Introduction
Breast cancer (BC) is the most common cancer type
affecting women worldwide (1).
Despite the benefits arising from expanding access to high-quality
prevention methods and early detection, as well as the rapid
development of treatment modalities, such as surgical resection,
endocrine therapy, chemotherapy and immunotherapy, the mortality
rate associated with BC remains relatively high and a further
decrease is warranted (2,3). Although certain molecular markers
have been extensively characterized, including immunohistochemical
markers (i.e., estrogen receptor, progesterone receptor, human
epidermal growth factor receptor 2 and Ki-67), genomic markers
[i.e., breast invasive carcinoma (BRCA)1, BRCA2 and
phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit α]
and immunomarkers (i.e., programmed death-ligand 1), new biomarkers
and new biomarker combinations still need to be developed, due to
the high tumor heterogeneity of BC.
Cancer stem cells (CSCs) retain the properties of
self-renewal, differentiation and drug-resistance, all of which are
considered to lead to a poor therapeutic response in BC, as well as
tumor recurrence and metastasis (4–6).
CSCs have been identified in various types of cancer, including BC.
The most common markers of BC stem cells (BCSCs) are
CD44+CD24− and aldehyde dehydrogenase
(ALDH)+ (7,8). Although a number of studies have
identified that targeting CSCs may be an effective approach to
anticancer treatment, more CSC-related markers and mechanisms need
to be elucidated.
Human cell division cycle-associated 5 (CDCA5; also
known as sororin or p35), which is located at 11q13.1, is a
critical regulator of sister chromatid in mitosis, stabilizing the
association between cohesion complex and chromatin, and is involved
in mitotic cell cycle and double-strand break repair (9,10).
In addition, CDCA5 is necessary for the stable cohesion of
chromatids during the S and G2/M phases and is then degraded
through anaphase-promoting complex-dependent ubiquitination during
the G0/G1 phase (11–13). Diseases associated with CDCA5
include Cornelia de Lange syndrome (14) and Robert-Sc phocomelia syndrome
(15). A significantly increased
CDCA5 expression has been reported in various human tumor tissues,
including lung cancer (16),
urothelial carcinoma (17), oral
squamous cell carcinoma (18),
gastric cancer (19,20) and hepatocellular carcinoma
(21) tissues. Hence, CDCA5 may be
a poor prognostic biomarker for patients with these types of
cancer. However, the clinical significance and biological function
of CDCA5 in BC are not yet fully understood.
In the present study, a link was identified between
aberrant CDCA5 expression and a poor prognosis of patients with BC
by analyzing The Cancer Genome Atlas (TCGA). Furthermore, it was
found that BC cells and BCSCs exhibited a higher CDCA5 expression
than normal breast and adherent cells, respectively. Finally, CDCA5
knockdown in BC cells inhibited cell proliferation, migration and
CSC activity. These findings thus suggest that CDCA5 may be a novel
prognostic biomarker for BC.
Materials and methods
Bioinformatics analysis
UALCAN (22) is a
comprehensive, user-friendly and interactive web resource for
analyzing OMICS data from the TCGA, MET500 and CPTAC databases.
CDCA5 mRNA expression was investigated in various tumor samples
(including BC samples) through a pan-cancer view of TCGA cancers
(using tumor and normal samples) and the expression of CDCA5 was
investigated, particularly in BRCA, based on sample types,
individual cancer stages and TP53 mutation status.
The Kaplan-Meier Plotter (23) is an online tool that was used
herein to perform a meta-analysis discovery and validation of
survival biomarkers, including >5,400 genes from the Gene
Expression Omnibus (GEO), European Genome-phenome Archive (EGA) and
TCGA databases, covering breast (n=7,830), ovarian (n=2,190), lung
(n=3,452) and gastric (n=1,440) cancer. CDCA5 (Affymetrix ID:
224753_at) was analyzed by selecting all probe sets per gene in
probe set options and a median split in the patient options. The
overall survival (OS), disease-free survival (DFS) and distant
metastasis-free survival (DMFS) of patients with BC were analyzed
by the use of the log-rank test.
LinkedOmics (24)
is a publicly available portal that includes three analytical
modules: LinkFinder, LinkInterpreter and LinkCompare. The
LinkInterpreter module performs enrichment analysis was based on
Gene Ontology (GO), biological pathways and network modules, among
other functional categories. CDCA5 was analyzed using the Pearson's
correlation coefficient with RNAseq data on the TCGA_BRCA sample
cohort for GO and Kyoto Encyclopedia of Genes and Genomes (KEGG)
analysis.
StarBase (25) is
an open-source platform that systematically identifies the RNA-RNA
and protein-RNA interaction networks from 108 CLIP-Seq (PAR-CLIP,
HITS-CLIP, iCLIP and CLASH) datasets generated by 37 independent
studies. The correlation between CDCA5 and stemness transcription
factors SRY-box transcription factor 2 (SOX2), POU class 5 homeobox
1 (POU5F1, also known as OCT4) and c-MYC in BRCA was analyzed.
Cells and cell culture
The human normal mammary epithelial cell line,
MCF10A (cat no. CL-0525), was purchased from Wuhan Procell Company
and cultured in MCF10A special medium (Procell Co., Ltd.). The
human BC cell lines, SKBR3 (cat no. 1101HUM-PUMC000085), MCF7 (cat
no: 1101HUM-PUMC000013), MDA-MB-231 (cat no. 1101HUM-PUMC000014)
and Hs578T (cat no. 1101HUM-PUMC000670) and MDA-MB-468 (cat no.
1101HUM-PUMC000249), were purchased from the National
Infrastructure of Cell line Resource (NICR). The SKBR3, MCF7,
MDA-MB-231 and Hs578T cells were cultured in DMEM (Meilun Biotech
Co., Ltd.) and the MDA-MB-468 cells were cultured in RPMI-1640
(Meilun Biotech Co., Ltd.) medium supplemented with 10% fetal
bovine serum (HyClone; Cytiva), 100 U/ml penicillin and 100 µg/ml
streptomycin (Gibco; Thermo Fisher Scientific, Inc.) in a 5%
CO2 incubator at 37°C.
Lentivivrus infection
shRNA vectors for sh-CDCA5 and a control vector were
purchased from Tsingke Biotechnology Co. Ltd. The shRNA sequences
were as follows: sh-CDCA5 forward,
5′-CCGGGGACGCCAGAGACTTGGAAATCTCGAGATTTCCAAGTCTCTGGCGTCCTTTTTG-3′
and reverse,
5′-AATTCAAAAAGGACGCCAGAGACTTGGAAATCTCGAGATTTCCAAGTCTCTGGCGTCC-3′.
The second-generation three-plasmid lentivirus system was used.
When the 293T (Procell Co., Ltd.) cells were in the logarithmic
growth phase, a total of 10 µg of the target plasmid and the helper
plasmid in a ratio of 5:3:2 were incubated with PEI transfection
reagent at 37°C for 30 min. After 72 h, the cell supernatant was
collected, cell debris was removed, the virus was collected by
centrifugation with PEG8000, incubated with target cells MCF7 and
SKBR3 for 4 h, the medium was changed, and puromycin was started
after 48 h. Screening, continuous drug screening for 2 weeks, to
obtain stable cell lines.
The SKBR3 and MCF7 BC cells were seeded in 6-well
plates (5×104 cells/well) at 60% confluency and then
infected with the sh-CDCA5 lentivirus. After 48 h, the stably
transfected cells were established using selection with 2 µg/ml
puromycin for 2 weeks.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from the BC cell lines using
the Ultrapure RNA kit (CWBio), according to the manufacturer's
instructions. Reverse transcription was conducted using
HiScript® II Q RT SuperMix for qPCR (Vazyme Biotech Co.,
Ltd.) at 50°C for 15 min and 85°C for 5 sec. The mRNA levels were
determined using Hieff® qPCR SYBR®-Green
Master Mix (Yeasen Biotechnology Co., Ltd.) on a Bio-Rad CFX
Maestro (Bio-Rad Laboratories, Inc.) with the following PCR cycling
conditions: 95°C 5 min; 95°C 15 sec; 55°C 15 sec; and 72°C 30 sec
for 30 cycles. GAPDH served as the quantitative control to
normalize the mRNA expression levels of the target genes. The
primer sequences for the detected genes are listed in the Table I.
 | Table I.Gene specific primers used for
reverse transcription-quantitative PCR. |
Table I.
Gene specific primers used for
reverse transcription-quantitative PCR.
| Gene | Forward primer (5′
to 3′) | Reverse primer (5′
to 3′) |
|---|
| GAPDH |
TTAAAAGCAGCCCTGGTGAC |
CTCTGCTCCTCCTGTTCGAC |
| SOX2 |
GCCGAGTGGAAACTTTTGTCG |
GGCAGCGTGTACTTATCCTTCT |
| C-MYC |
GGCTCCTGGCAAAAGGTCA |
CTGCGTAGTTGTGCTGATGT |
| OCT4 |
TTTTGGTACCCCAGGCTATGGGAG |
GTTTGAATGCATGGGAGAGCCCAG |
| CDCA5 |
GAGGTCCCAGCGGAAATCAG |
TCTTTAAGACGATGGGCTTTCTG |
Western blot analysis
Protein samples were prepared using RIPA lysis
buffer (Meilun Biotech Co., Ltd.) containing protease inhibitors on
ice for 15 min, and were then centrifuged at 12,000 × g for 15 min
at 4°C. Protein concentrations were measured using the BCA Protein
Assay kit (Beyotime Institute of Biotechnology). Equal amounts of
protein were separated by 10% SDS-PAGE, and transferred to PVDF
membranes (MilliporeSigma). They were then incubated with
TRIS-buffered saline (TBST) containing 5% skim milk at 37°C for 2
h, followed by incubation with primary antibodies targeting CDCA5
rabbit mAb (dilution, 1:2,000; cat no. A4044; ABclonal Science,
Inc.), OCT4 rabbit mAb (dilution, 1:2,000; cat no. T55781; Abmart
Inc.), c-MYC rabbit mAb (dilution, 1:2,000; cat no. T55150; Abmart
Inc.), SOX2 rabbit mAb (dilution, 1:2,000; cat no. T55268; Abmart
Inc.) and GAPDH rabbit mAb (dilution, 1:2,000; cat no. A19056;
ABclonal Science, Inc.) overnight at 4°C. Following three 5-min
washes in TBST, membranes were incubated with HRP goat anti-rabbit
secondary antibodies (dilution, 1:2,000; cat no. AS014; ABclonal
Science, Inc.) at room temperature for 1.5 h. Chemiluminescent
signals were detected using enhanced chemiluminescence detection
reagents (Meilun Biotech Co., Ltd.).
Cell proliferation assay
The MCF7 and SKBR3 cells were seeded in 96-well
plates at 2,000 cells/well, each well containing 100 µl culture
medium. A Cell Counting Kit-8 assay (Beyotime Institute of
Biotechnology) was performed at 24, 48, 72 and 96 h, according to
the manufacturer's instructions. The resulting solution was
quantified using a microplate reader (FC; Thermo Fisher Scientific,
Inc.). The light absorption value at 450 nm was detected, and the
cell growth curve was plotted with time as the abscissa and
absorbance as the ordinate.
Colony formation assay
Following selection with puromycin, the stably
transfected BC cells subjected to CDCA5 knockdown and the control
cells were seed in 6-well plates (1×103 cells/well) and
kept in culture for 14 days to yield cell colonies. Following
fixation with 4% paraformaldehyde for 15 min, the colonies were
stained at room temperature with 0.1% crystal violet solution
(Beyotime Institute of Biotechnology) for 10 min. Colonies were
photographed and counted.
Cell migration assay
BC cell migration was examined using a Transwell
assay with 8-µm well chambers (Corning, Inc.). The treated BC cells
were harvested and resuspended in medium to a final concentration
of 1×104 cells/ml. Subsequently, 0.2 ml BC cell
suspension was added to the upper chamber, and 0.5 ml medium
containing 20% FBS was added to the lower chamber. Following 24 h
of incubation at 37°C, the migrated BC cells were stained with 0.1%
crystal violet at room temperature for 10 min (Beyotime Institute
of Biotechnology) and quantified under a microscope (Olympus/IX83,
Japan).
Mammosphere formation assay
The BCSCs were cultured as previously described
(26–28). In brief, the CSCs were enriched in
ultra-low adsorption 6-well plates (Corning, Inc.) with SKBR3 and
MCF7 cells using serum-free DMEM/F12 medium supplemented with
epidermal growth factor (20 ng/ml, T&L Biological Technology),
basic fibroblast growth factor (10 ng/ml, T&L Biological
Technology) and 2% B27 (Gibco; Thermo Fisher Scientific, Inc.) for
14 days. The numbers and volume of CSCs were calculated.
Xenograft model in vivo
A total of 12 3-week-old female nude mice were
purchased from Beijing Vital River Laboratory Animal Technology
Co., Ltd. for xenotransplantation experiments. The mice were housed
in plastic cages (three mice/cage) under controlled conditions of
light (12-h light/dark cycle), temperature (23±2°C) and humidity
(55%), and were provided with free access to food and water. These
stable cells were trypsinized and suspended in PBS. A total volume
of 0.12 ml PBS containing 1×106 MCF7 cells was
subcutaneously injected into the right flanks of the mice. The 12
mice were divided into two groups (6 mice per group) as follows:
One group was injected with the MCF7-shNC cell line, and the other
group was injected with the MCF7-shCDCA5 cell line. All mice that
reached the study endpoint were euthanized by cervical dislocation
under 2–3% isoflurane anesthesia. The humane endpoints were deemed
as the time when the xenograft tumor diameter was >20 mm, the
xenograft tumor reached >20% of the animal body weight, a body
weight loss of >20% occurred due to tumor growth, and signs of
immobility, inability to eat, ulceration, infection or necrosis
were observed. Death was verified by the observation of pupil
dilation, as well as the cessation of breath and heartbeat. The
animal research protocol was reviewed and approved by the Animal
Protection Committee of Wuhan University of Science and Technology
(Wuhan, China).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5.0 (GraphPad, Inc.). The results are presented as the mean ±
SD from at least three independent experiments. Single comparisons
between two groups were performed using the unpaired Student's
t-test. Comparisons between three or more groups were determined
using ANOVA followed by Tukey's post hoc test. Kaplan-Meier
survival analysis was used to assess cumulative survival
probability by the use of the log-rank test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of CDCA5 in different types
of cancer
The mRNA expression levels of CDCA5 were examined in
different types of cancer from the UALCAN database (Fig. 1A). The results revealed that, in
the majority of cancer types, the expression of CDCA5 was
significantly increased, including bladder urothelial carcinoma
(BLCA), breast invasive carcinoma (BRCA), cervical squamous cell
carcinoma and endocervical adenocarcinoma (CESC),
cholangiocarcinoma (CHOL), esophageal carcinoma (ESCA), liver
hepatocellular carcinoma (LIHC), lung squamous cell carcinoma
(LUSC), sarcoma (SARC) and uterine corpus endometrial carcinoma
(UCEC). There were only a few cancer types where CDCA5 was not
clearly increased, such as thymoma (THYM; normal median value of
4.91 vs. tumor median value of 4.969) and skin cutaneous melanoma
(SKCM) with the same expression level of CDCA5. TCGA pan-cancer
abbreviations and noun comparisons are presented in Table SI.
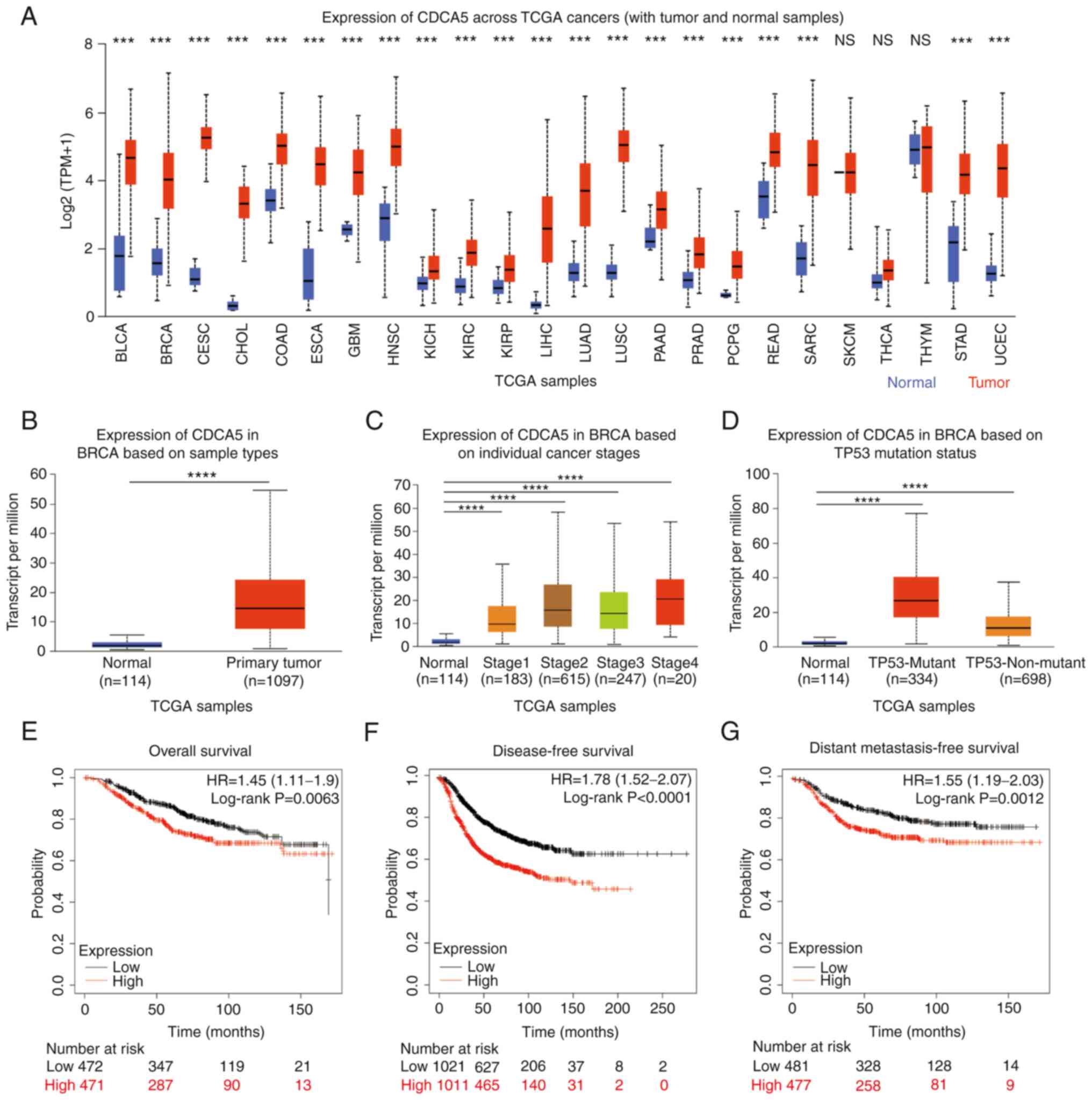 | Figure 1.High CDCA5 expression across TCGA
cancers and invasive breast cancer. (A) CDCA5 mRNA expression in
various types of cancer compared with normal tissues from the
UALCAN database. ***P<0.001. Expression of CDCA5 in BRCA based
on (B) sample types, (C) individual cancer stages and (D) TP53
mutation status from TCGA samples. (E) Overall, (F) disease-free
and (G) distant metastasis-free survival curves from the
Kaplan-Meier Plotter, and the evaluation of the impact of the low
(black line) and high (red line) CDCA5 expression. ****P<0.0001.
CDCA5, cell division cycle-associated 5; TCGA, The Cancer Genome
Atlas; BLCA, bladder urothelial carcinoma; BRCA, breast invasive
carcinoma; CESC, cervical squamous cell carcinoma and endocervical
adenocarcinoma; CHOL, cholangiocarcinoma; COAD, Colon
adenocarcinoma; ESCA, Esophageal carcinoma; GBM, glioblastoma;
HNSC, head and neck squamous cell carcinoma; KICH, kidney
chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP, kidney
renal papillary cell carcinoma; LIHC, liver hepatocellular
carcinoma; LUAD, lung adenocarcinoma; LUSC, lung squamous cell
carcinoma; PAAD, pancreatic adenocarcinoma; PRAD, prostate
adenocarcinoma; PCPG, pheochromocytoma and paraganglioma; READ,
rectal adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous
melanoma; THCA, thyroid carcinoma; STAD, stomach adenocarcinoma;
UCEC, uterine corpus endometrial carcinoma. |
Expression of CDCA5 in invasive
BC
Similarly, the expression of CDCA5 in BC and normal
tissues was analyzed using the UALCAN database. CDCA5 expression
was high in 1,097 BC primary tumor tissues compared with 114 normal
tissues (P<0.0001; Fig. 1B).
The expression of CDCA5 was associated with various clinical
features, such as individual cancer stages (normal vs. stage 1,
P<0.0001; stage 2, P<0.0001; stage 3, P<0.0001; and stage
4, P<0.0001; Fig. 1C), TP53
mutation status (normal vs. TP53 mutant, P<0.0001; and
TP53-non-mutant, P<0.0001, Fig.
1D).
Prognostic value of CDCA5 in patients
with BC
Based on the upregulated expression of CDCA5 in BC,
its prognostic value in patients with BC was analyzed using the
Kaplan-Meier Plotter. Compared with the low CDCA5 expression group,
the high CDCA5 expression group exhibited a worse OS (P=0.0063,
HR=1.45; Fig. 1E), DFS
(P<0.0001, HR=1.78; Fig. 1F)
and DMFS (P=0.0012, HR=1.55; Fig.
1G).
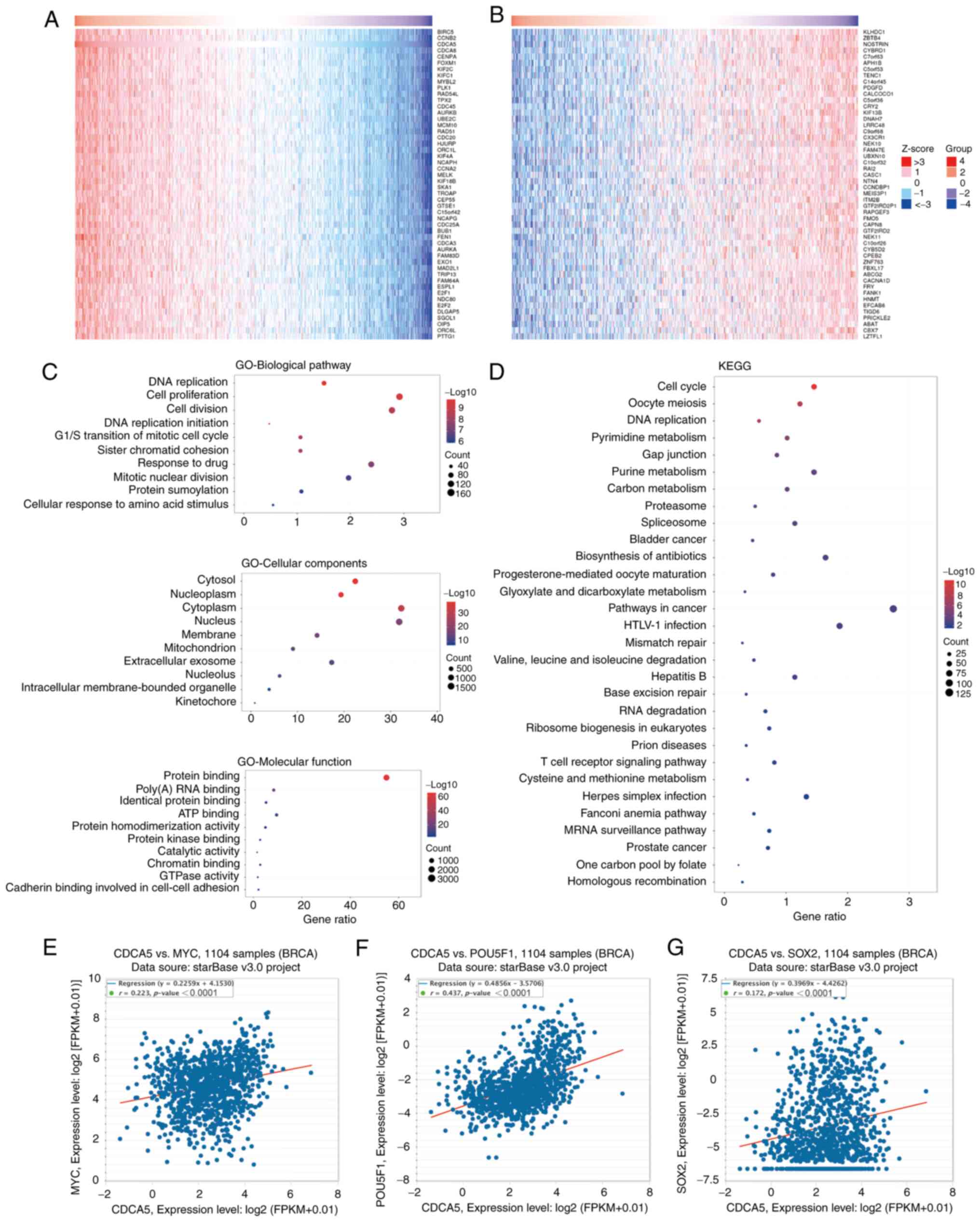
CDCA5 co-expressed genes and
functional enrichment analysis
Using the LinkedOmics database, the genes
co-expressed with CDCA5 were identified, and the top 50 positively
(Fig. 2A) and negatively (Fig. 2B) associated genes are presented in
the heat maps (Fig. 2A and B).
Further information about the top 50 positively and negatively
associated genes is presented in Tables SII and SIII. In addition, GO (Fig. 2C) and KEGG (Fig. 2D) functional enrichment analysis
was performed on the co-expressed genes. GO biological pathway
enrichment analysis (Table SIV)
indicated that these genes were involved in a variety of processes,
including DNA replication (P<0.0001), cell proliferation
(P<0.0001) and the G1/S transition of mitotic cell cycle
(P<0.0001). GO cellular component (Table SV) indicated various cellular
structures, including intracellular membrane-bounded organelles
(P<0.0001), extracellular exosomes (P<0.0001) and
mitochondria (P<0.0001). On the other hand, GO molecular
function (Table SVI) indicated
that these genes were involved in protein binding (P<0.0001),
poly(A) RNA binding (P<0.0001) and ATP binding (P<0.0001).
Moreover, KEGG functional enrichment analysis (Table SVII) revealed that these genes
were closely linked to pathways, such as T-cell receptor signaling,
Fanconi anemia and MRNA surveillance pathways. In addition, CDCA5
exhibited a strong positive correlation with the stemness
transcription factors, c-MYC (Fig.
2E), OCT4/POU5F1 (Fig. 2F) and
SOX2 (Fig. 2G).
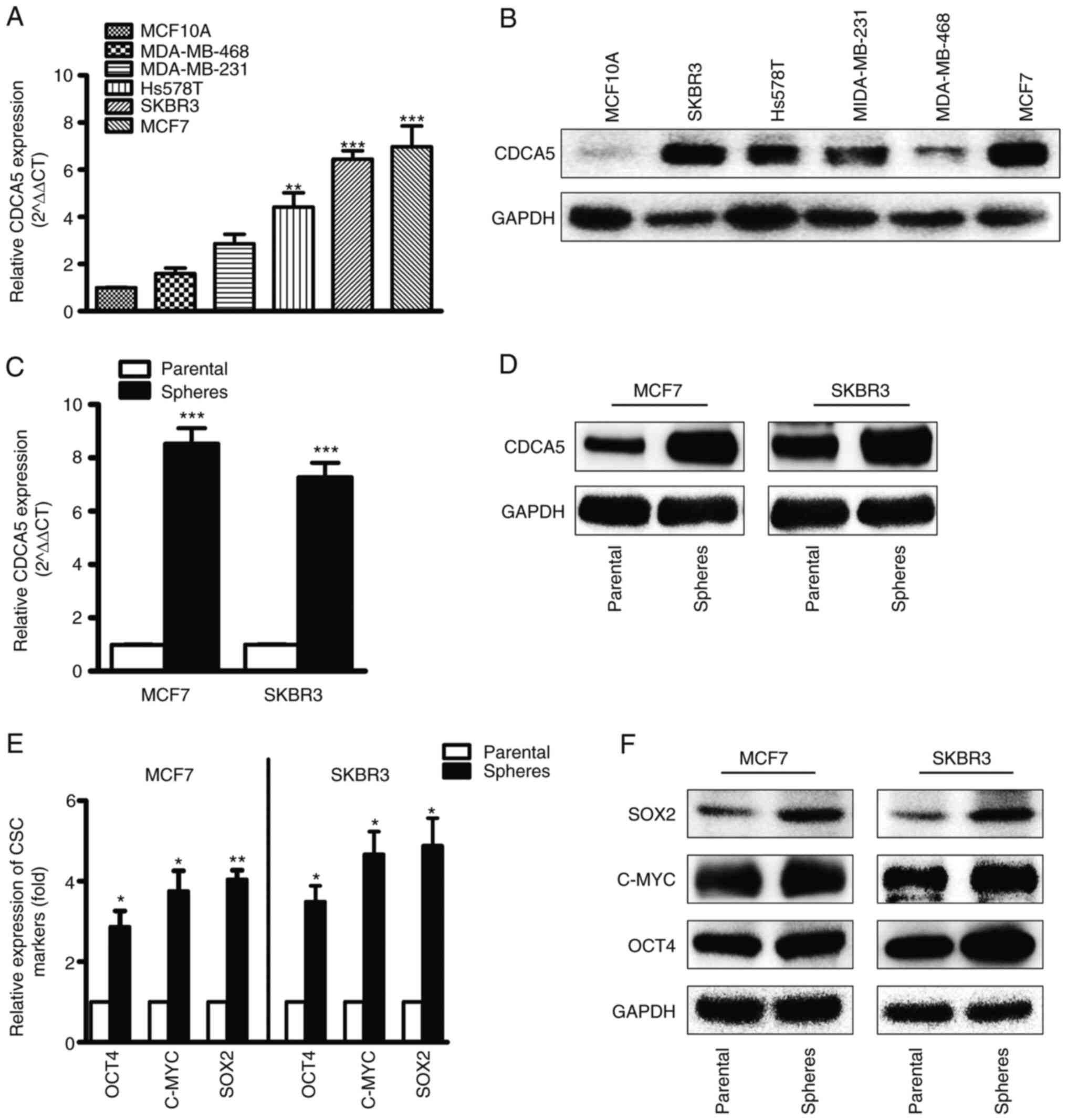
mRNA and protein expression of CDCA5
in BC cells and CSCs
CDCA5 expression was high in the MDA-MB-231,
MDA-MB-468, Hs578T, MCF7 and SKBR3 BC cell lines, and low in the
MCF10A normal breast cell lines. The mRNA (Fig. 3A) and protein (Fig. 3B) levels of CDCA5 were determined
using RT-qPCR and western blot analysis. By comparison, the SKBR3
and MCF7 cell lines exhibited higher CDCA5 expression levels
compared to the other cell lines; thus, these two cell lines were
selected for CDCA5 knockdown experiments. Furthermore, CDCA5
expression was significantly upregulated in mammospheres compared
with CDCA5 adherent cells (Fig. 3C and
D). To explore whether or not the mammospheres were CSCs, the
levels of CSC markers, including SOX2, c-MYC and OCT4 were
determined (Fig. 3E and F). The
increased levels of the CSC markers indicated that the mammospheres
contained a large number of CSCs.
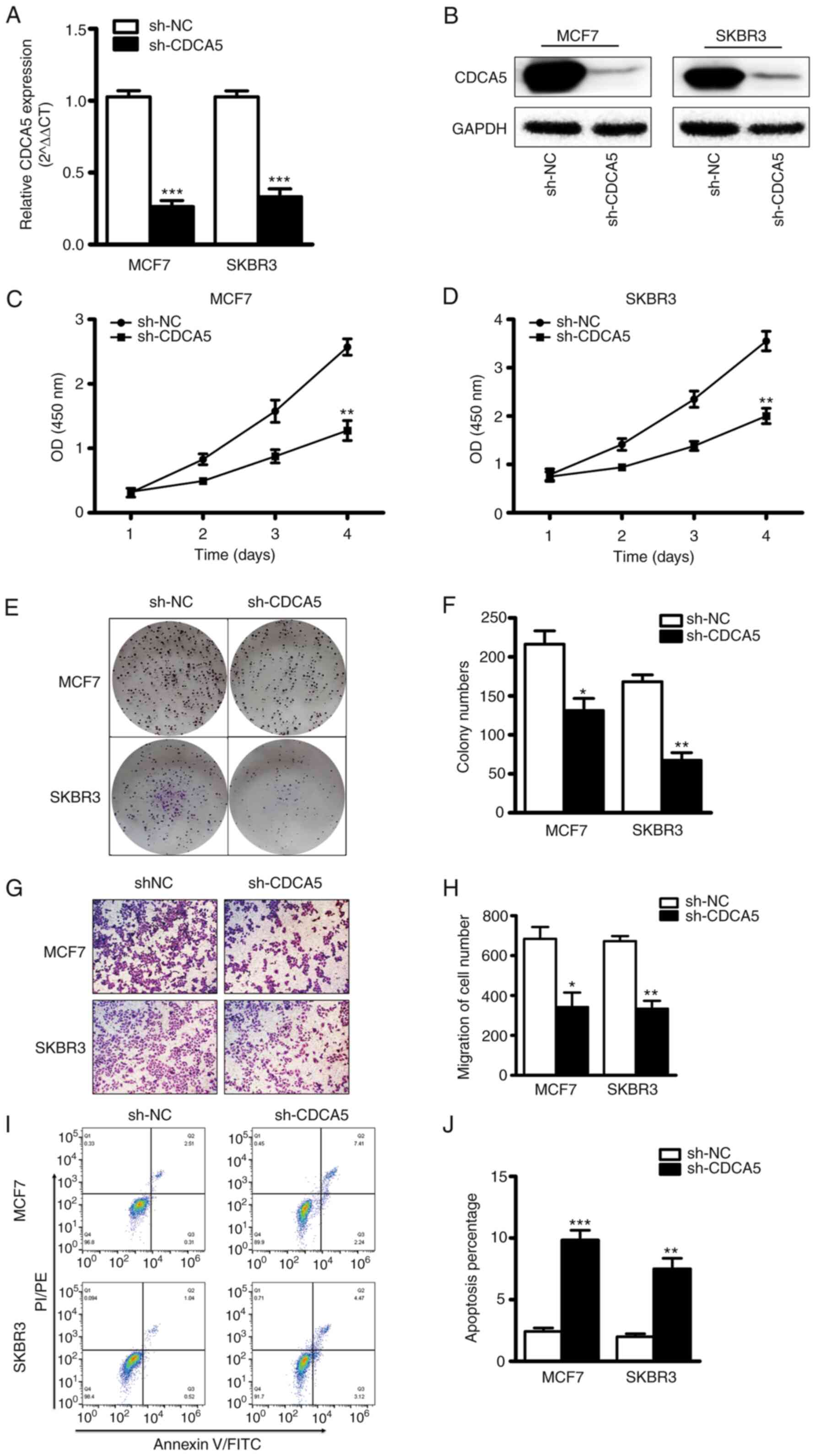
CDCA5 knockdown inhibits BC cell
proliferation and migration
The effect of CDCA5 on BC cell function was explored
in vitro. The MCF7 and SKBR3 cells were subjected CDCA5
knockdown and thus two stable cell lines with CDCA5 knockdown were
established (Fig. 4A and B). The
results of CCK-8 assay revealed that the proliferative capacity of
the MCF7 and SKBR3 cells was significantly decreased following
CDCA5 knockdown (Fig. 4C and D).
Furthermore, colony formation assays revealed that the clone
formation ability of the MCF7 and SKBR3 cells was significantly
reduced following CDCA5 knockdown (Fig. 4E). The statistical analysis results
of the number of colonies revealed a significant difference between
the two groups (Fig. 4F). The
experimental results also demonstrated that when CDCA5 was knocked
down in the SKBR3 or MCF7 BC cells, their migratory ability was
significantly decreased (Fig. 4G).
The statistical analysis results of the number of colonies revealed
a significant difference between the two groups (Fig. 4H). In addition, as shown in
Fig. 4I and J, CDCA5 knockdown
significantly promoted the apoptosis of the MCF7 and SKBR3 cells.
These data thus suggested that the proliferative and migratory
capacity of BC cells was reduced following CDCA5 knockdown.
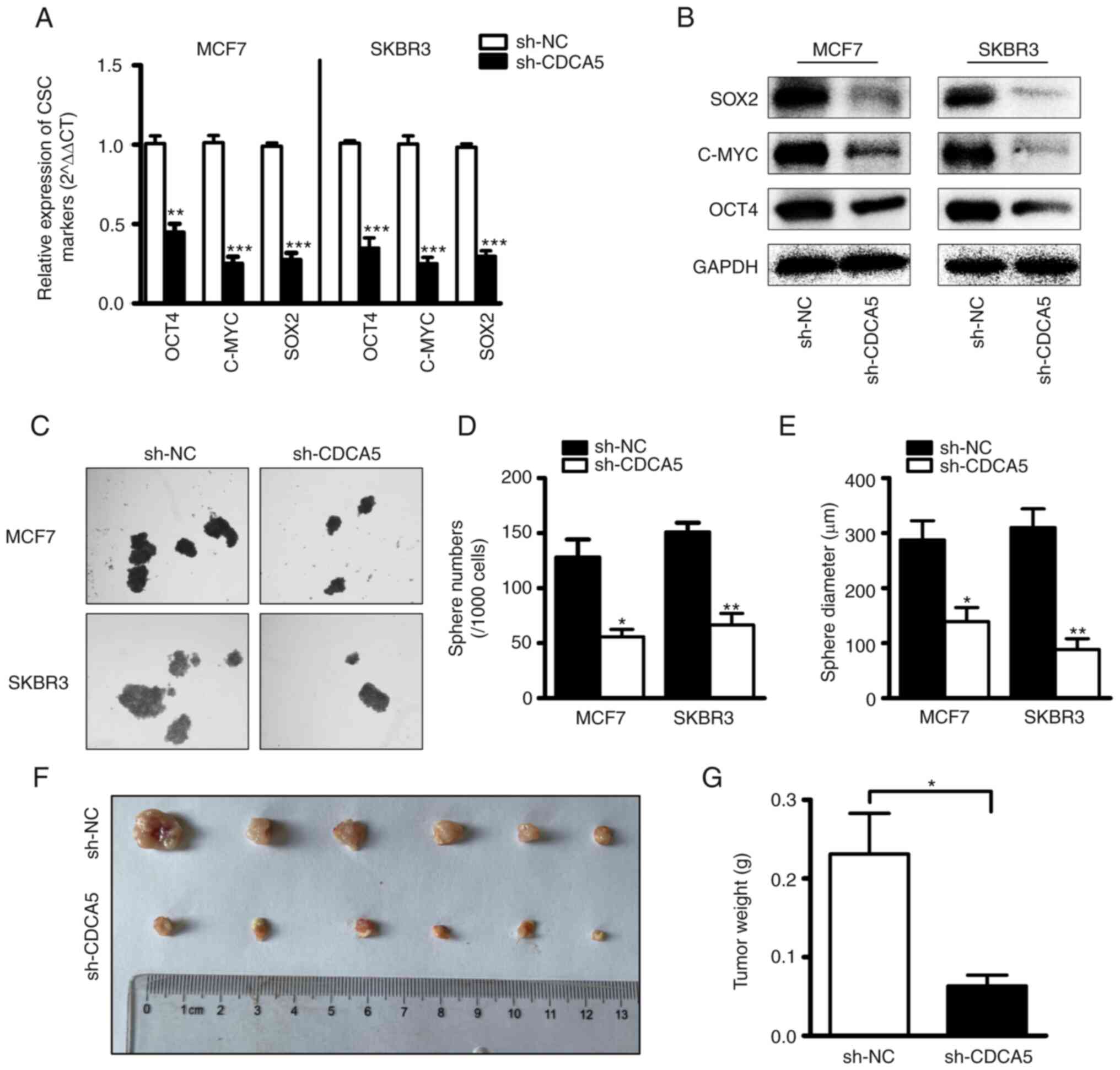
CDCA5 knockdown suppresses BC cell
stemness
To explore the role of CDCA5 in the regulation of
BCSCs, BC cells subjected to CDCA5 knockdown and their
corresponding control cells were utilized. CSC-related genes (SOX2,
OCT4 and c-MYC) have frequently been used to recognize CSCs in
clinical tissues and several cancer cell lines (29–31).
The mRNA (Fig. 5A) and protein
(Fig. 5B) levels of SOX2, OCT4 and
c-MYC were examined using RT-qPCR and western blot analysis.
Compared with the control cells, transfection with sh-CDCA5 led to
a markedly decreased expression of the stemness-related genes.
These findings indicated that CDCA induced alterations in the
levels of stem cell markers. Furthermore, the capacity of BC cells
in which CDCA5 was knocked down to form tumor spheroids was
examined. As shown in Fig. 5C, the
cells transfected with sh-CDCA5 formed less spheres than the vector
control cells, as was expected. The statistical analysis results of
the number and diameter of spheroids revealed a significant
difference between the two groups (Fig. 5D and E). It was thus concluded that
CDCA5 promotes BC cell stemness.
CDCA5 knockdown suppresses tumor
growth in vivo
To investigate the effects of CDCA5 expression on BC
growth, a mouse xenograft model was established. Since the protein
expression of CDCA5 was higher in the MCF7 cell line than in the
SKBR3 cells, the MCF7 cells were selected for use in vivo
experiments. Another reason for this selection was that the MCF7
cells are more capable of forming tumors in vivo. A total of
12 nude female mice were equally divided into two groups, and sh-NC
and sh-CDCA5 cells were subcutaneously injected. The detected ratio
of tumor growth to body weight was higher in the sh-NC group than
in the sh-CDCA5 group (Fig. 5F and
G). This indicated that the growth of MCF7 cells in vivo
was inhibited following CDCA5 knockdown.
Discussion
The mammalian cell cycle is induced by the
sequential activation or inactivation of proteins that regulate
various phases of the cell cycle (32). The loss of normal cell cycle due to
the dysregulation of several cell cycle-related genes can lead to
cancer development (33). CDCA5 is
a key regulator of DNA repair and chromosome segregation, and
belongs to a family of cell division cycle-related proteins. The
primary role of CDCA5 is to promote sister chromatid association
and ensure accurate sister chromatid segregation to maintain genome
integrity (34). CDCA5 is
aberrantly expressed in various types of cancer, such as
hepatocellular carcinoma (35–37),
bladder cancer (38), prostate
cancer (39) and renal clear cell
carcinoma (40), rendering it a
potentially important prognostic marker and potential therapeutic
target for cancer patients. However, only a limited number of
studies (41–43) to date have investigated the
expression and function of CDCA5 in BRCA.
In the present study, a significant upregulation of
CDCA5 expression was observed in BC cells. Moreover, patients with
a high expression of CDCA5 were found to have a worse survival. The
GO and KEGG function enrichment analysis revealed that CDCA5 was a
critical regulator of the cell cycle and DNA repair. Furthermore,
the knockdown of CDCA5 significantly inhibited cell growth and
tumorigenesis in vitro and in vivo. Therefore,
consistent with the findings of previous research (43), the results of the present study
support an oncogenic role for CDCA5 in BC progression and suggest
the possibility of the use of CDCA5 as a therapeutic target for
breast cancer.
The CSC theory highlights that CSCs, a minor
population of tumor cells, also referred to as tumor-initiating
cells, harbor the properties of self-renewal, differentiation and
drug-resistance, which contribute to metastasis and relapse
(44,45). The present study further explored
the association of CDCA5 with CSCs to examine whether CDCA5 is
involved in the regulation of CSCs. As was predicted, CDCA5 was
significantly upregulated in BCSCs, accompanied by the significant
upregulation of cancer stem cell transcription factors, including
SOX2, OCT4, c-MYC. CDCA5 knockdown significantly inhibited the
expression of CSC-related transcription factors, and significantly
inhibited mammosphere formation, such as the size and number of
tumor spheroids. Of course, the present study has limitations
compared to other studies on CDCA5. For example, the present study
did not further explore and examine the in-depth mechanisms
underlying the regulation of BCSCs by CDCA5, but only observed that
CDCA5 regulated BCSCs. In addition, it is unknown whether CDCA5
also has the ability to regulate CSCs in other types of cancer. In
the tumor microenvironment, the association between CDCA5 and
immune infiltration is also worthy of further study.
In conclusion, the present study indicated that
CDCA5 plays a critical role in BC progression by promoting BC cell
proliferation, migration and cellular stemness. Therefore, CDCA5
may serve as a novel prognostic biomarker and therapeutic target
for patients with BC.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
The authors would like to thank Dr Cao Yuan at the
Analytical and Testing Center of Wuhan University of Science and
Technology (Wuhan, China) for providing assistance with the flow
cytometric analysis.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant nos. 31501149, 31770815 and 31570764),
the Wuhan Health and Family Planning Scientific Research Project
(grant no. WX21Q49), the Hubei Natural Science Foundation (grant
nos. 2017CFB537, 2019CFB398, 2019CFB368 and 2021CFB230), the
Educational Commission of Hubei (grant no. B2020001), the Hubei
Province Health and Family Planning Scientific Research Project
(grant nos. WJ2021Q051, WJ2019M255 and ZY2021Q005), the Frontier
Project of Applied Basic Research in Wuhan (grant no.
2020020601012250) and the Hubei Province Technology Innovation
Special Major Project (Project no. 2019ACA168).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
XHL and TCZ participated in the research design. HH,
YX, XYZ, YD, FJW and YH performed the experiments and analyzed the
data, and HH was the major contributor to the writing of the
manuscript. HH and YX confirm the authenticity of all the raw data.
All the authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The animal research protocol was reviewed and
approved by the Animal Protection Committee of Wuhan University of
Science and Technology (Wuhan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Loibl S, Poortmans P, Morrow M, Denkert C
and Curigliano G: Breast cancer. Lancet. 397:1750–1769. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Ma J, Gaudet MM, Newman LA,
Miller KD, Goding Sauer A, Jemal A and Siegel RL: Breast cancer
statistics, 2019. CA Cancer J Clin. 69:438–451. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ginsburg O, Bray F, Coleman MP, Vanderpuye
V, Eniu A, Kotha SR, Sarker M, Huong TT, Allemani C, Dvaladze A, et
al: The global burden of women's cancers: A grand challenge in
global health. Lancet. 389:847–860. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kreso A and Dick JE: Evolution of the
cancer stem cell model. Cell Stem Cell. 14:275–291. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gupta PB, Fillmore CM, Jiang G, Shapira
SD, Tao K, Kuperwasser C and Lander ES: Stochastic state
transitions give rise to phenotypic equilibrium in populations of
cancer cells. Cell. 146:633–644. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Peng F, Li TT, Wang KL, Xiao GQ, Wang JH,
Zhao HD, Kang ZJ, Fan WJ, Zhu LL, Li M, et al: H19/let-7/LIN28
reciprocal negative regulatory circuit promotes breast cancer stem
cell maintenance. Cell Death Dis. 8:e25692017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Charafe-Jauffret E, Ginestier C, Bertucci
F, Cabaud O, Wicinski J, Finetti P, Josselin E, Adelaide J, Nguyen
TT, Monville F, et al: ALDH1-positive cancer stem cells predict
engraftment of primary breast tumors and are governed by a common
stem cell program. Cancer Res. 73:7290–7300. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rankin S, Ayad NG and Kirschner MW:
Sororin, a substrate of the anaphase-promoting complex, is required
for sister chromatid cohesion in vertebrates. Mol Cell. 18:185–200.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gaudet P, Livstone MS, Lewis SE and Thomas
PD: Phylogenetic-based propagation of functional annotations within
the Gene Ontology consortium. Brief Bioinform. 12:449–462. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schmitz J, Watrin E, Lénárt P, Mechtler K
and Peters JM: Sororin is required for stable binding of cohesin to
chromatin and for sister chromatid cohesion in interphase. Curr
Biol. 17:630–636. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nishiyama T, Ladurner R, Schmitz J, Kreidl
E, Schleiffer A, Bhaskara V, Bando M, Shirahige K, Hyman AA,
Mechtler K and Peters JM: Sororin mediates sister chromatid
cohesion by antagonizing Wapl. Cell. 143:737–749. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Watrin E, Demidova M, Watrin T, Hu Z and
Prigent C: Sororin pre-mRNA splicing is required for proper sister
chromatid cohesion in human cells. EMBO Rep. 15:948–955. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Boyle MI, Jespersgaard C, Nazaryan L, Ravn
K, Brøndum-Nielsen K, Bisgaard AM and Tümer Z: Deletion of
11q12.3-11q13.1 in a patient with intellectual disability and
childhood facial features resembling Cornelia de Lange syndrome.
Gene. 572:130–134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Safran M, Rosen N, Twik M, BarShir R,
Stein TI, Dahary D, Fishilevich S and Lancet D: The GeneCards
Suite. Practical Guide Life Sci Databases; pp. 27–56. 2021
|
|
16
|
Nguyen MH, Koinuma J, Ueda K, Ito T,
Tsuchiya E, Nakamura Y and Daigo Y: Phosphorylation and activation
of cell division cycle associated 5 by mitogen-activated protein
kinase play a crucial role in human lung carcinogenesis. Cancer
Res. 70:5337–5347. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang IW, Lin VC, He HL, Hsu CT, Li CC, Wu
WJ, Huang CN, Wu TF and Li CF: CDCA5 overexpression is an indicator
of poor prognosis in patients with urothelial carcinomas of the
upper urinary tract and urinary bladder. Am J Transl Res.
7:710–722. 2015.PubMed/NCBI
|
|
18
|
Tokuzen N, Nakashiro K, Tanaka H, Iwamoto
K and Hamakawa H: Therapeutic potential of targeting cell division
cycle associated 5 for oral squamous cell carcinoma. Oncotarget.
7:2343–2353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Shen M and Zhou G: Upregulation
of CDCA5 promotes gastric cancer malignant progression via
influencing cyclin E1. Biochem Biophys Res Commun. 496:482–489.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen T, Huang Z, Tian Y, Wang H, Ouyang P,
Chen H, Wu L, Lin B and He R: Role of triosephosphate isomerase and
downstream functional genes on gastric cancer. Oncol Rep.
38:1822–1832. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen Z, Yu X, Zheng Y, Lai X, Li J, Hong
Y, Zhang H, Chen C, Su Z and Guo R: CDCA5 regulates proliferation
in hepatocellular carcinoma and has potential as a negative
prognostic marker. Onco Targets Ther. 11:891–901. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi B and Varambally
S: UALCAN: A portal for facilitating tumor subgroup gene expression
and survival analyses. Neoplasia. 19:649–658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Győrffy B: Survival analysis across the
entire transcriptome identifies biomarkers with the highest
prognostic power in breast cancer. Comput Struct Biotechnol J.
19:4101–4109. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vasaikar SV, Straub P, Wang J and Zhang B:
LinkedOmics: Analyzing multi-omics data within and across 32 cancer
types. Nucleic Acids Res. 46:D956–D963. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:(Database Issue). D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dontu G, Abdallah WM, Foley JM, Jackson
KW, Clarke MF, Kawamura MJ and Wicha MS: In vitro propagation and
transcriptional profiling of human mammary stem/progenitor cells.
Genes Dev. 17:1253–1270. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saadin K and White IM: Breast cancer stem
cell enrichment and isolation by mammosphere culture and its
potential diagnostic applications. Expert Rev Mol Diagn. 13:49–60.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qin Y, Hou Y, Liu S, Zhu P, Wan X, Zhao M,
Peng M, Zeng H, Li Q, Jin T, et al: A Novel long non-coding RNA
lnc030 maintains breast cancer stem cell stemness by stabilizing
SQLE mRNA and increasing cholesterol synthesis. Adv Sci (Weinh).
8:20022322021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ai L, Mu S, Sun C, Fan F, Yan H, Qin Y,
Cui G, Wang Y, Guo T, Mei H, et al: Myeloid-derived suppressor
cells endow stem-like qualities to multiple myeloma cells by
inducing piRNA-823 expression and DNMT3B activation. Mol Cancer.
18:882019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang T, Guo C, Xia T, Zhang R, Zen K, Pan
Y and Jin L: LncCCAT1 promotes breast cancer stem cell function
through activating WNT/β-catenin signaling. Theranostics.
9:7384–7402. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma XL, Hu B, Tang WG, Xie SH, Ren N, Guo L
and Lu RQ: CD73 sustained cancer-stem-cell traits by promoting SOX9
expression and stability in hepatocellular carcinoma. J Hematol
Oncol. 13:112020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dominguez-Brauer C, Thu KL, Mason JM,
Blaser H, Bray MR and Mak TW: Targeting mitosis in cancer: Emerging
strategies. Mol Cell. 60:524–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
López-Lázaro M: The stem cell division
theory of cancer. Crit Rev Oncol Hematol. 123:95–113. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang N and Pati D: Sororin is a master
regulator of sister chromatid cohesion and separation. Cell Cycle.
11:2073–2083. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen H, Chen J, Zhao L, Song W, Xuan Z,
Chen J, Li Z, Song G, Hong L, Song P and Zheng S: CDCA5,
transcribed by E2F1, promotes oncogenesis by enhancing cell
proliferation and inhibiting apoptosis via the AKT pathway in
hepatocellular carcinoma. J Cancer. 10:1846–1854. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tian Y, Wu J, Chagas C, Du Y, Lyu H, He Y,
Qi S, Peng Y and Hu J: CDCA5 overexpression is an Indicator of poor
prognosis in patients with hepatocellular carcinoma (HCC). BMC
Cancer. 18:11872018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang J, Xia C, Pu M, Dai B, Yang X, Shang
R, Yang Z, Zhang R, Tao K and Dou K: Silencing of CDCA5 inhibits
cancer progression and serves as a prognostic biomarker for
hepatocellular carcinoma. Oncol Rep. 40:1875–1884. 2018.PubMed/NCBI
|
|
38
|
Fu G, Xu Z, Chen X, Pan H, Wang Y and Jin
B: CDCA5 functions as a tumor promoter in bladder cancer by
dysregulating mitochondria-mediated apoptosis, cell cycle
regulation and PI3k/AKT/mTOR pathway activation. J Cancer.
11:2408–2420. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ji J, Shen T, Li Y, Liu Y, Shang Z and Niu
Y: CDCA5 promotes the progression of prostate cancer by affecting
the ERK signalling pathway. Oncol Rep. 45:921–932. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Huang X, Huang Y, Lv Z, Wang T, Feng H,
Wang H, Du S, Wu S, Shen D, Wang C, et al: Loss of cell division
cycle-associated 5 promotes cell apoptosis by activating DNA damage
response in clear cell renal cell carcinoma. Int J Oncol.
61:872022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Phan NN, Wang CY, Li KL, Chen CF, Chiao
CC, Yu HG, Huang PL and Lin YC: Distinct expression of CDCA3,
CDCA5, and CDCA8 leads to shorter relapse free survival in breast
cancer patient. Oncotarget. 9:6977–6992. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jin X, Wang D, Lei M, Guo Y, Cui Y, Chen
F, Sun W and Chen X: TPI1 activates the PI3K/AKT/mTOR signaling
pathway to induce breast cancer progression by stabilizing CDCA5. J
Transl Med. 20:1912022. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang Y, Yao J, Zhu Y, Zhao X, Lv J and Sun
F: Knockdown of CDCA5 suppresses malignant progression of breast
cancer cells by regulating PDS5A. Mol Med Rep. 25:2092022.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gerger A, Zhang W, Yang D, Bohanes P, Ning
Y, Winder T, LaBonte MJ, Wilson PM, Benhaim L, Paez D, et al:
Common cancer stem cell gene variants predict colon cancer
recurrence. Clin Cancer Res. 17:6934–6943. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fabrizi E, di Martino S, Pelacchi F and
Ricci-Vitiani L: Therapeutic implications of colon cancer stem
cells. World J Gastroenterol. 16:3871–3877. 2010. View Article : Google Scholar : PubMed/NCBI
|