Introduction
Before precursor mRNA (pre-mRNA) becomes a protein,
it needs to be transformed into a mature mRNA by intron splicing
and exon splicing. This splicing process can be achieved by major
and secondary spliceosomes (1,2). The
major spliceosome comprises five small nuclear ribonucleoprotein
complexes (snRNPs) known as U1, U2, U4, U5 and U6. Of these, U2AF1
is the U2 small nuclear RNA auxiliary factor 1 protein coding
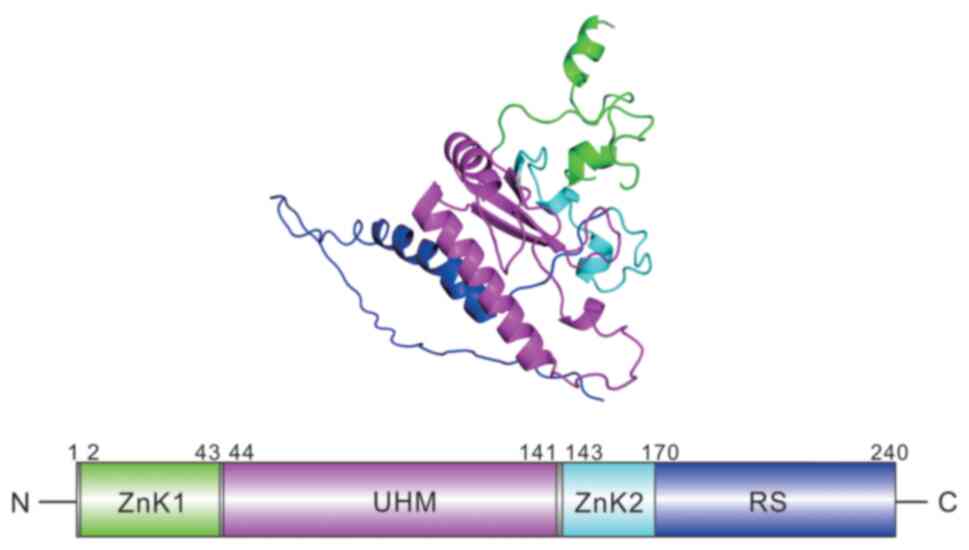
various spliceosome genes. This spliceosome is composed of 240
amino acids with a molecular weight of 35 kDa and is located on
chromosome 21q22.3 (3,4). The RNA binding region and C-terminal
serine/arginine-rich (RS) domain are composed of the U2AF1
structure, which binds to the AG dinucleotide in 3′ splicing sites
(3′ SS) at the boundary introns and exons. The 1 crystal structure
of U2AF1 includes zinc knuckles (ZnK1, ZnK2) and a central U2AF
homology motif (UHM). It is an integral part of both constitutive
and enhancer-subordinate RNA joining by straightforwardly
intervening in collaborations between the enormous subunit and the
proteins bound to the enhancers (Fig.
1) (5–8).
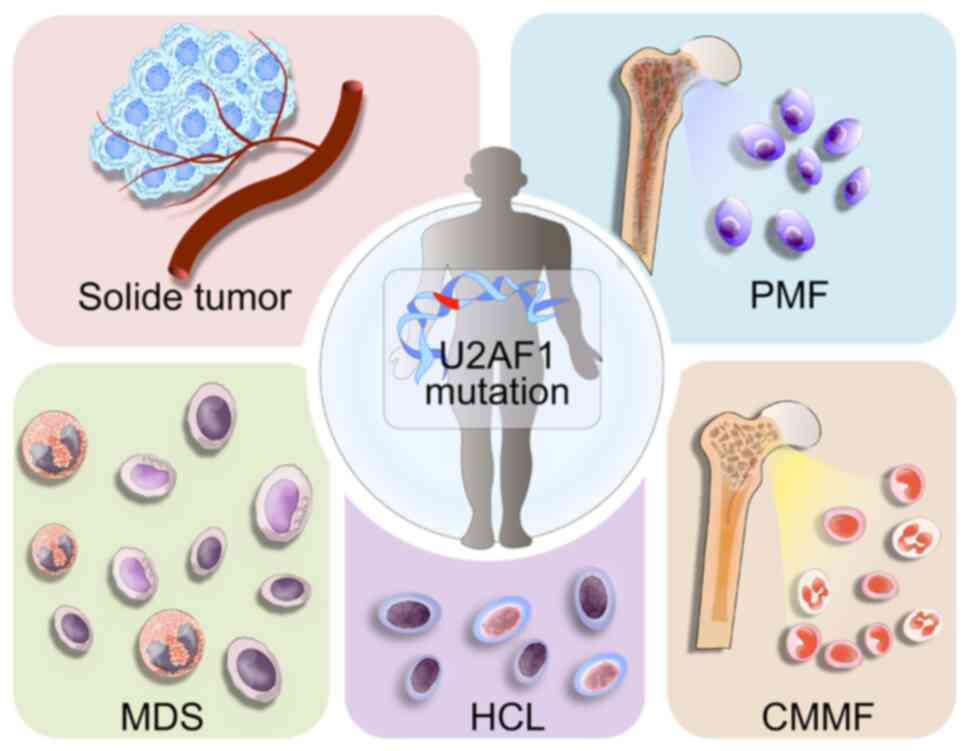
Mutations in U2AF1 have been found to be associated
with a wide range of diseases, most notably haematological
malignancies such as myelodysplastic syndromes (MDS) and acute
myeloid leukaemia (AML) as well as various solid tumours (9–11).
However, although U2AF1 mutations are commonly observed in the
former haematological malignancies, these mutations appear with a
low frequency in solid tumours (Fig.
2) (12–14). Additionally, mutations in other
splicing-associated genes, including SF3B1, SRSF2 and ZRSR2, have
been associated with haematological malignancies, albeit with
distinct and mutually exclusive tendencies across highly conserved
domains (15–19).
Given recent developments in sequencing technology,
an understanding of the molecular pathogenesis of U2AF1-associated
diseases has become increasingly important. Hence, the structure
and pathogenesis of U2AF1 mutations in different diseases were
outlined in the present review and potential therapeutic strategies
were presented.
U2AF1 mutations in malignant haematological
diseases
In MDS
MDS is a heterogeneous group of clonal
haematopoietic stem cell (HSC) disorders characterized primarily by
deficiencies in the myeloid, erythroid, and/or megakaryocytic
lineages, mono- or multilineage dysplasia, clonal dominance of
abnormal immature cells, and an increased risk of secondary AML.
The French, American and British divisions of AML subtypes are
categorized as M0 through M7 based on the type of cell the
leukaemia develops from and how mature the cells are. The World
Health Organization system classification is AML with certain
genetic abnormalities. The phenomenon of AML emerging from MDS has
prompted numerous researchers to suggest that the two disorders are
related, with similar pathogenic mechanisms and treatments
(20–24).
Heterozygous somatic mutations in splicing factor
genes (SF3B1, SRSF2, U2AF1 and ZRSR2) occur in >50% of all
patients with MDS (25–29), with U2AF1 mutations ranging from
7–11% and having the highest incidence in AML and the worst
prognosis among the four mutations (30,31).
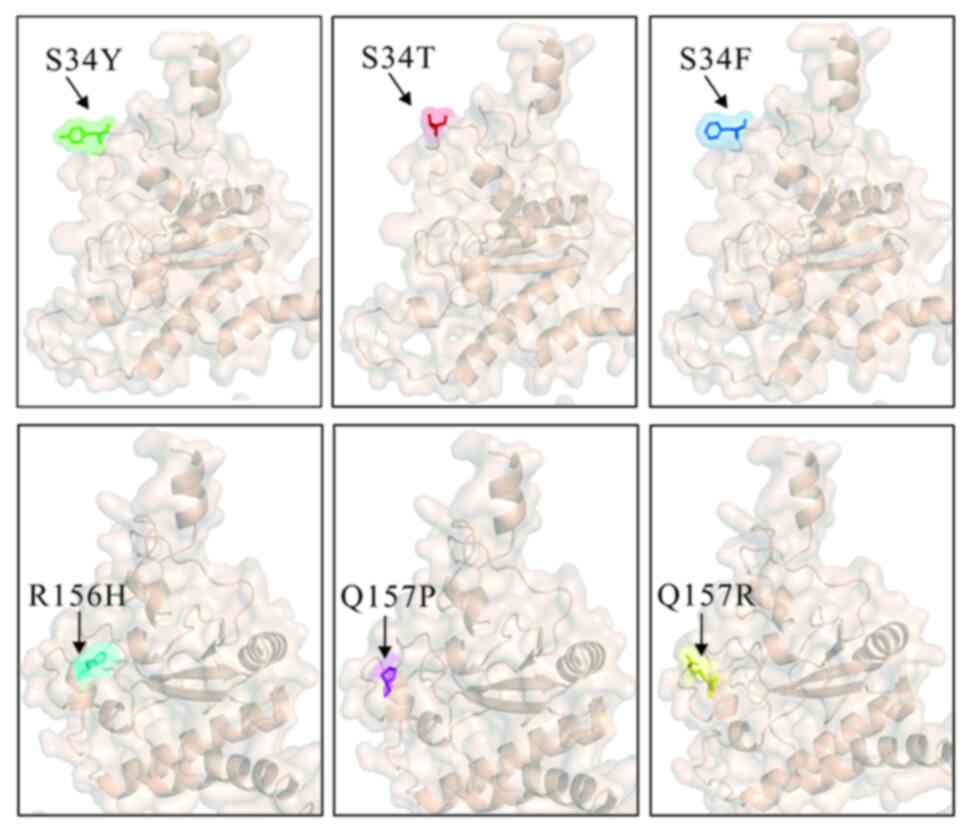
Structural changes caused by mutations in U2AF1 occur in two
hotspots-S34 and Q157-in the first and second zinc finger domains,
as illustrated in Fig. 3. These
mutations are capable of inducing gain-of-function changes, which
modify the 3′ SS preference in a sequence-specific manner,
determined by the nucleotides surrounding the AG dinucleotide
responsible for the 3′ SS core consensus motif (10,15,32–34).
This suggests that U2AF1 mutations directly affect RNA binding and
consequently influence exon inclusion. It has also been observed
that the effects of different mutations may vary on different
exons; for example, the U2AF1 S34 mutation prefers C or A more
frequently in the-3 position when analysed in AML patients, whereas
the Q157 mutation, involving the first zinc finger, shows
preferential recognition of G over A in the +1 position. By
combining comprehensive data from RNA sequencing and U2AF1
mutations with 3′ SS recognition ability, a clearer understanding
of the mechanism by which U2AF1 mutations affect splicing in
patients with MDS can be established (10,12,35,36).
Dysregulation of cell growth and immunity is a
hallmark of MDS (37). In
accordance with several research studies, aberrantly spliced genes
identified in MDS with U2AF1 mutations converge on disrupted
cellular differentiation processes, including RNA splicing, RNA
localization/transport, RNA binding, protein translation processes,
ribosomal pathways, and mitochondrial dysfunction (31,38–40).
Subsequently, an analysis of upstream transcriptional regulators
using the Ingenuity Pathway Analysis (IPA) tool suggested the
significant enrichment of genes regulated through transcriptional
regulators (RICTOR, E2F1, HNF4A, MYC, MYCN and RB1, the major
controllers of cell growth/cell cycle) in the aberrantly spliced
genes (38,41,42)
(Table I). Additionally, these
studies revealed that MDS with U2AF1 mutations has ten significant
transcriptional pathways, with the sirtuin signalling pathway
leading the rankings (38,43). Sirtuins, including sirtuin1,
sirtuin2 and other isoforms, are a class of proteins that regulate
diverse physiological processes and are considered to be partially
involved in MDS and other haematological malignancies (44–46).
Furthermore, mitochondrial dysfunction has generally been
associated with MDS pathophysiology. In one study, mice with
mitochondrial dysfunction exhibited characteristics of MDS,
including macrocytic anaemia (47).
Mitochondrial dysfunction can be induced by mitochondrial DNA
mutations, electron respiratory chain leakages, increased oxidative
stress, dysfunctional TCA cycle enzymes and abnormal tumour
suppressive signalling pathways (48). Park et al (49) reported that cells expressing the
U2AF1 S34F gene had defective autophagy along with mitochondrial
dysfunction and increased production of reactive oxygen species,
causing genomic instability and an elevated frequency of
spontaneous mutations. These sirtuins in the mitochondria, which
additionally function as histone deacetylases, are involved in
metabolism, inflammation, genome stability and cell proliferation
control. In addition, their role in ageing, cancer and survival has
been demonstrated in some studies (50,51).
 | Table I.The different mutation of U2 small
nuclear RNA auxiliary factor 1 in malignant diseases. |
Table I.
The different mutation of U2 small
nuclear RNA auxiliary factor 1 in malignant diseases.
| Malignant
diseases | Mutation site | Relevant gene | (Refs.) |
|---|
| MDS | S34, Q157 | RICTOR, E2F1,
HNF4A, MYC, | (10,12,15,31–60) |
|
|
| MYCN, RB1, ROS,
ERCC8, FANCM |
|
| PMF | S34, Q157 | JAK2, CALR,
MPL | (65–71) |
| CMML | S34F, Q157P/R, | ASXL1, TET2, CBL,
RAS, EZH2, TP53, | (74–76) |
|
| R156H | IDH1/2DNMT3A,
RUNX1, UTX |
|
| HCL | S34F, R156H | CD29, CD20, CD22,
CD25, TRAP, | (78,79) |
|
|
| Anx-A1, BRAF |
|
| Lung cancer | S34F | SLC34A2-ROS1 | (80) |
| Prostate
cancer | S34F | ARV7, MAPK,
EZH2 | (82,83) |
| Borderline ovarian
tumors | S34T | TP53 | (84) |
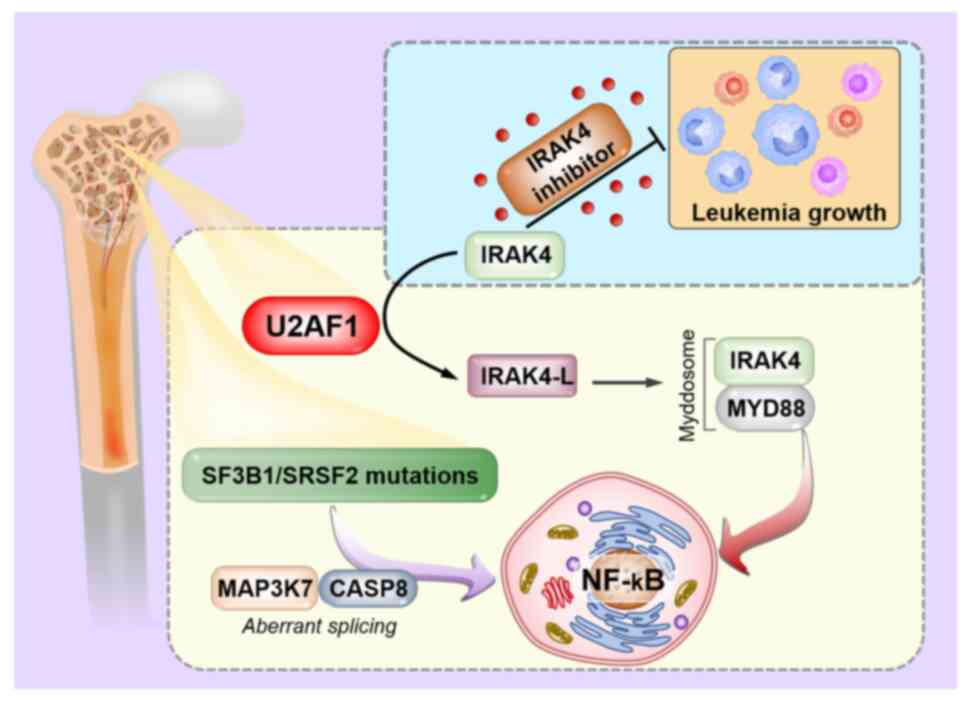
The NF-κB signalling pathway has a significant role
in determining the quiescent or active state of HSCs, and
aberrantly active NF-κB signalling may promote malignant HSPCs
(52). Balka et al (53) identified that U2AF1 mutations
resulted in aberrant splicing of IRAK4, a serine/threonine kinase
involved in toll-like receptor downstream signalling, giving rise
to a longer isoform named IRAK4-L, which preferentially assembles
signal transducers (for example, MyD88 and IRAK kinases) into a
myddosome, causing a higher degree of NF-κB activation. Moreover,
IRAK4-L participated in innate immunity and was found to be
associated with a poorer prognosis. Subsequent studies also
revealed that mutations in SF3B1 and SRSF2 had a similar effect,
leading to hyperactive NF-κB signals via aberrant splicing of
MAP3K7 and CASP8, respectively (19,54–56)
(Fig. 4). Overall, the
aforementioned evidence suggested that mutations in various
splicing factors alter different targets at the pre-mRNA splicing
level but share a common downstream signalling node leading to
heightened innate immune signalling.
The dysregulation of splicing factor genes is
associated with the formation of R loops, a pathogenic aberrant
RNA-DNA hybrid structure derived from nascent invasion. Crossley
et al (57) showed that the
expression of U2AF1 S34F mutations in human cancer cells led to
R-loop accumulation. Furthermore, treatment of U2AF1
S34F-expressing cells with ATR inhibitors has been identified to
induce DNA damage and cell death, which can be enhanced by
spliceosome inhibitors (58,59).
Chen et al (60) and
Pellagatti et al (38)
reported ERCC8 and FANCM mutations in MDS patients with U2AF1
mutations that may be involved in the suppression/regulation of
R-loop formation and the DNA damage response. Chen et al
(61) found that impaired
transcription inhibits release at transcription start sites (TSSs),
likely due to the inability to extract P-TEFb, a kinase that
phosphorylates the RNA polymerase II (Pol II) C-terminal domain,
from the inhibitory 7SK complex to transcriptionally engaged Pol II
at TSSs in SRSF2-mutant MDS, resulting in enhanced R loops.
Interestingly, Pellagatti et al (38) and Boultwood et al (62) identified aberrant splicing of
several genes, including SETX and ATR, involved in the
suppression/regulation of R-loop formation in bone marrow
CD34+ cells of MDS patients with splicing factor
mutations. Collectively, these data raise the possibility that
R-loop-induced DNA damage may lead to deleterious mutations that
promote clonal advantage in MDS pathogenesis.
In primary myelofibrosis (PMF)
PMF is classified as a chronic myeloproliferative
neoplasm characterized by bone marrow tissue fibrosis, enlargement
of the spleen, and the occurrence of anaemia associated with the
presence of nucleated and teardrop-shaped red blood cells. In PMF,
the osseous components of bone marrow undergo a process known as
osteosclerosis. Furthermore, fibroblasts secrete collagen and
reticulin proteins, collectively referred to as fibrosis. Both of
these pathological processes disrupt the usual functioning of bone
marrow, leading to reduced production of various blood cells,
including erythrocytes (red cells), granulocytes and
megakaryocytes; the latter cells are responsible for platelet
production. Thus, the combined effects of osteosclerosis and
fibrosis in PMF significantly impair the normal haematopoietic
activities of bone marrow (63–65).
Most cases of PMF are associated with mutations in
the JAK2, CALR or MPL genes (Table
I) (66). According to
sequencing results, >15–20% of patients with PMF suffer from
U2AF1 mutations, of which the two major mutation sites are Q157 and
S34. These findings contrast with MDS, in which the incidence of
S34 and Q157 mutations is low. Interestingly, the age distribution
of patients with U2AF1 mutations is also different in MDS and PMF,
with MDS patients tending to be younger and PMF patients tending to
be older (67). In terms of
clinical features, U2AF1 mutations are associated with anaemia,
thrombocytopenia and poor disease prognosis. In MDS, U2AF1S34
mutations are more specifically associated with thrombocytopenia,
while anaemia is more specifically associated with Q157 mutations
(Fig. 3). Both types of mutations
in PMF may cause anaemia, and Q157 mutations are more strongly
associated with thrombocytopenia (68–70).
In chronic myelomonocytic leukaemia
(CMML)
CMML is a haematopoietic disorder that involves
clonal proliferation of cells, presenting with diverse
myeloproliferative and myelodysplastic characteristics. This
complexity contributes to the heterogeneous nature of CMML. The
tyrosine kinase inhibitor imatinib remains the primary targeted
drug for the treatment of CMML, thus rendering it inevitable that
resistance to this inhibitor will decrease its efficacy (71,72).
An analysis of a large cohort of patients with CMML
revealed that the occurrence rate of U2AF1 mutations is ~9%.
Patnaik et al (73) reported
that the primary sites of U2AF1 mutations are S34F, Q157P/R and
R156H. Among patients with U2AF1 mutations, >50% carried the
S34F mutation, while a subset of patients exhibited the Q157P and
Q157R mutations, and only a minority had the R158H mutation
(Fig. 3). Additionally, it has been
demonstrated that other gene mutations, such as ASXL1, TET2, CBL,
RAS, EZH2, TP53, IDH1/2, DNMT3A, RUNX1 and UTX, interfere with
spliceosome genes and induce distinct amino acid missense mutations
in U2AF1 (73–75). These insights suggest that targeting
U2AF1 through drug design may hold promise for improving survival
rates among patients with U2AF1 mutations.
In hairy cell leukaemia (HCL)
HCL is a rare B-cell malignancy characterized by
pancytopenia and splenomegaly, with an incidence of ~2% in adults.
It is generally recognized as a subtype of chronic lymphocytic
leukaemia and can be divided into two categories: Variant HCL
(vHCL) and classical HCL (cHCL). vHCL is more prevalent, accounting
for nearly 10% of HCL patients, whereas cHCL is rare among mature
B-cell malignancies (76).
The presence of various markers, such as CD29, CD20,
CD22, CD25, tartrate-resistant acid phosphatase (TRAP), annexin 1A
(Anx-A1) and BRAF V600E mutations has been documented in the immune
response of leukaemia cells and summarized in Table I. In the case of vHCL, several
similarities with cHCL are notable, yet vHCL lacks expression of
CD25, CD123, ANXA1, TRAP and BRAF V600E. Additionally, vHCL has
been found to be more aggressive and less responsive to purine
analogues.
vHCL exhibits several similarities to HCL. However,
vHCL lacks the expression of CD25, CD123, ANXA1, TRAP and BRAF
V600E. Furthermore, vHCL has been observed to be more aggressive
and less purine analogue-sensitive (77). By detecting vHCL patients, Durham
et al (78) found that the
spliceosome U2AF1 was mutated, leading to its loss of function, and
the major mutation sites were S34F and R156H, as demonstrated in
Fig. 3. This is in contrast to
cHCL, which only included BRAF V600E mutations. Thus, there is an
important genetic difference between cHCL and vHCL. The
aforementioned research provides evidence for potential novel
therapeutic targets for vHCL, which may have implications for the
future personalized treatment of the disease.
U2AF1 mutation in different solid
tumours
In lung cancer (LC)
As revealed in separate studies, there are recurrent
U2AF1 mutations in LC, especially in lung adenocarcinoma (LUAD).
U2AF1 mutations have been observed in ~3% of patients with LUAD,
and the incidence of U2AF1 mutations in other LC subtypes appears
to be lower, including squamous cell carcinoma and small cell
carcinoma. In contrast to the aforementioned neoplastic disorders,
the S34F mutation is the most pervasive hotspot in LC targeting the
first zinc finger domain. Additionally, this mutation has been
linked to altered splicing of genes such as GNAS, H2AFY, STRAP and
PICALM in both LUAD and non-small cell LC patients. Based on an
analysis of A549 cells, U2AF1 S34F mutations can lead to aberrantly
spliced DNA damage repair factors (32,79).
In terms of pathogenesis, Esfahani et al
(32) reported that U2AF1 S34F
preferentially binds to and regulates splicing of introns
containing CAG trinucleotides at their 3′ splice junctions compared
with its wild-type counterpart. Moreover, overexpression of U2AF1
S34F upregulates the expression of genes associated with
epithelial-mesenchymal transition, thereby inducing cell invasion,
which is mediated by preferential splicing of SLC34A2-ROS1 long
isoforms (Table I) (32,80).
These studies have demonstrated that in lung tumours harbouring
U2AF1 S34F, a wild-type allele of the gene is invariably retained
and is critically dependent on cell survival. However, a previous
in vitro study observed that in cells harbouring multiple
U2AF1 alleles, a mutation in one allele stimulates the emergence of
the U2AF1 S34F mutation, leading to the development of U2AF1
‘wild-type’ cells. Consequently, these cells exhaust their
wild-type alleles and give rise to isogenic U2AF1 mutant cells
(80). This means that mutations in
U2AF1 appear to be an important contributing factor in the
formation of LC. Targeted therapies directed at U2AF1 mutations
could represent a promising clinical option in the treatment of
LC.
In other solid tumours
Prostate cancer is the second most commonly
occurring cancer in men. A recent study identified U2AF1 mutations
in prostate tumours, suggesting that this may be a putative
biosignature for this disease (81). According to a recent study,
downregulation of U2AF1 has been linked to poor prognosis and a
correlation with androgen receptor variant 7 (ARV7). This
downregulation can stimulate the proliferation of prostate cancer
cells and lead to bicalutamide resistance by regulating intron
splicing of ARV7. MAPK pathways have also been identified as having
an administrative impact on U2AF1 and its regulation of ARV7
splicing (Table I). In summary,
downregulation of U2AF1 can cause androgen resistance and increased
proliferation of prostate cancer cells through the regulation of
ARV7 splicing and an increase in MAPK1 expression, predicting a
poor prognosis in patients with this cancer (82). A further study based on prostate
tumour biopsies suggested, however, the overexpression of U2AF1
genes in association with EZH2 localized in mRNA and MAPK pathways
(83).
Borderline ovarian tumours account for ~15–20% of
all ovarian malignancies and arise from the tissue that
encapsulates the ovary. However, in clinical practice, it is
difficult to detect borderline mucinous tumours until advanced
stages. In a case study of one patient with a borderline mucinous
tumour, a novel mutation of U2AF1 was observed, with an alteration
in amino acid serine in place of tyrosine, although such mutations
are uncommon in ovarian tumours (occurring in <0.2% of cases)
(84). On the other hand, TP53
mutations remain highly common in ovarian cancers. Moreover, U2AF1
and TP53 mutations can predict poor prognosis and a lack of
remission. Consequently, inhibitors targeting U2AF1 are essential
for these patients.
Therapies targeting U2AF1
Small-molecule modulator: NSC
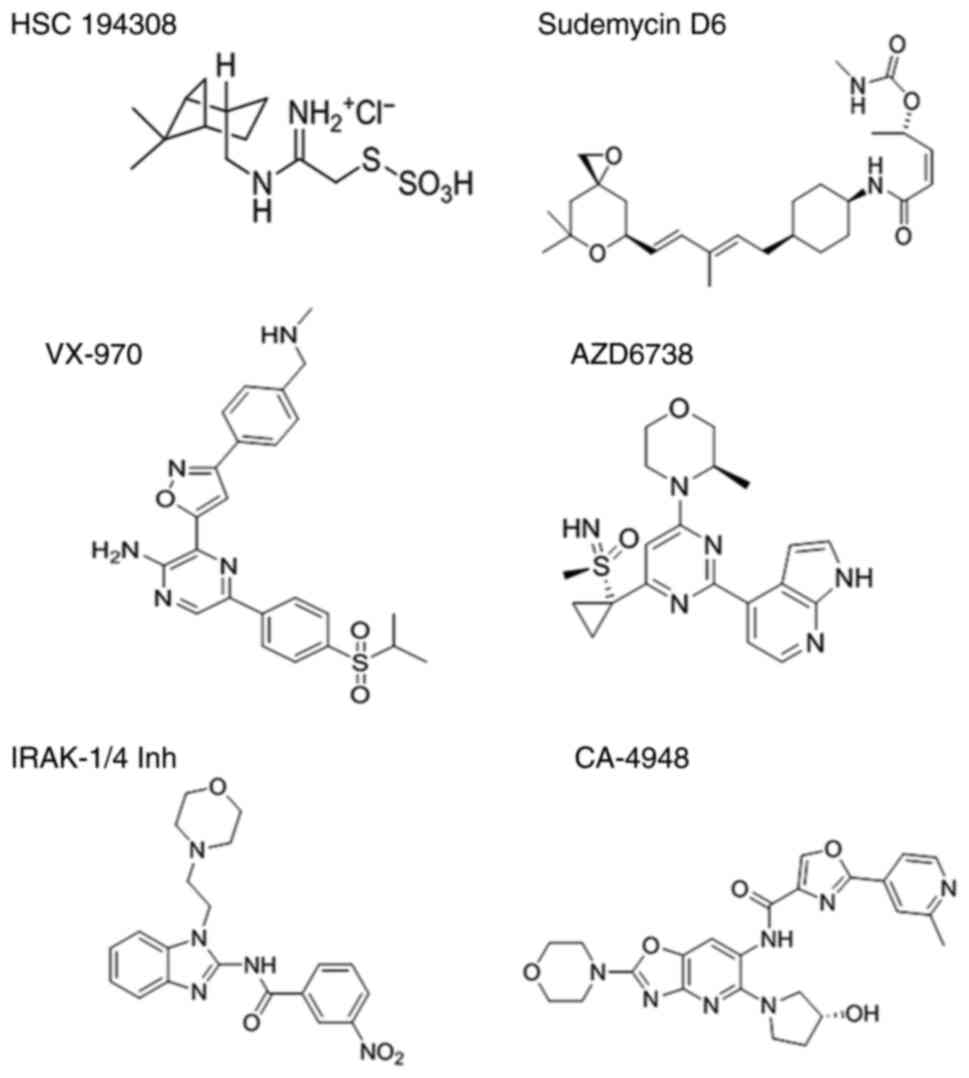
194308
The small-molecule inhibitor NSC 194308 was
synthesized by combining metallic Na and dry MeOH under an
exothermic reaction with the evolution of hydrogen. Subsequently,
(−)-cis-myrtanylamine was added and incubated for 16 h at room
temperature before being reacted with isopropyl alcohol (iPrOH) and
diethyl ether to form 2-chloro-N-(−)-cis-myrtanyl acetamidinium
hydrochloride. The solution was stirred in ethyl (Et) alcohol and
combined with Na2S2O3 in water R, and the compound's formation was
monitored using thin layer chromatography (85). As a result, NSC 194308 was
successfully synthesized and characterized as demonstrated in
Fig. 5.
The experiments of NSC 194308 in HeLa cells revealed
that its addition directly reduced the splicing of the adenovirus
major late promoter transcript substrate. An in-depth analysis of
the intermediate stages of spliceosome assembly revealed that NSC
194308 repressed the pre-mRNA splicing process by inhibiting the
gathering of spliceosomes at a U2AF-dependent checkpoint before
tri-snRNP enlistment and catalytic activity. Subsequent chemical
synthetic treatment with NSC194308 in K562 leukaemia cells, which
had S34F mutations of U2AF1, was followed by high cell apoptosis
(85). Collectively, these data
suggested that NSC194308 is a promising agent for targeted therapy
of leukaemia with U2AF1 S34F and other related mutations.
Synthesis of sudemycin D6
The Julia-Kocienski olefination is of paramount
importance in regard to the synthesis of sudemycin D6. This form is
derived from natural product spliceosome modulators (FR901464)
through intricate chemical reactions, culminating in the formation
of the
(S,Z)-5-(((1R,4R)-4-((2E,4E)-5-((3R,5S)-7,7-dimethyl-1,6-dioxaspiro[2.5]octan-5-yl)-3-methylpenta-2,4-dien-1-yl)cyclohexyl)amino)-5-oxopent-3-en-2-yl-methylcarbamate
structure of sudemycin D6. Notably, the biocatalytic synthesis of
sudemycin D6 was first achieved using the Julia-Kocienski
olefination methodology. In this way, the Julia-Kocienski
olefination is a pivotal step in the successful synthesis of
sudemycin D6 (Fig. 5) (86).
Haematopoietic progenitor cells expressing CD34 were
shown to be sensitive to sudemycin D6 treatment, resulting in
altered pre-mRNA splicing and reduced survival of K562 cells with
U2AF1 (S34F) mutants. The effects of this alteration on the cell
cycle profile were examined in vivo using transplanted mice
treated with sudemycin D6. The results demonstrated that expansion
of U2AF1 mutant progenitors was attenuated, and a beneficial effect
on the bone marrow of U2AF1-mutant mice was observed. Notable
improvements were observed in biological pathways related to the
immune response, leukocyte differentiation and cell proliferation
regulation (87). The
aforementioned study provided evidence that sudemycin D6 is a
promising therapeutic strategy for targeting U2AF1 mutants and may
have broader implications for the treatment of leukaemia.
Ataxia telangiectasia and Rad3-related
protein inhibitor (ATRi)
ATR is an essential protein-coding gene activated by
DNA damage and DNA replication stress. The two known ATR inhibitors
are VX-970 and AZD6738. VX-970, also known as berzosertib, has been
used in clinical trials of patients with ovarian serous tumours,
especially in U2AF1-mutant cancers. Its chemical name is
3-[3-[4-(methylaminomethyl)
phenyl]-5-isoxazolyl]-5-(4-propan-2-ylsulfonylphenyl)-2-pyrazinamine.
Furthermore, compound AZD 6738 (named ceralasertib) is a potent and
effective sulfoximine morpholinopyrimidine ATR inhibitor. The
detailed chemical structure of AZD 6738 is
4-(4-(1-((S(R))-S-methylsulfonimidoyl)cyclopropyl)-6-((3R)-3-methyl-4-morpholinyl)-2-pyrimidinyl)-1H-pyrrolo(2,3-b)pyridine
(Fig. 5). Moreover, AZD6738 can
inhibit ATR through the loss of downstream phosphorylation of CHK1
at Ser345 (88).
Adding ATRi to U2AF1S34F-expressing HeLa and
OCI-AML3 cells demonstrated an increased sensitivity to inhibit
cell growth by inducing DNA damage. Furthermore, HeLa cells
incubated with splicing modulators have been shown to generate a
dependency on ATR to counteract DNA damage, and the modulation of
RNA splicing can make tumour cells more susceptible to ATR
inhibitors. Human primary CD34+ haematopoietic cells
exposed to ATRi exhibit inhibited viability in a dose-dependent
manner. The findings of Nguyen et al (59) revealed that the expression of
U2AF1S34F in human primary CD34+ haematopoietic cells
induces a dramatic decrease in R-loop formation, making them
vulnerable to ATR inhibition. These findings should be further
investigated and validated in larger clinical trials.
IRAK4 inhibitors
A study on U2AF1 mutation and IRAK4 isoforms
indicated that transfecting U2AF1 (S34F) into HEK293 IRAK4exon4
cells generated an increase in IRAK4 exon 4 products from the
wild-type AAG splicing reporter compared with wild-type U2AF1.
However, it did not affect IRAK4 exon 4 from the mutant TAG
splicing reporter. This mutant U2AF1 was found to induce the
expression of IRAK4, which plays a role in the activation of MAPK
and NF-κB signalling to mediate the innate immune pathway (54). It was therefore suggested that
inhibiting IRK4 activity may be a useful therapeutic strategy for
diseases resulting from mutations in U2AF1 S34F.
IRAK1/4-Inh and CA-4948 are two distinct inhibitors
targeting IRAK4. Structurally, the chemical compound of IRAK1/4-Inh
is
N-[1-(2-morpholin-4-ylethyl)-benzimidaz-zol-2-yl]-3-nitrobenzamide,
while the clinical name of CA-4948 is emavusertib, formally named
6′-amino-N-[2-(4-morpholinyl) oxazolo [4, 5-b] pyridin-6-yl]-[2,
3′-bipyridine]-6-carboxamide (Fig.
5). The efficacy of both inhibitors was studied in K562 cells
expressing U2AF1 S34F mutations. Cells incubated with IRAK1/4-Inh
exhibited inhibited growth, which was dependent upon the
deactivation of NF-κB signalling, while primary cells derived from
AML or MDS patients with U2AF1 mutations incubated with CA-4948
underwent myeloid and erythroid differentiation. Taken together,
these results strongly suggest that U2AF1 (S34F) malignant
haematopoietic cells are sensitive to concentrations of IRAK4
inhibitors (89). Thus, inhibiting
IRAK4 may be effective in diseases linked to U2AF1 S34F
mutations.
Therapies targeting U2AF1 in clinical
trials
At present, only a few relevant U2AF1 clinical
studies have been included in clinical trials, namely, NCT04447651,
NCT04278768 and NCT02060409. Of these clinical trials, NCT04278768
was carried out in Phase 1 and Phase 2 for different trial
diseases, while NCT02060409 was completed in clinical trials
(Table II). These studies provide
a promising outlook for the clinical application of U2AF1 in the
future. Through rigorous study design and appropriate controls,
this treatment may bring new opportunities for both diagnosis and
therapy.
| Table II.Targeting U2 small nuclear RNA
auxiliary factor 1 drugs in clinical trials. |
Table II.
Targeting U2 small nuclear RNA
auxiliary factor 1 drugs in clinical trials.
| Drug | Target | Sponsor | Disease | Phase | Identifier |
|---|
| Immunotherapy | SF3B1, U2AF1 | Sidney Kimmel, | Metastatic
Solid | Interventional | NCT04447651 |
|
| SRSF2 | Comprehensive
Cancer, | Tumor |
|
|
|
|
| Center at Johns
Hopkins, |
|
|
|
|
|
| Vanderbilt
University, |
|
|
|
|
|
| Bristol-Myers
Squibb |
|
|
|
| Emavusertib | SF3B1, U2AF1 | Curis, Inc. | Acute
Myelogenous | Phase 1 | NCT04278768 |
| Azacitidine | SRSF2, ZRSR2 |
| Leukaemia | Phase 2 |
|
| Venetoclax |
|
|
Myelodysplastic |
|
|
|
|
|
| Syndrome |
|
|
| Decitabine | SRSF2, U2AF1 | Samsung
Medical |
Myelodysplastic | Interventional | NCT02060409 |
|
| ZRSR2 | Center | Syndrome |
|
|
NCT04447651 aimed to explore the use of spliceosome
mutational markers (PRISMMs) in the treatment of patients with
metastatic solid tumours and haematological malignancies or
lymphoma who may benefit from immunotherapy agents. NCT04278768 was
a dose alteration trial of the monotherapy combination of CA-4948
and venetoclax or azacitidine in the treatment of AML or MDS
patients with mutations in U2AF1, SRSF2, ZRSR2 and SF3B1. As a
response to the increasing use of hypomethylating agents in MDS
treatment, NCT02060409 investigated the prognostic impacts of
U2AF1, SRSF, and ZRSR2 mutations after treatment with decitabine.
Unfortunately, consistent prognostic relevance could not be
established in this trial (90).
Discussion and perspectives
The spliceosome is a dynamic RNP complex that
removes introns from nuclear pre-mRNAs and induces precise splicing
at the 5′ and 3′ splice sites. This complex is composed of five
small nuclear RNAs, U1, U2, U4, U5 and U6. Mutations in each of
these subunits have been associated with various diseases. Notably,
U2AF1, included in the U2 family, has a close correlation with
haematological diseases. Mutations in U2AF1 have been observed in
MDS, MDS-related AML, chronic myeloid leukaemia and PMF. The
pathogenic mechanism of U2AF1 mutation is consideredto impair
pre-mRNA splicing, thereby leading to DNA damage and loss of normal
haematopoietic function (Table I)
(91). Despite these findings,
therapies targeting U2AF1 are scarce in clinical practice.
At present, the prognosis of U2AF1 mutations in
malignant diseases, particularly in haematological diseases,
remains to be fully elucidated. Most U2AF1 mutations have been
observed to manifest as serine at site 34 being replaced by
phenylalanine, along with glutamine at site 157 instead of arginine
or proline. At sites 156 or 158, histidine can be replaced by
arginine. As per previous relevant studies, only NSC194308 has been
found to target U2AF1 directly, while other chemical compounds,
such as sudemycin D6, ATR inhibitor, IRAK1/4-Inh and CA4948
(emavusertib), have been reported to have an indirect effect on
U2AF1 mutations (92). Moreover,
immunotherapies, such as anti-PD-1, anti-PD-L1 and anti-CTLA-4, may
achieve favourable effects on U2AF1 mutations in MDS (93). It has been reported that IRAK4 may
activate NF-kB in U2AF1 mutations, and thus, it is essential to
assess the efficacy of NF-kB inhibitors in the therapy of U2AF1
mutations. Bortezomib or denosumab effectively broadly blocks the
activity of NF-kB. Other NF-kB inhibitors known as IκBs are
proteins that can mask nuclear localization signals of NF-κB to
restrain its function. Given the association of NF-kB with U2AF1
mutations, it is necessary to explore the role of these inhibitors
in therapies targeting U2AF1 mutations (94–96).
In a study on oestrogen receptor-positive breast cancer, it was
found that the silencing of U2AF1 in a zebrafish model might reduce
the activity of various cell adhesion molecules, which further
resulted in malignant differentiation. At the same time, natural
products deserve their own stage, leveraging a computer-aided drug
discovery online tool (http://cao.labshare.cn/drugrep/). A total of two
natural products with favourable affinity were screened, both of
them from Chinese herbal monomer extracts, named tiliroside and
1,3,6-tri-O-galloylglucose (Fig.
6). However, pre-silencing of U2AF1 RNAs may also promote new
therapeutic options (97).
Given the limited availability of targeted therapies
for spliceosomes, there is an urgent need to design more molecular
targeted agents that act on cell proliferation and regulate tumour
metabolism with the U2AF1 mutation representing a significant
target. Furthermore, further investigations into the targeting of
spliceosome mutations should be carried out to explore the
potential applications in the treatment of immunological or
malignant diseases. Further studies are necessary to improve
understanding of the implications of U2AF1 mutations.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Young Scientists Fund of
the National Natural Science Foundation of China (grant no.
82204858) and the Sichuan Science and Technology Program (grant no.
2023NSFSC1761).
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
QN, YHL, JWL and RXL wrote or contributed to the
writing of the manuscript. FRL, JHZ, LYZ, RXL and ZJY revised or
contributed to revising the manuscript. Data authentication is not
applicable. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ravi S, Schilder RJ and Kimball SR: Role
of precursor mRNA splicing in nutrient-induced alterations in gene
expression and metabolism. J Nutr. 145:841–846. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Saha K, Fernandez MM, Biswas T, Joseph S
and Ghosh G: Discovery of a pre-mRNA structural scaffold as a
contributor to the mammalian splicing code. Nucleic Acids Res.
49:7103–7121. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scotti MM and Swanson MS: RNA mis-splicing
in disease. Nat Rev Genet. 17:19–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Blijlevens M, Li J and van Beusechem VW:
Biology of the mRNA splicing machinery and its dysregulation in
cancer providing therapeutic opportunities. Int J Mol Sci.
22:51102021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cléry A, Sinha R, Anczuków O, Corrionero
A, Moursy A, Daubner GM, Valcárcel J, Krainer AR and Allain FH:
Isolated pseudo-RNA-recognition motifs of SR proteins can regulate
splicing using a noncanonical mode of RNA recognition. Proc Natl
Acad Sci USA. 110:E2802–E2811. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moraleva AA, Deryabin AS, Rubtsov YP,
Rubtsova MP and Dontsova OA: Eukaryotic ribosome biogenesis: The
60S subunit. Acta Naturae. 14:39–49. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carrocci TJ and Neugebauer KM: Pre-mRNA
splicing in the nuclear landscape. Cold Spring Harb Symp Quant
Biol. 84:11–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meyer K, Koester T and Staiger D: Pre-mRNA
splicing in plants: In vivo functions of RNA-binding proteins
implicated in the splicing process. Biomolecules. 5:1717–1740.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sette C and Paronetto MP: Somatic
mutations in core spliceosome components promote tumorigenesis and
generate an exploitable vulnerability in human cancer. Cancers
(Basel). 14:18272022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ilagan JO, Ramakrishnan A, Hayes B, Murphy
ME, Zebari AS, Bradley P and Bradley RK: U2AF1 mutations alter
splice site recognition in hematological malignancies. Genome Res.
25:14–26. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yoshimi A and Abdel-Wahab O: Molecular
pathways: Understanding and targeting mutant spliceosomal proteins.
Clin Cancer Res. 23:336–341. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fei DL, Zhen T, Durham B, Ferrarone J,
Zhang T, Garrett L, Yoshimi A, Abdel-Wahab O, Bradley RK, Liu P and
Varmus H: Impaired hematopoiesis and leukemia development in mice
with a conditional knock-in allele of a mutant splicing factor gene
U2af1. Proc Natl Acad Sci USA. 115:E10437–E10446. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen S, Benbarche S and Abdel-Wahab O:
Splicing factor gene mutations in hematologic malignancies. Blood.
129:1260–1269. 2017. View Article : Google Scholar
|
|
14
|
Ogawa S: Genetics of MDS. Blood.
133:1049–1059. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Inoue D, Bradley RK and Abdel-Wahab O:
Spliceosomal gene mutations in myelodysplasia: Molecular links to
clonal abnormalities of hematopoiesis. Genes Dev. 30:989–1001.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Taylor J and Lee SC: Mutations in
spliceosome genes and therapeutic opportunities in myeloid
malignancies. Genes Chromosomes Cancer. 58:889–902. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Przychodzen B, Jerez A, Guinta K, Sekeres
MA, Padgett R, Maciejewski JP and Makishima H: Patterns of
missplicing due to somatic U2AF1 mutations in myeloid neoplasms.
Blood. 122:999–1006. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Madan V, Li J, Zhou S, Teoh WW, Han L,
Meggendorfer M, Malcovati L, Cazzola M, Ogawa S, Haferlach T, et
al: Distinct and convergent consequences of splice factor mutations
in myelodysplastic syndromes. Am J Hematol. 95:133–143. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou Z, Gong Q, Wang Y, Li M, Wang L, Ding
H and Li P: The biological function and clinical significance of
SF3B1 mutations in cancer. Biomark Res. 8:382020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gupta G, Singh R, Kotasthane DS and
Kotasthane VD: Myelodysplastic syndromes/neoplasms: Recent
classification system based on World Health Organization
classification of tumors-international agency for research on
cancer for hematopoietic and lymphoid tissues. J Blood Med.
1:171–182. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Estey E, Hasserjian RP and Döhner H:
Distinguishing AML from MDS: A fixed blast percentage may no longer
be optimal. Blood. 139:323–332. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen J, Kao YR, Sun D, Todorova TI,
Reynolds D, Narayanagari SR, Montagna C, Will B, Verma A and Steidl
U: Myelodysplastic syndrome progression to acute myeloid leukemia
at the stem cell level. Nat Med. 25:103–110. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu J, Li Y, Li T, Li Y, Xing H, Sun H, Sun
L, Wan D, Liu Y, Xie X and Jiang Z: Gene mutational analysis by NGS
and its clinical significance in patients with myelodysplastic
syndrome and acute myeloid leukemia. Exp Hematol Oncol. 9:22020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Čolović N, Denčić-Fekete M, Peruničić M
and Jurišić V: Clinical characteristics and treatment outcome of
hypocellular acute myeloid leukemia based on WHO classification.
Indian J Hematol Blood Transfus. 36:59–63. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Visconte V, O Nakashima M and J Rogers H:
Mutations in Splicing Factor Genes In Myeloid Malignancies:
Significance and impact on clinical features. Cancers (Basel).
11:18442019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Follo MY, Pellagatti A, Ratti S,
Ramazzotti G, Faenza I, Fiume R, Mongiorgi S, Suh PG, McCubrey JA,
Manzoli L, et al: Recent advances in MDS mutation landscape:
Splicing and signalling. Adv Biol Regul. 75:1006732020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brunner AM and Steensma DP: Targeting
aberrant splicing in myelodysplastic syndromes: Biologic rationale
and clinical opportunity. Hematol Oncol Clin North Am. 34:379–391.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Douet-Guilbert N, Soubise B, Bernard DG
and Troadec MB: Cytogenetic and genetic abnormalities with
diagnostic value in myelodysplastic syndromes (MDS): Focus on the
pre-messenger RNA splicing process. Diagnostics (Basel).
12:16582022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dong Y and Li J, Zeng Z, Zhang X, Liang M,
Yi H, Luo J and Li J: Growth retardation and congenital heart
disease in a boy with a ring chromosome 6 of maternal origin. Mol
Cytogenet. 15:92022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li B, Zou D, Yang S, Ouyang G and Mu Q:
Prognostic significance of U2AF1 mutations in myelodysplastic
syndromes: A meta-analysis. J Int Med Res.
48:3000605198910132020.PubMed/NCBI
|
|
31
|
Awada H, Thapa B and Visconte V: The
genomics of myelodysplastic syndromes: Origins of disease
evolution, biological pathways, and prognostic implications. Cells.
9:25122020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Esfahani MS, Lee LJ, Jeon YJ, Flynn RA,
Stehr H, Hui AB, Ishisoko N, Kildebeck E, Newman AM, Bratman SV, et
al: Functional significance of U2AF1 S34F mutations in lung
adenocarcinomas. Nat Commun. 10:57122019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kielkopf CL: Insights from structures of
cancer-relevant pre-mRNA splicing factors. Curr Opin Genet Dev.
48:57–66. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Escobar-Hoyos L, Knorr K and Abdel-Wahab
O: Aberrant RNA splicing in cancer. Annu Rev Cancer Biol.
3:167–185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Biancon G, Joshi P, Zimmer JT, Hunck T,
Gao Y, Lessard MD, Courchaine E, Barentine AE, Machyna M, Botti V,
et al: Multi-omics profiling of U2AF1 mutants dissects pathogenic
mechanisms affecting RNA granules in myeloid malignancies. bioRxiv.
2021.2004.2022.441020. 2021.
|
|
36
|
Martínez-Valiente C, Garcia-Ruiz C, Rosón
B, Liquori A, González-Romero E, Fernández-González R,
Gómez-Redondo I, Cervera J, Gutiérrez-Adán A and Sanjuan-Pla A:
Aberrant alternative splicing in U2af1/Tet2 double mutant mice
contributes to major hematological phenotypes. Int J Mol Sci.
22:69632021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ivy KS and Brent Ferrell P Jr: Disordered
immune regulation and its therapeutic targeting in myelodysplastic
syndromes. Curr Hematol Malig Rep. 13:244–255. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pellagatti A, Armstrong RN, Steeples V,
Sharma E, Repapi E, Singh S, Sanchi A, Radujkovic A, Horn P,
Dolatshad H, et al: Impact of spliceosome mutations on RNA splicing
in myelodysplasia: dysregulated genes/pathways and clinical
associations. Blood. 132:1225–1240. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Akef A, McGraw K, Cappell SD and Larson
DR: Ribosome biogenesis is a downstream effector of the oncogenic
U2AF1-S34F mutation. PLoS Biol. 18:e30009202020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu Y and Ruggero D: The role of
translation control in tumorigenesis and its therapeutic
implications. Ann Rev Cancer Biol. 4:437–457. 2020. View Article : Google Scholar
|
|
41
|
Feliu N, Kohonen P, Ji J, Zhang Y,
Karlsson HL, Palmberg L, Nyström A and Fadeel B: Next-generation
sequencing reveals low-dose effects of cationic dendrimers in
primary human bronchial epithelial cells. ACS Nano. 9:146–163.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hallstrom TC, Mori S and Nevins JR: An
E2F1-dependent gene expression program that determines the balance
between proliferation and cell death. Cancer Cell. 13:11–22. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang H, Guo Y, Dong Z, Li T, Xie X, Wan D,
Jiang Z, Yu J and Guo R: Differential U2AF1 mutation sites, burden
and co-mutation genes can predict prognosis in patients with
myelodysplastic syndrome. Sci Rep. 10:186222020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Huang FT, Sun J, Zhang L, He X, Zhu YH,
Dong HJ, Wang HY, Zhu L, Zou JY, Huang JW and Li L: Role of SIRT1
in hematologic malignancies. J Zhejiang Univ Sci B. 20:391–398.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Carraway HE, Malkaram SA, Cen Y, Shatnawi
A, Fan J, Ali HEA, Abd Elmageed ZY, Buttolph T, Denvir J, Primerano
DA and Fandy TE: Activation of SIRT6 by DNA hypomethylating agents
and clinical consequences on combination therapy in leukemia. Sci
Rep. 10:103252020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bhalla S and Gordon LI: Functional
characterization of NAD dependent de-acetylases SIRT1 and SIRT2 in
B-cell chronic lymphocytic leukemia (CLL). Cancer Biol Ther.
17:300–309. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen ML, Logan TD, Hochberg ML, Shelat SG,
Yu X, Wilding GE, Tan W, Kujoth GC, Prolla TA, Selak MA, et al:
Erythroid dysplasia, megaloblastic anemia, and impaired
lymphopoiesis arising from mitochondrial dysfunction. Blood.
114:4045–4053. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Luo Y, Ma J and Lu W: The significance of
mitochondrial dysfunction in cancer. Int J Mol Sci. 21:55982020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park SM, Ou J, Chamberlain L, Simone TM,
Yang H, Virbasius CM, Ali AM, Zhu LJ, Mukherjee S, Raza A and Green
MR: U2AF35(S34F) promotes transformation by directing aberrant ATG7
pre-mRNA 3′ end formation. Mol Cell. 62:479–490. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhao L, Cao J, Hu K, He X, Yun D, Tong T
and Han L: Sirtuins and their biological relevance in aging and
age-related diseases. Aging Dis. 11:927–945. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bosch-Presegué L and Vaquero A: The dual
role of sirtuins in cancer. Genes Cancer. 2:648–662. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nakagawa MM, Chen H and Rathinam CV:
Constitutive activation of NF-κB pathway in hematopoietic stem
cells causes loss of quiescence and deregulated transcription
factor networks. Front Cell Dev Biol. 6:1432018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Balka KR and De Nardo D: Understanding
early TLR signaling through the myddosome. J Leukoc Biol.
105:339–351. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Smith MA, Choudhary GS, Pellagatti A, Choi
K, Bolanos LC, Bhagat TD, Gordon-Mitchell S, Von Ahrens D, Pradhan
K, Steeples V, et al: U2AF1 mutations induce oncogenic IRAK4
isoforms and activate innate immune pathways in myeloid
malignancies. Nat Cell Biol. 21:640–650. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pellagatti A and Boultwood J: SF3B1 mutant
myelodysplastic syndrome: Recent advances. Adv Biol Regul.
79:1007762021. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lee SC, North K, Kim E, Jang E, Obeng E,
Lu SX, Liu B, Inoue D, Yoshimi A, Ki M, et al: Synthetic lethal and
convergent biological effects of cancer-associated spliceosomal
gene mutations. Cancer Cell. 34:225–241.e8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Crossley MP, Bocek M and Cimprich KA:
R-loops as cellular regulators and genomic threats. Mol Cell.
73:398–411. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen C, Zhou P, Zhang Z and Liu Y: U2AF1
mutation connects DNA damage to the alternative splicing of RAD51
in lung adenocarcinomas. Clin Exp Pharmacol Physiol. 49:740–747.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Nguyen HD, Leong WY, Li W, Reddy PNG,
Sullivan JD, Walter MJ, Zou L and Graubert TA: Spliceosome
mutations induce R loop-associated sensitivity to ATR inhibition in
myelodysplastic syndromes. Cancer Res. 78:5363–5374. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen H, Libring S, Ruddraraju KV, Miao J,
Solorio L, Zhang ZY and Wendt MK: SHP2 is a multifunctional
therapeutic target in drug resistant metastatic breast cancer.
Oncogene. 39:7166–7180. 2019. View Article : Google Scholar
|
|
61
|
Chen L, Chen JY, Huang YJ, Gu Y, Qiu J,
Qian H, Shao C, Zhang X, Hu J, Li H, et al: The augmented R-loop is
a unifying mechanism for myelodysplastic syndromes induced by
high-risk splicing factor mutations. Mol Cell. 69:412–425.e6. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Boultwood J and Pellagatti A: The impact
of spliceosome mutations in MDS. Hemasphere. 3 (Suppl):S132–S134.
2019. View Article : Google Scholar
|
|
63
|
Vallapureddy RR, Mudireddy M, Penna D,
Lasho TL, Finke CM, Hanson CA, Ketterling RP, Begna KH, Gangat N,
Pardanani A and Tefferi A: Leukemic transformation among 1306
patients with primary myelofibrosis: Risk factors and development
of a predictive model. Blood Cancer J. 9:122019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tefferi A, Siragusa S, Hussein K, Schwager
SM, Hanson CA, Pardanani A, Cervantes F and Passamonti F:
Transfusion-dependency at presentation and its acquisition in the
first year of diagnosis are both equally detrimental for survival
in primary myelofibrosis-prognostic relevance is independent of
IPSS or karyotype. Am J Hematol. 85:14–17. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Alhuraiji A, Naqvi K, Huh YO, Ho C,
Verstovsek S and Bose P: Acute lymphoblastic leukemia secondary to
myeloproliferative neoplasms or after lenalidomide exposure. Clin
Case Rep. 6:155–161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Tefferi A, Lasho TL, Finke CM, Knudson RA,
Ketterling R, Hanson CH, Maffioli M, Caramazza D, Passamonti F and
Pardanani A: CALR vs JAK2 vs MPL-mutated or triple-negative
myelofibrosis: Clinical, cytogenetic and molecular comparisons.
Leukemia. 28:1472–1477. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chaligné R, James C, Tonetti C, Besancenot
R, Le Couédic JP, Fava F, Mazurier F, Godin I, Maloum K, Larbret F,
et al: Evidence for MPL W515L/K mutations in hematopoietic stem
cells in primitive myelofibrosis. Blood. 110:3735–3743. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Tefferi A, Finke CM, Lasho TL, Hanson CA,
Ketterling RP, Gangat N and Pardanani A: U2AF1 mutation types in
primary myelofibrosis: Phenotypic and prognostic distinctions.
Leukemia. 32:2274–2278. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Tefferi A, Mudireddy M, Finke CM, Nicolosi
M, Lasho TL, Hanson CA, Patnaik MM, Pardanani A and Gangat N: U2AF1
mutation variants in myelodysplastic syndromes and their clinical
correlates. Am J Hematol. 93:E146–E148. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wu SJ, Tang JL, Lin CT, Kuo YY, Li LY,
Tseng MH, Huang CF, Lai YJ, Lee FY, Liu MC, et al: Clinical
implications of U2AF1 mutation in patients with myelodysplastic
syndrome and its stability during disease progression. Am J
Hematol. 88:E277–E282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Patnaik MM and Tefferi A: Chronic
myelomonocytic leukemia: 2020 Update on diagnosis, risk
stratification and management. Am J Hematol. 95:97–115. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Machherndl-Spandl S, Jäger E, Barna A,
Gurbisz M, Marschon R, Graf T, Graf E, Geissler C, Hoermann G,
Nösslinger T, et al: Impact of age on the cumulative risk of
transformation in patients with chronic myelomonocytic leukaemia.
Eur J Haematol. 107:265–274. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Patnaik MM, Lasho TL, Finke CM, Hanson CA,
Hodnefield JM, Knudson RA, Ketterling RP, Pardanani A and Tefferi
A: Spliceosome mutations involving SRSF2, SF3B1, and U2AF35 in
chronic myelomonocytic leukemia: Prevalence, clinical correlates,
and prognostic relevance. Am J Hematol. 88:201–206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Patnaik MM and Tefferi A: Cytogenetic and
molecular abnormalities in chronic myelomonocytic leukemia. Blood
Cancer J. 6:e3932016. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
McClure RF, Ewalt MD, Crow J,
Temple-Smolkin RL, Pullambhatla M, Sargent R and Kim AS: Clinical
significance of DNA variants in chronic myeloid neoplasms: A report
of the association for molecular pathology. J Mol Diagn.
20:717–737. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Grever MR, Abdel-Wahab O, Andritsos LA,
Banerji V, Barrientos J, Blachly JS, Call TG, Catovsky D, Dearden
C, Demeter J, et al: Consensus guidelines for the diagnosis and
management of patients with classic hairy cell leukemia. Blood.
129:553–560. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kreitman RJ: Hairy cell leukemia: Present
and future directions. Leuk Lymphoma. 60:2869–2879. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Durham BH, Getta B, Dietrich S, Taylor J,
Won H, Bogenberger JM, Scott S, Kim E, Chung YR, Chung SS, et al:
Genomic analysis of hairy cell leukemia identifies novel recurrent
genetic alterations. Blood. 130:1644–1648. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Rahman MA, Krainer AR and Abdel-Wahab O:
SnapShot: Splicing alterations in cancer. Cell. 180:208–208.e1.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fei DL, Motowski H, Chatrikhi R, Prasad S,
Yu J, Gao S, Kielkopf CL, Bradley RK and Varmus H: Wild-type U2AF1
antagonizes the splicing program characteristic of U2AF1-mutant
tumors and is required for cell survival. PLOS Genetics.
12:e10063842016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Sanchez A, El Ouardi D, Houfaf Khoufaf FZ,
Idrissou M, Boisnier T, Penault-Llorca F, Bignon YJ, Guy L and
Bernard-Gallon D: Role of JMJD3 demethylase and its inhibitor
GSK-J4 in regulation of MGMT, TRA2A, RPS6KA2, and U2AF1 genes in
prostate cancer cell lines. OMICS. 24:505–507. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Cao H, Wang D, Gao R, Chen L and Feng Y:
Down regulation of U2AF1 promotes ARV7 splicing and prostate cancer
progression. Biochem Biophys Res Commun. 541:56–62. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
El Ouardi D, Idrissou M, Sanchez A,
Penault-Llorca F, Bignon YJ, Guy L and Bernard-Gallon D: The
inhibition of the histone methyltransferase EZH2 by DZNEP or SiRNA
demonstrates its involvement in MGMT, TRA2A, RPS6KA2, and U2AF1
gene regulation in prostate cancer. OMICS. 24:116–118. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Je EM, Yoo NJ, Kim YJ, Kim MS and Lee SH:
Mutational analysis of splicing machinery genes SF3B1, U2AF1 and
SRSF2 in myelodysplasia and other common tumors. Int J Cancer.
133:260–265. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Chatrikhi R, Feeney CF, Pulvino MJ,
Alachouzos G, MacRae AJ, Falls Z, Rai S, Brennessel WW, Jenkins JL,
Walter MJ, et al: A synthetic small molecule stalls pre-mRNA
splicing by promoting an early-stage U2AF2-RNA complex. Cell Chem
Biol. 28:1145–1157.e6. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Lagisetti C, Palacios G, Goronga T,
Freeman B, Caufield W and Webb TR: Optimization of antitumor
modulators of pre-mRNA splicing. J Med Chem. 56:10033–10044. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Shirai CL, White BS, Tripathi M, Tapia R,
Ley JN, Ndonwi M, Kim S, Shao J, Carver A, Saez B, et al: Mutant
U2AF1-expressing cells are sensitive to pharmacological modulation
of the spliceosome. Nat Commun. 8:140602017. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Middleton MR, Dean E, Evans TRJ, Shapiro
GI, Pollard J, Hendriks BS, Falk M, Diaz-Padilla I and Plummer R:
Phase 1 study of the ATR inhibitor berzosertib (formerly M6620,
VX-970) combined with gemcitabine ± cisplatin in patients with
advanced solid tumours. Br J Cancer. 125:510–519. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Powers JP, Li S, Jaen JC, Liu J, Walker
NP, Wang Z and Wesche H: Discovery and initial SAR of inhibitors of
interleukin-1 receptor-associated kinase-4. Bioorg Med Chem Lett.
16:2842–2845. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Thol F, Kade S, Schlarmann C, Löffeld P,
Morgan M, Krauter J, Wlodarski MW, Kölking B, Wichmann M, Görlich
K, et al: Frequency and prognostic impact of mutations in SRSF2,
U2AF1, and ZRSR2 in patients with myelodysplastic syndromes. Blood.
119:3578–3584. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Griffin C and Saint-Jeannet JP:
Spliceosomopathies: Diseases and mechanisms. Dev Dyn.
249:1038–1046. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Zhao Y, Cai W, Hua Y, Yang X and Zhou J:
The biological and clinical consequences of RNA splicing factor
U2AF1 mutation in myeloid malignancies. Cancers (Basel).
14:44062022. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Lee P, Yim R, Yung Y, Chu HT, Yip PK and
Gill H: Molecular targeted therapy and immunotherapy for
myelodysplastic syndrome. Int J Mol Sci. 22:102322021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Jacobs MD and Harrison SC: Structure of an
IkappaBalpha/NF-kappaB complex. Cell. 95:749–758. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Raedler L: Velcade (bortezomib) receives 2
new FDA indications: For retreatment of patients with multiple
myeloma and for first-line treatment of patients with mantle-cell
lymphoma. Am Health Drug Benefits. 8:135–140. 2015.PubMed/NCBI
|
|
96
|
Hamdy NA: Denosumab: RANKL inhibition in
the management of bone loss. Drugs Today (Barc). 44:7–21. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Vazquez Rodriguez G, Abrahamsson A,
Turkina MV and Dabrosin C: Lysine in combination with estradiol
promote dissemination of estrogen receptor positive breast cancer
via upregulation of U2AF1 and RPN2 proteins. Front Oncol.
10:5986842020. View Article : Google Scholar : PubMed/NCBI
|