Introduction
In 2020, prostate cancer (PCa) became the most
frequently diagnosed cancer in 112 countries and the leading cause
of cancer-related mortality in 48 countries among men (1). In recent decades, researchers have
found that the activation of androgen receptor (AR) signalling
plays a crucial role in the development of PCa (2). Thus, androgen deprivation therapy
(ADT) is an important and standard treatment for PCa. In China, the
majority of patients with PCa have progressed to an advanced stage
at the time of diagnosis, and ADT has been adopted as the
first-line treatment when seeking treatment (3). However, the effect of ADT on
prolonging the survival time of patients with PCa is not
satisfactory. Upon ADT, almost all patients will further develop
castration-resistant prostate cancer (CRPC), with a mean survival
time of only 14.5 months (4,5).
Hence, there is an urgent need to explore the specific mechanism of
CRPC and ameliorate the prognosis of patients with CRPC.
Most patients with CRPC exhibit a continuous
increase in prostate-specific antigen (PSA) levels despite the use
of ADT and testosterone levels. The increase in PSA levels
indicates that intratumoural androgen biosynthesis is functional in
CRPC cells (6). Intratumoural
androgen is considered the key factor for the continued activation
of AR in CRPC. After binding with intratumoural androgen, the
inactive AR protein changes its conformation, translocates from the
cytoplasm to the nucleus, and forms AR dimers (7). Furthermore, the AR dimer is able to
bind to androgen response elements (AREs), such as the promoter or
enhancer regions of downstream genes (7), which promote the development of PCa.
Although new hormonal agents (e.g., enzalutamide or abiraterone)
have been shown to prolong the overall survival time of patients
with CRPC by blocking intratumoural androgen biosynthesis and
activity, most of the patients will develop resistance to hormonal
agents in only several months (8,9). The
reincrease in PSA in drug-resistant patients with CRPC suggests
that the AR signalling pathway is reactivated in
androgen-independent conditions (6,9). A
previous study by the authors confirmed that the AR protein could
still be dimerized and bind to ARE as well as promote the
transcription of its downstream genes under androgen deprivation
conditions, which conferred CRPC cell growth. By contrast, the
dimerization and activity of AR as well as the growth of the tumour
cells were inhibited after mutation of the dimerization sites (F23,
27A/L26A and A596T/S597T) (10).
Therefore, the AR protein still plays a key role in the
reactivation of the AR signalling pathway even under androgen
deprivation conditions. It is important to identify target genes
that can interact with AR proteins and regulate the ability of AR
proteins to bind to AREs.
High-mobility group protein B1 (HMGB1), which plays
an important role in the activation of transcription factor
signalling pathways, has been identified to be involved in the
occurrence and development of a variety of tumours, including
non-small-cell lung, cervical, and colorectal cancer (11). Moreover, an increasing number of
studies have demonstrated that the overexpression of HMGB1 is
related to the poor prognosis of PCa and CRPC (12,13).
Although these studies initially demonstrated that HMGB1 was
associated with the progression, clinical pathological
characteristics, and poor prognosis of PCa, the molecular mechanism
by which HMGB1 promotes the development of CRPC remains unclear. A
previous study suggested that HMGB1 enhances oestrogen receptor
(ER) dimer binding to the oestrogen response element (ERE) and then
enhances the transcription of its downstream genes (14). Notably, the protein structure
domain and active form (dimerization) of ER were similar to those
of AR (15), which implied that
the activation of ER and AR may be regulated by the same regulatory
factors. Additionally, some studies have indicated that estrone
sulfate and dehydroepiandrosterone sulfate could enhance the
transcriptional activity of ER and AR simultaneously (16). RO48-8071 was demonstrated to
inhibit the transcriptional activity of ER as well as AR (17). Hence, it is surmised that HMGB1 may
interact with AR protein, promote the ability of AR protein to bind
to ARE, and ultimately promote the development of CRPC.
To the best of our knowledge, there are no other
studies reporting that HMGB1 protein could regulate AR
transactivating activity, which is also well known as AR
transcriptional activity, in PCa. The aim of the present study was
to investigate the association between HMGB1 and/or AR expression
and the pathological features of patients with PCa to verify the
regulatory effect and mechanisms of HMGB1 on AR transactivating
activity in order to provide a theoretical basis for applying novel
targeted drugs for CRPC treatment.
Materials and methods
Bioinformatics analysis
The Oncomine (https://www.oncomine.org/resource/login.html), GEPIA
(http://gepia.cancer-pku.cn/index.html), and DepMap
Portal (https://depmap.org/portal/interactive/) databases were
used to analyse the gene expression levels of HMGB1 and AR in the
prostate gland, PCa tissues, androgen-dependent PCa cells (LNCaP
and MDAPCA2B), and CRPC cells (VCaP and 22RV1). The data retrieved
from Oncomine originated from GSE6099 (18), GSE68907 (19), GSE6919 (20) and a study of Vanaja et al
(21). Furthermore, the
correlation between HMGB1 mRNA expression and AR mRNA expression
was analysed using the GEPIA database.
Specimens
The present study was approved by the Ethics
Committee of the School of Nursing, Jilin University (Changchun,
China). Paraffin-embedded specimens were collected from 2001–2009
at the Prostate Diseases Prevention and Treatment Research Centre
of Jilin University (Changchun, China). All patients were diagnosed
during a PCa screening study for high-risk males. The patients were
informed and agreed to participate in the PCa-relative study. The
inclusion criteria were as follows: i) Male and over 50 years old;
ii) the level of serum PSA exceeded 4 pg/ml or an existing prostate
tumor suggested by imaging; iii) initially diagnosed with PCa; and
iv) the pathological diagnoses were determined as prostate
adenocarcinoma by an experienced urological pathologist working at
the Second Hospital of Jilin University. The exclusion criteria
were as follows: i) Individual history of other malignant tumors;
ii) the patients had previously undergone radical prostatectomy or
other treatments, such as hormone treatment or radiotherapy
(22). Five patients with benign
prostate hyperplasia (BPH) were included as control subjects. The
clinicopathological features of the patients included are presented
in Table I. The
clinicopathological features of prostatic needle biopsy samples are
shown in Table II.
 | Table I.Clinicopathological features of
patients with PCa. |
Table I.
Clinicopathological features of
patients with PCa.
| Clinical
features | No. of cases
(%) |
|---|
| Age, years |
|
|
<73 | 25 (49.02) |
|
≥73 | 26 (50.98) |
| Pathological
grade |
|
|
BPH | 5 (9.80) |
|
GS=6 | 10 (19.61) |
|
GS=7 | 10 (19.61) |
|
GS=8 | 11 (21.57) |
|
GS=9 | 12 (23.53) |
|
GS=10 | 3 (5.88) |
| PSA level,
pg/ml |
|
|
<54.3 | 25 (49.02) |
|
≥54.3 | 26 (50.98) |
| Table II.Clinicopathological features of
prostatic needle biopsy samples. |
Table II.
Clinicopathological features of
prostatic needle biopsy samples.
| Samples | Case no. | Mean age
(range) | Mean PSA level
(range) |
|---|
| BPH | 5 | 75 (67–84) | 19.3
(9.1-29.44) |
| GS=6 | 10 | 77.4 (63–83) | 33.24 (4-96.8) |
| GS=7 | 10 | 68.4 (56–82) | 83.95
(9.4-300) |
| GS=8 | 11 | 70.9 (64–84) | 82.96
(28-254.3) |
| GS=9 | 12 | 73.25 (54–90) | 75.48
(3-126.1) |
| GS=10 | 3 | 75 (70–78) | 29.9 (1.7-49) |
Immunohistochemistry
Immunohistochemical (IHC) staining was performed as
previously described (23).
Briefly, after being fixed in 10% formalin at room temperature for
24 h, PCa samples or mouse tumour samples were embedded in paraffin
and cut into 3-µm-thick sections. After deparaffinization by xylol
and rehydration by a descending alcohol series, the sections were
soaked in EDTA retrieval buffers and heated in a microwave oven.
Nonspecifc binding was blocked by 5% bovine serum albumin (product
code AR1006; Boster Biological Technology, Inc.) at room
temperature for 20 min. Subsequently, the histological sections
were stained with rabbit anti-AR antibody (product code ab74272;
1:200; Abcam) and anti-HMGB1 antibody (product code A00066-1;
1:200; Boster Biological Technology, Inc.) at 4°C overnight. Goat
anti-rabbit IgG conjugated with horseradish peroxidase (cat. no.
SA00001-2; 1:200; ProteinTech Group, Inc.) was used as the
secondary antibody, and this staining procedure was carried out at
37°C for 30 min. Reactive products were visualized with
3,3′-diaminobenzidene (Boster Biological Technology, Inc.) as the
chromogen, and the sections were counterstained with 0.1%
haematoxylin (Boster Biological Technology, Inc.) at room
temperature for 2 min. Histological images were captured under a
light microscope (Olympus Corporation) with an objective
magnification of ×200 or ×400. The positive cell density was
assessed using Image-Pro Plus 6.0 software (Media Cybernetics,
Inc.), and the results are presented as the mean optical density
(MOD) values (22).
PCa cell lines and reagents
PC3 (ATCC no. CRL-1435), C4-2 (ATCC no. CRL-3314),
VCaP (ATCC no. CRL-2876), and DU145 (ATCC no. HTB-81) cells were
obtained from the American Type Culture Collection. LNCaP95 cells
were provided from the College of Life Science of Jilin University
(Changchun, China). LNCaP (cat. no. CL-0143) and 22RV1 (cat. no.
CL-0004) cells were obtained from the Procell Life Science &
Technology Co., Ltd. These cells were cultured in RPMI-1640 medium
(HyClone; Cytiva) supplemented with 10% foetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) in a humidified atmosphere
with 5% CO2 at 37°C.
Lentiviral vector plasmid pLVX-Puro (cat. no.
632164; Clontech; Takara Bio USA, Inc.) was used to construct
pLVX-HMGB1 plasmid and pLVX-shRNA1 (cat. no. 632177; Clontech;
Takara Bio USA, Inc.) was used to construct PLVX-shHMGB1,
PLVX-shAR, and PLVX-shScramble plasmids. The interference sequences
were as follows: shHMGB1, 5′-GGACAAGGCCCGTTATGAA-3′; shAR,
5′-TAGTGCAATCATTTCTGCTGGC-3′; and shScramble,
5′-GTATAAGTCAACTGTTGAC-3′. The lentiviruses used in this study were
packaged using a 4th generation lentiviral packaging system (cat.
no. 631275; Clontech; Takara Bio USA, Inc.) according to the
manufacturer's instructions. Briefly, after mixing the lentiviral
vector plasmids with premixed complex including five lentiviral
packaging plasmids and transfection reagent, the nanoparticle
complex solution was added into Lenti-X 293T cells (cat. no.
632180; Clontech; Takara Bio USA, Inc.). After 24 and 48 h, the
media was collected and concentrated using the Lenti-X Concentrator
(cat. no. 631231; Clontech; Takara Bio USA, Inc.) according to the
manufacturer's instructions. The concentration medium was used to
detect the titer of the virus using a Lenti-X qRT-PCR Titration Kit
(cat. no. 631235; Clontech; Takara Bio USA, Inc.) according to the
manufacturer's instructions, and then stored at −80°C.
To generate LNCaP-neo and LNCaP-HMGB1 cells, LNCaP
cells were infected with 30 MOI of pLVX-HMGB1 containing the HMGB1
expression gene or the empty vector pLVX (the final titer of the
virus was 7.5×105 IFU/ml). The media were changed 48 h
later. For stable transfection, the cells were selected in medium
containing puromycin (2 µM) and maintained in medium containing
puromycin (1 µM). Puromycin selection was continued for 1 week
prior to harvesting cells for downstream analysis, or 2 weeks prior
to further knock down AR. To detect the effect of HMGB1 on AR
transcriptional activity, LNCaP-HMGB1 cells were further infected
with 30 MOI of pLVX-shAR containing AR shRNA or pLVX-shScramble
(the final titer of the virus was 7.5×105 IFU/ml). To
generate 22RV1-shScramble and 22RV1-shHMGB1 cells, 22RV1 cells were
infected with 30 MOI of pLVX-shHMGB1 containing HMGB1 shRNA or
pLVX-shScramble (the final titer of the virus was
7.5×105 IFU/ml).
Cell Counting Kit-8 (CCK-8) assay
LNCaP-neo, LNCaP-HMGB1, 22RV1-shScramble and
22RV1-shHMGB1 cells were seeded into 96-well plates (1,000
cells/well) and cultured in RPMI-1640 medium (HyClone; Cytiva)
supplemented with 10% charcoal-stripped FBS (Thermo Fisher
Scientific, Inc.). Subsequently, 48 and 96 h later, CCK-8 reagent
(10 µl/well; BIOSS) was added to each well. Following culture at
37°C for 1.5 h, the absorbance of each well was measured at 450 nm
using a microplate reader (Cytation 5; BioTek Instruments,
Inc.).
Reporter gene assay
The reporter gene assay was performed according to
the manufacturer's instructions. Briefly, LNCaP-neo, LNCaP-HMGB1,
22RV1-shScramble and 22RV1-shHMGB1 cells were transiently
transfected with an ARE-luciferase reporter plasmid using
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.).
The ARE-luciferase reporter plasmid was provided by Dr Robert
Matusik at Vanderbilt University School of Medicine (Nashville,
USA) as indicated in a previous study by the authors (24). After incubating with the
transfection mixture at 37°C for 3 h, the cells were divided
equally into 24-well plates and cultured in charcoal-stripped
medium for 96 h. Subsequently, the cells were lysed in reporter
lysis buffer (Promega Corporation), and the luciferase activity was
determined using the Luciferase Assay System (Promega Corporation)
and normalized based on the protein concentrations for each sample
(24).
Reverse transcription-quantitative PCR
(RT-qPCR)
LNCaP-neo, LNCaP-HMGB1, 22RV1-shScramble and
22RV1-shHMGB1 cells were seeded into 6-well plates at a density of
3×105 cells/well. After being treated with
charcoal-stripped medium for 96 h, the total RNA was extracted
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
and first-strand cDNA synthesis was performed using a cDNA
synthesis kit (cat. no. AT311-02; TransGen Biotech Co., Ltd.)
according to the manufacturer's instructions. Real-time PCR was
used to detect the transcription levels of PSA and TMPRSS2 using
the SYBR Green qPCR kit (cat. no. AQ132; TransGen Biotech Co.,
Ltd.) according to the manufacturer's instructions, and GAPDH was
used as an internal control gene. The following primers were used:
TMPRSS2 forward, 5′-ACACACCGATTCTCGTCCT-3′ and reverse,
5′-TGGCCTACTCTGGAAGTTCA-3′; PSA forward, 5′-CGATTCTTCAGGAGGCTCAT-3′
and reverse, 5′-GCTGCCCACTGCATCAG-3′; and GAPDH forward,
5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse, 5′-GAAGATGGTGATGGGATTTC-3′.
PCR was performed at 94°C for 30 sec, followed by 45 cycles of
amplification at 94°C for 5 sec, 51°C for 15 sec and 72°C for 10
sec using ABI-Q3 (Thermo Fisher Scientific, Inc.). mRNA expression
levels were quantified using 2−ΔΔCq method (25).
Western blotting
Soluble proteins were extracted from LNCaP, 22RV1,
and PC3 cells treated by the indicated manner using RIPA lysis
(Beyotime Institute of Biotechnology). The protein concentrations
were measured by a Coomassie Plus (Bradford) Assay Kit (Thermo
Fisher Scientific, Inc.). Proteins, at a concentration of 15 µg,
were then subjected to each lane of a 4–15% Precast Tris-Glycine
Gel (TransGen Biotech Co., Ltd.). Prestained standards (TransGen
Biotech Co., Ltd.) were used as molecular weight markers. Separated
proteins were electrophoretically transferred onto polyvinylidene
difluoride (PVDF) membranes (iBlot system; Invitrogen; Thermo
Fisher Scientific, Inc.). The membranes were then blocked with 5%
skim milk for 1 h at room temperature, and incubated with primary
antibodies, including anti-AR (product code ab74272; 1:100; Abcam),
anti-HMGB1 (product code A00066-1; 1:1,000; Boster Biological
Technology, Inc.), and anti-GAPDH (product code ab245355; 1:5,000;
Abcam), at room temperature for 2 h. Subsequently, following
incubation with secondary-HRP antibodies (cat. no. SA00001-2;
1:20,000; ProteinTech Group, Inc.) at room temperature for 1 h, the
protein levels were detected with an ECL plus kit and developed via
an imaging densitometer (both from Clinx Science Instruments Co.,
Ltd.). The assay was repeated three times independently.
Bioluminescence resonance energy
transfer (BRET) assay
To further detect the interaction between HMGB1 and
AR protein, PC3 cells were co-transfected with an HMGB1-RLuc BRET
fusion plasmid (Donor) and an AR-Turbo BRET fusion plasmid
(Receptor). After being cultured in 10% charcoal-stripped medium
for 48 h, the cells were detached with 5 mM EDTA in PBS and
resuspended in PBS with 1% sucrose. Subsequently, the cells were
counted and seeded in triplicate into a 96-well white-wall
microplate at 1×105 cells/well. Freshly prepared
coelenterazine (Nanolight Technologies; Prolume Ltd.) in water was
added to the cells at a final concentration of 25 µM. BRET readings
at 528 and 635 nm were obtained immediately with a Synergy 2
microplate reader (BioTek Instruments, Inc.). The BRET ratio was
calculated by subtracting the ratio of the 635-nm emission and the
528-nm emission obtained from cells coexpressing the HMGB1-RLuc and
AR-Turbo fusion proteins from the background BRET ratio resulting
from cells expressing the RLuc fusion protein alone in the same
experiment: BRET ratio=(emission at 635 nm)/(emission at 528
nm)-(emission at 635 nm RLuc only)/(emission at 528 nm RLuc only)
(10).
Chromatin immunoprecipitation (ChIP)
assay
To illustrate the effect of HMGB1 on the
reactivation of the AR signalling pathway, a Magnetic Bead ChIP
assay kit (cat. no. 26157; Thermo Fisher Scientific, Inc.) was used
with an AR antibody (product code ab108341; Abcam) to determine the
effect of HMGB1 on AR binding events at known AREs of the PSA and
TMPRSS2 genes. The ChIP assay was performed according to the
manufacturer's instructions. Briefly, after being treated with
charcoal-stripped medium for 48 h, the cells were incubated with 1%
formaldehyde at room temperature for 10 min. The nuclear
DNA-protein complexes of LNCaP-neo, LNCaP-HMGB1, 22RV1-shScramble,
and 22RV1-shHMGB1 cells were extracted using Lysis Buffer (200 µl
for each IP reaction) of the kit, and then were centrifuged at
9,000 × g for 5 min (4°C). Subsequently, the nuclei was sonicated
to produce 200–500 bp fragments. The chromatin extract was
incubated with antibodies against AR (2 µg for each IP reaction),
10 µl RNA Polymerase II (Thermo Fisher Scientific, Inc.), or 2 µl
IgG (Thermo Fisher Scientific, Inc.) overnight at 4°C. A uniform
suspension of 20 µl containing Protein A/G Magnetic Beads was added
to each IP reaction and incubated for 2 h at 4°C with mixing.
Following incubation, the beads were collected and washed three
times with IP Wash Buffer 1. After immunoprecipitation, the IP
Elution Buffer was used to elute the protein-chromatin complex from
the beads at 65°C for 30 min, and the complex was then treated with
proteinase K at 65°C for 1.5 h. Finally, the purified DNA was
analysed using RT-qPCR.
In vivo effect of HMGB1 on the
development of PCa
A total number of 20 male BALB/c nu/nu mice (age,
seven weeks old; weight, 15–20 g) were purchased from Beijing Vital
River Laboratory Animal Technology Co., Ltd. The animal experiments
were approved by the Animal Ethics Committee of Basic Medical
College of Jilin University (Changchun, China), complied with the
recommendations of the ARRIVE guidelines, and were carried out in
accordance with the Good Laboratory Practice Regulations.
Approximately 0.1 ml of LNCaP-neo or LNCaP-HMGB1 cells was mixed
with 50% Matrigel and subcutaneously implanted into the right flank
region of BALB/c nu/nu mice at a dosage of 4×106
cells/mouse. When the tumour volume reached 100 mm3, the
mice were subjected to sham surgery (intact group) or castration
(castration group). The tumour volume was calculated as follows:
V=L × W2/2, where L and W represent the length and width
of the tumour, respectively (26).
The weight and tumour volume were measured weekly. When the volume
of tumours in the intact group or castration group reached 2,000
mm3, all mice were euthanized by cervical dislocation
under 2–3% isoflurane anaesthesia, and tumour samples were weighed
and collected.
ELISA
After mice were anaesthetized with isoflurane (a
concentration of 2 to 3% for induction and 1.5 to 2% for
maintenance), mouse serum was collected and used for PSA detection.
The level of PSA was determined by an ELISA kit (cat. no.
MM-0421H1; Meimian; http://www.mmbio.cn/) according to the manufacturer's
instructions. Finally, the absorbance was detected at 450 nm using
a microplate reader (BioTek Instruments, Inc.).
Statistical analysis
Statistical analysis was performed with multiple
comparisons using one-factor analysis of variance (ANOVA) followed
by Dunnett's post hoc test or Bonferroni's correction test, and the
differences among two dependent samples were analysed using an
unpaired Student's t-test. The linear relationships between two
continuous variables were analyzed by Pearson's correlation
analysis. Spearman's correlation analysis was used to determine the
correlations between rank variables and continuous variables. All
data were analysed with the statistical software SPSS 17.0 (SPSS,
Inc.), and the results are expressed as the means ± SD. The results
were repeated three times independently. P<0.05 was considered
to indicate a statistically significant difference.
Results
Association of AR and HMGB1 mRNA
levels in PCa
The associations between the mRNA levels of AR and
HMGB1 in PCa were analysed using the Oncomine and GEPIA databases.
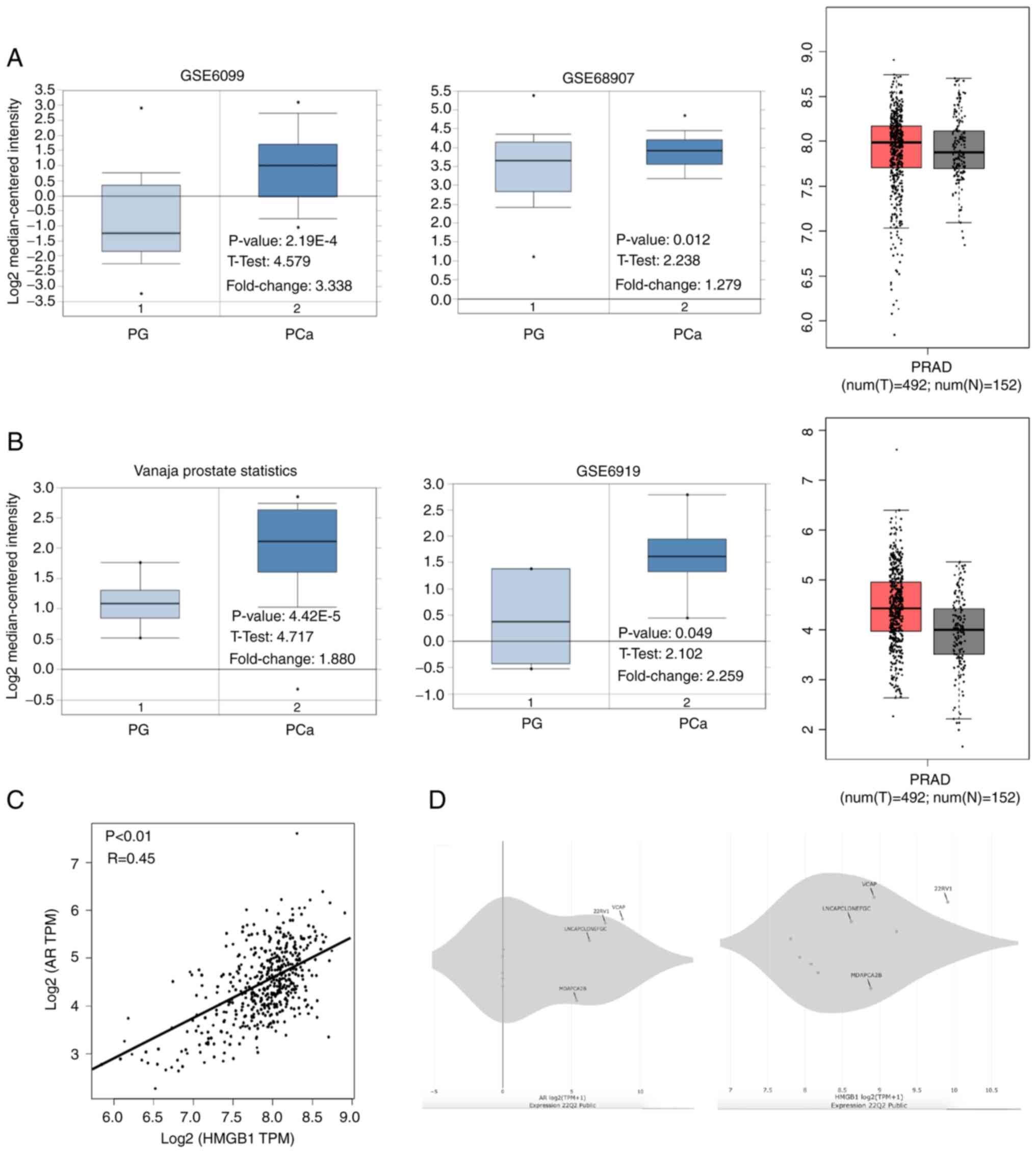
As shown in Fig. 1A and B, the
mRNA levels of AR and HMGB1 were increased in PCa tissues compared
with normal prostate glands (P<0.05). Interestingly, a
significant correlation between AR and HMGB1 mRNA expression in
prostate tissues was found using Pearson analysis in the GEPIA
database (R=0.45, P<0.01; Fig.
1C). Additionally, the data showed that the mRNA level of AR
and HMGB1 was increased in CPRC cells (VCaP and 22RV1) compared
with androgen-dependent PCa cells (LNCaP and MDAPCA2B) (Fig. 1D). The data also revealed that the
expression levels of AR protein and HMGB1 protein were higher in
22RV1 cells than that in LNCaP cells (Fig. S1). LNCaP cells were employed to
overexpress HMGB1 because of its low expression of HMGB1 and
androgen-dependent feature. Moreover, although HMGB1 was
upregulated in both 22RV1 and LNCaP95 cells compared with the other
PCa cell lines, it was determined that 22RV1 cells exhibited a
higher HMGB1 expression than that in LNCaP95 cells, indicating that
22RV1 cells may be more suitable for silencing HMGB1 in the
subsequent experiments (Fig.
S1).
Association of AR and HMGB1 proteins
in human prostate tissues
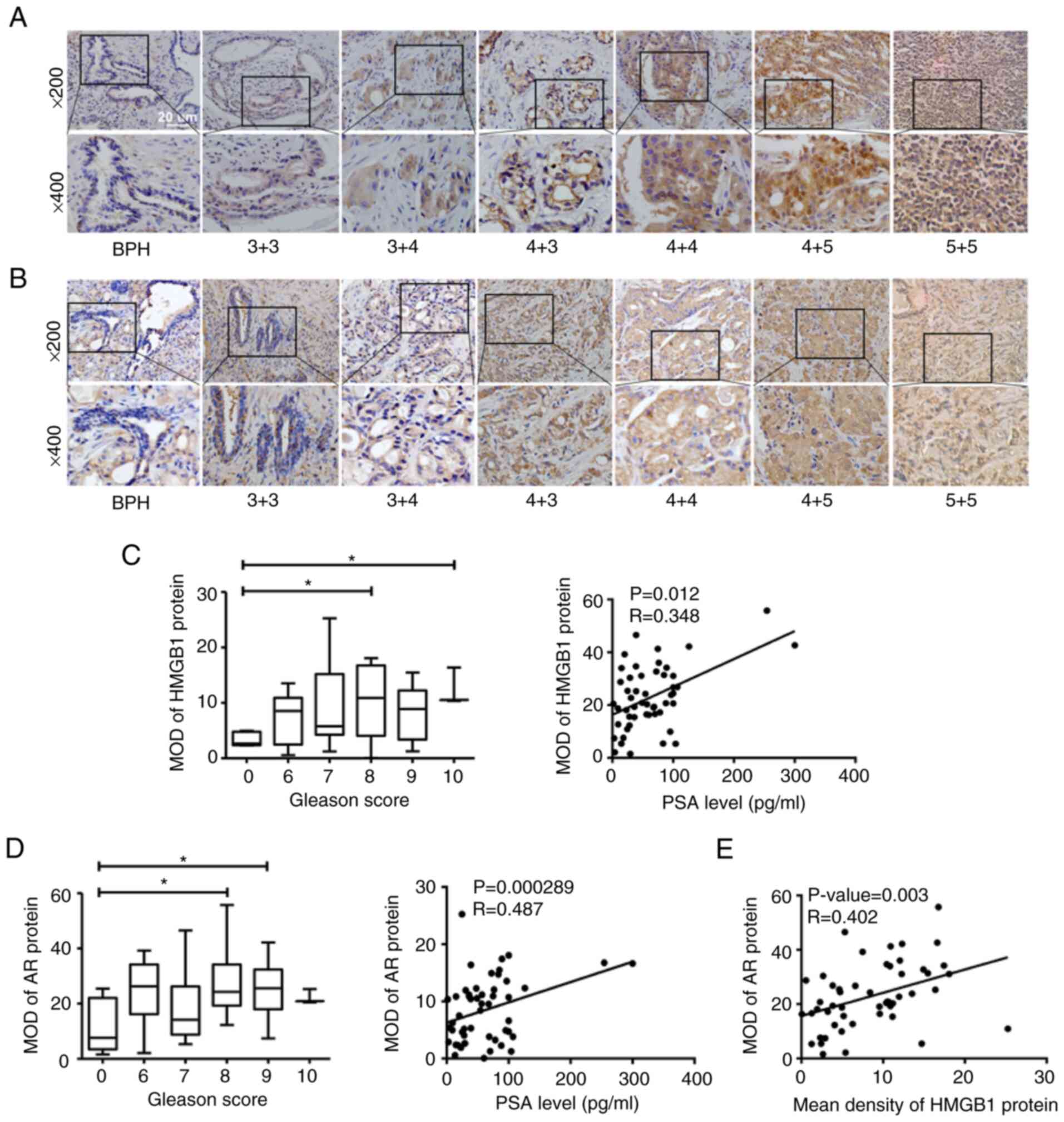
In this study, IHC staining was carried out to
detect the protein expression of HMGB1 and AR in human prostate
needle biopsy tissue specimens, including BPH and PCa. Positive
staining of HMGB1 and AR in various pathologic categories is
presented in Fig. 2A and B. By
contrast to a few disseminated cells in BPH samples, cytoplasmic
brown staining for HMGB1 and AR protein was widely observed in PCa
tissues. For PCa specimens, the protein expression levels of HMGB1
and AR gradually increased in the epithelial cytoplasm of PCa cells
according to the increasing PSA level (HMGB1: r=0.348, P=0.012; AR:
r=0.487, P=0.000289) as well as the Gleason Score (GS) (Fig. 2C and D). These results indicated
that both HMGB1 and AR are closely associated with the development
of PCa. Moreover, the data showed a significant positive
correlation between HMGB1 and AR expression in PCa tissues
(r=0.402, P=0.003; Fig. 2E). The
correlations of AR and HMGB1 expression with Gleason score, PSA and
age are also presented in Table
III.
| Table III.Correlation of AR and HMGB1
expression with Gleason score, PSA and age. |
Table III.
Correlation of AR and HMGB1
expression with Gleason score, PSA and age.
|
| Prostate tumor |
|---|
|
|
|
|---|
| Variable | Values | Gleason
scorea | PSAb | Ageb |
|---|
| MOD of AR
Protein | R | 0.259 | 0.487b | −0.018 |
|
| P-value | 0.067 | 0.000 | 0.9 |
| MOD of HMGB1
Protein | R | 0.298a | 0.348b | −0.182 |
|
| P-value | 0.033 | 0.012 | 0.202 |
HMGB1 promotes the proliferation of
PCa cells
Following transfection with HMGB1 or shHMGB1 using
the pLVX plasmid, the expression of HMGB1 protein was successfully
overexpressed or silenced in LNCaP or 22RV1 cells (Fig. 3A and B), respectively. The effect
of HMGB1 expression on LNCaP and 22RV1 cells was examined using a
CCK-8 assay under charcoal-stripped conditions. As revealed in
Fig. 3C, HMGB1 overexpression
significantly promoted the growth ability of LNCaP cells, while
silencing of HMGB1 inhibited the viability of 22RV1 cells. However,
no significant effect of HMGB1 on AR protein expression was
detected (Fig. 3A and B).
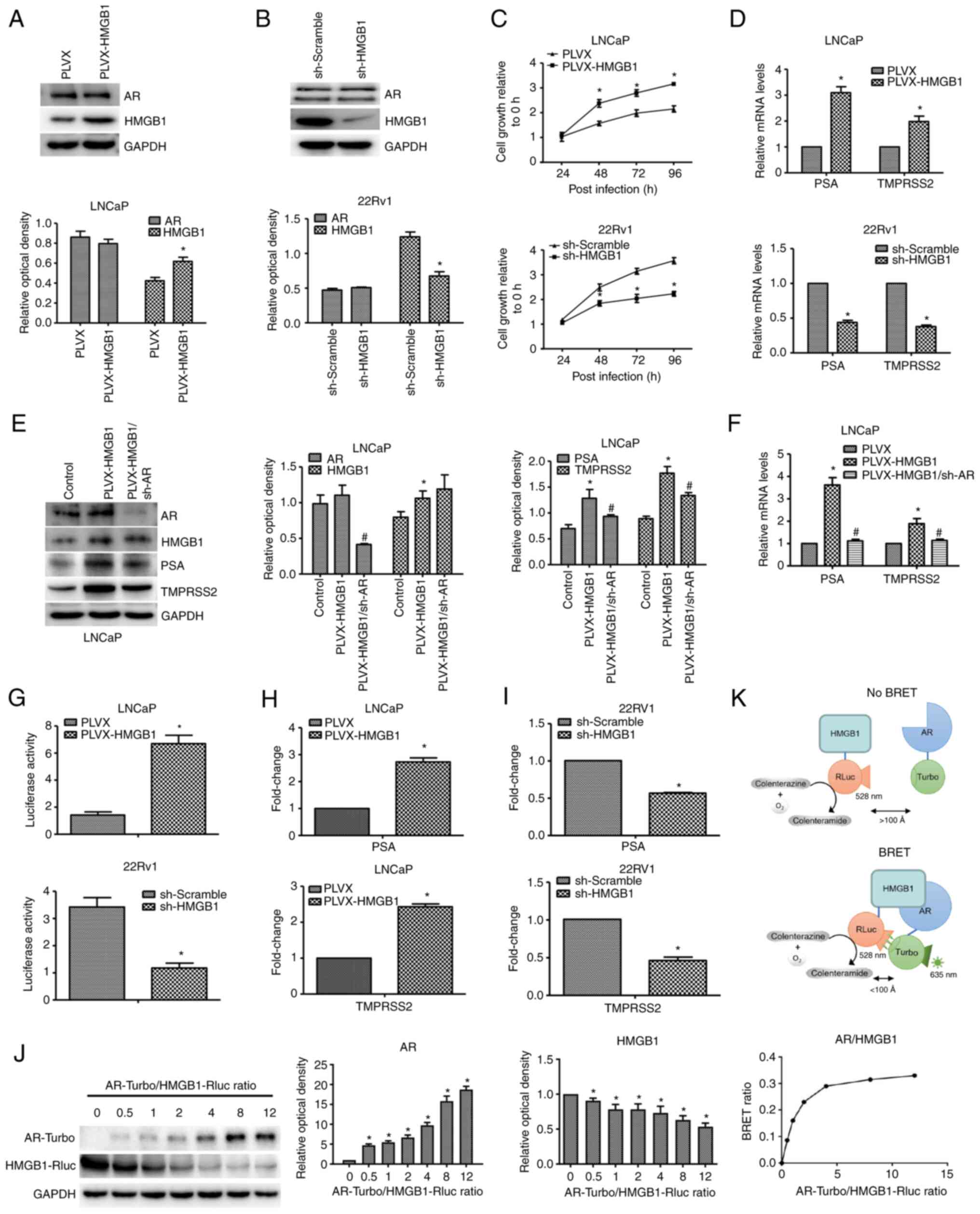 | Figure 3.HMGB1 protein activates the AR
signalling pathway by directly interacting with AR protein in PCa
in vitro. (A) Following transfection with exogenous HMGB1, the
expression levels of AR and HMGB1 in LNCaP were detected by western
blotting. *P<0.05 compared with pLVX or sh-Scramble. (B)
Following transfection with sh-HMGB1, the expression levels of AR
and HMGB1 in 22RV1 cells were detected by western blotting.
*P<0.05 compared with pLVX or sh-Scramble. (C) The growth
abilities of LNCaP-neo, LNCaP-HMGB1, 22RV1-shScramble, and
22RV1-shHMGB1 cells were detected by CCK-8 assay. *P<0.05
compared with the 0-h group. (D) The effects of HMGB1 on the
transcription levels of PSA and TMPRSS2 in LNCaP-neo, LNCaP-HMGB1,
22RV1-shScramble, and 22RV1-shHMGB1 cells were examined by RT-qPCR.
*P<0.05 compared with pLVX or sh-Scramble. (E) After treatment
with pLVX-shAR, AR, HMGB1, PSA, and TMPRSS2 protein expression
levels in LNCaP-neo, LNCaP-HMGB1 and LNCaP-HMGB1/shAR cells were
detected by western blotting. *P<0.05 compared with the control
group; #P<0.05 compared with the pLVX-HMGB1 group. (F) The
effects of HMGB1 and AR on the transcription levels of PSA and
TMPRSS2 in LNCaP-neo, LNCaP-HMGB1 and LNCaP-HMGB1/shAR cells were
examined by RT-qPCR. *P<0.05 compared with the pLVX group;
#P<0.05 compared with the pLVX-HMGB1 group. (G) Following
transfection with exogenous HMGB1 or sh-HMGB1, the transactivating
activity of AR in LNCaP-neo, LNCaP-HMGB1, 22RV1-shScramble, and
22RV1-shHMGB1 cells was determined by a reporter gene assay.
*P<0.05 compared with the pLVX group. (H and I) ChIP assay
followed by RT-qPCR was used to detect the effect of HMGB1 on the
ability of AR to bind to the promoters of PSA and TMPRSS2 in LNCaP
and 22RV1 cells. *P<0.05 compared with the pLVX group or
sh-Scramble group. (J) AR-Turbo and HMGB1-Rluc, the BRET fusion
constructs, were cotransfected into PC3 cells, and the BRET signal
was measured after the addition of the coelenterazine substrate.
Western blotting revealed the fold changes of the expression levels
of the fusion proteins. *P<0.05 compared with the
AR-Turbo/HMGB1-Rluc ratio=0 group. (K) A schematic of the principle
of the BRET assay. HMGB1, high-mobility group protein B1; AR,
androgen receptor; PCa, prostate cancer; PSA, prostate specific
antigen; TMPRSS2, transmembrane protease, serine 2; RT-qPCR,
reverse transcription-quantitative PCR; ChIP, chromatin
immunoprecipitation; BRET, bioluminescence resonance energy
transfer. |
HMGB1 promotes the
androgen-independent transactivating activity of AR
As demonstrated in Fig.
3D, overexpression of HMGB1 significantly increased the levels
of PSA and TMPRSS2 mRNA in LNCaP-HMGB1 cells under
charcoal-stripped conditions, which indicated that HMGB1 promotes
the transactivating activity of AR. Moreover, the increasing levels
of PSA and TMPRSS2 mRNA and protein were almost completely
diminished by AR knockdown (Fig. 3E
and F). Furthermore, overexpression of HMGB1 significantly
promoted the luciferase activity of ARE in LNCaP-HMGB1 cells under
charcoal-stripped conditions, while the luciferase activity of ARE
was inhibited after silencing of HMGB1 in 22RV1 cells (Fig. 3G). These results indicated that
HMGB1 promoted the androgen-independent transactivating activity of
AR by increasing the recruitment of AR to ARE-luc.
HMGB1 promotes the binding of AR to
the promoter/enhancer of AR downstream genes
To explore the underlying molecular mechanism of
HMGB1 on androgen-independent reactivation of the AR signalling
pathway, a ChIP assay was used to detect the binding levels of the
promoter/enhancer of AR downstream genes. As shown in Fig. 3H, the overexpression of HMGB1 in
LNCaP cells significantly upregulated the binding levels of the
promoter/enhancer of PSA and TMPRSS2 in the ChIP assay, indicating
that HMGB1 could strengthen the ability of AR to bind with the
promoter/enhancer of its downstream genes PSA and TMPRSS2 under
androgen-free conditions. These results were further confirmed by
the data which revealed that silencing of HMGB1 in 22RV1 cells
significantly downregulated the binding levels of the
promoter/enhancer of PSA and TMPRSS2 in the ChIP assay (Fig. 3I).
HMGB1 initiates the reactivation of
the AR signalling pathway by directly interacting with the AR
protein
To further demonstrate the way in which HMGB1
initiates the reactivation of the AR signalling pathway, a BRET
assay was performed to identify the interaction between AR and
HMGB1 proteins. The BRET saturation curve for the combination of
AR/HMGB1 fusion proteins in PC3 cells is presented in Fig. 3J. The BRET ratios increased
hyperbolically and rapidly saturated with the increase in the ratio
of energy acceptor to energy donor, indicating specific protein
interactions (10). The BRET data
revealed the ability of HMGB1 protein to interact with AR protein.
The mechanisms of the BRET assay are shown in Fig. 3K.
HMGB1 promotes the development of PCa
in vivo
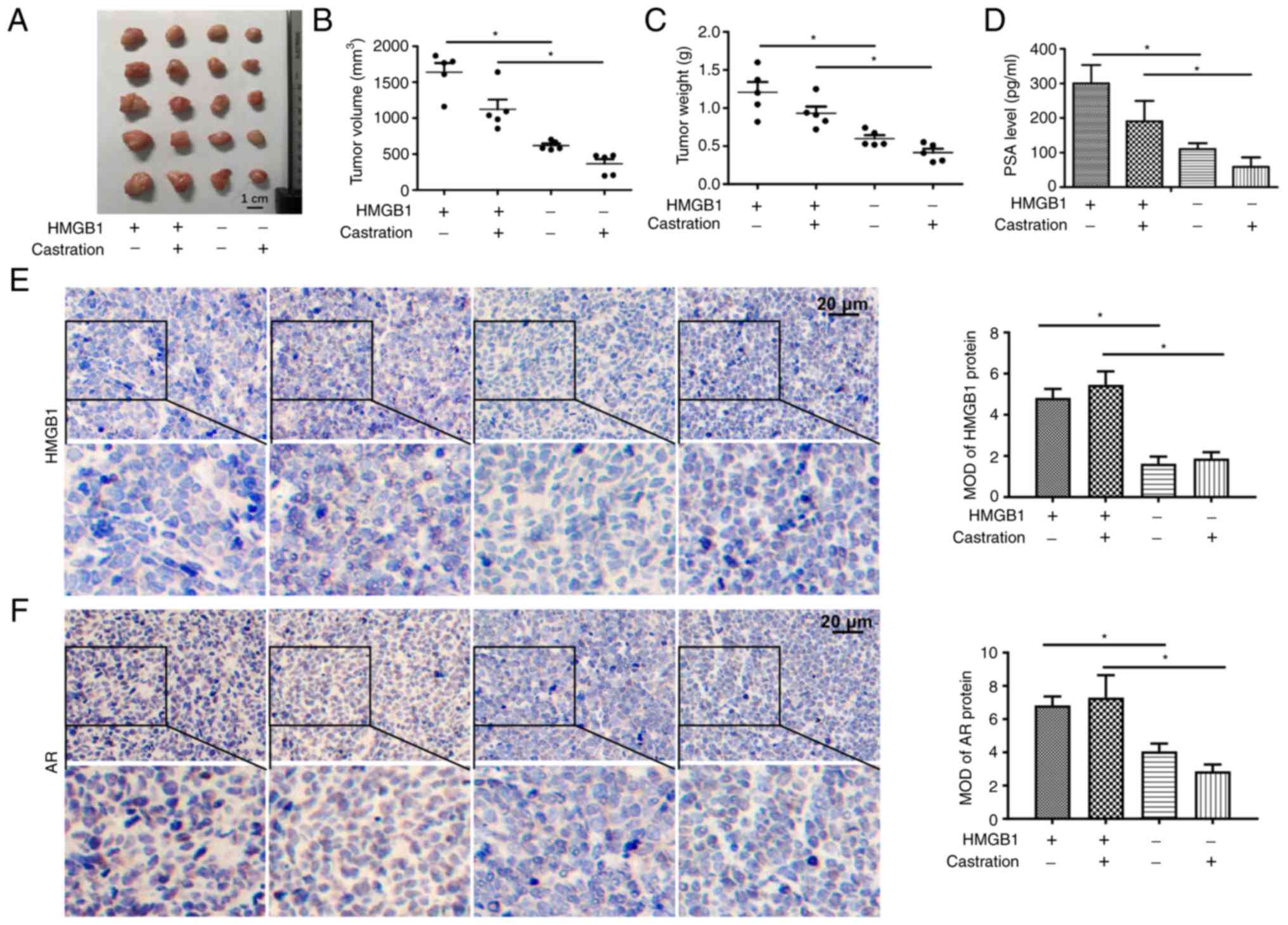
To verify whether overexpression of HMGB1 could
promote the development of PCa, a LNCaP xenograft model was
developed. The results revealed that overexpression of HMGB1
significantly increased the volume and weight of prostate tumours
(Fig. 4A-C). Furthermore, the
serum PSA level of mice exhibited the same tendency as the growth
of the tumours (Fig. 4D). The data
from IHC demonstrated that the expression of HMGB1 in tumour tissue
was significantly increased in the two groups injected with
LNCaP-HMGB1 cells (Fig. 4E). As
shown in Fig. 4F, overexpression
of HMGB1 significantly increased AR protein expression in the cell
nuclei of the tumour tissues in both the intact and castration
groups. These results suggested that HMGB1 promoted the development
of PCa in an androgen-independent manner.
Discussion
The present study is the first, to the best of our
knowledge, to reveal the association between HMGB1 and AR protein
in CRPC cells and PCa patients. Using bioinformatics analysis and
IHC, it was demonstrated that the expression levels of AR and HMGB1
were not only associated to the malignant progression of PCa but
also correlated with each other. By overexpressing HMGB1 in LNCaP
cells and silencing HMGB1 in 22RV1 cells, and performing a ChIP
assay, it was further revealed that the ability of AR protein to
bind to ARE is mediated by HMGB1 protein upon androgen deprivation.
Notably, using BRET, the interaction effect between HMGB1 protein
and AR protein, was directly identified, indicating the requirement
of HMGB1 protein for important AR protein functions in CRPC.
The finding in the present study, on the correlation
between HMGB1 expression and the malignant progression of PCa
patients is in accordance with a previous study (27). Wang et al determined that
the expression of HMGB1 protein was induced by enzalutamide in
mouse CRPC tissues (28). These
findings provide clinical and experimental evidence for the
hypothesis proposed by the authors, that HMGB1 may be a promising
target for CRPC treatment. Furthermore, it was revealed that the
expression levels of HMGB1 in PCa samples were significantly
correlated with the levels of AR. Together with the consideration
of the similarity between the structure of ER and AR (15) as well as the effect of HMGB1 on ER
signalling pathway activation (29), it was hypothesized that HMGB1
protein contributed to CRPC development by interacting with AR
protein and then reactivating the AR signalling pathway.
A previous study revealed that HMGB1 knockdown
inhibited the development of enzalutamide-induced CRPC in a mouse
model (28). This is supported by
the results of the present study showing that HMGB1 knockdown
inhibited the growth of androgen-independent 22RV1 cells and
promoted the growth of androgen-dependent LNCaP cells without the
existence of androgen. Moreover, the in vivo data showed that
overexpression of the HMGB1 protein attenuated the treatment effect
of ADT on LNCaP-bearing mice. This indicated that HMGB1 could
promote the transformation of PCa into CRPC in the presence of ADT.
Thus, it is suggested that targeting HMGB1 may be a potential
strategy for CRPC prevention. Interestingly, it was determined that
the expression of AR was not regulated by HMGB1 protein in vitro,
whereas the expression levels were significantly associated in the
tissues of patients PCa. A plausible explanation is that
correlation in expression does not signify that AR expression is
directly regulated by HMGB1, and HMGB1 may only promote AR activity
instead of expression in vitro. The in vivo results that HMGB1
overexpression promoted AR protein expression in the cell nucleus
of tumour tissues provided a further explanation. It is well known
that AR protein is predominantly located in the cytoplasm. Only
after a nuclear translocation process can the activity of AR
protein be activated with or without the presence of androgens
(7,30). From this perspective, the in vivo
data of the present study indicated that HMGB1 could activate AR
activity by promoting the nuclear translocation process of AR.
However, the detailed mechanism requires further investigation.
Furthermore, it is suggested that this may also be due to the short
regulation time of HMGB1 on AR in vitro. This short time is only
enough for HMGB1 to promote AR activity instead of expression.
However, this assumption requires more experiments for
verification. Although the conclusion that HMGB1 protein could
regulate the development of CRPC was in accordance with a previous
study, a different mechanism of the effect of HMGB1 protein on CRPC
was also identified in the present study.
Based on the data that HMGB1 protein could induce
androgen-independent increases in the mRNA expression levels of PSA
and TMPRSS2 mRNA, two AR downstream genes, in an AR-dependent
manner, it is suggested that the mechanism may be related to the
regulation of AR transactivating activity to its downstream genes.
Interestingly, the results of the reporter gene assay revealed that
HMGB1 could enhance AR transactivating activity without the
existence of androgen, which confirmed the aforementioned
hypothesis. Therefore, the potential mechanism of these results is
continually being explored. To date, studies of AR transactivating
activity in CRPC have focused mostly on its ability to bind to ARE.
Selenium suppresses the binding of AR to the ARE and then inhibits
the AR signalling pathway (31).
Collak et al determined that Yes associated protein 1 (YAP1)
and AR could form a complex and ultimately bind to the ARE of PSA
(32). Interestingly, it was
determined that HMGB1 promoted the ability of AR binding to the
AREs of PSA and TMPRSS2 genes in a ChIP assay, which directly
identified the promoting effect of HMGB1 protein on AR protein
activity.
AR activity is affected not only by ligand binding
but also by homodimerization and by interactions with cofactors
(7). Over 170 cofactors of AR have
been found to bind to AR and influence the transactivating activity
of AR at low levels of androgen (6). The data of BRET performed in the
present study, indicated that HMGB1 protein could be a cofactor and
directly interact with AR protein through the formation of an
HMGB1/AR complex in an androgen-independent manner in CRPC. This,
together with the finding that the binding of AR protein to ARE was
promoted by HMGB1 protein, demonstrated that the HMGB1/AR complex
could contribute to the activation of the AR signalling pathway,
which may be the key mechanism by which the HMGB1 protein promotes
the malignant progression of PCa. Thus, disrupting AR/HMGB1
interactions may be an effective strategy for CRPC prevention as
well as treatment.
In conclusion, the present study revealed the
effects of HMGB1 on the development of CRPC in vitro and in vivo
and described its mechanisms. The results demonstrated that HMGB1
promoted the development of CRPC by interacting with AR. Targeting
HMGB1 may be a novel and effective treatment strategy for CPRC
prevention and treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China Project (grant no. 81602228) and Jilin Province
Development and Reform Commission (grant no. 2019C050-5).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DX, JC, and LZ conceived and designed the
experiments. JC and TW performed the experiments including data
collection and data analysis. ZY and KH analyzed the data. JC and
TW wrote the original manuscript. ZY and YY contributed to revising
the manuscript for intellectual content and language editing. LZ
and KH supervised the study and were involved in project
management. LZ and JC confirm the authenticity of the raw data. All
authors read and approved the final manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the School of Nursing, Jilin University (Changchun,
China), and the animal experiments were approved by the Animal
Ethics Committee of Basic Medical College of Jilin University
(Changchun, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
financial interests or personal relationships that could have
influenced the research reported in the present study.
Glossary
Abbreviations
Abbreviations:
|
CRPC
|
castration-resistant prostate
cancer
|
|
AR
|
androgen receptor
|
|
AREs
|
androgen response elements
|
|
HMGB1
|
high-mobility group protein B1
|
|
PCa
|
prostate cancer
|
|
ADT
|
androgen deprivation therapy
|
|
PSA
|
prostate specific antigen
|
|
ER
|
estrogen receptor
|
|
ERE
|
estrogen response element
|
|
IHC
|
immunohistochemical
|
|
MOD
|
mean optical density
|
|
BRET
|
bioluminescence resonance energy
transfer
|
|
ChIP
|
Chromatin immunoprecipitation
|
|
BPH
|
benign prostate hyperplasia
|
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Glina S, Rivero MA, Morales A and
Morgentaler A: Studies on prostatic cancer I. The effect of
castration, of estrogen and of androgen injection on serum
phosphatases in metastatic carcinoma of the prostate by Charles
Huggins and Clarence V. Hodges. J Sex Med. 7:640–644. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guan W, Li F, Zhao Z, Zhang Z, Hu J and
Zhang Y: Tumor-associated macrophage promotes the survival of
cancer cells upon docetaxel chemotherapy via the
CSF1/CSF1R-CXCL12/CXCR4 axis in castration-resistant prostate
cancer. Genes (Basel). 12:7732021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kluetz PG, Ning YM, Maher VE, Zhang L,
Tang S, Ghosh D, Aziz R, Palmby T, Pfuma E, Zirkelbach JF, et al:
Abiraterone acetate in combination with prednisone for the
treatment of patients with metastatic castration-resistant prostate
cancer: U.S. Food and drug administration drug approval summary.
Clin Cancer Res. 19:6650–6656. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Scher HI, Fizazi K, Saad F, Taplin ME,
Sternberg CN, Miller K, de Wit R, Mulders P, Chi KN, Shore ND, et
al: Increased survival with enzalutamide in prostate cancer after
chemotherapy. N Engl J Med. 367:1187–1197. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Egan A, Dong Y, Zhang H, Qi Y, Balk SP and
Sartor O: Castration-resistant prostate cancer: Adaptive responses
in the androgen axis. Cancer Treat Rev. 40:426–433. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
van Royen ME, van Cappellen WA, de Vos C,
Houtsmuller AB and Trapman J: Stepwise androgen receptor
dimerization. J Cell Sci. 125:1970–1979. 2012.PubMed/NCBI
|
|
8
|
Tran C, Ouk S, Clegg NJ, Chen Y, Watson
PA, Arora V, Wongvipat J, Smith-Jones PM, Yoo D, Kwon A, et al:
Development of a second-generation antiandrogen for treatment of
advanced prostate cancer. Science. 324:787–790. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ryan CJ, Smith MR, Fizazi K, Saad F,
Mulders PF, Sternberg CN, Miller K, Logothetis CJ, Shore ND, Small
EJ, et al: Abiraterone acetate plus prednisone versus placebo plus
prednisone in chemotherapy-naive men with metastatic
castration-resistant prostate cancer (COU-AA-302): Final overall
survival analysis of a randomised, double-blind, placebo-controlled
phase 3 study. Lancet Oncol. 16:152–160. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu D, Zhan Y, Qi Y, Cao B, Bai S, Xu W,
Gambhir SS, Lee P, Sartor O, Flemington EK, et al: Androgen
receptor splice variants dimerize to transactivate target genes.
Cancer Res. 75:3663–3671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xue J, Suarez JS, Minaai M, Li S, Gaudino
G, Pass HI, Carbone M and Yang H: HMGB1 as a therapeutic target in
disease. J Cell Physiol. 236:3406–3419. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jung AR, Kim GE, Kim MY, Ha US, Hong SH,
Lee JY, Kim SW and Park YH: HMGB1 promotes tumor progression and
invasion through HMGB1/TNFR1/NF-κB axis in castration-resistant
prostate cancer. Am J Cancer Res. 11:2215–2227. 2021.PubMed/NCBI
|
|
13
|
Chou YE, Yang PJ, Lin CY, Chen YY, Chiang
WL, Lin PX, Huang ZY, Huang M, Ho YC and Yang SF: The impact of
HMGB1 polymorphisms on prostate cancer progression and
clinicopathological characteristics. Int J Environ Res Public
Health. 17:72472020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Das D, Peterson RC and Scovell WM: High
mobility group B proteins facilitate strong estrogen receptor
binding to classical and half-site estrogen response elements and
relax binding selectivity. Mol Endocrinol. 18:2616–2632. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Powell E and Xu W: Intermolecular
interactions identify ligand-selective activity of estrogen
receptor alpha/beta dimers. Proc Natl Acad Sci USA.
105:19012–19017. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bjerregaard-Olesen C, Ghisari M, Kjeldsen
LS, Wielsøe M and Bonefeld-Jørgensen EC: Estrone sulfate and
dehydroepiandrosterone sulfate: Transactivation of the estrogen and
androgen receptor. Steroids. 105:50–58. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mafuvadze B, Liang YY and Hyder SM:
Cholesterol synthesis inhibitor RO 48-8071 suppresses
transcriptional activity of human estrogen and androgen receptor.
Oncol Rep. 32:1727–1733. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tomlins SA, Mehra R, Rhodes DR, Cao X,
Wang L, Dhanasekaran SM, Kalyana-Sundaram S, Wei JT, Rubin MA,
Pienta KJ, et al: Integrative molecular concept modeling of
prostate cancer progression. Nat Genet. 39:41–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Singh D, Febbo PG, Ross K, Jackson DG,
Manola J, Ladd C, Tamayo P, Renshaw AA, D'Amico AV, Richie JP, et
al: Gene expression correlates of clinical prostate cancer
behavior. Cancer Cell. 1:203–209. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu YP, Landsittel D, Jing L, Nelson J, Ren
B, Liu L, McDonald C, Thomas R, Dhir R, Finkelstein S, et al: Gene
expression alterations in prostate cancer predicting tumor
aggression and preceding development of malignancy. J Clin Oncol.
22:2790–2799. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vanaja DK, Cheville JC, Iturria SJ and
Young CY: Transcriptional silencing of zinc finger protein 185
identified by expression profiling is associated with prostate
cancer progression. Cancer Res. 63:3877–3882. 2003.PubMed/NCBI
|
|
22
|
Tian Y, Zhao L, Zhang H, Liu X, Zhao L,
Zhao X, Li Y and Li J: AKR1C3 overexpression may serve as a
promising biomarker for prostate cancer progression. Diagn Pathol.
9:422014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J, Xia Y, Peng Y, Wu S, Liu W, Zhang
H, Wang T, Yang Z, Zhao S and Zhao L: Analysis of the association
between KIN17 expression and the clinical features/prognosis of
epithelial ovarian cancer, and the effects of KIN17 in SKOV3 cells.
Oncol Lett. 21:4752021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang Y, Liu T, Hu C, Xia H, Liu W, Chen J,
Wu S, Jiang Y, Xu Y, Liu W and Zhao L: Ferroptosis inducer erastin
downregulates androgen receptor and its splice variants in
castration-resistant prostate cancer. Oncol Rep. 45:252021.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen J, Zhao S, Tan W, Wang T, Wu S, Wang
C, Jiang Y, Zhou T, Zhang Z and Zhao L: Attenuated Salmonella
carrying plasmid co-expressing HPV16 L1 and siRNA-E6 for cervical
cancer therapy. Sci Rep. 11:200832021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li T, Gui Y, Yuan T, Liao G, Bian C, Jiang
Q, Huang S, Liu B and Wu D: Overexpression of high mobility group
box 1 with poor prognosis in patients after radical prostatectomy.
BJU Int. 110:E1125–E1130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang C, Peng G, Huang H, Liu F, Kong DP,
Dong KQ, Dai LH, Zhou Z, Wang KJ, Yang J, et al: Blocking the
feedback loop between neuroendocrine differentiation and
macrophages improves the therapeutic effects of enzalutamide
(MDV3100) on prostate cancer. Clin Cancer Res. 24:708–723. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ogawa S, Inoue S, Watanabe T, Hiroi H,
Orimo A, Hosoi T, Ouchi Y and Muramatsu M: The complete primary
structure of human estrogen receptor beta (hER beta) and its
heterodimerization with ER alpha in vivo and in vitro. Biochem
Biophys Res Commun. 243:122–126. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cao B, Qi Y, Zhang G, Xu D, Zhan Y,
Alvarez X, Guo Z, Fu X, Plymate SR, Sartor O, et al: Androgen
receptor splice variants activating the full-length receptor in
mediating resistance to androgen-directed therapy. Oncotarget.
5:1646–1656. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dong Y, Lee SO, Zhang H, Marshall J, Gao
AC and Ip C: Prostate specific antigen expression is down-regulated
by selenium through disruption of androgen receptor signaling.
Cancer Res. 64:19–22. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Collak FK, Demir U and Sagir F: YAP1 is
involved in tumorigenic properties of prostate cancer cells. Pathol
Oncol Res. 26:867–876. 2020. View Article : Google Scholar : PubMed/NCBI
|