Introduction
Cancer cachexia is a devitalizing multifactorial
syndrome that affects several metabolic processes in multiple
organs. It is characterized by progressive weight loss due to the
depletion of muscle with or without loss of adipose tissue mass,
which cannot be recovered by nutritional therapy (1). Patients with cachexia exhibit
decreased responses to chemotherapy and a diminished tolerance to
anticancer treatments (1).
Skeletal muscle wasting is a hallmark of cachexia
(2) and induces pathophysiological
changes, such as a weak and fragile body (3). Moreover, patients with cachexia
experience dysregulations in protein metabolism, decreased protein
synthesis and upregulated whole-body protein turnover (4). The onset of muscle atrophy is a
consequence of an imbalance between the synthesis and breakdown of
skeletal muscle proteins (5). The
muscle-specific E3 ubiquitin ligases, MAFbx/atrogin-1 (atrogin-1)
and muscle RING-finger protein-1 (MuRF1) are the key regulators of
muscle atrophy in cancer cachexia (6). Notably, they are upregulated in the
skeletal muscles of patients with cancer cachexia, thereby
indicating the presence of muscular atrophy (6). In addition, the knockdown of atrogin-1
or MuRF1 ameliorated muscle wasting in patients with cancer
cachexia (6,7).
In cases where tumor and muscle tissues exist
distantly from each other, proinflammatory cytokines, such as IL-6
and TNF-α, mediate signals to promote the breakdown of proteins,
while also inhibiting protein synthesis (8). Systemic inflammation and the resulting
catabolic stimuli cause cachexia by suppressing the synthesis of
muscle protein and promoting muscle catabolism and atrophy
(9). The liver is also involved in
regulating metabolic processes in cancer cachexia (10). In patients with cancer cachexia,
amino acids released by muscle wasting can be used for hepatic
gluconeogenesis (11). Moreover,
inflammatory cytokines, such as IL-6 produced by activated
macrophages stimulate the liver to produce an acute phase response
(12).
Muscle stemness can be damaged when cancer cachexia
impedes muscle stem cell differentiation (13). In cancer cachexia, impaired muscle
stem cell function can cause skeletal muscle atrophy by decreasing
the regeneration of muscle myofibers (13). When quiescent muscle satellite cells
are activated, they coexpress paired box 7 (Pax7) and myoblast
determination protein 1 (MyoD), two major transcription factors of
myogenic differentiation (14,15).
The PI3K/Akt signaling pathway is reportedly
involved in protein turnover in muscle tissues (16). Whereby the phosphorylation of PI3K
causes the insulin-like growth factor (IGF)-1 to activate Akt and
mTOR, which promotes protein synthesis and muscle proliferation
(17). Thereby, cancer cachexia
commonly exhibits an inhibited PI3K/Akt signaling pathway during
muscle atrophy (18).
β-carotene (BC) is a provitamin A carotenoid and a
strong antioxidant, which can scavenge free radicals (19,20).
Oxidative stress can cause a reduction in the intracellular
antioxidant pool, which results in the disturbance of the myogenic
differentiation and muscle atrophy (21,22).
Moreover, since BC acts as a strong antioxidant, it can defend
against oxidative stress and DNA damage (23). In fact, BC exerted protective
effects against oxidative stress (100 µM of
H2O2)-induced muscle atrophy in C2C12
myotubes and soleus muscle atrophy in mice (24). In addition, combination treatments
of the representative antioxidants including BC, astaxanthin, and
resveratrol, increased protein synthesis in muscles during
hypertrophy following atrophy in mice (25).
The concentration of serum carotenoid concentration
has been revealed to be negatively associated with the risk of
decreased muscle strength and walking pace (26,27).
However, it has been reported that supplementation with BC
decreased the soleus muscle mass loss in mice caused by denervation
(24). It has been demonstrated
that BC administration elevated muscle mass and functional
hypertrophy of the soleus muscle in mice under physiological
conditions (28). A combination of
BC, astaxanthin and resveratrol increased protein synthesis and
counteracted muscle hypertrophy in mice (25). BC supplementation also upregulated
myoblast differentiation in chicken (29). In addition, the mean serum level of
BC was significantly reduced in patients with cachexia (30). However, the effects of BC on the
regulation of muscle atrophy and myogenic differentiation during
cancer cachexia have yet to be investigated.
Thus, the aim of the present study was to
investigate whether: i) BC can suppress aberrant muscle
differentiation and atrophy induced by cancer cachexia; ii) the
PI3K/Akt pathway can contribute to this effect; and iii) increased
hepatic gluconeogenesis and systemic inflammation caused by cancer
cachexia can be countered by BC supplementation.
Materials and methods
Cell culture and reagents
Lewis lung carcinoma (LLC) cells (CRL-1642,
http://www.atcc.org/products/crl-1642) and C2C12
myoblasts (CRL-1772, http://www.atcc.org/products/crl-1772) were purchased
from American Type Culture Collection and cultured in a growth
medium containing Dulbecco's modified Eagle's medium (DMEM;
Welgene, Inc.) with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) and 1% streptomycin-penicillin (Invitrogen;
Thermo Fisher Scientific, Inc.) in an incubator at 37°C with a
humidified atmosphere of 5% CO2. BC was purchased from
MilliporeSigma and dissolved in tetrahydrofuran (THF;
MilliporeSigma). Mycoplasma testing was conducted for these cell
lines and it was confirmed that they were mycoplasma-free. Fresh BC
stock solution was prepared before each application and stored
under dim light.
Cancer cachexia mouse model
Male C57BL/6J mice were purchased at 5 weeks of age
from Central Lab Animal Inc. All mice were individually housed in a
12:12-h light-dark cycle and fed AIN-93G purified rodent pellet
diets (RaonBio Inc.) ad libitum. After 1 week of
acclimatization, the mice were randomized and sorted into four
groups: i) Control mice (CTRL; n=12); ii) LLC cell-induced cancer
cachexia mice (CC; n=12); iii) LLC cell-induced cancer cachexia
mice supplemented with BC at 4 mg/kg body weight (BW) (BC 4; n=12);
and iv) LLC cell-induced cancer cachexia mice supplemented with BC
at 8 mg/kg BW (BC 8; n=12). To induce cancer cachexia,
1×106 LLC cells were diluted in 100 µl of
phosphate-buffered saline and subcutaneously inoculated into the
right hindlimb of each mouse. The mice were orally administered BC
4 and BC 8 dissolved in 100 µl of corn oil twice a week throughout
the experimental period.
BW and tumor volume were measured every other day.
Tumor volume was calculated as width (mm) × length2/2,
with dimensions presented in millimeters (31). The diameter and volume of the
largest tumor of mice on the day of sacrifice was 2.0 cm and 2.9
cm3. Diameter measurements were decreased by 0.1 cm for
skin and subcutaneous fat. Notably, 22 days after tumor cell
inoculation, all mice were sacrificed by CO2 inhalation,
and the tissues and blood were collected. When sacrificing the
mice, the CO2 flow rate for euthanasia was calculated.
CO2 was delivered at three different flow rates: 30, 50
and 70% of the cage volume per minute, corresponding to rates of
5.9, 9.9 and 13.9 l/min, respectively. Serum samples were obtained
by centrifugation at 25,553 × g for 15 min at 4°C. Moreover, the
weights of the organs (spleen, liver, etc.), muscles
(gastrocnemius, pectoralis, etc.) and fats (subcutaneous,
perirenal, etc.) were measured. Liver and gastrocnemius muscles
were extracted for PCR, western blot and histological analyses.
The weights of the mouse carcasses and tumors were
measured immediately after sacrifice. ‘Carcass-tumor weight’ was
obtained by subtracting the tumor weight from the mouse carcass
weight.
The present study was conducted according to the
guidelines of the National Institutes of Health (NIH publication
no. 8023, revised 1978), and approved (IACUC approval no. EWHA
IACUC 21-003-1) by the Institutional Animal Care and Use Committee
of Ewha Womans University (Seoul, Republic of Korea).
Grip strength assessment
Grip strength was determined using a grip strength
meter (Jeung-Do Bio & Plant Co., Ltd.). After allowing the
mouse to grip a mesh bar linked to a force transducer, the mouse
tail was pulled gently until its grip was released, and the peak
force (g) generated was recorded by the transducer. Reported values
are the average of the measurements for each mouse, with a 1-min
interval between sets.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from cells and gastrocnemius
muscles using TRIzol reagent (Thermo Fisher Scientific, Inc.).
Reverse transcription was then performed according to the
manufacturer's protocol using a cDNA Reverse-Transcription kit
(Thermo Fisher Scientific, Inc.). The resulting cDNA was used to
perform quantitative polymerase chain reaction (PCR) amplification
using a SYBR Green master mix (Qiagen GmbH). Briefly, cDNAs were
mixed with 2X Rotor-Gene SYBR Green PCR Master Mix and the cycling
program consisted of one cycle at 95°C for 5 min, followed by 40
cycles at 95°C for 5 sec, and 60°C for 10 sec. The expression of
all genes was normalized relative to the expression of
glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The sequences of
the primers for RT-qPCR are listed in Table I. The relative quantification was
performed by the common 2−ΔΔCq method (32).
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Mouse species
gene | Forward primer (5′
to 3′) | Reverse primer (5′
to 3′) |
|---|
| Murf1 |
TGACATCTACAAGCAGGAGTGC |
TCGTCTTCGTGTTCCTTGC |
|
Atrogin-1 |
AGTGAGGACCGGCTACTGTG |
GATCAAACGCTTGCGAATCT |
| MyoD |
CTACAGTGGCGACTCAGATG |
TGTAGTAGGCGGTGTCGTAG |
| Pax7 |
CTGGATGAGGGCTCAGATGT |
GGTTAGCTCCTGCCTGCTTA |
| G6pase |
AGGAAGGATGGAGGAAGGAA |
TGGAACCAGATGGGAAAGAG |
| Pepck |
AGAGCAGAGAGACACAGTGC |
AGGGCGAGTCTGTCAGTTCA |
| Gapdh |
AACTTTGGCATTGTGGAAGG |
TGTGAGGGAGATGCTCAGTG |
Western blot analysis
Protein samples from cells and gastrocnemius muscles
were extracted using a radioimmunoprecipitation assay (RIPA) lysis
buffer [150 mM NaCl, 50 mM Tris-hydrochloride (pH 7.5), 1% Nonidet,
P-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS),
1 mM phenylmethylsulfonyl fluoride, 1 mM
Na3VO4, and 1 mM sodium fluoride]. Protein
concentrations were determined by Bradford protein assay (Bio-Rad
Laboratories, Inc.) and western blotting was then performed. The
proteins (60 µg) were loaded and separated on a sodium dodecyl
sulfate (SDS) denaturing polyacrylamide 12% gel and transferred to
a PVDF membrane. The proteins were blocked with 5% skim milk for 1
h at room temperature and incubated with the respective primary
antibodies overnight at 4°C. Primary antibodies included anti-MuRF1
(1:200; cat. no. sc-398608; Santa Cruz Biotechnology, Inc.),
anti-atrogin-1 (1:200; cat. no. sc-166806; Santa Cruz
Biotechnology, Inc.), anti-phosphorylated (p)-Akt (Ser473)
(1:1,000; cat. no. 9271; Cell Signaling Technology, Inc.), anti-Akt
(1:1,000; cat. no. 9272; Cell Signaling Technology, Inc.),
anti-p-PI3K p85 (Tyr458)/p55 (Tyr199) (1:1,000; cat no. 4228; Cell
Signaling Technology, Inc.), anti-PI3K (1:1,000; cat no. 4292; Cell
Signaling Technology, Inc.) and anti-α-tubulin (1:10,000; cat no.
T5168; Sigma-Aldrich; Merck KGaA). Subsequently, the membranes were
washed 3 times for 5 min with TBST containing 0.05% Tween-20, and
then, incubated with the respective secondary anti-IgG (mouse and
rabbit) antibodies [(1:2,000; cat no. 610-1302; Rockland
Immunochemicals, Inc.) and (1:5,000; cat no. sc-2357; Santa Cruz
Biotechnology, Inc.), respectively], which were purchased from
Bio-Rad Laboratories, Inc. The protein of interest was visualized
using an enhanced chemiluminescence reagent (cat no. SM801-0500;
Gene DireX, Inc.). α-Tubulin was used as the internal control.
Quantifications of blots were performed using an ImageJ software
(v1.8.0; National Institutes of Health).
Enzyme-linked immunosorbent (ELISA)
assay
The concentrations of serum IL-6 (cat. no. KMC0061)
and TNF-α (cat. no. BMS607-3; Invitrogen; Thermo Fisher Scientific,
Inc.) were measured by ELISA according to the manufacturer's
protocols.
Hematoxylin and eosin (H&E)
staining
The gastrocnemius muscle tissues were fixed in 4%
formaldehyde for 48 h at room temperature, and cut into
5-micron-thick sections. Subsequently, the slices were stained with
H&E at room temperature using an automated stainer (Thermo
Fisher Scientific, Inc.). The duration for staining was 10 min for
eosin Y and 5 min for Harris hematoxylin. Images were then captured
using a light microscope before being analyzed by ImageJ software
(v 1.8.0).
Immunohistochemistry (IHC)
The gastrocnemius muscle tissues were fixed in 4%
neutral buffered formaldehyde for 48 h at room temperature. The
fixed muscle tissues were embedded in paraffin and sectioned into
4-µm thick sections. To perform immunohistocemical analysis, the
sections were incubated at 37°C with proteinase K (Invitrogen;
Thermo Fisher Scientific, Inc.) for 30 min for antigen retrieval
and incubated at room temperature with 0.3% hydrogen peroxide in
methanol for 30 min to block endogenous peroxidase/phosphatase
activity. All sections were incubated with blocking reagent (2.5%
normal horse serum; Vector Laboratories, Inc.) at room temperature
for 1 h. The sections were incubated with anti-MyoD (1:500; cat.
no. sc-377460; Santa Cruz Biotechnology, Inc.) and anti-atrogin-1
(1:80,00; cat. no. 67172-1-Ig; ProteinTech Group, Inc.) at 4°C
overnight. The sections were then incubated with the secondary
antibody (biotinylated goat anti-mouse IgG; cat. no. PK-6102 ABC
kit; Vector Laboratories, Inc.) at room temperature for 1 h,
followed by incubation with avidin-biotin reagent (Vector
Laboratories, Inc.) at room temperature for 30 min. After
incubation, all sections were incubated with 1X
3,3-diaminobenzidine tetrahydrochloride (DAB; Thermo Fisher
Scientific, Inc.). The sections were counterstained with Mayer's
hematoxylin for 30 sec at room temperature. At least three randomly
selected image fields per sample were examined under a light
microscope (Olympus Corporation) and imaged.
Cell culture and conditioned medium
collection
LLC cells were seeded at 5×106 cells per
dish of 75 cm2 and cultured to >90% confluence, after
which, their growth medium was removed and replenished with
serum-free DMEM. After 48 h of incubation, the conditioned medium
(CM) was collected into a centrifuge tube and centrifuged at 3,062
× g for 15 min at 4°C. The collected CM was passed through a
syringe filter with 0.2-µm pores (Sartorius AG) and aliquoted for
storage at −20°C until required for further use. Cell cultures were
maintained in a 5% CO2/95% air atmosphere at 37°C.
Differentiation and LLC CM treatment
of C2C12 cells
C2C12 cells seeded at a density of 50,000 cells/well
in a 6-well plate and were incubated in a growth medium (DMEM with
10% FBS and 1% streptomycin-penicillin) until reaching 90–100%
confluence. The growth medium was then replaced by a
differentiation medium [high-glucose DMEM containing 2% horse serum
(Thermo Fisher Scientific, Inc.) and 1% penicillin-streptomycin] to
induce myogenic differentiation for 2 days. Subsequently, the
differentiation medium was replaced by a 1:1 mixture of DMEM
supplemented with 2% horse serum and LLC CM for 3 days in the
presence of BC or THF. The medium was changed every day during the
experiment. All the cellular experiments were performed under 5%
CO2/95% air atmosphere at 37°C.
Myotube length assessment
Cellular differentiation was analyzed by measuring
the myotube length using images captured by a microscope at
magnifications of ×10 or ×20 (Leica, DMI 6000 B; Leica
Microsystems, Inc.). Myotube length (µm) was assessed using ImageJ
(v1.8.0) software.
Cell viability assay
Cell viability was estimated using MTT
(MilliporeSigma) assay. C2C12 myotubes were seeded at a
concentration of 10,000 cells/well and differentiated in 96-well
plates and exposed to LLC CM for 4 days with BC or THF at 37°C. The
media were removed, and 100 µl MTT solution was added to each well
and allowed to incubate for 3 h at 37°C. The plate was analyzed
using a plate reader at 560 nm (Molecular Devices, LLC).
Statistical analysis
All data are expressed as the mean ± standard error
of the mean (SEM). One-way analysis of variance (ANOVA) followed by
the Newman-Keuls post hoc test was conducted using GraphPad PRISM
software (version 3.02; GraphPad Software; Dotmatics). A P-value
threshold of <0.05 was considered to indicate a statistically
significant difference. Each experiment involved at least three
replicates.
Results
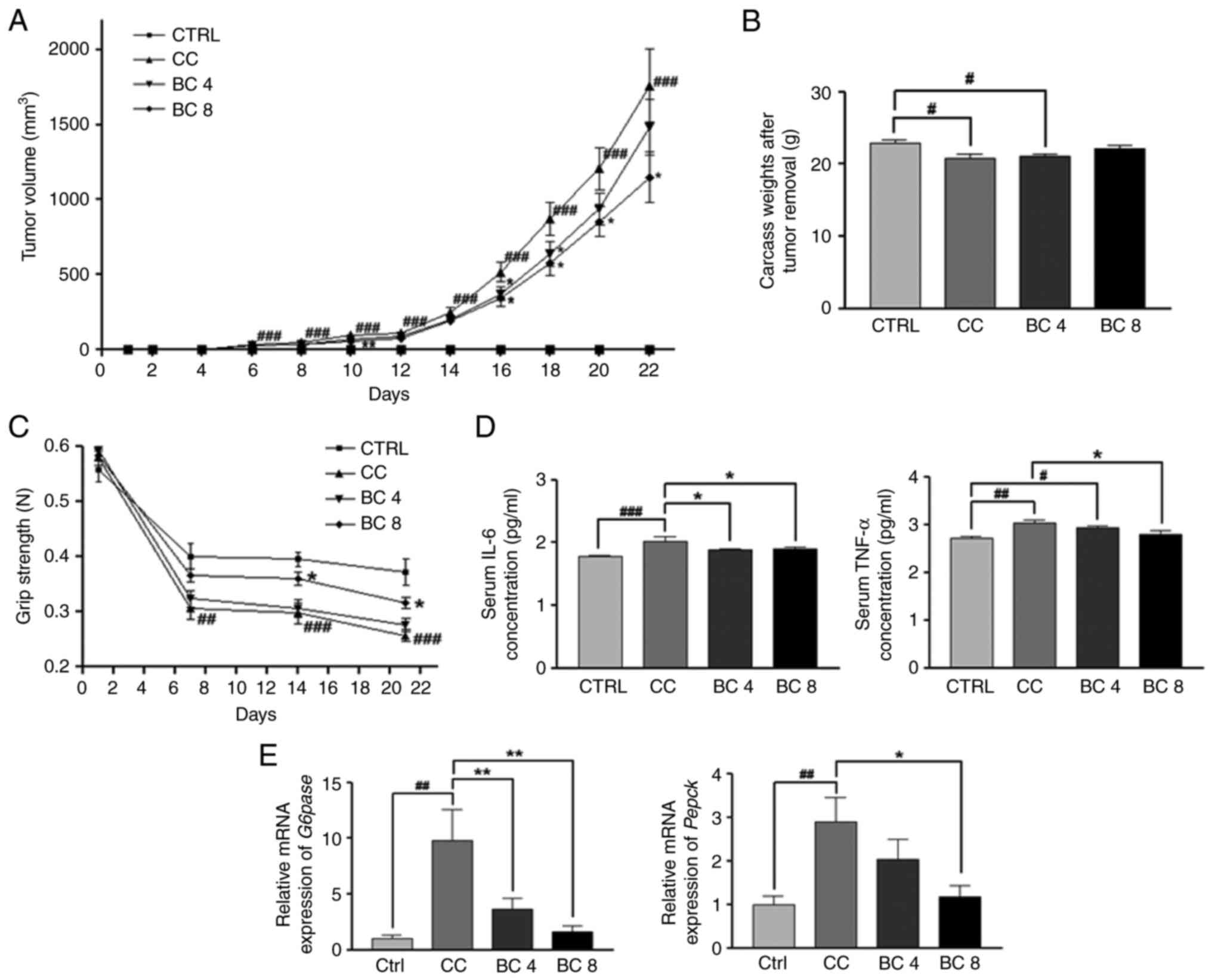
Effects of BC on cachectic progression
in an LLC-induced cancer cachexia mouse model
On days 10, 16, 18, 20 and 22 after cancer cell
inoculation, tumor volumes had decreased significantly in the
BC-supplemented groups compared with the tumor volumes in the CC
group (Fig. 1A). An MTT assay was
then conducted to evaluate the direct effects of BC on the number
of viable LLC cells. After 3 days of treatment, BC induced
significant reductions in the viability of LLC cells, in a
dose-dependent manner. Thus, it can be inferred that BC has direct
antitumor effects on LLC cancer cells (Fig. S1). Carcass weight after tumor
removal (carcass-tumor weight) was significantly decreased in the
CC group compared with the CTRL group (P<0.05). The body weight
changes during the experiment period were measured (Fig. S2A). While the body weights of the
BC 4 and BC 8 groups were significantly higher than the CC group on
day 18, there were no statistically significant differences among
groups on the last day of the experiment. Carcass-tumor weight
tended to be increased by BC supplementations at 4 and 8 mg/kg BW
compared with the CC group, although the levels of statistical
significance were both P>0.05 (Fig.
1B). The weights of the mice spleens, hearts, livers, kidneys
and lungs from the mice were also measured. BC supplementation did
not exert significant effects on the organ weight recovery compared
with the CC group weights (Table
SI). As anorexia is normally accompanied by cachexia, food
intake was measured every two days. However, there were no
significant differences throughout the experimental period among
the groups for food intake (Fig.
S2B).
 | Figure 1.Effects of BC on cachectic
progression in an LLC cancer cachexia mouse model. (A) Tumor
volumes, (B) carcass weights after tumor removal (carcass-tumor
weight) and (C) grip strength were analyzed. (D) Serum IL-6 and
TNF-α levels were assessed using ELISA. (E) mRNA expression of
gluconeogenesis-related genes, Pepck and G6pase in the liver was
determined by reverse trascription-quantitative PCR. Gapdh was used
as a loading control. All data are shown as the mean ± standard
error of the mean and were analyzed using one-way ANOVA with a
Newman-Keuls post hoc test. #P<0.05,
##P<0.01 and ###P<0.001 vs. the CTRL;
*P<0.05 and **P<0.01 vs. CC. BC, β-carotene; LLC, Lewis lung
carcinoma; CTRL, control; CC, LLC-induced cancer cachexia; BC 4,
LLC-induced cancer cachexia + 4 mg/kg BW of β-carotene; BC 8,
LLC-induced cancer cachexia + 8 mg/kg BW of β-carotene; IL-6,
interleukin-6; TNF-α, tumor necrosis factor-α; G6pase, glucose
6-phosphatase; Pepck, phosphoenol-pyruvate carboxykinase. |
Subsequently, the effects of BC on cancer
cachexia-induced muscle and adipose tissue weight losses were
observed. As illustrated in Table
II, LLC tumor inoculation caused significant reductions in the
weights of the gastrocnemius (P<0.001), pectoralis (P<0.01),
triceps (P<0.05) and quadriceps (P<0.001) compared with the
CTRL group. However, the weights in the BC 8 group were
significantly increased for the gastrocnemius muscle (P<0.01),
triceps (P<0.05) and quadriceps (P<0.05) compared with the CC
group. Moreover, the weight of the gastrocnemius muscle in the BC 4
group was also significantly increased (P<0.001) compared with
the CC group. In addition, the weights of the adipose tissues,
including subcutaneous (P<0.001), perirenal (P<0.001),
mesenteric (P<0.01) and epididymal fats (P<0.01) of the CC
group were decreased compared with the CTRL group. By contrast, BC
supplementation at 8 mg/kg BW significantly restored the mesenteric
fat weight loss (P<0.05) compared with the CC group. These
results indicated that BC effectively attenuated cancer
cachexia-related muscle wasting.
| Table II.Effects of BC on various types of fat
and muscle weights in an LLC cancer-cachexia mouse model. |
Table II.
Effects of BC on various types of fat
and muscle weights in an LLC cancer-cachexia mouse model.
| Muscle or fat
tissue | CTRL | CC | BC 4 | BC 8 |
|---|
| Gastrocnemius
muscle (g) | 0.222±0.011 |
0.152±0.010c |
0.205±0.012f |
0.208±0.006e |
| Pectoralis muscle
(g) | 0.055±0.005 |
0.035±0.004b | 0.045±0.003 | 0.043±0.004 |
| Triceps (g) | 0.113±0.010 |
0.085±0.005a | 0.093±0.006 |
0.113±0.008d |
| Quadriceps (g) | 0.153±0.006 |
0.110±0.007c |
0.118±0.007b |
0.135±0.008d |
| Tibialis anterior
muscle (g) | 0.066±0.005 | 0.050±0.005 | 0.058±0.005 | 0.061±0.005 |
| Subcutaneous fat
(g) | 0.245±0.025 |
0.099±0.009c |
0.098±0.011c |
0.105±0.014c |
| Perirenal fat
(g) | 0.107±0.007 |
0.067±0.006c |
0.067±0.005c |
0.085±0.007a |
| Mesenteric fat
(g) | 0.213±0.019 |
0.136±0.011b |
0.158±0.009a |
0.193±0.018d |
| Epididymal fat
(g) | 0.297±0.023 |
0.211±0.017b |
0.222±0.013a | 0.265±0.019 |
| Brown fat (g) | 0.094±0.007 | 0.075±0.004 | 0.076±0.005 | 0.082±0.009 |
To identify whether the increased muscle mass in the
mice fed the BC supplement was related to functional improvement,
the grip strength was analyzed at days 0, 7, 14 and 21 after
inoculating the LLC cells (Fig.
1C). The grip strength in all mice decreased after day 7, an
effect that was considered to be due to the stress caused by
repeated practices. Moreover, compared with the CTRL group, there
was a greater reduction in grip strength in the CC group 7 days
after the LLC cell inoculation (P<0.01 on day 7; P<0.001 on
days 14 and 21). Furthermore, the mice in the BC 8 group
significantly recovered their grip strength on days 14 and 21 after
LLC cell inoculation by 21 and 23%, respectively (P<0.05 for
both).
Serum IL-6 and TNF-α levels were measured using
ELISA (Fig. 1D). The levels of
these proinflammatory cytokines were significantly increased in the
CC group compared with the CTRL group, whereby IL-6 increased by
14.1% (P<0.001) and TNF-α increased by 11.5% (P<0.01).
Nonetheless, supplementation with BC at 4 and 8 mg/kg BW
significantly decreased the IL-6 serum levels by 7.0 and 6.1%,
respectively, compared with those observed in the CC group
(P<0.05 for both). In addition, a BC supplementation of 8 mg/kg
BW effectively decreased the TNF-α serum levels by 7.5% compared
with the CC group (P<0.05).
Hepatic gluconeogenesis regulation by BC in LLC
cancer cachexia mice is depicted in Fig. 1E. The mRNA expression levels of both
G6pase and Pepck were significantly increased in the
CC group compared with the CTRL group (P<0.01 for both,
respectively). In addition, compared with the CC group, the
expression of both genes was significantly downregulated with BC
supplementation at 8 mg/kg BW, by 83.3 and 59.2%, compared with the
CC group (P<0.01 and P<0.05, respectively). The G6pase
mRNA expression was also significantly decreased by 63.1% in the BC
supplementation group of 4 mg/kg BW compared with the CC group
(P<0.01).
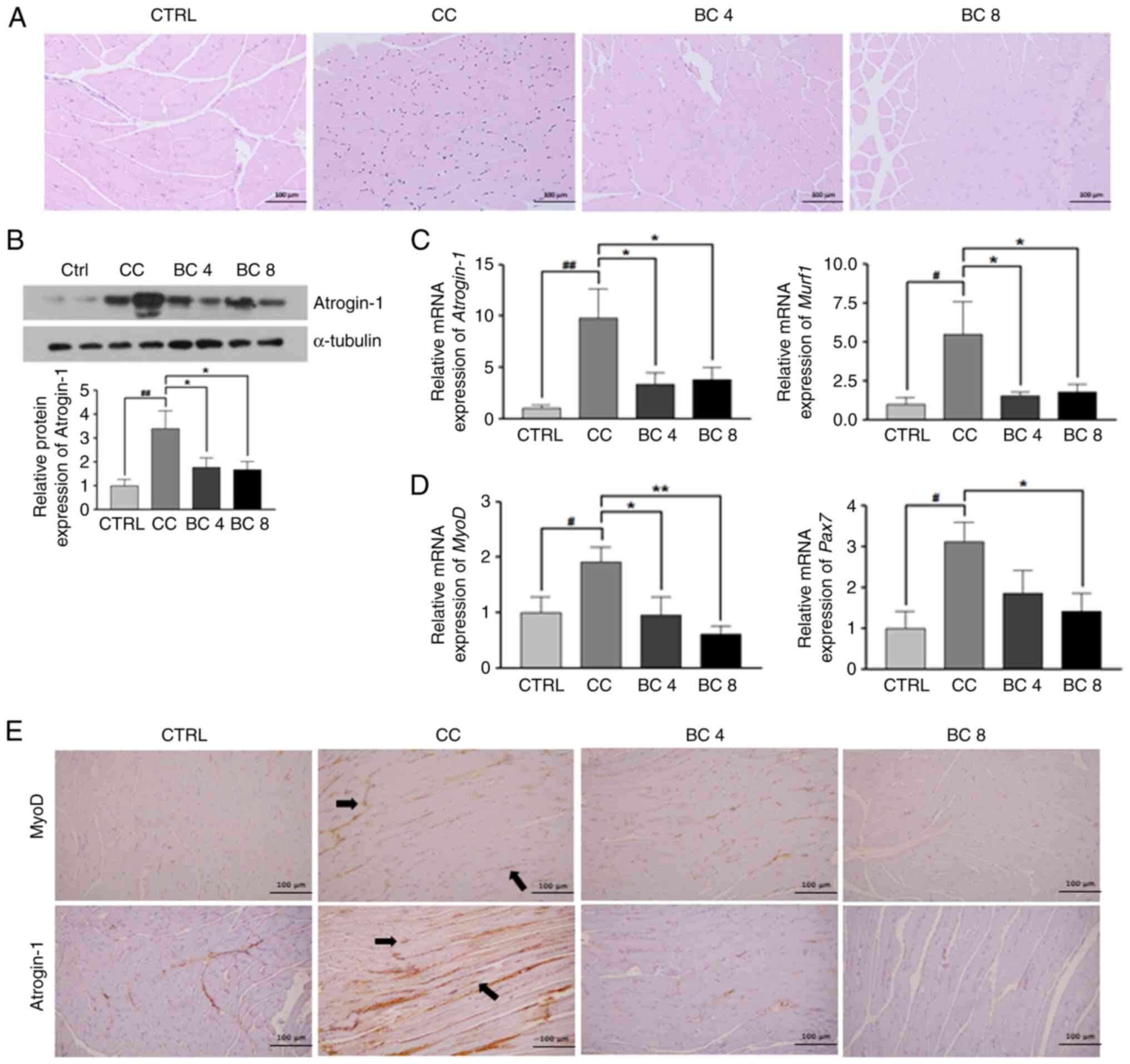
Effects of BC on gastrocnemius muscle
atrophy in an LLC-induced cancer cachexia mouse model
As revealed in Fig.
2A, H&E staining of the gastrocnemius muscles in the CC
group revealed a decrease in the size of the muscle fibers, while
this was restored by BC supplementation. The myofiber size
distribution tended to shift rightward in the BC 8 group (the most
frequent value range was 1,500-2,000 µm2) compared with
the CC group (the most frequent value range was 1,000-1,500
µm2), indicating the myofibers in the BC group were
longer than those in the CC group (data not shown).
 | Figure 2.Effects of BC on gastrocnemius muscle
atrophy in an LLC cancer cachexia mouse model. (A) Representative
images of hematoxylin-eosin stained sections of the gastrocnemius
muscle (magnification, ×100; scale bar, 100 µm) (B) Protein
expression level of atrogin-1 was analyzed by western blotting. (C)
mRNA expression levels of muscle atrophy-related genes, atrogin-1
and Murf1, and (D) muscle stem cell-related genes, MyoD and Pax7 in
gastrocnemius muscle were determined by reverse
trascription-quantitative PCR. Gapdh was used as a loading control.
(E) Representative images of IHC staining of MyoD and atrogin-1 in
gastrocnemius muscle (magnification, ×100). All data are shown as
the mean ± standard error of the mean and were analyzed using
one-way ANOVA with a Newman-Keuls post hoc test.
#P<0.05 and ##P<0.01 vs. the CTRL;
*P<0.05 and **P<0.01 vs. CC. BC, β-carotene; LLC, Lewis lung
carcinoma; CTRL, control; CC, LLC-induced cancer cachexia; BC 4,
LLC-induced cancer cachexia + 4 mg/kg BW of β-carotene; BC 8,
LLC-induced cancer cachexia + 8 mg/kg BW of β-carotene; Murf1,
muscle RING-finger protein-1; MyoD, myoblast determination protein
1; Pax7, paired box 7. |
In Fig. 2B, the
atrogin-1 protein expression in gastrocnemius muscles was
upregulated in the CC group compared with the CTRL group
(P<0.01), which indicated an increase in muscle atrophy.
However, this expression was significantly suppressed by 52.2 and
48.6% in the BC supplementation groups of 4 and 8 mg/kg BW,
respectively, compared with the CC group (both P<0.05).
As demonstrated in Fig.
2C, the atrogin-1 and Murf-1 mRNA levels were
significantly increased in the CC group compared with the CTRL
group (P<0.01 and P<0.05, respectively), while BC
supplementation of 4 or 8 mg/kg BW significantly suppressed these
increases (P<0.05 for both). As revealed in Fig. 2D, mRNA expression of muscle stem
cell markers, MyoD and Pax7 in gastrocnemius muscles
was significantly higher in the CC group compared with the CTRL
group (P<0.05 for both). Conversely, the MyoD mRNA level
was significantly reduced compared with the CTRL level following BC
supplementation of 4 and 8 mg/kg BW (P<0.05 and P<0.01,
respectively), while the Pax7 mRNA level was significantly
downregulated by BC supplementation of 8 mg/kg BW (P<0.05).
In Fig. 2E, the IHC
staining of gastrocnemius muscles revealed that both MyoD and
atrogin-1 were highly overexpressed in the nuclei of the CC group
compared with those of the CTRL group. The expression of MyoD and
atrogin-1, however, was decreased and weakened in the BC 4 and BC 8
groups when compared with the CC group (magnification, ×100).
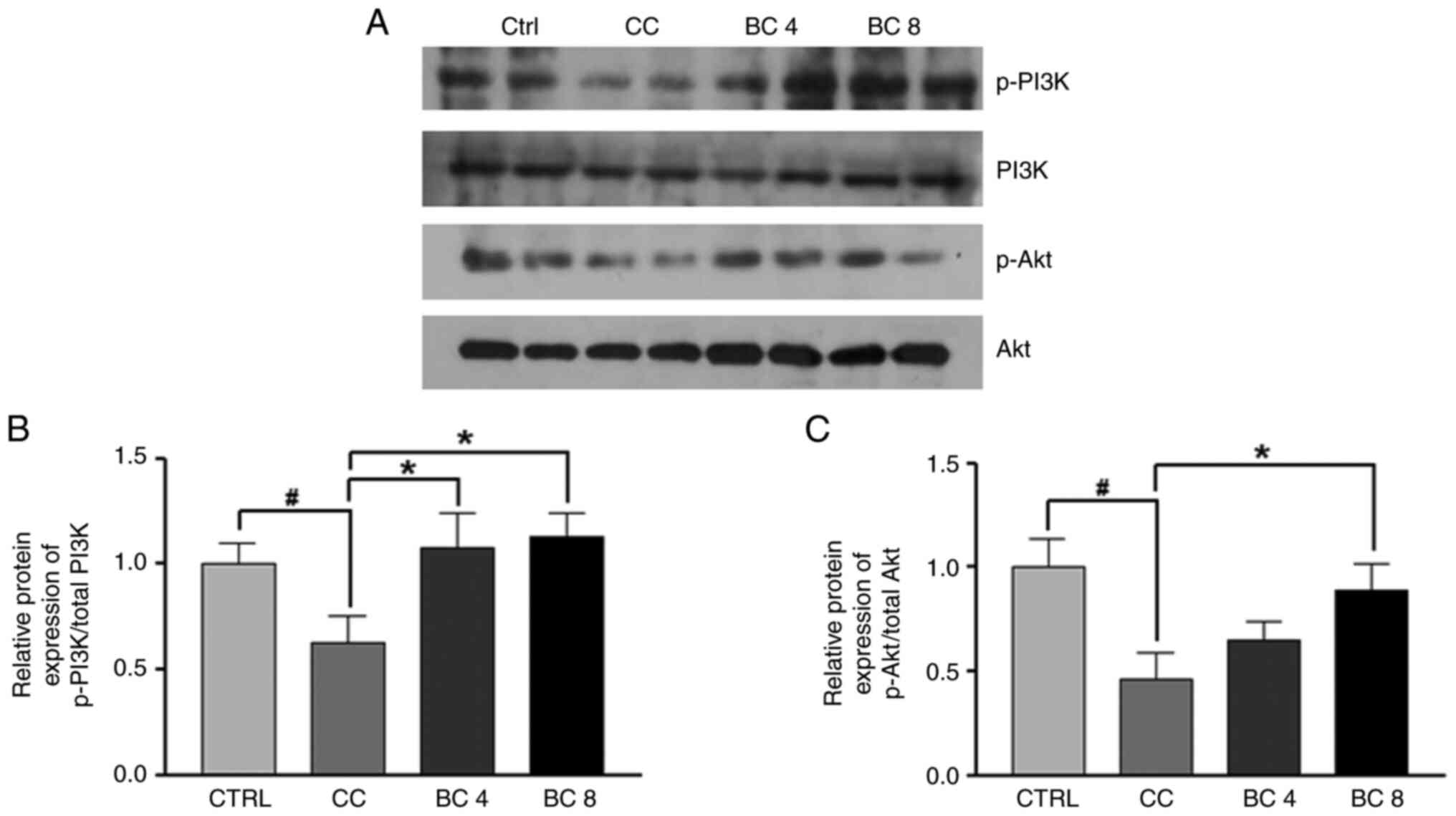
Effects of BC on the PI3K/Akt pathway
in gastrocnemius muscles of LLC-induced cancer cachexia mice
To examine the effects of BC on the PI3K/Akt pathway
in the gastrocnemius muscle in cancer cachexia mice, p-PI3K and
p-Akt protein expression was analyzed. As shown in Fig. 3, the ratio of p-PI3K/PI3K and
p-Akt/Akt in the CC group was lower than that of the CTRL group
(both P<0.05). However, BC supplementation at 4 mg/kg suppressed
the downregulation of phosphorylation of PI3K by 71.9%, whereas
PI3K and Akt phosphorylation was increased in the BC 8 group by
80.4 and 282.7%, respectively (both P<0.05). These results
demonstrated that BC stimulated the PI3K/Akt signaling pathway to
inhibit muscle wasting in the gastrocnemius muscle induced by
cancer cachexia.
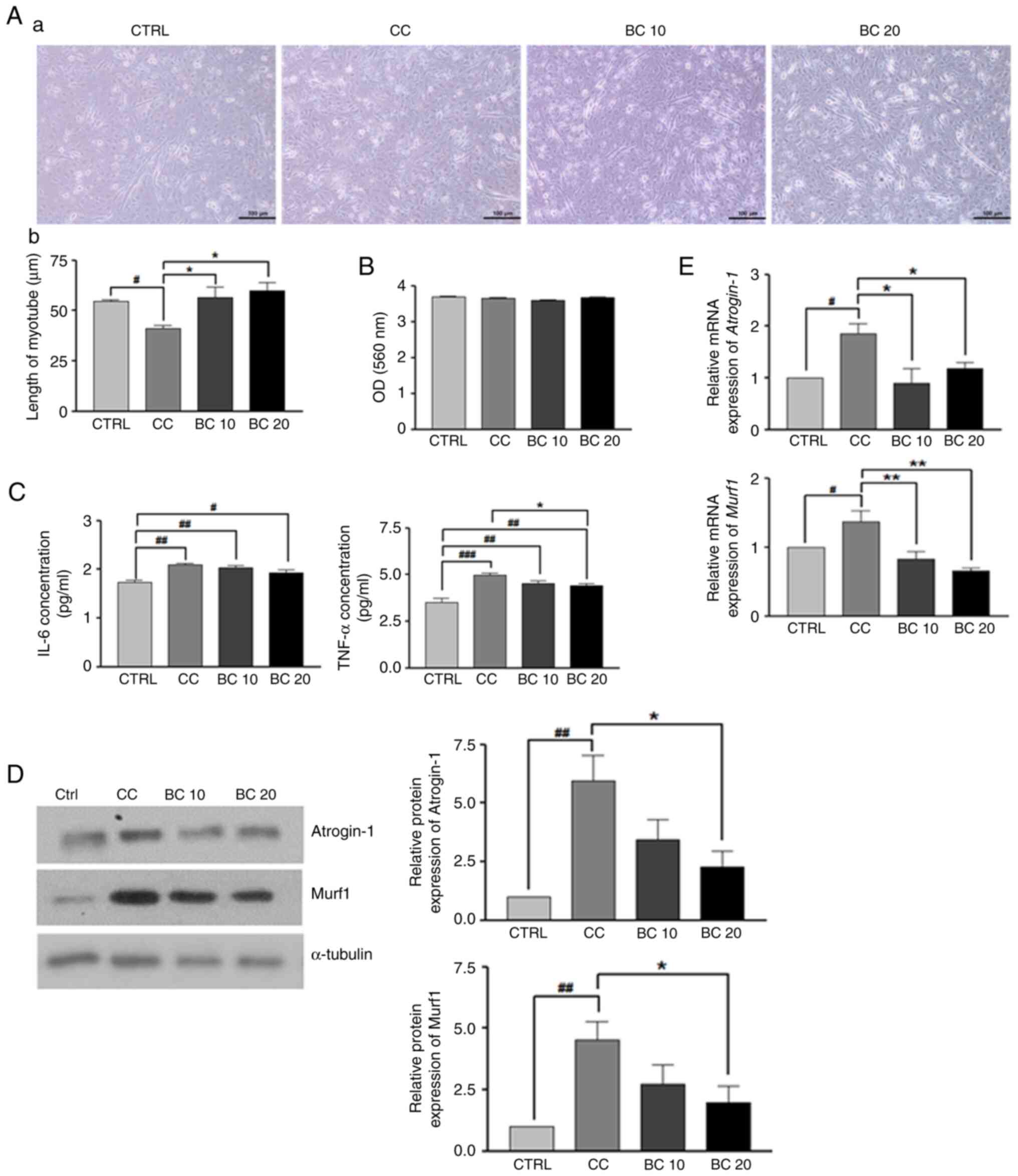
Effects of BC on myogenesis and muscle
atrophy in C2C12 myoblasts treated with LLC CM
As demonstrated in Fig.
4A-a, the addition of LLC CM inhibited the formation of normal
myotubes. Specifically, LLC CM incubation shortened myotubes
(P<0.05), suggesting that LLC CM caused myotube atrophy.
However, treatment with 10 and 20 µM of BC significantly suppressed
myotube length shortening by 38.2 and 46.7%, respectively, compared
with the CC group (P<0.05 for both) (Fig. 4A-b). In Fig. 4B, the absorbance values from the MTT
assay indicated that BC (10 or 20 µM) did not affect the cellular
viability of the myotubes within 3 days of treatment. Moreover, no
cytotoxic effects were observed on the C2C12 myotubes at any of the
BC concentrations up to 20 µM.
 | Figure 4.Effects of BC on myogenesis and
muscle atrophy in C2C12 myotubes treated with LLC CM. (A-a) Cells
were observed under a microscope. (A-b) Length of myotube as formed
by differentiated C2C12 cells. (B) Cell viability in C2C12 myotubes
treated with LLC CM was evaluated with 10 and 20 µM BC for 3 days
using an MTT assay. (C) Secreted IL-6 and TNF-α levels in the media
were determined. (D) Protein levels of atrogin-1 and MuRF1 were
measured by western blotting and the relative band intensities were
calculated after normalization to a-tubulin expression. (E) The
mRNA levels of atrogin-1 and Murf1 were measured by reverse
trascription-quantitative PCR. Gapdh was used as a loading control.
All data are shown as the mean ± standard error of the mean and
were analyzed using one-way ANOVA with a Newman-Keuls post hoc
test. #P<0.05, ##P<0.01, and
###P<0.001 vs. the CTRL; *P<0.05 and **P<0.01
vs. CC. BC, β-carotene; LLC, Lewis lung carcinoma; CM, conditioned
medium; CTRL, Control; CC, Cancer-cachexia; BC 10, 10 µM of BC; BC
20, 20 µM of BC; Murf1, muscle RING-finger protein-1; IL-6,
interleukin-6; TNF-α, tumor necrosis factor-α. |
The culture medium was isolated to measure the IL-6
and TNF-α levels. As revealed in Fig.
4C, the secretion of IL-6 and TNF-α was significantly increased
by 20.7 (P<0.01) and 41.4%, respectively in the CC group
(P<0.001) compared with the CTRL group. Interestingly, while the
IL-6 level tended to be restored by BC treatments, albeit without
statistical significance, the elevated TNF-α level caused by LLC CM
incubation was significantly suppressed by 20 µM BC
(P<0.05).
Next, the regulatory roles of BC on the levels of
muscle atrophy-related markers in C2C12 myotubes, while incubated
with LLC CM, were analyzed. As revealed in Fig. 4D, C2C12 myotubes treated with LLC CM
exhibited an upregulation of protein expression levels of the
muscle atrophy-related markers atrogin-1 and Murf1 compared with
the CTRL group (P<0.05 for both). However, these expression
levels were suppressed by treatment with BC at 20 µM (P<0.05).
In Fig. 4E, LLC CM treatment also
significantly upregulated atrogin-1 and Murf1 mRNA
expression levels compared with the CTRL group (P<0.05 for
both). However, these expression levels were significantly
downregulated by treatment with BC at 10 and 20 µM (P<0.05 for
atrogin-1 and P<0.01 for Murf1). These data
indicated that BC could restore the LLC CM-induced dysregulations
of myogenesis, muscle atrophy and proinflammatory cytokines in
C2C12 myotubes.
In addition, the protein expression levels of
atrogin-1 and Murf1 in C2C12 myotubes treated with BC at 20 µM were
evaluated in the presence or absence of LLC CM. As a result,
treatment with BC at 20 µM in the absence of LLC CM tended to
suppress the expression levels of Murf1 and atrogin-1 compared with
the CTRL group, but they were not statistically significant
(P>0.05 for both). Nevertheless, as already described in the
present study, LLC CM-induced upregulation of atrogin-1 and Murf1
compared with the CTRL group were significantly suppressed by 20 µM
BC treatment (P<0.01 for atrogin-1 and P<0.05 for
Murf1). These results indicated that BC is only effective in
inhibiting the muscle atrophy in C2C12 myotubes with LLC CM
incubation (Fig. S3).
Discussion
The present study identified the protective effects
of BC on cancer cachexia-induced muscle wasting both in
vitro and in vivo. BC supplementation prevented the loss
of muscle mass and grip strength in the LLC cell-bearing cancer
cachexia mouse model. Moreover, BC administration also suppressed
muscle atrophy by regulating the PI3K/Akt pathway and muscle
stemness. In vitro analysis confirmed that LLC CM caused
reductions in myotube length and myogenesis and increased muscle
atrophy and inflammatory cytokine secretions compared with the CTRL
group. However, these alterations were effectively restored by
treatments with BC.
LLC cells were used to induce cancer cachexia both
in vitro and in vivo (33–35).
Consistent with the model of the present study, a previous study
revealed that LLC cell injection effectively induced cancer
cachexia with muscle wasting in mice (36). Moreover, LLC CM caused a reduction
in C2C12 myotube size and the upregulation of muscle atrophy
markers, atrogin-1 and MuRF1 (37,38).
Since overgrown and large tumors can cause great
pain in mice, the mice were sacrificed before the average length of
their tumor reached 2 cm in accordance with IACUC regulations.
Moreover, based on the preliminary study results, 22 days of
1×106 LLC cell inoculation could successfully induce
cancer cachexia phenotypes, such as muscle atrophy and adipose
depletion. Therefore, mice were sacrificed 22 days after cancer
cell inoculation in the present study.
While a previous study investigating the effects of
BC on muscle metabolism mainly focused on the soleus muscle
(39), the gastrocnemius muscle was
the primary focus in the present study because it represents the
most common skeletal muscle associated with cachexia. Additionally,
it is more involved in cancer cachexia pathogenesis, such as
myogenesis and muscle atrophy, than the soleus muscle. Myogenic
transcription and muscle-specific E3 ubiquitin ligases, which are
important in regulating cancer cachexia, were more prominently
expressed in the gastrocnemius muscle than in soleus muscles after
nerve injury (40). Muscle atrophy,
which was accompanied by a reduction in muscle fiber size and
upregulation of MuRF1 and atrogin-1 expression levels, was also
more predominant in the gastrocnemius muscle than in the soleus
muscle after injury (40).
Similar to the finding that supplementation with BC
attenuated the LLC-induced cancer cachexia by downregulating the
expression levels of atrophy markers (atrogin-1 and MuRF1), it was
previously reported that BC attenuated soleus muscle loss by
suppressing the expression levels of atrogin-1 and
Murf1 against denervation-induced muscle atrophy (24). BC also reportedly promoted protein
synthesis in the soleus muscles and suppressed ubiquitin conjugates
under normal conditions, however, it did not regulate mRNA
expression of the atrogenes, atrogin-1 and Murf1
(28). Since the muscle-specific E3
ubiquitin ligases, atrogin-1 and MuRF1, are regulated by BC, these
findings suggest that BC exerts protective effects on muscle
wasting in cancer cachexia through the ubiquitin-proteasome
pathway.
However, in contrast to the findings in the present
study, the previous studies showed that BC administration was only
effective in increasing the mass of the soleus muscle against the
denervation (24) and under
physiological conditions (28),
whereas no effect was observed in the gastrocnemius muscle. In
addition, BC treatment inhibited the denervation-induced
upregulation of ubiquitin conjugates in the soleus muscle, yet not
in the gastrocnemius muscle (24).
The gastrocnemius and soleus have distinct anatomical and
physiological features, whereby the gastrocnemius is mainly
composed of fast twitch muscle fibers, whereas the soleus muscle
mostly comprises slow twitch muscle fibers (41). In contrast to the gastrocnemius,
there is also less risk of injury in the soleus muscle, while
should injury occur, soleus injuries have a tendency of being less
severe in clinical presentation and less acute compared with
gastrocnemius injuries (42). Thus,
it is assumed that the gastrocnemius was more affected by cancer
cachexia, which is a devastating muscle-wasting disease than the
soleus. Similar to the findings of the present study, previous
studies have reported that fast twitch muscle fibers, such as the
gastrocnemius muscles, were selectively targeted in the cancer
cachexia rodent model (43,44).
TNF-α, IL-1 and IL-6 are major proinflammatory
cytokines that are released from immune or cancer cells and promote
muscle loss in patients with cachexia (45). The present study also demonstrated
that BC effectively suppresses serum cytokines (TNF-α and IL-6) in
cancer cachexia, implying that BC may have effects on immune or
cancer cells, and their interactions with muscle tissues. The
ability of BC to modulate the immune system and attenuate
inflammatory diseases has been well studied (46). Previously, it was reported that
treatments with BC significantly reduced the production of the
proinflammatory cytokines, TNF-α and IL-6, following induction by
lipopolysaccharides in macrophages (47). Pretreatment with BC significantly
decreased cytokines levels of TNF-α and IL-6 in the kidneys of
bromobenzene-induced rats (48).
Additionally, another study reported that subjects with lower
plasma BC levels exhibited increased IL-6 levels compared with
healthy children (49).
As a result of muscle depletion in cachexia or
muscle wasting, amino acids are released from skeletal muscles and
function as precursors for hepatic gluconeogenesis (10,50).
In addition, the upregulation of gluconeogenesis also results in a
low concentration of skeletal muscle amino acids, leading to muscle
wasting. Thus, this aggressive feedback loop worsens the disease
(51). During muscle atrophy, the
activity of the glycolytic enzymes is increased in the muscle,
while gluconeogenesis is increased in the liver (52). Hepatic gluconeogenesis is also
affected by inflammatory cytokines, such as TNF-α, IL-6 and IL-1.
Production of inflammatory proteins, such as acute-phase proteins,
in the liver also requires muscle deterioration as a source of
amino acids in cancer cachexia (11,53).
In the present study, treatment with BC suppressed IL-6 and TNF-α
serum levels, which may inhibit hepatic gluconeogenesis. Thus,
further investigation into the hepatic levels of inflammatory
cytokines and other related mechanisms are required in future
studies.
In the present study, the PI3K/Akt signaling pathway
was suppressed by cancer cachexia. It was previously reported that
myotubes become hypertrophic via the PI3K/Akt pathway, which
increases protein synthesis and decreases the expression of the
muscle atrophy-related markers, MAFbx and MuRF1 (54,55).
BC supplementation was observed to upregulate the PI3K/Akt
signaling pathway in cancer cachexia-induced mouse gastrocnemius
muscles. Oral BC administration enhanced skeletal muscle mass by
upregulating IGF-1 (28), which is
known to activate the PI3K/Akt signaling pathway (54).
The mRNA levels of the muscle stem cell markers,
Pax7 and MyoD, were upregulated following the
induction of cancer cachexia; however, these levels were recovered
after treatment with BC. Further stem cell-based regenerative
treatment was used to restore the muscle homeostasis following
injury (56). The muscle satellite
cells were activated to participate in regenerative procedures
following cancer cachexia-associated muscle damage (13). This implies that the upregulation of
the stem cell markers caused by cancer cachexia was a consequence
of the feedback system to counteract muscle wasting. A previous
study reported that BC inhibited proliferation and promoted the
differentiation of Pax7-enriched chicken myoblasts via b-carotene
oxygenase 1 (29), thereby
supporting the findings of the present study. Additionally, BC
supplementation suppressed cancer stemness in colon cancer
(57) and neuroblastoma (58,59).
In the present study, BC effectively restored LLC
CM-induced myogenesis inhibition, while it has also been reported
in a previous study regarding neuroblastoma, that BC induced
neuronal cell differentiation in SK-N-BE(2)C neuroblastoma cells
(59). Retinoic acid (RA), produced
by cleavage of BC, also reportedly upregulates the differentiation
of embryonic and cancer stem cells (60). The present study revealed that
treatments with BC downregulated atrogin-1 and Murf1
levels against LLC CM-induced wasting in C2C12 myotubes. Similar to
the results of the present study, treatments with BC decreased
hydrogen peroxide-induced increases in atrogin-1 and
Murf1 levels in C2C12 myotubes (24). BC was also converted into RA, which
suppressed proliferation and increased the differentiation of
chicken myoblasts (29). Treatments
with BC have been shown to alleviate
H2O2-induced ubiquitin ligases of
atrogin-1 and Murf1, dose-dependently in C2C12
myotube cells (24).
For in vivo analysis, mice were treated with
BC supplements of 4 and 8 mg/kg BW, administered twice weekly, for
22 days. These doses corresponded to 1.12 and 2.24 mg/kg BW per
day, respectively, and can be converted to 5.44 and 10.88 mg/day
for a 60-kg person (61). As
orange-colored carrots (Daucus carota L.) contain BC, in
quantities of 7 to 17 mg/100 g (62), the current doses used in the present
study were physiological. At 10 and 20 µM, BC treatments did not
result in significant toxic effects in differentiated C2C12
cells.
Recently, it was reported that relatively low doses
of BC (0.5 and 1 µM for in vitro experiments and 0.5 and 2
mg/kg BW for in vivo experiments) were effective in
restoring adipose wasting in cancer cachexia induced by CT26 cancer
cell inoculation (63). However, in
the present study, relatively high doses of BC (10 and 20 µM for
in vitro experiments and 4 and 8 mg/kg BW for in vivo
experiments) restored muscle wasting-associated dysregulations in
cancer cachexia. Likewise, it was observed that BC plays a
preventive role against cancer cachexia at different concentration
ranges depending on the model. This suggests that relatively low BC
concentrations were effective in improving the recovery of a
patient from early cancer cachexia-induced impairments. CT26 and
LLC cancer cells are known to induce different molecular mechanisms
causing cancer cachexia (64). A
combination treatment of exercise training and erythropoietin
appeared to have different anticancer cachexia effects in CT26- and
LLC-induced mouse models, respectively (65). The precise underlying mechanism by
which BC provides protective effects in each of the CT26- and
LLC-induced cancer cachexia models and with their respective
concentration ranges should be studied further.
A limitation of the present study was that the
protein expression of Murf1 was only measured in in vitro
but not in vivo experiments. Examination of the protein
expression of Murf1 in the cancer cachexia mouse model was
attempted. In spite of numerous attempts and troubleshooting, the
uniformly distributed high background in the western blot films
continued to appear. It was hypothesized that this was due to the
non-specific bindings of the proteins with the primary or secondary
antibodies. Thus, since it was difficult to draw reliable
conclusions from the blurred and unclear western Murf1 bands from
the mouse samples, the data could not be presented.
In conclusion, the present study determined that BC
supplementation can alleviate muscle wasting in cancer cachexia by
regulating myogenesis and muscle atrophy dysregulations in
gastrocnemius muscles. These effects may be mediated by the
regulation of PI3K/Akt phosphorylation. BC supplementation also
effectively downregulated the increased hepatic gluconeogenesis and
systemic inflammation caused by cancer cachexia. Cachexia also
occurs in several diseases, including heart failure, kidney disease
and chronic obstructive pulmonary disease (66). Thus, the present study provides
insights into the possible roles of BC as a novel therapeutic agent
for other diseases associated with muscle wasting.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by the Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education (grant no.
NRF-2019R1F1A1059287), the NRF Grant funded by the Korean
Government (NRF-2019-Global Ph.D. Fellowship Program), the BK21
Fostering Outstanding Universities for Research (FOUR) funded by
the Ministry of Education (MOE, Korea) and the National Research
Foundation of Korea (grant no. NRF-5199990614253; Education
Research Center for 4IR-Based Health Care).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YuK contributed to the conception, design of the
study, and acquisition of funding. YeK, YO, YSK, JHS, YSL and YuK
analyzed and interpreted the data, and wrote and reviewed the
manuscript. YeK, YO and YSL performed the experiments. YSK, JHS,
and YuK provided technical support to perform the experiments. YeK
and YuK confirm the authenticity of all the raw data. All authors
have read and agreed to the published version of the
manuscript.
Ethics approval and consent to
participate
The present study was conducted according to the
guidelines of the National Institutes of Health (NIH publication
no. 8023, revised 1978), and approved (IACUC approval no. EWHA
IACUC 21-003-1) by the Institutional Animal Care and Use Committee
of Ewha Womans University (Seoul, Republic of Korea).
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fearon K, Strasser F, Anker SD, Bosaeus I,
Bruera E, Fainsinger RL, Jatoi A, Loprinzi C, MacDonald N,
Mantovani G, et al: Definition and classification of cancer
cachexia: An international consensus. Lancet Oncol. 12:489–495.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Evans WJ, Morley JE, Argilés J, Bales C,
Baracos V, Guttridge D, Jatoi A, Kalantar-Zadeh K, Lochs H,
Mantovani G, et al: Cachexia: A new definition. Clin Nutr.
27:793–799. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Armstrong VS, Fitzgerald LW and Bathe OF:
Cancer-associated muscle wasting-candidate mechanisms and molecular
pathways. Int J Mol Sci. 21:92682020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brown JL, Lee DE, Rosa-Caldwell ME, Brown
LA, Perry RA, Haynie WS, Huseman K, Sataranatarajan K, Van Remmen
H, Washington TA, et al: Protein imbalance in the development of
skeletal muscle wasting in tumour-bearing mice. J Cachexia
Sarcopenia Muscle. 9:987–1002. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Burckart K, Beca S, Urban RJ and
Sheffield-Moore M: Pathogenesis of muscle wasting in cancer
cachexia: Targeted anabolic and anti-catabolic therapies. Curr Opin
Clin Nutr Metab Care. 13:4102010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yuan L, Han J, Meng Q, Xi Q, Zhuang Q,
Jiang Y, Han Y, Zhang B, Fang J and Wu G: Muscle-specific E3
ubiquitin ligases are involved in muscle atrophy of cancer
cachexia: An in vitro and in vivo study. Oncol Rep.
33:2261–2268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adams V, Gußen V, Zozulya S, Cruz A,
Moriscot A, Linke A and Labeit S: Small-molecule chemical knockdown
of MuRF1 in melanoma bearing mice attenuates tumor cachexia
associated myopathy. Cells. 9:22722020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cole CL, Kleckner IR, Jatoi A, Schwarz EM
and Dunne RF: The role of systemic inflammation in
cancer-associated muscle wasting and rationale for exercise as a
therapeutic intervention. JCSM Clin Rep. 3:e000652018.PubMed/NCBI
|
|
9
|
Webster JM, Kempen LJ, Hardy RS and Langen
RC: Inflammation and skeletal muscle wasting during cachexia. Front
physiol. 11:5976752020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rohm M, Zeigerer A, Machado J and Herzig
S: Energy metabolism in cachexia. EMBO Rep. 20:e472582019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bonetto A, Aydogdu T, Kunzevitzky N,
Guttridge DC, Khuri S, Koniaris LG and Zimmers TA: STAT3 activation
in skeletal muscle links muscle wasting and the acute phase
response in cancer cachexia. PLoS One. 6:e225382011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gauldie J, Richards C and Baumann H: IL6
and the acute phase reaction. Res Immunol. 143:755–758. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He WA, Berardi E, Cardillo VM, Acharyya S,
Aulino P, Thomas-Ahner J, Wang J, Bloomston M, Muscarella P, Nau P,
et al: NF-κB-mediated Pax7 dysregulation in the muscle
microenvironment promotes cancer cachexia. J Clin Investig.
123:4821–4835. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zammit PS, Golding JP, Nagata Y, Hudon V,
Partridge TA and Beauchamp JR: Muscle satellite cells adopt
divergent fates: A mechanism for self-renewal? J Cell Biol.
166:347–357. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Megeney LA, Kablar B, Garrett K, Anderson
JE and Rudnicki MA: MyoD is required for myogenic stem cell
function in adult skeletal muscle. Genes Dev. 10:1173–1183. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schiaffino S and Mammucari C: Regulation
of skeletal muscle growth by the IGF1-Akt/PKB pathway: Insights
from genetic models. Skelet Muscle. 1:42011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang W, Huang J, Wu H, Wang Y, Du Z, Ling
Y, Wang W, Wu Q and Gao W: Molecular mechanisms of cancer
cachexia-induced muscle atrophy. Mol Med Rep. 22:4967–4980. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Quan-Jun Y, Yan H, Yong-Long H, Li-Li W,
Jie L, Jin-Lu H, Jin L, Peng-Guo C, Run G and Cheng G: Selumetinib
attenuates skeletal muscle wasting in murine cachexia model through
ERK inhibition and AKT activation. Mol Cancer Ther. 16:334–343.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Toti E, Chen CO, Palmery M, Villaño
Valencia D and Peluso I: Non-provitamin A and provitamin A
carotenoids as immunomodulators: recommended dietary allowance
therapeutic index, or personalized nutrition? Oxid Med Cell Longev.
2018.46378612018.PubMed/NCBI
|
|
20
|
Huang J, Weinstein SJ, Yu K, Männistö S
and Albanes D: Serum beta carotene and overall and cause-specific
mortality: A prospective cohort study. Circul Res. 123:1339–1349.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ardite E, Barbera JA, Roca J and
Fernández-Checa JC: Glutathione depletion impairs myogenic
differentiation of murine skeletal muscle C2C12 cells through
sustained NF-κB activation. Am. J Pathol. 165:719–728.
2004.PubMed/NCBI
|
|
22
|
Powers SK, Smuder A and Judge A: Oxidative
stress and disuse muscle atrophy: Cause or consequence? Curr Opin
Clin Nutr Metab Care. 15:2402012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen QH, Wu BK, Pan D, Sang LX and Chang
B: Beta-carotene and its protective effect on gastric cancer. World
J Clin Cases. 9:65912021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ogawa M, Kariya Y, Kitakaze T, Yamaji R,
Harada N, Sakamoto T, Hosotani K, Nakano Y and Inui H: The
preventive effect of β-carotene on denervation-induced soleus
muscle atrophy in mice. Br J Nutr. 109:1349–1358. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawamura A, Aoi W, Abe R, Kobayashi Y,
Wada S, Kuwahata M and Higashi A: Combined intake of astaxanthin,
β-carotene, and resveratrol elevates protein synthesis during
muscle hypertrophy in mice. Nutrition. 69:1105612020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lauretani F, Semba RD, Bandinelli S,
Dayhoff-Brannigan M, Giacomini V, Corsi AM, Guralnik JM and
Ferrucci L: Low plasma carotenoids and skeletal muscle strength
decline over 6 years. J Gerontol A Biol Sci Med Sci J. 63:376–383.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alipanah N, Varadhan R, Sun K, Ferrucci L,
Fried L and Semba RD: Low serum carotenoids are associated with a
decline in walking speed in older women. J Nutr Health Aging.
13:170–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kitakaze T, Harada N, Imagita H and Yamaji
R: β-Carotene increases muscle mass and hypertrophy in the soleus
muscle in mice. J Nutr Sci Vitaminol. 61:481–487. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Praud C, Al Ahmadieh S, Voldoire E, Le
Vern Y, Godet E, Couroussé N, Graulet B, Le Bihan Duval E, Berri C
and Duclos MJ: Beta-carotene preferentially regulates chicken
myoblast proliferation withdrawal and differentiation commitment
via BCO1 activity and retinoic acid production. Exp Cell Res.
358:140–146. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Robboy MS, Sato AS and Schwabe AD: The
hypercarotenemia in anorexia nervosa: A comparison of vitamin A and
carotene levels in various forms of menstrual dysfunction and
cachexia. Am J Clin Nutr. 27:362–367. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Carlsson G, Gullberg B and Hafström L:
Estimation of liver tumor volume using different formulas-an
experimental study in rats. J Cancer Res Clin Oncol. 105:20–23.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Geppert J, Walth AA, Terrón Expósito R,
Kaltenecker D, Morigny P, Machado J, Becker M, Simoes E, Lima JDCC,
Daniel C, et al: Aging aggravates cachexia in tumor-bearing mice.
Cancers. 14:902021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hain BA, Xu H, VanCleave AM, Gordon BS,
Kimball SR and Waning DL: REDD1 deletion attenuates cancer cachexia
in mice. J Appl Physiol. 131:1718–1730. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao S and Carson JA: Lewis lung carcinoma
regulation of mechanical stretch-induced protein synthesis in
cultured myotubes. Am J Physiol Cell Physiol. 310:C66–C79. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun R, Zhang S, Lu X, Hu W, Lou N, Zhao Y,
Zhou J, Zhang X and Yang H: Comparative molecular analysis of early
and late cancer cachexia-induced muscle wasting in mouse models.
Oncol Rep. 36:3291–3302. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chiappalupi S, Sorci G, Vukasinovic A,
Salvadori L, Sagheddu R, Coletti D, Renga G, Romani L, Donato R and
Riuzzi F: Targeting RAGE prevents muscle wasting and prolongs
survival in cancer cachexia. J Cachexia Sarcopenia Muscle.
11:929–946. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang G, Jin B and Li YP: C/EBPβ mediates
tumour-induced ubiquitin ligase atrogin1/MAFbx upregulation and
muscle wasting. EMBO J. 30:4323–4335. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang J, Zheng J, Chen H, Li X, Ye C,
Zhang F, Zhang Z, Yao Q and Guo Y: Curcumin targeting
NF-κB/ubiquitin-proteasome-system axis ameliorates muscle atrophy
in triple-negative breast cancer cachexia mice. Mediators Inflamm.
2022:25671502022.PubMed/NCBI
|
|
40
|
Wiberg R, Jonsson S, Novikova LN and
Kingham PJ: Investigation of the expression of myogenic
transcription factors, microRNAs and muscle-specific E3 ubiquitin
ligases in the medial gastrocnemius and soleus muscles following
peripheral nerve injury. PLoS One. 10:e01426992015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ro A, Kageyama N and Mukai T:
Pathophysiology of venous thromboembolism with respect to the
anatomical features of the deep veins of lower limbs: A review. Ann
Vasc Dis. 10:99–106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bryan Dixon J: Gastrocnemius vs. soleus
strain: How to differentiate and deal with calf muscle injuries.
Curr Rev Musculoskelet Med. 2:74–77. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baracos VE, DeVivo C, Hoyle D and Goldberg
AL: Activation of the ATP-ubiquitin-proteasome pathway in skeletal
muscle of cachectic rats bearing a hepatoma. Am J Physiol.
268:E996–E1006. 1995.PubMed/NCBI
|
|
44
|
Acharyya S, Butchbach ME, Sahenk Z, Wang
H, Saji M, Carathers M, Ringel MD, Skipworth RJ, Fearon KC,
Hollingsworth MA, et al: Dystrophin glycoprotein complex
dysfunction: A regulatory link between muscular dystrophy and
cancer cachexia. Cancer Cell. 8:421–432. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fearon KC, Glass DJ and Guttridge DC:
Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell
Metab. 16:153–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chew BP: Vitamin A and β-carotene on host
defense. J Dairy Sci. 70:2732–2743. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li R, Hong P and Zheng X: β-Carotene
attenuates lipopolysaccharide-induced inflammation via inhibition
of the NF-κB, JAK2/STAT3 and JNK/p38 MAPK signaling pathways in
macrophages. Anim Sci J. 90:140–148. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Akkara PJ and Sabina EP: Pre-treatment
with beta carotene gives protection against nephrotoxicity induced
by bromobenzene via modulation of antioxidant system,
pro-inflammatory cytokines and pro-apoptotic factors. Appl Biochem
Biotechnol. 190:616–633. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rodríguez-Rodríguez E, López-Sobaler AM,
Navia B, Andrés P, Jiménez-Ortega AI and Ortega RM: β-Carotene
concentration and its association with inflammatory biomarkers in
Spanish schoolchildren. Ann Nutr Metab. 71:80–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rui L: Energy metabolism in the liver.
Compr Physiol. 4:1772014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kumar R, Prakash SS, Priyadarshi RN and
Anand U: Sarcopenia in chronic liver disease: A metabolic
perspective. J Clin Transl Hepatol. 10:1213–1222. 2022.PubMed/NCBI
|
|
52
|
Stein T and Wade C: Metabolic consequences
of muscle disuse atrophy. J Nutr. 135:1824S–1828S. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Poulia KA, Sarantis P, Antoniadou D,
Koustas E, Papadimitropoulou A, Papavassiliou AG and Karamouzis MV:
Pancreatic cancer and cachexia-metabolic mechanisms and novel
insights. Nutrients. 12:15432020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Stitt TN, Drujan D, Clarke BA, Panaro F,
Timofeyva Y, Kline WO, Gonzalez M, Yancopoulos GD and Glass DJ: The
IGF-1/PI3K/Akt pathway prevents expression of muscle
atrophy-induced ubiquitin ligases by inhibiting FOXO transcription
factors. Mol Cell. 14:395–403. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rodriguez J, Vernus B, Chelh I,
Cassar-Malek I, Gabillard JC, Hadj Sassi A, Seiliez I, Picard B and
Bonnieu A: Myostatin and the skeletal muscle atrophy and
hypertrophy signaling pathways. Cell Mol Life Sci. 71:4361–4371.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhu H, Lin X and Diao Y: Function and
regulation of muscle stem cells in skeletal muscle development and
regeneration: A narrative review. J bio-X Res. 4:89–96. 2021.
|
|
57
|
Lee KE, Kwon M, Kim YS and Kim Y, Chung
MG, Heo SC and Kim Y: β-carotene regulates cancer stemness in colon
cancer in vivo and in vitro. Nutr Res Pract. 16:161–172. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lim JY, Kim YS, Kim KM, Min SJ and Kim Y:
Beta-carotene inhibits neuroblastoma tumorigenesis by regulating
cell differentiation and cancer cell stemness. Biochem Biophys Res
Commun. 450:1475–1480. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee HA, Park S and Kim Y: Effect of
beta-carotene on cancer cell stemness and differentiation in
SK-N-BE(2)C neuroblastoma cells. Oncol Rep. 30:1869–1877. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tighe AP and Gudas LJ: Retinoic acid
inhibits leukemia inhibitory factor signaling pathways in mouse
embryonic stem cells. J Cell Physiol. 198:223–229. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Reagan-Shaw S, Nihal M and Ahmad N: Dose
translation from animal to human studies revisited. FASEB J.
22:659–661. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mech-Nowak A, Swiderski A, Kruczek M,
Luczak I and Kostecka-Gugała A: Content of carotenoids in roots of
seventeen cultivars of Daucus carota L. Acta Biochim Pol.
59:139–141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kim Y, Jung S, Park G, Shin H, Heo SC and
Kim Y: β-Carotene suppresses cancer cachexia by regulating the
adipose tissue metabolism and gut microbiota dysregulation. J Nutr
Biochem. 114:1092482022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ballarò R, Costelli P and Penna F: Animal
models for cancer cachexia. Curr Opin. 10:281–287. 2016.PubMed/NCBI
|
|
65
|
Pin F, Busquets S, Toledo M, Camperi A,
Lopez-Soriano FJ, Costelli P, Argilés JM and Penna F: Combination
of exercise training and erythropoietin prevents cancer-induced
muscle alterations. Oncotarget. 6:432022015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Baracos VE, Martin L, Korc M, Guttridge DC
and Fearon KC: Cancer-associated cachexia. Nat Rev Dis Primers.
4:171052018. View Article : Google Scholar : PubMed/NCBI
|