig. 4E and F).
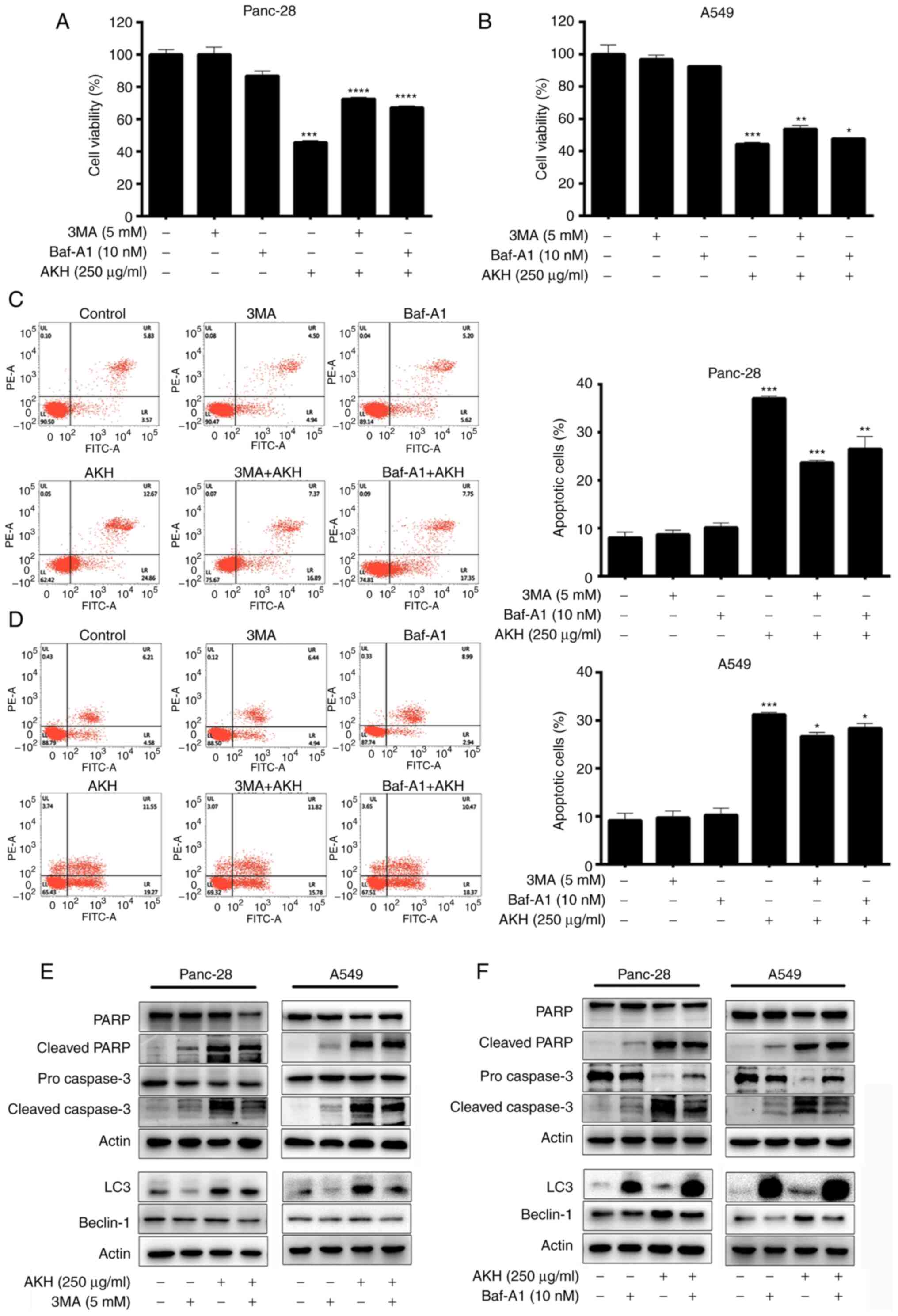 | Figure 4.Effects of the autophagy inhibitors,
3MA and Baf-A1, on AKH-induced cell growth inhibition, apoptosis,
and the expression of apoptosis- and autophagy-related proteins in
Panc-28 and A549 cells. (A) The viability of the Panc-28 cells by
CCK-8 assay. (B) The viability of the A549 cells by CCK-8 assay.
(C) Flow cytometric analysis of Panc-28 cells. (D) Flow cytometric
analysis of A549 cells. (E) The bands of apoptosis-related proteins
PARP, cleaved-PARP, pro-caspase-3 and cleaved-caspase-3;
autophagy-related proteins LC3, Beclin-1 and actin following
treatment of 3MA and AKH alone or in combination in Panc-28 and
A549 cells by western blot analysis. (F) The bands of
apoptosis-related proteins PARP, cleaved-PARP, pro-caspase-3 and
cleaved-caspase-3, autophagy-related proteins LC3, Beclin-1 and
actin following treatment of Baf-A1 and AKH alone or in combination
in Panc-28 and A549 cells by western blot analysis. Actin was used
as a loading control. The cells were pre-treated with the vehicle
control, 3MA (5 mM) or Baf-A1 (10 nM) for 24 h followed by the
vehicle control or AKH (250 µg/ml) for 48 h. The data are
representative of three independent experiments (n=3) run in
triplicate and are expressed as the mean ± SD. *P<0.05,
**P<0.01 and ***P<0.001 vs. vehicle control (determined using
one-way ANOVA). AKH, Alpinia katsumadai Hayata; PARP,
poly(ADP-ribose)polymerase; 3MA, 3-methyladenine; Baf-A1,
baflomycin A1. |
Effects of AKH on AMPK and
Akt/mTOR/p70S6K in Panc-28 and A549 cells
AMPK and Akt/mTOR/p70S6K play a critical role in
regulating both the apoptosis and autophagy of cancer cells
(31–34). Therefore, the present study
investigated the effects of AKH on the expression of proteins
related to the AMPK and Akt/mTOR/p70S6K signaling pathways in
Panc-28 and A549 cells by western blot analysis. The data
demonstrated that AKH significantly increased the levels of p-AMPK
and decreased those of p-Akt (P<0.01 or P<0.001) and
subsequently decreased the levels of downstream target proteins of
Akt, such as p-mTOR and p-70S6K (P<0.01 or P<0.001) in a
concentration-dependent manner in both the Panc-28 and A549 cells
(Fig. 5).
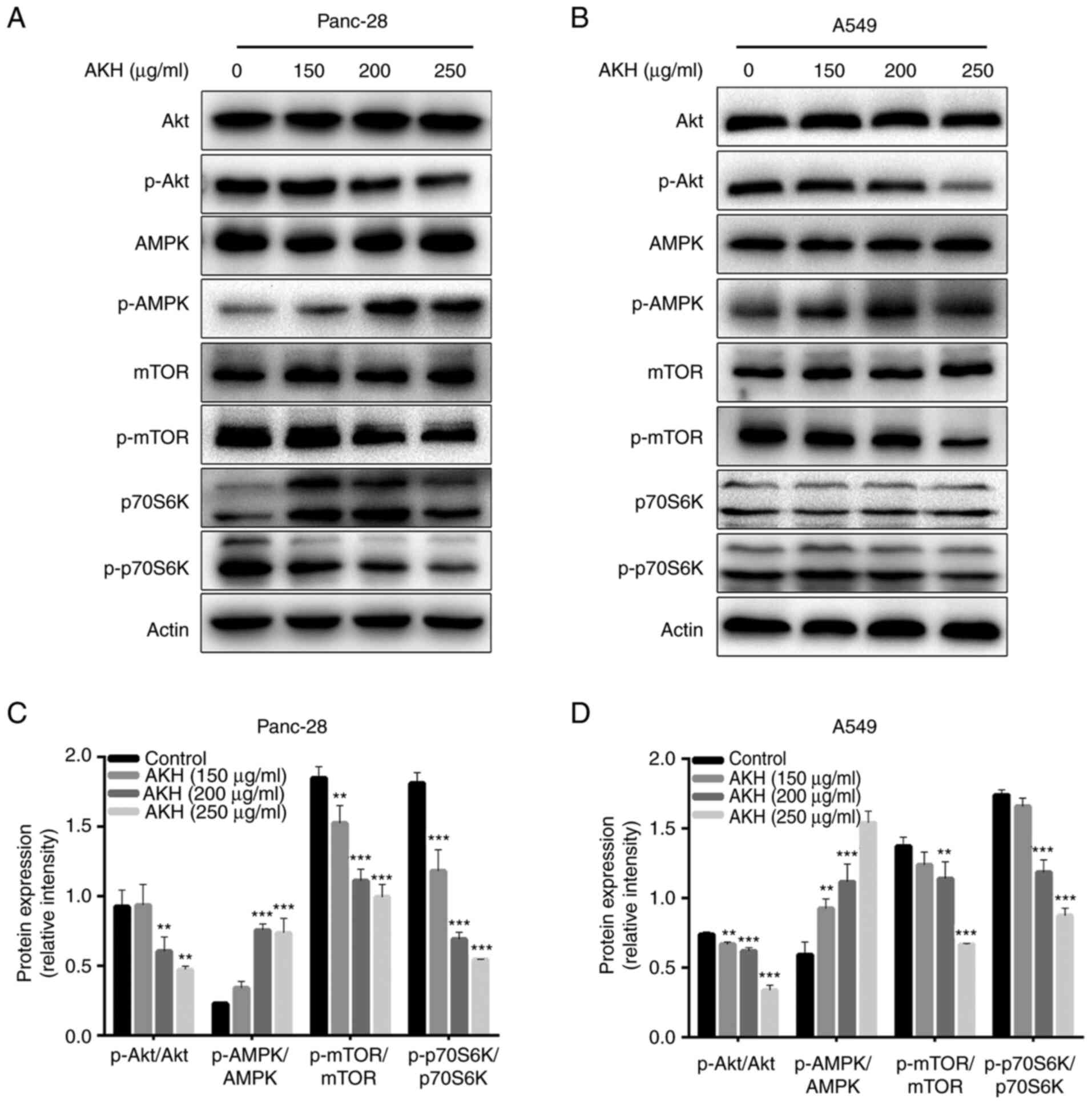 | Figure 5.The effect of AKH on the expression
of proteins related to the AMPK and Akt/mTOR/p70S6K singling
pathways in Panc-28 and A549 cells by western blot analysis. (A)
Bands of Akt, pAkt, AMPK, pAMPK, mTOR, pmTOR, 70S6K p70S6K and
Actin in Panc-28 cells. (B) Bands of Akt, pAkt, AMPK, pAMPK, mTOR,
pmTOR, 70S6K p70S6K and Actin in A549 cells. Actin was used as a
loading control. (C) Relative protein expression of pAkt/Akt,
pAMPK/AMPK, pmTOR/mTOR and p70S6K/70S6K in Panc-28 cells. (D)
Relative protein expression of pAkt/Akt, pAMPK/AMPK, pmTOR/mTOR and
p-p70S6K/p70S6K in A549 cells. The cells were treated with the
vehicle control or AKH at 150, 200, and 250 µg/ml for 48 h. The
data are representative of three independent experiments (n=3) run
in triplicate and are expressed as the mean ± SD. **P<0.01 and
***P<0.001 vs. vehicle control (determined using one-way ANOVA).
AKH, Alpinia katsumadai Hayata; AMPK, AMP-activated protein
kinase. |
These data indicated that AKH significantly
increased the levels of p-AMPK and decreased those of p-Akt, and
its downstream target proteins, such as p-mTOR and p-70S6K in the
Panc-28 and A549 cells.
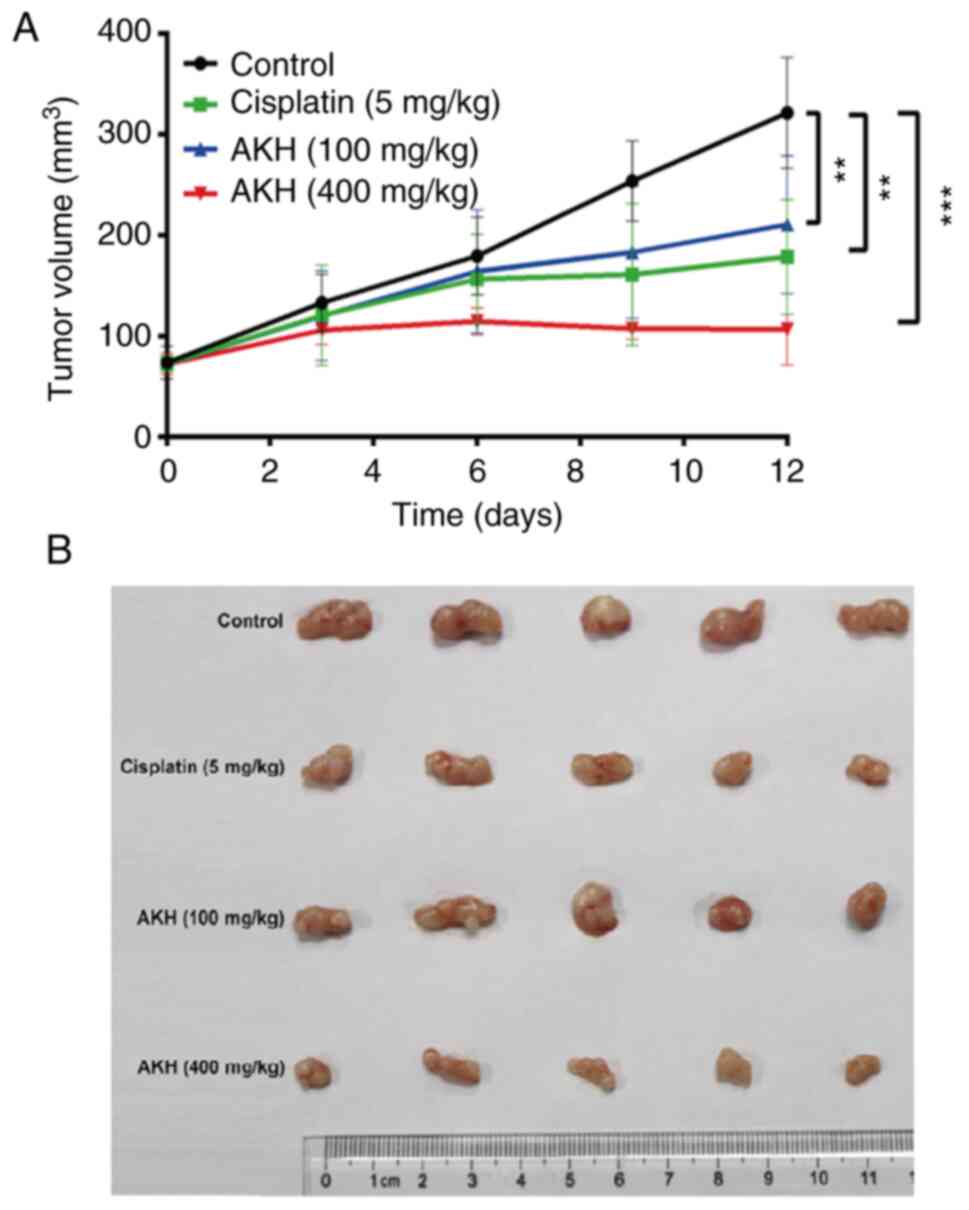
AKH inhibits the growth of A549 tumor
xenografts in nude mice in vivo
After completing the experiments to determine the
effects of AKH on the growth inhibition and apoptosis/autophagy
induction of the Panc-28 and A549 cells in vitro, the
present study further evaluated the in vivo antitumor
activity of AKH (100 and 400 mg/kg) compared to that of normal
saline (negative vehicle control) and cisplatin (5 mg/kg, positive
control) in nude mice bearing A549 tumor xenografts; the results
are illustrated in Fig. 6. The
data indicated the kinetics of tumor growth (Fig. 6A) and the images of tumors
(Fig. 6B) at the end of experiment
following the completion of treatment with normal saline, cisplatin
or AKH in nude mice bearing A549 tumor xenografts. The results
indicated that while normal saline had no significant antitumor
effect, cisplatin and AKH significantly inhibited the growth of
A549 tumor xenografts (P<0.01 or P<0.001). The mean tumor
inhibitory rates were 50.40±8.23, 42.40±9.87 and 72.72±4.96% for
cisplatin, 100 mg/kg AKH and 400 mg/kg AKH, respectively. Of note,
the antitumor effects of AKH occurred in a dose-dependent manner
and the antitumor efficacy of high-dose AKH (400 mg/kg) was
significantly more prominent than that of cisplatin (P<0.01).
Furthermore, no treatment-related death occurred, and the body
weight loss of the mice was much less with AKH treatment (<10%)
compared to that of cisplatin treatment (>15%; data not shown).
These data indicated that AKH was effective against A549 tumor
xenografts and safe to the hosts with better antitumor activity and
less toxicity compared to that of cisplatin in vivo.
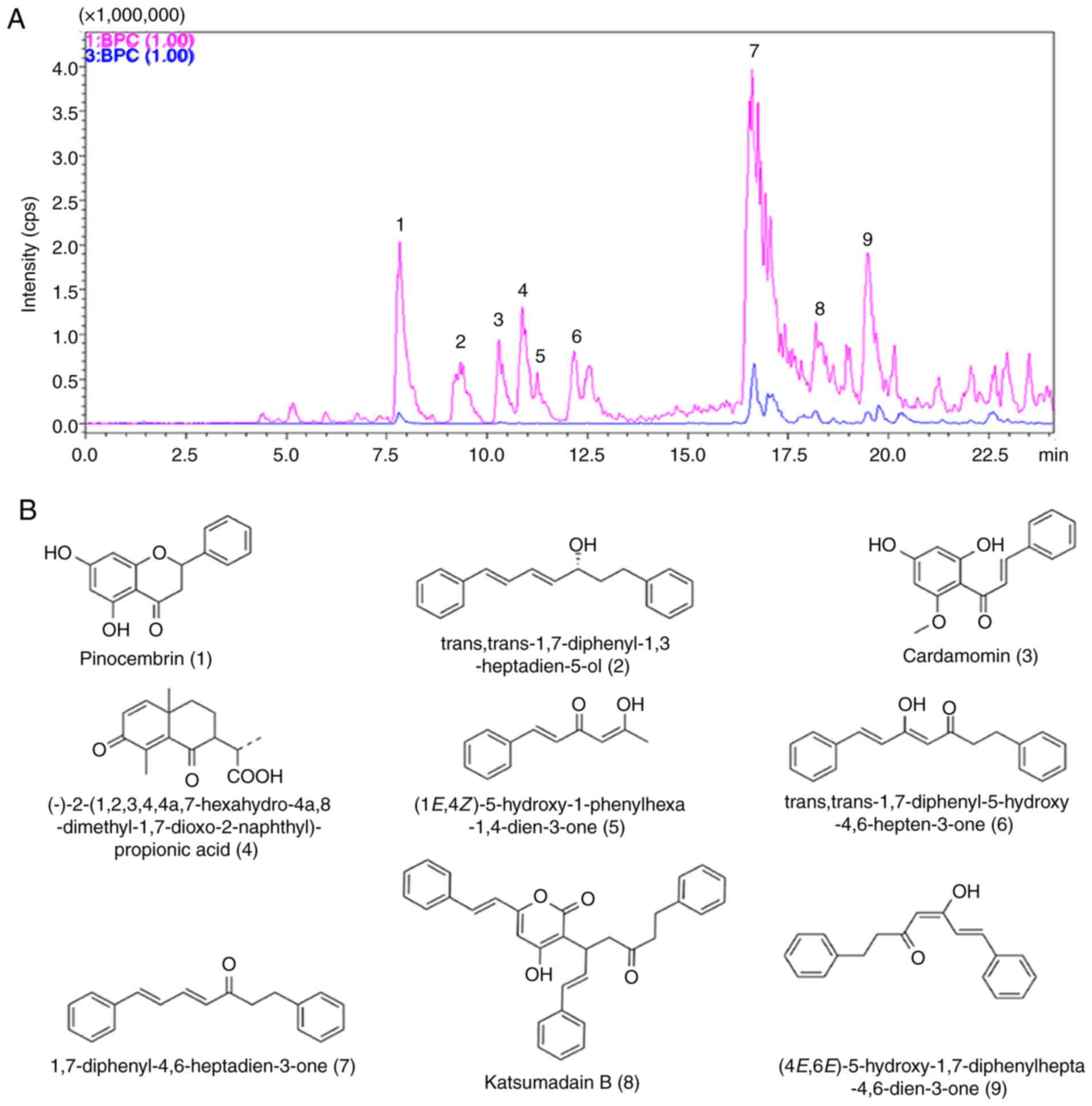
Analysis of the compositions of AKH by
LCMS-IT-TOF
Finally, the present study determined the
compositions of AKH using LCMS-IT-TOF assay with 0.5% formic acid
in water and acetonitrile for separation. A total of nine
components were detected from AKH using this assay (Fig. 7A). The nine compounds on their UV
spectra were identified as pinocembrin,
trans,trans-1,7-diphenyl–1,3-heptadien-5-ol,cardamomin,(−)-2-(1,2,3,4,4a,7-hexahydro-4a,8-dimethyl-1,7-dioxo-2-naphthyl)-propionic
acid, (1E,4Z)–5-hydroxy–1–phenylhexa-1,4-dien-3-one,
trans,trans-1,7-diphenyl–5-hydroxy–4,6-hepten-3-one,
1,7-diphenyl-4,6-heptadien-3-one, katsumadain B and
(4E,6E)-5-hydroxy-1,7-diphenylhepta-4,6-dien-3-one, respectively.
The chemical structures and characteristics of compounds identified
from AKH are summarized in (Fig.
7B and Table II).
| Table II.Analysis of the characterized
compounds from AKH. |
Table II.
Analysis of the characterized
compounds from AKH.
| Compound no. | Retention time
(min) | Molecular weight
(Da) | Molecular
formula | Unsaturation | MS fragmentation
(error in mDa) | Ultraviolet
absorption (λmax) nm | Structure or type
of compound |
|---|
| 1 | 7.78 | 256 |
C15H12O4 | 10 | Pos: 257.0889
[(M+H)+, 8.1] | 216, 289 | Pinocembrin |
|
|
|
|
|
| Neg: 255.0680
[(M-H)−, 1.7] |
|
|
| 2 | 9.33 | 264 |
C19H20O | 10 | Pos: 265.1605
[(M+H)+, 1.8] | 250, 331 |
Trans,trans-1,7-diphenyl-1,3-heptadien-5-ol |
|
|
|
|
|
| MS/MS 247.1500
[(M+H)+, 1.9] |
|
|
| 3 | 10.28 | 270 |
C16H14O4 | 10 | Pos: 271.0995
[(M+H)+,3.0] | 215, 344 | Cardamomin |
|
|
|
|
|
| Neg:269.0787
[(M-H)-, −3.2] |
|
|
| 4 | 10.92 | 262 |
C15H18O4 | 7 | Pos: 263.1350
[(M+H)+, 7.2] | 218, 250 |
(−)-2-(1,2,3,4,4a,7-Hexahydro-4a,8-dimethyl-1,7-dioxo-2-naphthyl)-propionic
acid |
| 5 | 11.26 | 188 |
C12H12O2 | 7 | Pos: 189.0877
[(M+H)+, 3.3] | 225, 340 |
(1E,4Z)-5-Hydroxy-1-phenylhexa-1,4-dien-3-one |
| 6 | 12.17 | 278 |
C19H18O2 | 11 | Pos: 279.1347
[(M+H)+, 2.0] | 219, 331 |
Trans,trans-1,7-diphenyl-5-hydroxy-4,6-hepten-3-one |
| 7 | 16.65 | 262 |
C19H18O | 11 | Pos: 263.1498
[(M+H)+, 6.8] | 231, 355 |
1,7-Diphenyl-4,6-heptadien-3-one |
| 8 | 18.18 | 476 |
C32H28O4 | 19 | Pos: 477.2004
[(M+H)+, −5.6] | 229, 325 | Katsumadain B |
|
|
|
|
|
| Neg:475.1913
[(M-H)−, −0.2] |
|
|
| 9 | 19.51 | 278 |
C19H18O2 | 11 | Pos: 279.1346
[(M+H)+, 2.0] | 229, 361 |
(4E,6E)-5-Hydroxy-1,7-diphenylhepta-4,6-dien-3-one |
|
|
|
|
|
| Neg: 277.1174
[(M-H)−, −6.0] |
|
|
Discussion
Cancer is one of the most critical public health
concerns and a leading cause of morbidity and mortality worldwide
(2). It caused almost 10 million
deaths worldwide in 2020 (35).
Lung cancer is the second most diagnosed cancer with an estimated
2.2 million new cases and the leading cause of cancer death (1.8
million) globally in 2020 (35).
Pancreatic cancer has a poor prognosis and a low survival rate,
with 466,000 deaths reported 2020 worldwide (35). It has been indicated that only ~15%
of patients with pancreatic cancer are diagnosed at the early
stages upon disease progression (36). Therefore, the present study focused
on the anticancer effects of AKH and associated molecular
mechanisms in A549 lung cancer and Panc-28 pancreatic cancer
cells.
Firstly, the present study investigated the
anticancer effects of AKH in seven cancer cell lines, including
Panc-28, A549, MDA-MB-468, Hela, A875, U87 and HCT-116, and
compared these to those in normal human hepatic stellate LX-2 cells
using CCK-8 assay. The results revealed that AKH displayed potent
cytotoxicity against the tested cancer cells with IC50
values of 203–284 µg/ml; the Panc-28 and A549 cells were the most
sensitive cells as regards the response to AHK treatment (Fig. 1A and Table I). Notably, AHK was much less
cytotoxic against normal liver LX-2 cells with an IC50
of value of 395 µg/ml (Fig. 1A and
Table I). These results indicated
that AKH could selectively inhibit cancer cells. Subsequently, the
time-dependent effects of AKH on the growth inhibition in Panc-28
and A549 cells were examined following treatment with AKH at 0
(control), 100, 150, 200, 250, 300 and 400 µg/ml for 24, 48 and 72
h, respectively. The results revealed that the inhibitory effects
of AKH occurred in a concentration- and time-dependent manner; AKH
was more effective at 48 and 72 h than at 24 h (P<0.001) in the
Panc-28 and A549 cells (Fig. 1Ba and
Bb).
Apoptosis and autophagy play a critical role in the
fate of cancer and cancer therapy (19,31,37).
Previous studies have demonstrated that apoptosis and autophagy are
highly connected, and their association is very complex (28,35).
Some active components from TCM have been proven to be effective
against various types of cancer by inducing apoptosis and
autophagy-related apoptosis (19,31,38–41).
The present study found that AKH induced typical morphological
features of apoptosis, such as chromatin condensation, nuclear
shrinkage and apoptotic body formation in the Panc-28 and A549
cells by Hoechst 33342/PI staining assay (Fig. 2A and B). AKH also significantly
increased the apoptotic rate in a concentration-dependent manner in
the Panc-28 and A549 cells by quantitative analysis of flow
cytometry with Annexin V-FITC/PI double staining assay (Fig. 2C-F). Furthermore, AKH significantly
increased the expression of apoptosis-related proteins, such as
cleaved PARP, caspase-8, cleaved caspase-3 and caspase-9 in a
concentration-dependent manner in the Panc-28 and A549 cells, as
demonstrated by western blot analysis (Fig. 2G and H). Moreover, treatment with
AKH (150–250 µg/ml) for 48 h significantly induced autophagy, as
evidenced by increased diffuse green spots, the formation and
conversion of LC3-II and the increased expression of Beclin-1 in a
concentration-dependent manner (Fig.
3).
It has been demonstrated that oridonin-induced
apoptosis is attenuated by 3MA (an autophagy specific inhibitor) in
human breast cancer cells, indicating that oridonin-induced
autophagy participated in the upregulation of apoptosis (38). Another study also demonstrated that
treatment with 3MA and Baf-A1 (another autophagy specific
inhibitor) enhanced the salidroside-induced apoptosis of HT-29
colon cancer cells, indicating that salidroside-mediated autophagy
may negatively regulate apoptosis to protect cancer cells from
programmed cell death (40). In
the present study, it was demonstrated that 3MA and Baf-A1
significantly decreased the effects of AKH on cell growth
inhibition, apoptosis and autophagy induction, and the expression
of apoptosis- and autophagy-related proteins, including cleaved
PARP, pro- and cleaved caspase-3 and Beclin-1 in the Panc-28 and
A549 cells (Fig. 4). Therefore,
these results confirmed that AKH induced autophagy-related
apoptosis and the data are consistent with those of previous
studies (38,40).
Akt/mTOR/p70S6K is one of the major pathways of
autophagy and the activation of Akt/mTOR/p70S6K can inhibit
autophagy (41). mTOR plays a
central role by negatively controlling autophagy and
phosphorylation of AMPK inhibits the mTOR signaling pathway,
thereby activating autophagy (15,42).
AMPK can also regulate apoptosis and autophagy by affecting the
activation of p53, Bax, Bak and the caspase cascade (37). The present study found that AKH
significantly increased the levels of p-AMPK and decreased those of
p-Akt and its downstream target proteins, p-mTOR and p-70S6K in the
Panc-28 and A549 cells (Fig. 5).
Therefore, the AMPK and Akt/mTOR/p70S6K signaling pathways play key
roles in the AKH-induced growth inhibition and apoptosis/autophagy
induction in cancer cells.
To successfully develop an effective novel
anticancer drug clinically, it must be validated using in
vivo animal studies and clinical trials in order to confirm
whether it is highly active against tumors with tolerated toxicity
to the host. The establishment of transplanted human tumors in nude
mice is an ideal approach to carry out studies on antitumor
activity and subsequently, pharmacology in vivo (43). Therefore, the present study
established the A549 ×enograft model to investigate the antitumor
effects of AKH in vivo. The result revealed that AKH was
active against A549 human lung cancer tumor xenografts with
tolerated toxicity to the host (Fig.
6). AKH displayed significant in vivo antitumor activity
in a dose-dependent manner and high-dose AKH (400 mg/kg) exerted
more prominent antitumor effects and less toxicity compared with
cisplatin, with the tumor inhibition rates of 72.72% for AKH and
50.40% for cisplatin. According to the formula of human equivalent
dose (HED) calculation as HED (mg/kg)=Animal does (mg/kg) × (animal
Km/human Km) Eq, 400 mg/kg of AKH in mice is equal to 1,626 mg in
an individual of 50 kg (44). AKH
is a preparation of TCM and the used dosage of the ingredients of
TCM is relatively high for the treatment of various diseases
clinically. For example, the daily dose of Jinlida granule is ~27 g
for diabetes mellitus treatment (45). Hua Shi Bai Du granule is used at a
daily dose of 20 g in the treatment of coronavirus disease
(46). Therefore, the preparations
of TCM, including AKH can be applied to a dose of 20 g clinically.
However, the clinical doses of AKH and/or its active components
should be based on the results of preclinical pharmacodynamics,
pharmacokinetics, efficacy and toxicology, as well as phase I
clinical trials. Therefore, AKH has the potential to be further
developed for the treatment of patients with lung cancer
clinically. However, further studies on AKH and/or its active
fractions for the antitumor efficacy and toxicity are warranted,
with various animal models of human tumor xenografts in vivo
and validation by clinical trials. In the in vivo
experiments from the present study, AKH only significantly
suppressed the growth of A549 lung cancer xenografts but did not
shrink the original size of the tumors. In general, the combination
of an anticancer agent extracted from TCM and a chemotherapeutic
drug could enhance the antitumor efficacy and reduce the
side-effects of chemotherapy. Therefore, the clinical application
of AKH in cancer therapy is expected to be combined with
chemotherapeutic, targeted therapeutic and/or immunotherapeutic
agents.
Furthermore, the compositions of AKH were determined
by LCMS-IT-TOF analysis and nine compounds were detected from AKH
namely pinocembrin, trans,trans-1,7-diphenyl–1,3-heptadien-5-ol,
cardamomin,
(−)-2-(1,2,3,4,4a,7-haxahydro-4a,8-dimethyl-1,7-dioxo-2-naphthyl)-propionic
acid, (1E,4Z)-5-hydroxy-1-phenylhexa-1,4-dien-3-one,
trans,trans-1,7-diphenyl-5-hydroxy-4,6-hepten-3-one,
1,7-diphenyl-4,6-heptadien-3-one, katsumadain B and
(4E,6E)-5-hydroxy-1,7-diphenylhepta-4,6-dien-3-on-e, respectively
(Fig. 7 and Table II). These findings indicated that
AKH mainly contains chalcones and diarylheptanoids. Pinocembrin
exerts anti-inflammatory, anti-bacterial and anticancer effects
(47,48). Studies have shown that pinocembrin
inhibits the proliferation, migration and invasion of breast cancer
cells at non-cytotoxic concentrations by inhibiting the STAT3
signaling pathway (48).
Cardamonin is the main flavonoid derived from AKH and exerts
anti-proliferative effects against various cancer cells (49). Another study demonstrated that the
anticancer effects of cardamonin were associated with autophagy in
HCT-116 cells (23). In a previous
study by the authors, it was also shown that cardamonin induced the
apoptosis of glioblastoma stem cells by suppressing the STAT3
signaling pathway (30). However,
the study of katsumadain B is only limited to its anti-emetic
effect (50). Therefore,
pinocembrin and cardamonin may play key roles in the anticancer
activity of AKH. IT has been indicated that alpinetin is one of the
main constituents of AKH and that it is involved in its antitumor
effect (51,52). However, the present study could not
detect alpinetin using LCMS-IT-TOF, which is inconsistent with the
aforementioned studies.
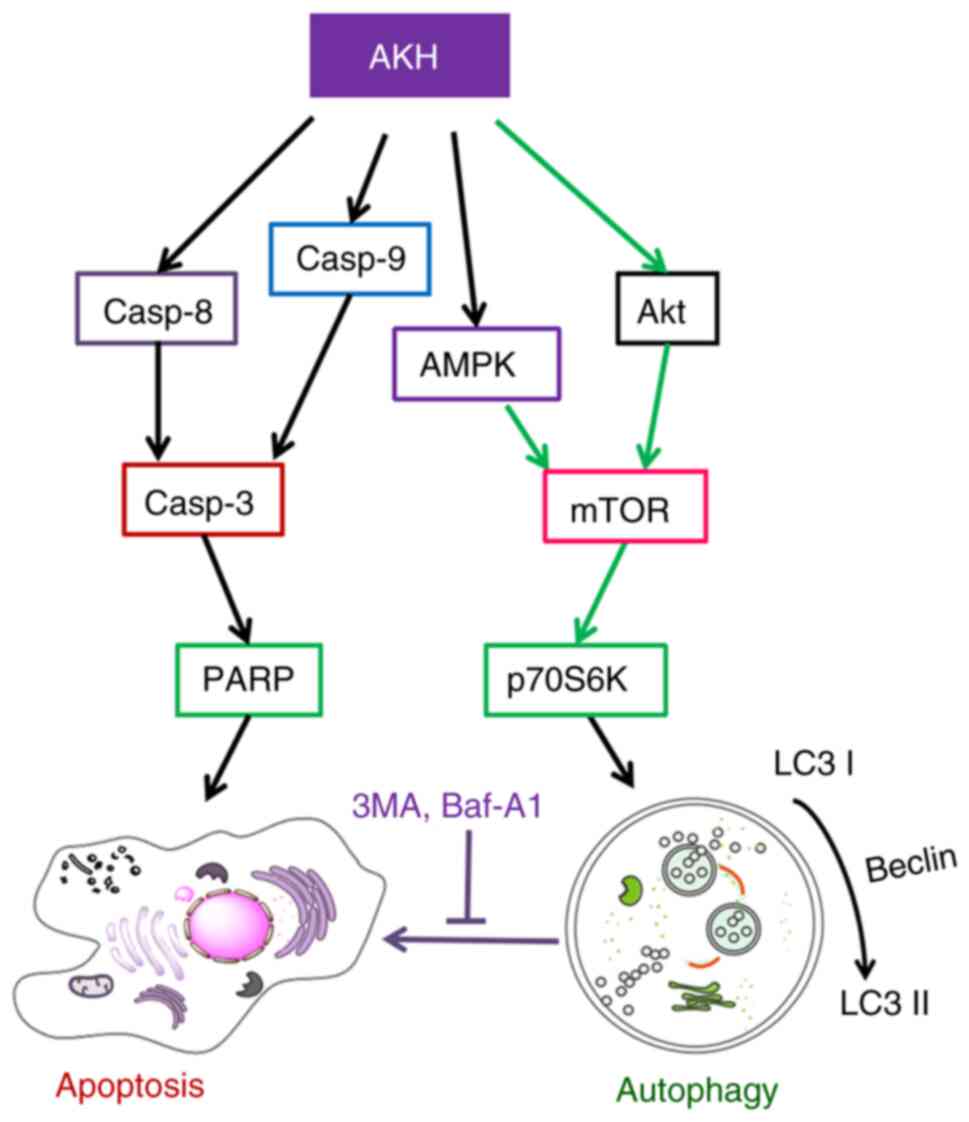
Finally, the model of the possible underlying
molecular mechanisms associated with the effects of AKH on the
apoptosis and autophagy in human cancer cells was proposed in
(Fig. 8). AKH induces
caspase-mediated apoptosis by increasing the expression of
caspase-8, caspase-9, cleaved caspase-3 and cleaved PARP. Moreover,
AKH not only promotes apoptosis, but also activates autophagy. AKH
inhibits the activation of the Akt/mTOR/p70S6K signaling pathway by
increasing the levels of p-AMPK, and decreasing those of p-Akt,
p-mTOR and p-70S6K, thereby activating autophagy.
In conclusion, the present study demonstrated that
AKH selectively inhibited the proliferation of various cancer
cells, whereas it exerted much less potent inhibitory effects on
normal liver cells. AKH significantly induced cellular apoptosis
and autophagy by regulating the AMPK and Akt/mTOR/p70S6K signaling
pathways in Panc-28 pancreatic cancer and A549 lung cancer cells
in vitro. Furthermore, AKH significantly inhibited tumor
growth with better antitumor efficacy and less toxicity compared to
that of cisplatin (a commonly used anticancer drug in clinic) in
nude mice bearing A549 tumor xenografts in vivo. In
addition, nine components were identified from AKH by LCMS-IT-TOF
assay. Therefore, the findings presented herein may provide the
scientific rationale for the understanding of the anticancer
effects of AKH and associated mechanisms of action. AKH and its
active fractions may have the potential to be developed as novel
anticancer agents clinically.
Acknowledgements
Not applicable.
Funding
The present study was supported by the International
Collaborative Project of the MOST of China (grant no.
2017YFE0195000), the Taishan Talents project of Shandong province
and the Natural Science Foundation in Shandong Province of China
(grant nos. ZR2020MH420, ZR2020MH421 and ZR2020QH360), and the
Distinguished Professor Research Startup Funding from Southwest
Medical University (grant no. 2015-RCYJ0002).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SC and XL provided the funding and designed the
study, supervised the experiments, and analyzed the data. WA, YZ,
HL and YZ performed the experiments. HZ, GZ, YL and ML analyzed the
data and prepared the figures. WA, YZ and SC wrote the manuscript.
WA and HL confirm the authenticity of all the raw data. All authors
have read and approved the final version of the manuscript.
Ethics approval and consent to
participate
All animal experiments were approved (permit no.
201802-112) by the Institutional Animal Care and Use Committee of
Southwest Medical University (Luzhou, China) and strictly followed
the guidelines for the investigation of experimental pain in
conscious animals for improving animal welfare to minimize animal
suffering (29).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Adams C, Grey N, Magrath I, Miller A and
Torode J: The world cancer declaration: Is the world catching up?
Lancet Oncol. 11:1018–1020. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Qiu X and Jia J: Research advances on TCM
anti-tumor effects and the molecular mechanisms. J Cancer Res Ther.
10 (Suppl 1):S8–S13. 2014. View Article : Google Scholar
|
|
4
|
Li Z, Feiyue Z and Gaofeng L: Traditional
Chinese medicine and lung cancer-from theory to practice. Biomed
Pharmacother. 137:1113812021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lim HS, Seo CS, Ha H, Lee H, Lee JK, Lee
MY and Shin H: Effect of Alpinia katsumadai Hayata on house
dust mite-induced atopic dermatitis in NC/Nga mice. Evid Based
Complement Alternat Med. 2012:7051672012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu H, Che X, Zheng Q, Wu A, Pan K, Shao A,
Wu Q, Zhang J and Hong Y: Caspases: A molecular switch node in the
crosstalk between autophagy and apoptosis. Int J Biol Sci.
10:1072–1083. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Z and Klionsky DJ: An overview of the
molecular mechanism of autophagy. Curr Top Microbials Immunol.
335:1–32. 2009.PubMed/NCBI
|
|
10
|
Panda PK, Mukhopadhyay S, Das DN, Sinha N,
Naik PP and Bhutia SK: Mechanism of autophagic regulation in
carcinogenesis and cancer therapeutics. Semin Cell Dev Biol.
39:43–55. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
White E: Deconvoluting the
context-dependent role for autophagy in cancer. Nat Rev Cancer.
12:401–410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ávalos Y, Canales J, Bravo-Sagua R,
Criollo A, Lavandero S and Quest AF: Tumor suppression and
promotion by autophagy. Biomed Res Int. 2014:6039802014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meijer AJ and Codogno P: Regulation and
role of autophagy in mammalian cells. Int J Biochem Cell Biol.
36:2445–2462. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Codogno P and Meijer AJ: Autophagy and
signaling: Their role in cell survival and cell death. Cell Death
Differ. 12 (Suppl 2):S1509–S1518. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsui Y, Takagi H, Qu X, Abdellatif M,
Sakoda H, Asano T, Levine B and Sadoshima J: Distinct roles of
autophagy in the heart during ischemia and reperfusion: Roles of
AMP-activated protein kinase and beclin 1 in mediating autophagy.
Circ Res. 100:914–922. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scott RC, Juhász G and Neufeld TP: Direct
induction of autophagy by Atg1 inhibits cell growth and induces
apoptotic cell death. Curr Biol. 17:1–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bousman CA, Chana G, Glatt SJ, Chandler
SD, Lucero GR, Tatro E, May T, Lohr JB, Kremen WS, Tsuang MT and
Everall IP: Preliminary evidence of ubiquitin proteasome system
dysregulation in schizophrenia and bipolar disorder: Convergent
pathway analysis findings from two independent samples. Am J Med
Genet B Neuropsychiatr Genet. 153B:494–502. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang CY, Bai XY and Wang CH: Traditional
Chinese medicine: A treasured natural resource of anticancer drug
research and development. Am J Chin Med. 42:543–559. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
An W, Lai H, Zhang Y, Liu M, Lin X and Cao
S: Apoptotic pathway as the therapeutic target for anticancer
traditional Chinese medicines. Front Pharmacol. 10:7582019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chinese Pharmacopoeia Commission: Chinese
Pharmacopoeia, 2020. Beijing, China: Chinese Medicine Science and
Technology Press. Part 1; pp. pp2492020
|
|
21
|
Lee MY, Lee NH, Seo CS, Lee JA, Jung D,
Kim JH and Shin HK: Alpinia katsumadai seed extract
attenuate oxidative stress and asthmatic activity in a mouse model
of allergic asthma. Food Chem Toxicol. 48:1746–1752. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Du J, Tang B, Wang J, Sui H, Jin X, Wang L
and Wang Z: Antiproliferative effect of alpinetin in BxPC-3
pancreatic cancer cells. Int J Mol Med. 29:607–612. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim YJ, Kang KS, Choi KC and Ko H:
Cardamonin induces autophagy and an antiproliferative effect
through JNK activation in human colorectal carcinoma HCT116 cells.
Bioorg Med Chem Lett. 25:2559–2564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang XB, Yang CS, Luo JG, Zhang C, Luo J,
Yang MH and Kong LY: Experimental and theoretical calculation
studies on the structure elucidation and absolute configuration of
calyxins from Alpinia katsumadai. Fitoterapia. 119:121–129.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu M, Zhao G, Zhang D, An W, Lai H, Li X,
Cao S and Lin X: Active fraction of clove induces apoptosis via
PI3K/Akt/mTOR-mediated autophagy in human colorectal cancer HCT-116
cells. Int J Oncol. 53:1363–1373. 2018.PubMed/NCBI
|
|
26
|
Cao S, McGuire JJ and Rustum YM: Antitumor
activity of ZD1694 (tomudex) against human head and neck cancer in
nude mouse models: Role of dosing schedule and plasma thymidine.
Clin Cancer Res. 5:1925–1934. 1999.PubMed/NCBI
|
|
27
|
Cao S, Durrani FA, Tóth K and Rustum YM:
Se-methylselenocysteine offers selective protection against
toxicity and potentiates the antitumour activity of anticancer
drugs in preclinical animal models. Br J Cancer. 110:1733–1743.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kanzawa T, Zhang L, Xiao L, Germano IM,
Kondo Y and Kondo S: Arsenic trioxide induces autophagic cell death
in malignant glioma cells by upregulation of mitochondrial cell
death protein BNIP3. Oncogene. 24:980–991. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zimmermann M: Ethical guidelines for
investigations of experimental pain in conscious animals. Pain.
16:109–110. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu N, Liu J, Zhao X, Yan Z, Jiang B, Wang
L, Cao S, Shi D and Lin X: Cardamonin induces apoptosis by
suppressing STAT3 signaling pathway in glioblastoma stem cells.
Tumour Biol. 36:9667–9676. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han C, Xing G, Zhang M, Zhong M, Han Z, He
C and Liu X: Wogonoside inhibits cell growth and induces
mitochondrial-mediated autophagy-related apoptosis in human colon
cancer cells through the PI3K/AKT/mTOR/p70S6K signaling pathway.
Oncol Lett. 15:4463–4470. 2018.PubMed/NCBI
|
|
32
|
Shinojima N, Yokoyama T, Kondo Y and Kondo
S: Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in
curcumin-induced autophagy. Autophagy. 3:635–637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dasgupta B and Chhipa RR: Evolving lessons
on the complex role of AMPK in normal physiology and cancer. Trends
Pharmacol Sci. 37:192–206. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moore J, Megaly M, MacNeil AJ, Klentrou P
and Tsiani E: Rosemary extract reduces Akt/mTOR/p70S6K activation
and inhibits proliferation and survival of A549 human lung cancer
cells. Biomed Pharmacother. 83:725–732. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moutinho-Ribeiro P, Macedo G and Melo SA:
Pancreatic cancer diagnosis and management: Has the time come to
prick the bubble? Front Endocrinol (Lausanne). 9:7792019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chaabane W, User SD, El-Gazzah M, Jaksik
R, Sajjadi E, Rzeszowska-Wolny J and Los MJ: Autophagy, apoptosis,
mitoptosis and necrosis: Interdependence between those pathways and
effects on cancer. Arch Immunol Ther Exp (Warsz). 61:43–58. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Y, Wang Y, Wang S, Gao Y, Zhang X and
Lu C: Oridonin phosphate-induced autophagy effectively enhances
cell apoptosis of human breast cancer cells. Med Oncol. 32:3652015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou ZW, Li XX, He ZX, Pan ST, Yang Y,
Zhang X, Chow K, Yang T, Qiu JX, Zhou Q, et al: Induction of
apoptosis and autophagy via sirtuin1- and PI3K/Akt/mTOR-mediated
pathways by plumbagin in human prostate cancer cells. Drug Des
Devel Ther. 9:1511–1554. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fan XJ, Wang Y, Wang L and Zhu M:
Salidroside induces apoptosis and autophagy in human colorectal
cancer cells through inhibition of PI3K/Akt/mTOR pathway. Oncol
Rep. 36:3559–3567. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang HW, Hu JJ, Fu RQ, Liu X, Zhang YH,
Li J, Liu L, Li YN, Deng Q, Luo QS, et al: Flavonoids inhibit cell
proliferation and induce apoptosis and autophagy through
downregulation of PI3Kγ mediated PI3K/AKT/mTOR/p70S6K/ULK signaling
pathway in human breast cancer cells. Sci Rep. 8:112552018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Inoki K, Kim J and Guan KL: AMPK and mTOR
in cellular energy homeostasis and drug targets. Annu Rev Pharmacol
Toxicol. 52:381–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu CF, Wu CY, Chiou RY, Yang WC, Lin CF,
Wang CM, Hou PH, Lin TC, Kuo CY and Chang GR: The anti-cancer
effects of a zotarolimus and 5-fluorouracil combination treatment
on A549 cell-derived tumors in Balb/c nude mice. Int J Mol Sci.
22:45622021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nair AB and Jacob S: A simple practice
guide for dose conversion between animals and human. J Basic Clin
Pharm. 7:27–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shi YL, Liu WJ, Zhang XF, Su WJ, Chen NN,
Lu SH, Wang LY, Shi XL, Li ZB and Yang SY: Effect of Chinese herbal
medicine Jinlida granule in treatment of patients with impaired
glucose tolerance. Chin Med J (Engl). 129:2281–2286. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu J, Yang W, Liu Y, Lu C, Ruan L, Zhao
C, Huo R, Shen X, Miao Q, Lv W, et al: Combination of Hua Shi Bai
Du granule (Q-14) and standard care in the treatment of patients
with coronavirus disease 2019 (COVID-19): A single-center,
open-label, randomized controlled trial. Phytomedicine.
91:1536712021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rasul A, Millimouno FM, Ali Eltayb W, Ali
M, Li J and Li X: Pinocembrin: A novel natural compound with
versatile pharmacological and biological activities. Biomed Res
Int. 2013:3798502013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhu X, Li R, Wang C, Zhou S, Fan Y, Ma S,
Gao D, Gai N and Yang J: Pinocembrin inhibits the proliferation and
metastasis of breast cancer via suppression of the PI3K/AKT
signaling pathway. Front Oncol. 11:6611842021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gonçalves LM, Valente IM and Rodrigues JA:
An overview on cardamonin. J Med Food. 17:633–640. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yang Y, Kinoshita K, Koyama K, Takahashi
K, Tai T, Nunoura Y and Watanabe K: Two novel anti-emetic
principles of Alpinia katsumadai. J Nat Prod. 62:1672–1674.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Huo M, Chen N, Chi G, Yuan X, Guan S, Li
H, Zhong W, Guo W, Soromou LW, Gao R, et al: Traditional medicine
alpinetin inhibits the inflammatory response in Raw 264.7 cells and
mouse models. Int Immunopharmacol. 12:241–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao X, Guo X, Shen J and Hua D: Alpinetin
inhibits proliferation and migration of ovarian cancer cells via
suppression of STAT3 signaling. Mol Med Rep. 18:4030–4036.
2018.PubMed/NCBI
|