Introduction
Breast cancer, with ~2.3 million new cases and
670,000 mortalities worldwide in 2022 (1,2),
remains the most prevalent malignancy among women globally,
exhibits remarkable clinical diversity and molecular heterogeneity
(3). This disease not only
demonstrates aggressive growth patterns but also involves complex
pathogenic mechanisms regulated through multiple signaling
pathways, presenting considerable therapeutic challenges. Despite
recent advancements in diagnostic and therapeutic technologies,
chemotherapy resistance and tumor recurrence remain key factors
affecting patient outcomes (4).
Current chemotherapy regimens predominantly suffer from limitations
such as low therapeutic index and severe side effects.
Consequently, developing novel anti-tumor drugs with both high
efficacy and safety has become a key research focus (5). In this context, drug repurposing, a
strategy that leverages the established pharmacological properties
of existing medications to explore new therapeutic applications,
has emerged as a potential frontier in oncology research.
Ivermectin (IVM), a classic antiparasitic agent, has
demonstrated pharmacological effects that extend beyond its
traditional use in deworming (6). A
recent study has revealed its anti-tumor efficacy through
multi-target regulatory mechanisms, including the modulation of
multidrug resistance proteins, interference with the Akt/mTOR-WNT
signalling axis and regulation of purinergic receptor systems
(7–9). Notably, metformin (MET), a commonly
used clinical hypoglycemic agent, has also exhibited
cross-indication anticancer properties (10), showing notable tumor-suppressive
effects in various malignancies such as breast cancer (11), colorectal cancer (12), glioma (13) and oral cancer (14).
Current research on combining IVM and MET remains
limited (15). Although both drugs
act on the PI3K/AKT/mTOR signalling pathway, they differ markedly
in their molecular targets: MET primarily inhibits mitochondrial
complex I to reduce the ATP/AMP ratio and regulate mTORC1 activity,
while IVM modulates cell cycle progression by affecting
cyclin-dependent kinases and autophagy-related proteins (16). This complementary mechanism provides
a theoretical basis for synergistic antitumor effects. From a drug
development perspective, the combination of IVM and MET, both
clinically validated drugs, offers advantages: i) Well-defined
pharmacokinetic parameters; ii) known adverse reaction profiles;
iii) avoidance of toxicity risks in new drug development. This drug
repurposing strategy not only shortens research and development
cycles but also considerably reduces development risks. It is
expected to provide a new intervention for cancer treatment
(17).
In previous years, the dual role of autophagy in
tumor development has garnered increasing academic attention
(18). As a highly conserved
mechanism for maintaining cellular homeostasis, autophagy
eliminates damaged organelles and misfolded proteins through
lysosomal-mediated degradation pathways, carrying out a key role in
determining the fate of tumor cells. Existing evidence suggests
that moderate activation of autophagy can markedly inhibit tumor
cell proliferation and induce programmed cell death (19). Notably, the central metabolic and
survival signaling hub, PI3K/AKT/mTOR, serves as a key molecular
switch for autophagy. Inhibiting this pathway has been shown to
effectively activate the autophagic cascade, thereby exerting
anti-tumor effects (20).
Particularly noteworthy is the discovery that platelet-derived
serum protein 1 (THBS1) acts as a notable upstream regulator of the
PI3K/AKT/mTOR pathway (21).
Meanwhile, reactive oxygen species (ROS), serving as important
intracellular second messengers, bidirectionally regulate both the
activity of this pathway and autophagy levels through
redox-sensitive mechanisms (22,23).
Building on these complementary molecular
mechanisms, the present study proposes an innovative strategy that
combines IVM with MET, synergistically targeting the PI3K/AKT/mTOR
pathway to enhance autophagy induction and thereby inhibit
malignant biological behaviors in breast cancer MCF-7 cells.
Materials and methods
Reagents and antibodies
The reagents used are as follows: IVM (purity ≥98%)
purchased from MilliporeSigma; MET (purity ≥98%), obtained from
Macklin Inc., 3-Methyladenine (3-MA; purity ≥98%; MilliporeSigma);
N-acetylcysteine (NAC; Macklin Inc.), matrix adhesive (Hangzhou
Lianke Meixun Biomedical Technology Co., Ltd.) and crystal violet
(Biosharp Life Sciences). p62/SQSTM Rabbit Ab (cat. no.
118420-1-AP; Proteintech Group Inc.); Phospho-AKT (cat. no. WLP001;
Wanleibio Co., Ltd.); THBS1 (cat. no. A75291; Nature Biosciences
LtD.); β-actin (cat. no. AC038; ABclonal Biotech Co., Ltd.); LC3B
(cat. no. A19665; ABclonal Biotech Co., Ltd.); Beclin1 (cat. no.
A21191; ABclonal Biotech Co., Ltd.); Bcl-2 (cat. no. A19693;
ABclonal Biotech Co., Ltd.); PI3 Kinase p85 (cat. no. A4992;
ABclonal Biotech Co., Ltd.); Phospho-PI3KP85α (cat. no. AP0854;
ABclonal Biotech Co., Ltd.); AKT (cat. no. A20779; ABclonal Biotech
Co., Ltd.); mTOR (cat. no. A11345; ABclonal Biotech Co., Ltd.);
Phospho-mTOR (cat. no. AP0115; ABclonal Biotech Co., Ltd.). For
Western blotting and immunofluorescence, HRP-conjugated Goat Rabbit
IgG (H+L) (cat. no. AS014; ABclonal Biotech Co., Ltd.) and Alexa
Flour 594-Goat Anti-Rabbit IgG (AD9279; cat. no. ABbox) were used
as secondary antibodies. All antibodies were stored at −20°C in
accordance with the conditions recommended in the instructions.
Cell culture
The MCF-7 human breast cancer cell line used in the
present study was kindly provided by Professor Dong Zhiqiang's
laboratory at Huazhong Agricultural University, Wuhan, China. The
cells were cultured in high-glucose DMEM medium (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(BIOVISTECH PTY. LTD) and 1% penicillin-streptomycin dual
antibiotic solution (MilliporeSigma) and maintained in a
constant-temperature incubator at 37°C, 5% CO2 and 95%
humidity. When the cell confluence reached 80–90%, they were
passaged using 0.25% trypsin (EDTA-containing) for 1 min at 37°C,
then divided into culture flasks at a 1:3 ratio. All experiments
strictly limited cell passage to ≤15 generations, with regular
mycoplasma detection and morphological observation to ensure cell
viability.
Cell viability determination
MCF-7 cells in the logarithmic growth phase were
prepared as single-cell suspensions (1×104 cells/ml) and
inoculated into 96-well culture plates (100 µl per well), with
three replicates per group. After 12 h of cell adhesion (37°C, 5%
CO2) to achieve 50% confluency, the following treatments
were administered: Blank control (with PBS only), IVM monotherapy
(5–45 µmol/l), MET monotherapy (5–50 mmol/l) and combined treatment
(IVM + MET; aforementioned concentrations). Treatment durations
were set at 12, 24 and 48 h at 37°C in a 5% CO2
incubator. For mechanistic studies, cells were pre-treated with
3-MA (5 mM; 2 h) or NAC (10 mM; 2 h) at 37°C in a 5% CO2
incubator before receiving the final drug at 24 h. After culture
termination, 100 µl working solution was added per well according
to the Cell Counting Kit-8 (CCK-8; Wuhan Huiyucheng Biotechnology
Co., Ltd.) protocol. The mixture was incubated at 37°C for 30 min,
followed by absorbance measurement using a Bio-Rad Laboratories,
Inc. microplate reader at a wavelength of 450 nm (OD value) to
assess cell viability. Three independent replicates were performed
throughout the experiment to ensure data reliability.
Cell invasion test
The present study employed the Transwell invasion
assay to evaluate the combined effects of IVM and MET on the
invasion of MCF-7 cells. A 100 µl Matrigel matrix gel (Xi'an
Zhongtuan Biotechnology Co., Ltd.) was uniformly coated on the
upper chamber of Transwell chambers (8 µM pore size; Corning,
Inc.). After 4 h of polymerization at 37°C, 100 µl of cell
suspension (in serum-free DMEM) containing different treatment
groups (density 5×104 cells/ml) was inoculated into the
upper chamber. By contrast, 800 µl DMEM medium with 10% fetal
bovine serum served as a chemotactic inducer in the lower chamber.
The cultures were incubated at 37°C with 5% CO2 for 24
h. Following medium removal, cells were washed with PBS, fixed with
100% methanol for 30 min at 37°C, stained with 0.1% crystal violet
for 30 min at 37°C and membrane-penetrating cell counts were
quantified by randomly selecting 5 fields under an inverted Olympus
microscope (Olympus Corporation).
Assessment of cell migration ability
(scratch test)
The present study evaluated the combined effects of
IVM and MET on MCF-7 cell migration using a scratch assay. The
protocol involved seeding MCF-7 cells at 2×105 cells per
well in 6-well plates in DMEM supplemented with 10% fetal bovine
serum, incubating them at 37°C with 5% CO2 for 80%
fusion. A 200 µl sterile pipette was used to create vertical
scratch lines perpendicular to the plate surface, followed by
gentle washing with PBS to remove detached cells. Four experimental
groups were established: Serum-free DMEM control, 6 µM IVM
monotherapy, 6 mM MET monotherapy and IVM (6 µM) + MET (6 mM)
combined intervention. All groups were cultured under identical
conditions for 24 h, with the progress of scratch closure monitored
every 8 h using an inverted microscope (100×; Olympus CKX41;
Olympus Corporation). Migration area changes were quantified using
ImageJ software (version 1.53e; National Institutes of Health) to
evaluate treatment effects.
Measurement of intracellular ROS
The ROS levels in MCF-7 cells were detected using
the 2′,7′-Dichlorodihydrofluorescein diacetate (DCFH-DA)
fluorescence probe method. The experimental protocol was as
follows: MCF-7 cells were seeded at 2×105 cells per well
and cultured in 6-well plates until 70% confluency, then subjected
to the following treatments: i) Different drug groups (PBS, IVM,
MET and combination therapy) for 12 or 24 h at 37°C in a 5%
CO2 incubator; ii) drug administration after 2 h
pretreatment with 10 mM NAC; iii) positive control [Ros up (a
compound mixture that induces ROS production); 50 µg/ml stimulation
for 20 min at 37°C; Beyotime Biotechnology]. Cells were collected
and adjusted to a density of 1×109 cells/ml. Then, they
were incubated with a 10 µM DCFH-DA probe (Wuhan Huiyucheng
Biotechnology Co., Ltd.) at 37°C in the dark for 30 min. After
washing 3× with PBS and resuspension, the fluorescence intensity
was measured using a flow cytometer (CytoFLEX; Beckman Coulter)
with excitation/emission wavelengths of 488 nm and 530 nm,
respectively. Each group had 3 duplicate wells, and the experiment
was independently repeated 3 times. Data were expressed as mean
fluorescence intensity ± SD.
Transmission electron microscopy
observation
The present study employed a transmission electron
microscope (accelerated voltage 100 kV; Hitachi H7650; Hitachi,
Ltd.) to investigate the effects of IVM, MET and their combination
on the ultrastructure of MCF-7 cells. Cells were inoculated into
6-well plates (1×105 cells per well). After achieving
60–70% fusion, they were treated with the respective drugs (PBS,
IVM, MET or IVM + MET) for 24 h at 37°C in a 5% CO2
incubator. After fixation with 2.5% glutaraldehyde for 2 h at 4°C,
the samples underwent an ethanol gradient dehydration process (50,
70, 90 and 100%) followed by epoxy resin embedding and
polymerization at 60°C for 48 h and ultra-thin sectioning (80 nm,
using a Leica UC6 microtome; Leica Microsystems). Uranium acetate
staining for 30 min at room temperature in the dark was applied to
enhance contrast. A total of 10 random fields from each group were
systematically observed, with a particular focus on analyzing
autophagosome formation and morphological changes in organelles,
such as mitochondria and the endoplasmic reticulum.
Western blotting
The present study systematically evaluated the
effects of various treatment protocols on protein expression
profiles in MCF-7 cells using western blot analysis. The
experimental protocol was as follows: MCF-7 cells were cultured in
6-well plates until 80% confluency, after which the following
groups were established: i) PBS control group, IVM single drug
group (6 µM), MET single drug group (6 mM) and combined treatment
group, all treated for 24 h at 37°C in a 5% CO2
incubator; ii) Autophagy inhibitor pretreatment group (5mM 3-MA
administered 2 h prior to treatment at 37°C in a 5% CO2
incubator). Experimental procedures included collecting cells,
washing them 3 times with pre-chilled PBS, adding RIPA lysis buffer
(containing protease inhibitors; cat. no. P0013B; Beyotime
Biotechnology) for lysing on ice for 30 min. Protein concentrations
were measured using the BCA Protein Quantification Kit (Thermo
Fisher Scientific Inc.) and adjusted to a uniform concentration of
15 µg/well. Protein samples were separated by 10% SDS-PAGE and
transferred to a PVDF membrane using the gel-membrane sandwich that
was submerged in transfer buffer and electrophoresed at constant
voltage. Subsequent steps included blocking (5% skim milk; room
temperature for 2 h), primary antibody incubation at 4°C overnight
with the following dilutions: p62/SQSTM (1:1,000), Phospho-AKT
(1:500), THBS1 (1:500), β-actin (1:2,000), LC3B (1:1,000), Beclin1
(1:1,000), Bcl-2 (1:1,000), PI3K p85 (1:1,000), Phospho-PI3K p85
(1:500), AKT (1:1,000), mTOR (1:1,000) and Phospho-mTOR (1:500)
secondary antibody incubation (HRP-labelled; 1:5,000 dilution; room
temperature for 2 h). Finally, signals were detected using a Fusion
FX7 chemiluminescence imaging system and quantified with ImageJ
software (version 1.53e; National Institutes of Health) to analyze
the expression ratio of the target protein to the internal control
β-actin. To ensure experimental reliability, all experiments
included three independent biological replicates.
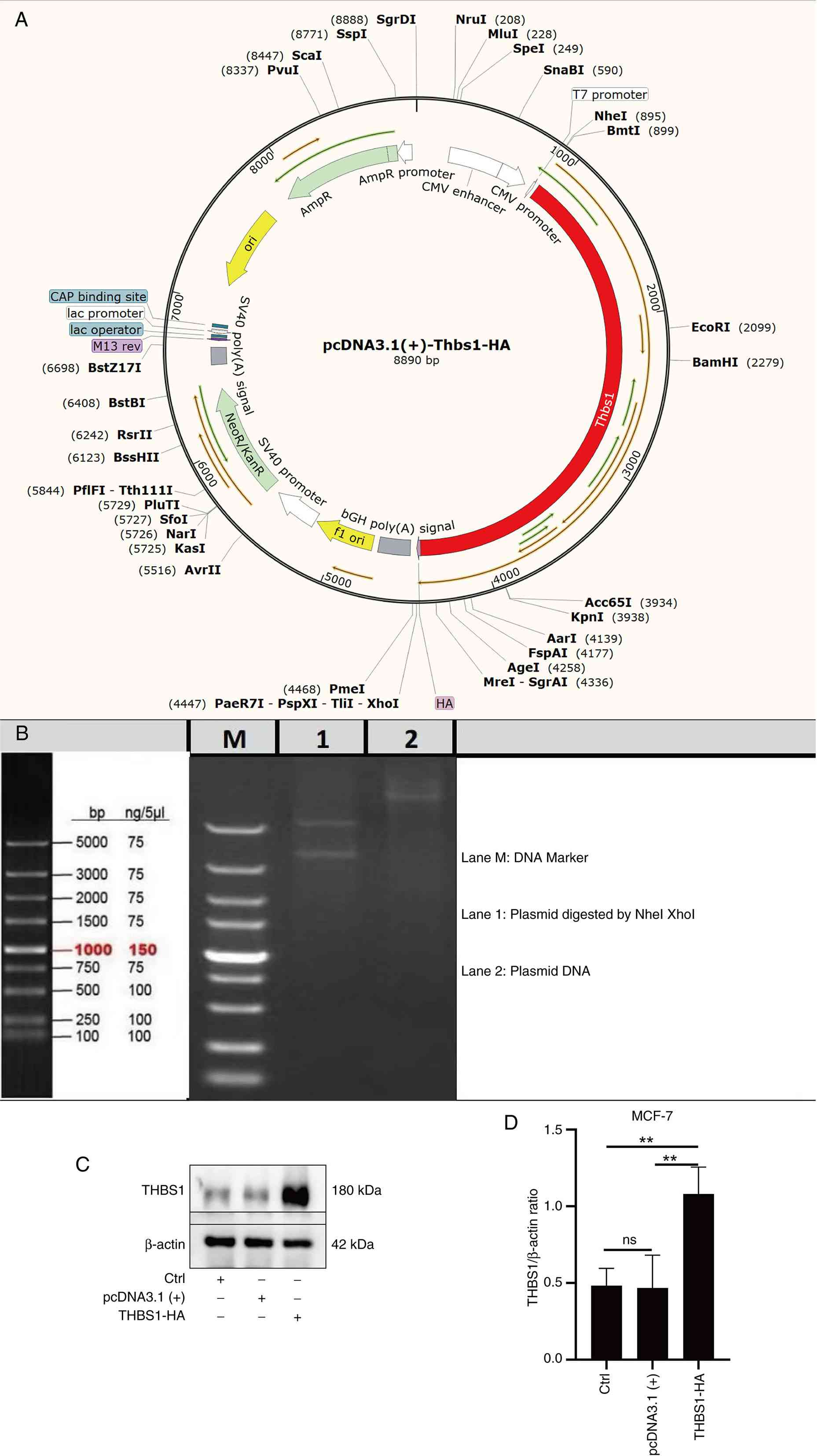
Construction of overexpressed
plasmids
The present study employed molecular cloning
techniques to construct an overexpression vector for the THBS1
gene. Specific primers based on the mouse THBS1 coding sequence
(accession no. NP_035710.2) from the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/)
were first designed to amplify the target gene. The primer
sequences used for amplifying the mouse THBS1 CDS were as follows:
Forward,
5′-CTATAGGGAGACCCAAGCTGGCTAGCGCCACCATGGAGCTCCTGCGGGGACTAGGTGTCCTGTTCCTGTTGCATATG-3′;
reverse,
5′-GTTTAAACGGGCCCTCTAGACTCGAGTTAAGCGTAGTCTGGGACGTCGTATGGGTAGGAATCTCGACACTCGTATTTC-3′.
The reverse primer sequence is presented in its full-length
synthesized form, which includes a 5′ vector homology arm (for
In-Fusion cloning) and a sequence encoding a C-terminal HA tag
(Tyr-Pro-Tyr-Asp-Val-Pro-Asp-Tyr-Ala) immediately upstream of the
THBS1 gene-specific region. For sequence verification purposes,
only the gene-specific portion should be used for BLAST alignment
against the mouse THBS1 reference sequence. Using MCF-7 cell cDNA
as a template, PCR amplification was performed with PrimeSTAR HS
high-fidelity DNA polymerase (Takara Bio, Inc.) to ensure genetic
accuracy. The amplified products were digested with NheI/XhoI
restriction endonucleases (Thermo Fisher Scientific, Inc.) and
ligated to the homologous pcDNA3.1 (+) eukaryotic expression
vector. The recombinant plasmid was transformed into DH5α competent
cells (Beijing Quanshijin Biotechnology Co., Ltd.) via the heat
shock method, followed by ampicillin resistance screening to
identify positive clones. After Sanger sequencing (Shenzhen BGI
Genomics Co., Ltd.) confirmed the correct insertion sequences, the
vector (2 µg per well of a 6-well plate) was introduced into MCF-7
cells using Lipofectamine® 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.) via liposome transfection at 37°C in a 5%
CO2 incubator for 6 h, after which the medium was
replaced with fresh complete medium. Western blotting was used to
verify the expression levels of THBS1 protein. In these
experiments, cells transfected with the empty pcDNA3.1(+) vector
served as the negative control.
Immunofluorescence experiment
The present study utilized immunofluorescence
technology to assess the effects of various treatments on MCF-7
cells. MCF-7 cells were cultured at a density of 1×104
cells per well in 24-well plates until they reached 80% confluency.
A total of four treatment groups were established: Blank control
(PBS), 6 µM IVM (IVM monotherapy), 6 mM MET (MET monotherapy) and
combined treatment group (6 µM IVM + 6 mM MET). After 24-h
treatment, cells were washed with PBS, fixed in 4% polyformaldehyde
(Biosharp Life Sciences) for 30 min at room temperature,
permeabilized with 0.2% Triton X-100 for 15 min at room temperature
and blocked with 5% goat serum for 2 h at room temperature. Samples
were incubated with specific primary antibodies at the following
dilutions: Anti-LC3B (1:200) and anti-p62 (1:200) overnight at 4°C,
followed by a 2-h incubation at room temperature in the dark with a
FITC-labelled secondary antibody (1:500). DAPI staining (Biosharp
Life Sciences) was performed for 5 min at room temperature. The
cells were observed under a fluorescence microscope (Olympus IX73;
Olympus Corporation).
Statistical analysis
This study employed standardized statistical methods
to process and analyze the experimental data. All quantitative data
were derived from three independent biological replicate
experiments, presented as mean ± SD. Data analysis was conducted
using GraphPad Prism 10.1 (Dotmatics) professional statistical
software, with one-way ANOVA primarily used for intergroup
comparisons, supplemented by Tukey's post hoc tests for multiple
comparisons. Statistical significance thresholds were defined as
follows: P<0.05 indicates a statistically significant
difference, P<0.01 denotes highly significant differences,
P<0.001 signifies extremely significant differences, while
P>0.05 (marked as ns) indicates no statistically significant
difference.
Results
IVM combined with MET inhibits the
proliferative activity of MCF-7 cells in vitro
Results of the present study demonstrated that both
IVM and MET monotherapy reduced MCF-7 cell viability, exhibiting
typical concentration-dependent and time-dependent characteristics
(Fig. 1A and B). Through the
calculation of half-inhibitory concentrations (I) and orthogonal
experimental analysis, the optimal combination concentration was
determined to be 6 µM IVM combined with 6 mM MET, which confirmed
the enhanced inhibitory effect of the combination (Fig. 1C). Notably, the combined treatment
group exhibited a significant synergistic effect, demonstrating
markedly improved inhibitory efficacy when compared with either
monotherapy (Fig. 1D). This
optimized formulation was subsequently employed in subsequent
experimental studies.
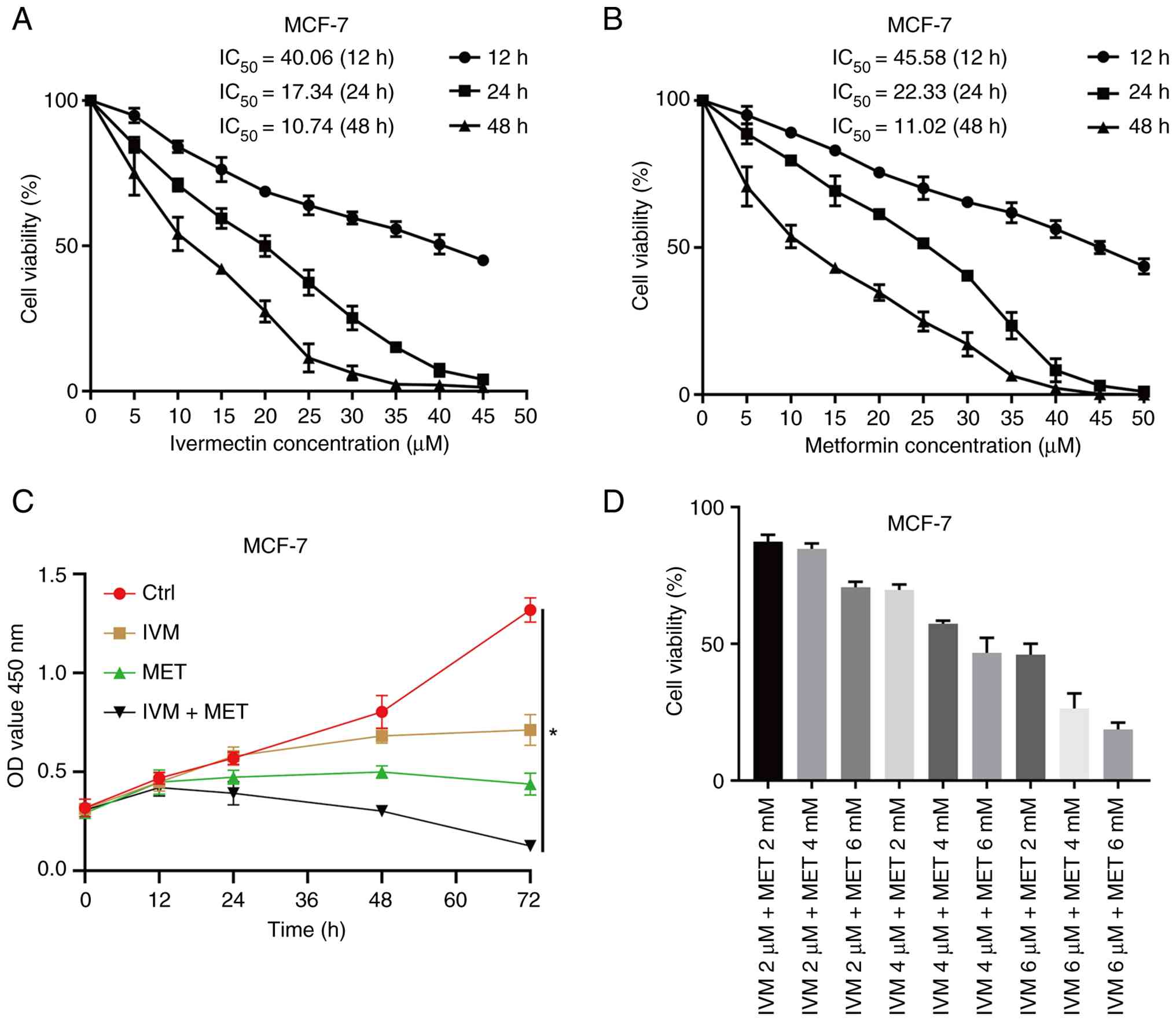 | Figure 1.Effects of IVM, MET and the
combination of the two drugs on the toxicity and proliferation of
MCF-7 cells. (A) Cell viability assays after treatment of MCF-7
cells with IVM. (B) Cell viability assays after treatment of MCF-7
cells with MET. (C) CCK-8 assay evaluating cell viability in MCF-7
cells treated with PBS, IVM, MET and IVM + MET at 0, 12, 24, 48 and
72 h; (D) Comparison of MCF-7 cell viability after co-treatment
with 2, 4 µM and 6 mMdoses of IVM combined with 2, 4 and 6 mmol/l
doses of MET using the CCK-8 method. CCK-8, Cell Counting Kit-8;
IVM, ivermectin; MET, metformin, MCF-7, human mammary tumor cell
line; Ctrl, control. All data were measured as mean ± SEM by three
independent experiments. *P<0.05. |
IVM combined with MET inhibits the
invasion and migration of MCF-7 cells in vitro
The combination of IVM and MET demonstrated
significant inhibitory effects on the malignant phenotype of MCF-7
cells. Transwell invasion assays revealed that, compared with the
control group and single-drug treatment groups, the combined
treatment group exhibited a markedly reduced cell invasion capacity
(Fig. 2A and B). The
scratch-healing assay further indicated that the drug-coated
treatment group exhibited significantly slower cell migration rates
compared with both the control and single-drug treatment groups
(Fig. 2C and D). These data
collectively demonstrate that the synergistic application of IVM
and MET effectively suppresses the invasive migration capabilities
of breast cancer cells.
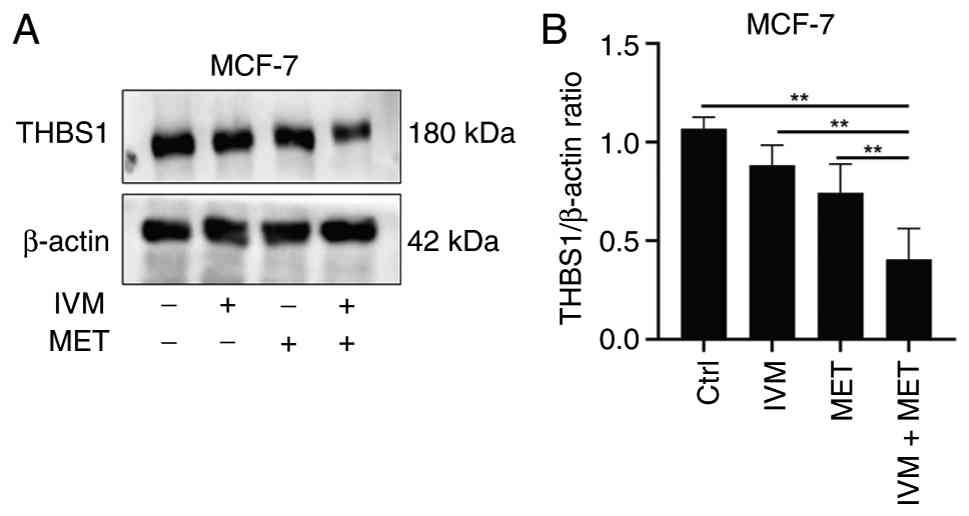
IVM combined with MET decreases the
expression level of THBS1 protein
During the development of breast tumors, various
cytokines and protein molecules carry out key roles. THBS1, a key
extracellular matrix protein, is associated with tumor cell
proliferation, migration and invasion (24). Western blot analysis revealed that,
compared with the control group and single-drug treatment groups,
the IVM combined with MET treatment significantly reduced THBS1
expression in MCF-7 cells, with statistically significant
difference; (Fig. 3A and B). These
results demonstrate that the IVM combined with MET effectively
suppresses THBS1 expression in breast cancer cells.
IVM combined with MET reduced the
phosphorylation level of the PI3K/AKT/mTOR pathway
The PI3K/AKT/mTOR signaling pathway plays a key role
in the development of tumors (25).
The present study employed western blotting to evaluate the effects
of different drug combinations on phosphorylation levels within
this pathway. Results demonstrated that when IVM was combined with
MET treatment, intracellular phosphorylation levels of
phosphorylated (p)-PI3K, p-AKT and p-mTOR were significantly
reduced in MCF-7 cells compared with control groups and single-drug
treatments (Fig. 4A and B). These
findings suggest that the IVM-MET combination may target the
PI3K/AKT/mTOR pathway and induce tumor cell death.
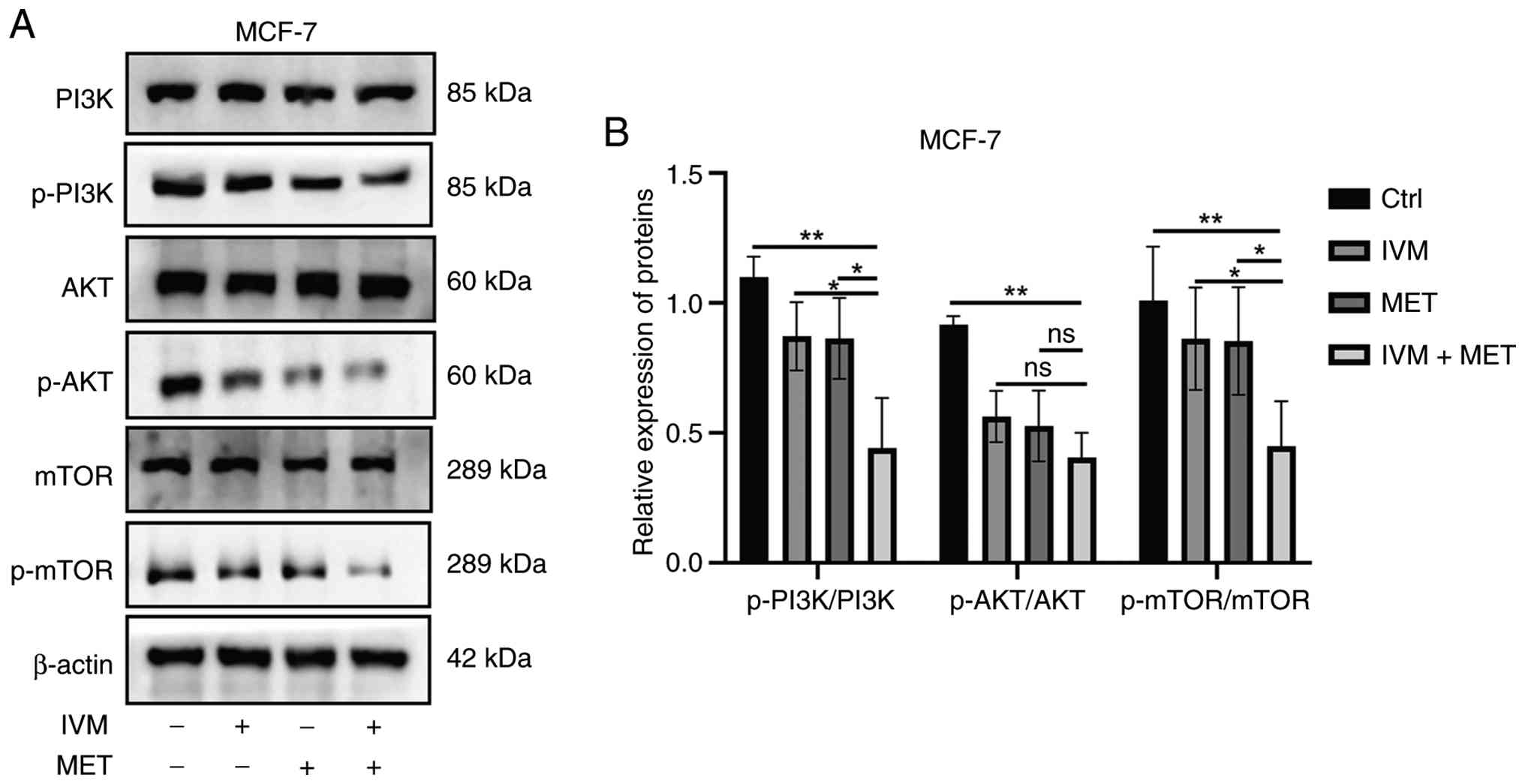 | Figure 4.IVM combined with MET reduces the
phosphorylation levels of proteins in the PI3K/AKT/mTOR pathway.
(A) Representative images of western blotting and, (B) The
expression levels of PI3K, AKT, mTOR and their phosphorylated
proteins in MCF-7 cells treated with PBS, IVM, and IVM + MET. IVM,
ivermectin; MET, metformin, MCF-7, human mammary tumor cell line;
Ctrl, control; p, phosphorylated. All data were measured as mean ±
SEM by three independent experiments. *P<0.05, **P<0.01. |
Overexpression of THBS1 enhances the
levels of phosphorylated PI3K, AKT and mTOR
Studies have demonstrated that the expression of
THBS1 is regulated by the PI3K/AKT/mTOR signaling pathway (21,26).
To investigate whether THBS1 exerts feedback regulation on this
pathway, the present study utilized transcriptome data
(NP_035710.2; NCBI database). The present study identified the CDS
region of THBS1 from the National Centre for Biotechnology
Information (NCBI) database. Using pcDNA3.1 (+) as the vector, an
overexpression plasmid for THBS1 was constructed (Fig. 5A). Agarose gel electrophoresis
revealed amplified gene fragments from the THBS1 CDS region,
measuring 3558 bp in length (Fig.
5B).
The THBS1 overexpression construct used in this
study was originally generated for parallel investigations in mouse
and canine mammary tumor models, leveraging the high cross-species
conservation of THBS1. Its application in human MCF-7 cells is
supported by the >97% amino acid sequence identity between mouse
and human THBS1, ensuring functional relevance (27).
Western blotting was used to detect the
overexpression efficiency of THBS1 in MCF-7 cells, transfected with
the empty pcDNA3.1(+) vector serving as the negative control. The
results showed that, compared with the Ctrl group and the negative
control [pcDNA3.1(+)] group, the overexpression efficiency of THBS1
was significantly increased (Fig. 5C
and D).
IVM combined with MET induces
autophagy in MCF-7 cells
Disruption of autophagy is associated with the
development and progression of various diseases. To investigate the
effects of IVM combined with MET on autophagy in breast tumors, the
present study employed cellular immunofluorescence to detect
autophagy-related proteins LC3B and p62 in MCF-7 cells. The results
showed that compared with the control group, the combined treatment
group exhibited elevated LC3B expression (Fig. 6A) and decreased p62 levels (Fig. 6B), indicating that IVM combined with
MET can induce excessive autophagy in breast tumor cells. Excessive
autophagy leads to over-degradation of multiple cellular
components. Transmission electron microscopy analysis revealed
that, compared with the control group and single-drug treatment
groups, the IVM + MET group contained numerous
double-membrane-structured autophagosomes and an increased number
of autophagic lysosomes (Fig. 6C).
Western blot analysis revealed that, compared with the PBS control
group, LC3B and Beclin1 protein expressions were upregulated in
MCF-7 cells after IVM, MET and IVM + MET treatments, while p62 and
Bcl2 were downregulated (Fig. 6D and
E). These results demonstrate that IVM, when combined with MET,
can promote autophagy in MCF-7 cells.3-MA is an early-stage
autophagy inhibitor (28). To
investigate how autophagy inhibition affects cytotoxicity caused by
IVM combined with MET, the present used the CCK-8 assay to evaluate
the effects of co-treatment with 3-MA and/or IVM + MET on cell
viability. The results showed that the co-treated group exhibited a
significant rebound in cell activity compared with the IVM +
MET-only group (Fig. 6F).
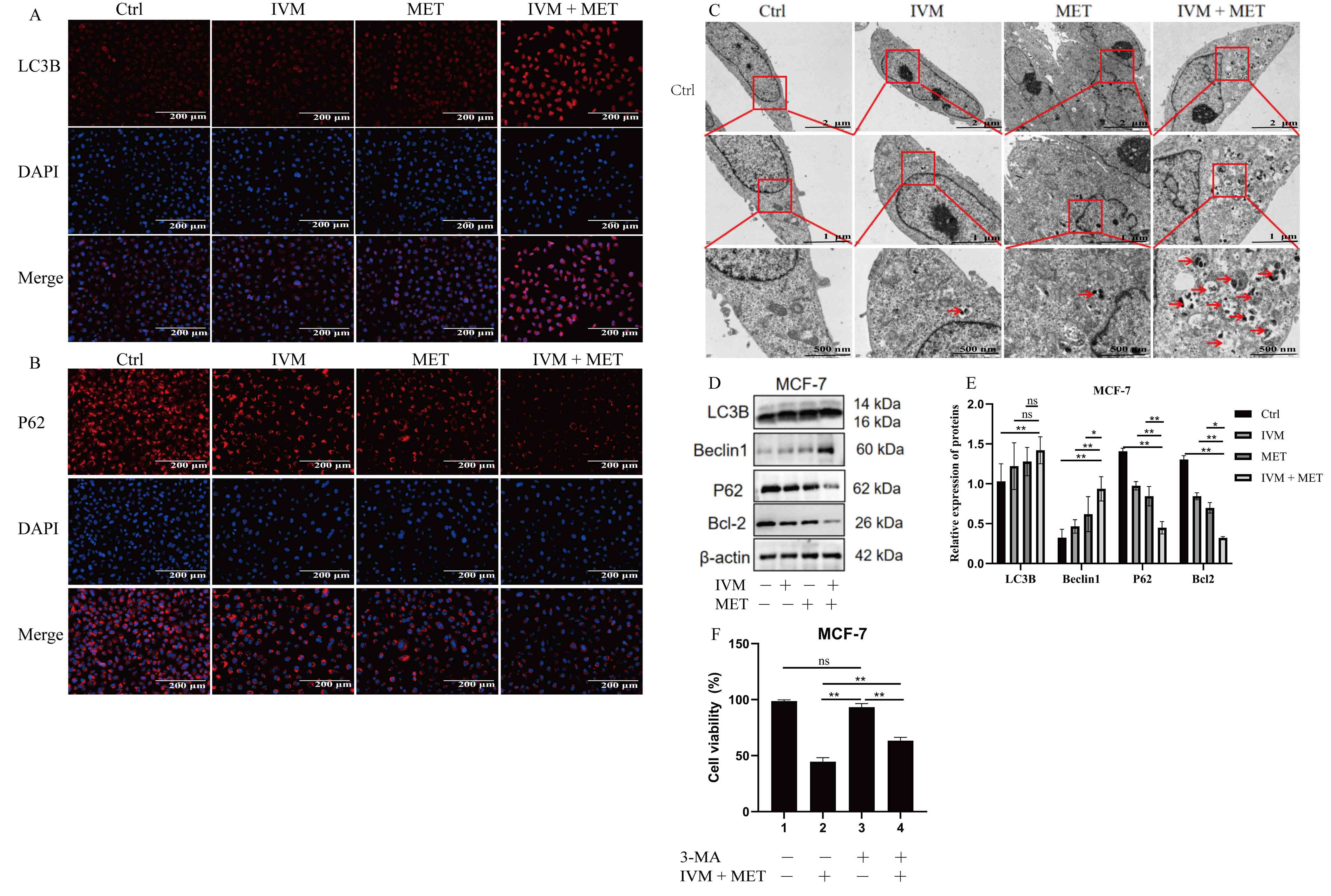 | Figure 6.Autophagy induction in MCF-7 cells by
IVM combined with MET. (A) Representative immunofluorescence images
showing the expression and localization of the autophagosome marker
LC3B (red) in MCF-7 cells treated with PBS (Ctrl), IVM (6 µM), MET
(6 mM) or IVM + MET for 24 h. Nuclei are stained with DAPI (blue).
The pronounced increase in LC3B puncta formation (indicated by
arrows) in the IVM + MET-treated group indicates autophagosome
accumulation, a hallmark of autophagy induction. Scale bar, 200 µm.
(B) Representative immunofluorescence images showing the expression
and localization of the autophagic flux marker p62/SQSTM1 (red) in
MCF-7 cells under the same treatment conditions as in (A). Nuclei
are stained with DAPI (blue). The marked reduction in p62
fluorescence intensity in the IVM + MET-treated group reflects
enhanced autophagic degradation, confirming active autophagic flux.
Scale bar, 200 µm. (C) Representative transmission electron
microscopy images of MCF-7 cells following 24 h of treatment.
Images are shown at increasing magnifications with scale bars of 2
µm (top), 1 µm (middle), and 500 nm (bottom) for each treatment
group (Ctrl, IVM, MET and IVM + MET). Numerous double-membrane
autophagosomes (indicated by red arrows) are evident in the IVM +
MET-treated cells, providing ultrastructural confirmation of
autophagy activation. (D) Western blot analysis of key
autophagy-related proteins (LC3B-I/II, Beclin1, p62 and Bcl-2) in
MCF-7 cells treated as indicated for 24 h. β-actin serves as the
loading control. (E) Densitometric semi-quantification of the
protein levels. (F) Cell viability assessment by Cell Counting
Kit-8 assay. Pre-treatment with 3-MA significantly rescued the loss
of cell viability induced by IVM + MET, confirming the functional
contribution of autophagy to the cytotoxic effect. IVM, ivermectin;
MET, metformin, MCF-7, human mammary tumor cell line; Ctrl,
control. All data were measured as mean ± SEM by three independent
experiments. *P<0.05, **P<0.01. |
IVM combined with MET promotes
autophagy of breast tumor cells through ROS
Flow cytometry analysis of ROS in MCF-7 cells
revealed significantly elevated intracellular ROS levels following
IVM and MET treatments compared with the control group, with the
combined IVM + MET treatment showing even higher ROS levels
(P<0.05; Fig. 7A and B). This
suggests that the IVM + MET regimen may induce oxidative stress by
increasing intracellular ROS, thereby affecting cell survival and
function. NAC, an antioxidant that reduces intracellular ROS
(29).
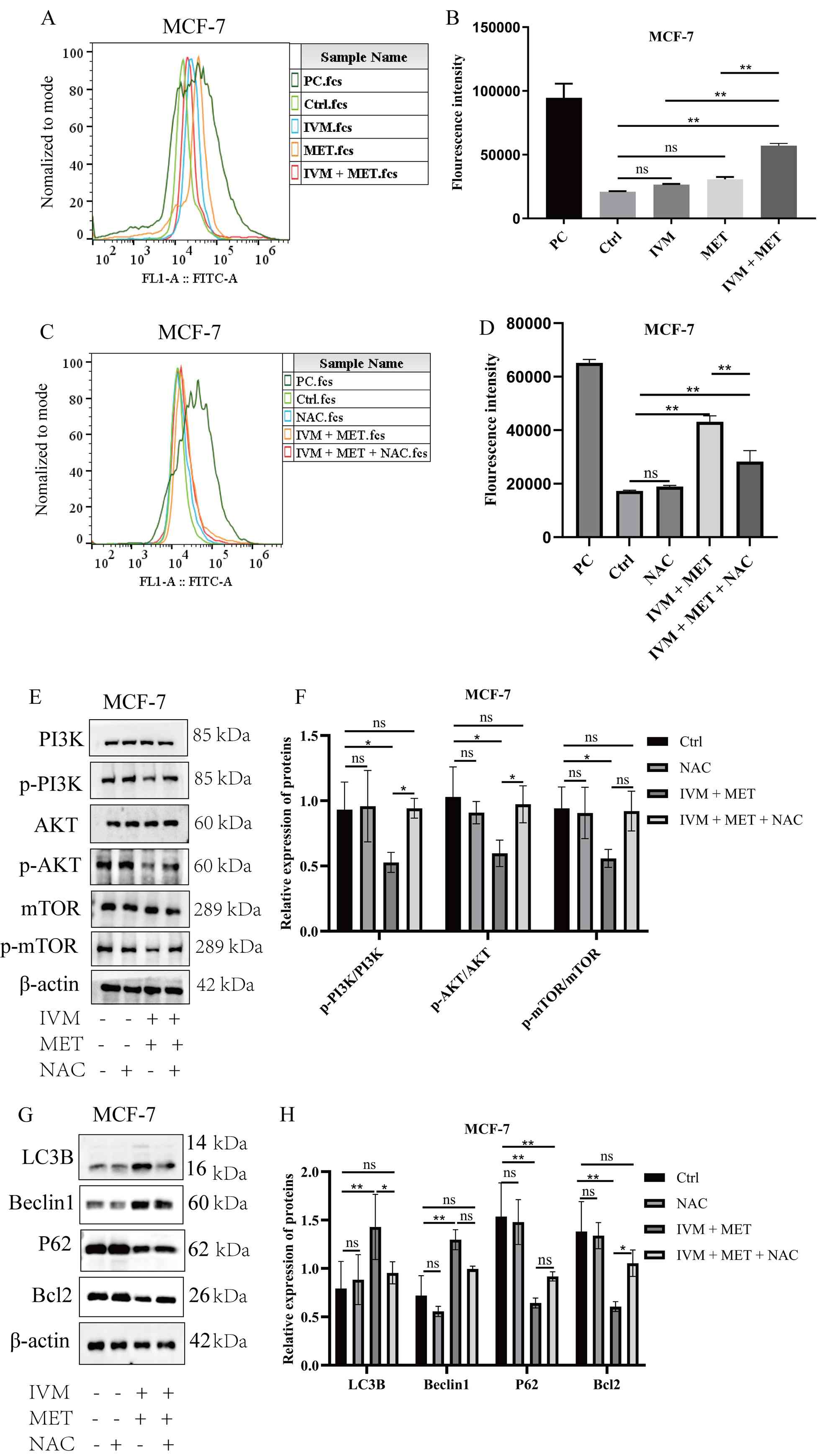 | Figure 7.IVM combined with MET promotes the
accumulation of ROS in MCF-7 cells. (A) Representative flow
cytometry plot and (B) quantification of ROS expression levels in
MCF-7 cells after treatment with different experimental groups. PC,
Ctrl, IVM, MET and IVM + MET groups are shown. (C) Representative
flow cytometry plot and (D) quantification of ROS expression levels
in MCF-7 cells after treatment with NAC. PC, Ctrl, IVM, MET, and
IVM + MET groups are shown. (E) Representative western blotting
images and (F) semi-quantification of p-PI3K, p-AKT and p-mTOR
protein expression levels in MCF-7 cells after treatment with
different experimental groups; (G) Representative western blotting
images and (H) semi-quantification of autophagy-related protein
expression levels in MCF-7 cells treated with PBS, IVM, MET and IVM
+ MET. ROS, reactive oxygen species; p, phosphorylated; IVM,
ivermectin; MET, metformin, MCF-7, human mammary tumor cell line;
Ctrl, control; PC, positive control. All data were measured as mean
± SEM by three independent experiments. *P<0.05,
**P<0.01. |
MCF-7 cells were pre-treated with 10 mM NAC in the
present study (NAC group). Subsequent experimental groups included
Ctrl, IVM + MET and IVM + MET + NAC. After 24 h of stimulation, ROS
levels were measured using flow cytometry. Results demonstrated a
partial reduction in ROS levels in the IVM + MET + NAC group
compared with the IVM + MET group (Fig.
7C and D). To investigate whether IVM + MET affects
PI3K/AKT/mTOR pathway phosphorylation through ROS, western blotting
was employed to detect p-PI3K, p-AKT and p-mTOR protein expression
in MCF-7 cells. The results demonstrated that IVM + MET inhibited
phosphorylation activity in the PI3K/AKT/mTOR signaling pathway
compared with Ctrl and single-drug treatments, and this inhibition
was associated with elevated intracellular ROS levels. Furthermore,
compared with the IVM + MET group, the IVM + MET + NAC group showed
a partial elevation in protein expression levels of p-PI3K, p-AKT
and p-mTOR (Fig. 7E and F). This
suggests that the combination of IVM and MET may suppress
phosphorylation levels in the PI3K/AKT/mTOR signaling pathway by
inducing ROS accumulation. When intracellular ROS levels increase,
key proteins in the PI3K/AKT/mTOR signaling pathway undergo
oxidative modifications, resulting in reduced activity (30). To investigate whether the IVM + MET
combination could alter autophagy-related proteins via ROS, western
blot analysis was performed on MCF-7 cells for LC3B, Beclin1, p62
and Bcl-2. Results revealed that, compared with the IVM + MET
group, the IVM + MET + NAC group exhibited partial decreases in
LC3B and Beclin1 protein expression, while showing partial
increases in p62 and Bcl-2 expression (Fig. 7G and H). These findings suggest that
the IVM + MET combination inhibits the phosphorylation of the
PI3K/AKT/mTOR signaling pathway by inducing excessive ROS
accumulation within cells, thereby triggering tumor cell autophagy
and exerting antitumor effects.
Discussion
Biosynthetic inhibition (IVM) combined with meta
protection (MET) demonstrated potent antitumor effects on MCF-7
breast cancer cells, effectively suppressing their proliferation,
migration and invasiveness. These findings suggest that the
combined therapy may exert its anti-tumor efficacy by modulating
key biological processes in tumor cells. RNA sequencing has emerged
as a powerful and versatile tool for gene expression analysis
(31). Transcriptomic analysis of
drug combinations in mouse models from NCBI revealed 53
differentially expressed genes across the PBS-treated control
(Ctrl), IVM (6 µM) + Ctrl, MET (6 mM) + Ctrl, and IVM (6 µM) + MET
(6 mM) groups. The grouping strategy included comparisons of the
combination treatment against the Ctrl, as well as against each
single agent (IVM or MET alone), as shown in the experimental
design.
IVM + PBS-treated control (Ctrl), MET + Ctrl and IVM
+ MET groups. GO enrichment analysis identified key functional
pathways, including protein trafficking, cell differentiation, DNA
repair, signaling pathways and ubiquitination. KEGG pathway
analysis revealed considerable enrichment in PI3K/AKT/mTOR, MAPK
signaling and tumor-related pathways. Our transcriptional studies
further demonstrate that IVM and MET synergistically regulate
THBS1. As THBS1 is associated with activation of the PI3K/AKT/mTOR
pathway, overexpression of THBS1 enhances PI3K/AKT phosphorylation,
thereby promoting glioma cell migration and proliferation (32,33).
Conversely, THBS1 knockdown inhibits activation of the PI3K/AKT
signaling pathway (34). In the
present study, the combination of IVM and MET reduced the protein
expression level of THBS1 and inhibited the activation of the
PI3K/AKT/mTOR signaling pathway. To further validate these
findings, the THBS1-pCDNA3.1(+) vector was transfected into MCF-7
cells to achieve overexpression of THBS1 protein. Experimental
results demonstrated that the overexpression of THBS1 partially
restored phosphorylation levels in this signaling pathway. These
findings suggest that the combination of IVM and MET may target
THBS1 to inhibit tumor cell proliferation and survival, indicating
a novel approach for developing anti-tumor drugs that target the
PI3K/AKT/mTOR signaling pathway. however, further in vitro
and in vivo experiments would first be required.
PI3K/AKT/mTOR is highly associated with autophagy.
Studies have shown that hawthorn acid can inhibit the protein
expression of PI3K-p110α, p-Akt and p-mTOR, thereby inducing
autophagy in nasopharyngeal carcinoma cells (35,36).
Glycyrrhizin A can suppress the phosphorylation of the
PI3K/AKT/mTOR signaling pathway, promoting autophagy in renal
cancer cells (37). Berberine II
inhibits the PI3K/AKT/mTOR pathway, enhancing autophagy in MCF-7
cells (38). Therefore, the present
study investigated the PI3K/AKT/mTOR pathway to elucidate the
molecular mechanisms underlying the interaction between IVM and
MET, as well as the role of autophagy. Western blotting results
revealed that IVM combined with MET inhibited the phosphorylation
in the PI3K/AKT/mTOR pathway, consistent with findings from another
tumor study (39).
Immunofluorescence analysis of LC3B and p62 proteins in MCF-7 cells
revealed increased LC3B levels and decreased p62 levels.
Transmission electron microscopy observations of autophagosome
formation in Ctrl, IVM, MET and IVM + MET groups demonstrated that
the IVM + MET group had the highest volume of autophagosomes.
Western blot analysis of LC3B and p62 expression in all three cell
types revealed that IVM, combined with MET, upregulated LC3B
expression while downregulating p62.
Based on these findings, the present study concluded
that the combination of IVM and MET inhibits the phosphorylation of
protein in the PI3K/AKT/mTOR pathway, thereby promoting excessive
autophagy in tumor cells. The present study revealed that both the
activation level of the PI3K/AKT/mTOR pathway and autophagy levels
can be regulated by ROS. The effects of IVM combined with MET on
ROS levels in breast tumor cells using in vitro experiments
were first examined. Results revealed that, compared with using
either IVM or MET alone, the ROS levels in breast tumor cells were
increased under the combined treatment group. This suggests that
the synergistic effect of IVM and MET may stem from their ability
to elevate intracellular ROS levels. ROS, as products of oxidative
stress within cells, exhibit diverse biological functions. Under
normal physiological conditions, intracellular ROS levels remain
relatively stable. However, in tumor cells, disrupted redox balance
often leads to elevated ROS concentrations (40). Previous research has demonstrated
that star fruit root can inhibit cardiomyocyte apoptosis by
regulating ROS-mediated PI3K/AKT/mTOR autophagy pathways (41). Findings of the present study
indicated that IVM combined with MET induces excessive ROS
accumulation and further promotes autophagy in breast tumor cells,
suggesting that ROS regulation may constitute another key mechanism
by which IVM and MET suppress tumor biology.
Autophagy, a process that involves the degradation
and recycling of intracellular material, can inhibit tumor
development during its early stages (42). The accumulation of ROS can induce
cellular oxidative stress, impairing cell proliferation and
survival by triggering autophagy. To investigate their interplay,
pre-treatment with NAC (a ROS scavenger) was employed. Results
demonstrated that NAC alleviates ROS overproduction caused by the
combination of IVM and MET, partially restores phosphorylation
levels in the PI3K/AKT/mTOR signaling pathway and reduces autophagy
in breast cancer cells. However, the present study has limitations.
First, the in vitro model, while controlled, does not fully
recapitulate the complex tumor microenvironment in vivo.
Second, while NAC was used to establish the functional role of ROS,
future investigations could characterize the specific alterations
in the antioxidant defense system (for example, superoxide
dismutase, catalase or glutathione peroxidase activity) in response
to IVM + MET treatment to fully delineate the redox adaptations
involved. Another limitation of the present study is the lack of a
pharmacological positive control (for example rapamycin) in the
immunofluorescence assay for autophagy. However, the conclusion of
the present study is supported by the convergence of evidence from
transmission electron microscopy (direct observation of
autophagosomes), western blot analysis of autophagic flux markers
and a functional rescue experiment using the autophagy inhibitor
3-MA. These findings suggest that the synergistic effect of IVM and
MET may stem from the elevation of ROS, subsequent suppression of
the PI3K/AKT/mTOR pathway phosphorylation and induction of
autophagy. This discovery provides a scientific foundation for
exploring the therapeutic applications of IVM and MET in the
treatment of breast cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by a research grant ‘Dandelion
Sterol Inhibits the Proliferation and Migration of Human and Canine
Breast Tumor Cells’ (grant no. 2505NDLX06), Henan Vocational
College of Agriculture, 2025.5. ‘Clinical Diagnosis and Prevention
Techniques for Canine Breast Tumor in Veterinary Medicine’ (grant
no. 2505RCLX02), Henan Vocational College of Agricultural, 2025.5.
‘Innovative Research Team in Pet Medical Technology’ (grant no.
2505TDLX05), Henan Vocational College of Agricultural, 2025.5.
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
HF and CQ contributed to conceptualization; HF, LH
and TU contributed to methodology; HW, LY and GC contributed to
data analysis; PS, WZ and HL contributed to formal analysis; CQ
contributed to resources, investigation, supervision, project
administration, data curation, validation and funding acquisition;
GD contributed to data curation, visualization and validation; HF
contributed to writing of the original draft and TU contributed to
reviewing and editing. HF and CQ confirm the authenticity of data.
All authors have read and agreed to the published version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
3-MA
|
3-methyladenine
|
|
CCK-8
|
cell counting kit-8
|
|
IVM
|
Ivermectin
|
|
NAC
|
N-Acetyl-L-cysteine
|
|
ROS
|
reactive oxygen species
|
|
THBS1
|
thrombospondin-1
|
References
|
1
|
Kim J, Harper A, McCormack V, Sung H,
Houssami N, Morgan E, Mutebi M, Garvey G, Soerjomataram I and
Fidler- Benaoudia MM: Global patterns and trends in breast cancer
incidence and mortality across 185 countries. Nat Med.
31:1154–1162. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249.
2021.PubMed/NCBI
|
|
3
|
Harbeck N, Penault-Llorca F, Cortes J,
Gnant M, Houssami N, Poortmans P, Ruddy K, Tsang J and Cardoso F:
Breast cancer. Nat Rev Dis Primers. 5:662019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kannan K, Sivakumar S, Venkatesan T and
Ali N: The underlying mechanisms and emerging strategies to
overcome resistance in breast cancer. Cancers (Basel). 17:29382025.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Al Khzem AH, Gomaa MS, Alturki MS, Tawfeeq
N, Sarafroz M, Alonaizi SM, Al Faran A, Alrumaihi LA, Alansari FA
and Alghamdi AA: Drug repurposing for cancer treatment: A
comprehensive review. Int J Mol Sci. 25:124412024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Seyyedabadi B, Babataheri S, Naseri M,
Laher I and Soraya H: Ivermectin and non-parasitic disorders: An
update. Curr Opin Pharmacol. 85:1025742025. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kaur B, Blavo C and Parmar MS: Ivermectin:
A multifaceted drug with a potential beyond anti-parasitic therapy.
Cureus. 16:e560252024.PubMed/NCBI
|
|
8
|
Hayashi A, Kamio K, Miyanaga A, Yoshida K,
Noro R, Matsuda K, Tozuka T, Omori M, Hirao M, Fukuizumi A, et al:
Ivermectin enhances paclitaxel efficacy by overcoming resistance
through modulation of ABCB1 in non-small cell lung cancer.
Anticancer Res. 44:5271–5282. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Zhang J, Feng Q, Jiang X, Yang Y,
Liu W, Xiao J, Feng J, Wang Z, Pan M, et al: Purinergic receptor
nanoimmunoamplifiers potentiate chemoimmunotherapy efficacy in
hepatocellular carcinoma. Biomater Res. 29:02782025. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kuznetsov KO, Safina ER, Gaimakova DV,
Frolova YS, Oganesyan IY, Sadertdinova AG, Nazmieva KA, Islamgulov
AH, Karimova AR, Galimova AM and Rizvanova EV: Metformin and
malignant neoplasms: A possible mechanism of antitumor action and
prospects for use in practice. Probl Endokrinol (Mosk). 68:45–55.
2022.(In Russian). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song C, Jung D, Kendi AT, Rho JK, Kim EJ,
Horn I, Curran GL, Ghattamaneni S, Shim JY, Kang PS, et al:
Metformin prevents tumor cell growth and invasion of human hormone
receptor-positive breast cancer (HR+ BC) Cells via FOXA1
inhibition. Int J Mol Sci. 25:74942024. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Higurashi T and Nakajima A: Metformin and
colorectal cancer. Front Endocrinol (Lausanne). 9:6222018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Basheer HA, Salman NM, Abdullah RM,
Elsalem L and Afarinkia K: Metformin and glioma: Targeting
metabolic dysregulation for enhanced therapeutic outcomes. Transl
Oncol. 53:1023232025. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li JH, Hsin PY, Hsiao YC, Chen BJ, Zhuang
ZY, Lee CW, Lee WJ, Vo TTT, Tseng CF, Tseng SF and Lee IT: A
narrative review: Repurposing metformin as a potential therapeutic
agent for oral cancer. Cancers (Basel). 16:30172024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feng H, He L, Umar T, Wang X, Li W, Zhang
B, Zhu X, Deng G and Qiu C: Synergistic antitumor effects of
ivermectin and metformin in canine breast cancer via PI3K/AKT/mTOR
pathway inhibition. Curr Issues Mol Biol. 47:4032025. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mahfauz M, Yuruker O and Kalkan R:
Repurposing metformin as a potential anticancer agent using in
silico technique. Daru. 32:549–555. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xia Y, Sun M, Huang H and Jin WL: Drug
repurposing for cancer therapy. Signal Transduct Target Ther.
9:922024. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ayub A, Hasan MK, Mahmud Z, Hossain MS and
Kabir Y: Dissecting the multifaceted roles of autophagy in cancer
initiation, growth, and metastasis: From molecular mechanisms to
therapeutic applications. Med Oncol. 41:1832024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Liu S, Jin L, Ma Y and Liu T: NOD2
inhibits the proliferation of esophageal adenocarcinoma cells
through autophagy. J Cancer Res Clin Oncol. 149:639–652. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu Z, Han X, Ou D, Liu T, Li Z, Jiang G,
Liu J and Zhang J: Targeting PI3K/AKT/mTOR-mediated autophagy for
tumor therapy. Appl Microbiol Biotechnol. 104:575–587. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang ZM, Fang ZY, Yang X, Ji XX, Zhao YY,
Lin BY, Weng ZB and Liu EH: Glycyrrhetinic acid ameliorates gastric
mucosal injury by modulating gut microbiota and its metabolites via
Thbs1/PI3K-Akt/p53 pathway. Phytomedicine. 142:1567452025.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong S, Liang S, Cheng Z, Zhang X, Luo L,
Li L, Zhang W, Li S, Xu Q, Zhong M, et al: ROS/PI3K/Akt and
Wnt/β-catenin signalings activate HIF-1α-induced metabolic
reprogramming to impart 5-fluorouracil resistance in colorectal
cancer. J Exp Clin Cancer Res. 41:152022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsai SC, Chang PC, Lin YT, Huang PT, Chen
JY, Lin CS, Wu BN, Chang HM, Wu WJ, Chang CI and Lee CH:
Repurposing of the antipsychotic trifluoperazine induces
SLC7A11/GPX4-mediated ferroptosis of oral cancer via the
ROS/autophagy pathway. Int J Biol Sci. 20:6090–6113. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Corbella E, Fara C, Covarelli F, Porreca
V, Palmisano B, Mignogna G, Corsi A, Riminucci M, Maras B and
Mancone C: THBS1 and THBS2 enhance the in vitro proliferation,
adhesion, migration and invasion of intrahepatic cholangiocarcinoma
cells. Int J Mol Sci. 25:17822024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang M, Zhang K, Zhang Z, Zeng X, Huang
Z, Qin P, Xie Z, Cai X, Ashrafizadeh M, Tian Y and Wei R:
PI3K/AKT/mTOR axis in cancer: From pathogenesis to treatment.
MedComm (2020). 6:e702952025. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bu X, Sha S, Zhang Z, Bian S, Feng S, Li
C, Wang L and Chen H: Thrombospondin 1 aggravates cardiac
remodeling in heart failure with preserved ejection fraction by
inhibiting mitophagy. iScience. 29:1146392026. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sayers EW, Beck J, Bolton EE, Brister JR,
Chan J, Connor R, Feldgarden M, Fine AM, Funk K, Hoffman J, et al:
Database resources of the National Center for Biotechnology
Information in 2025. Nucleic Acids Res. 53:D20–D29. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu A, Wang H, Lv H, Yang H, Li Y and Qian
J: Autophagy inhibitor 3-methyladenine mitigates the severity of
colitis in aged mice by inhibiting autophagy. Exp Gerontol.
201:1126972025. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Galicia-Moreno M, Monroy-Ramirez HC,
Caloca-Camarena F, Arceo-Orozco S, Muriel P, Sandoval-Rodriguez A,
García-Bañuelos J, García-González A, Navarro-Partida J and
Armendariz-Borunda J: A new opportunity for N-acetylcysteine. An
outline of its classic antioxidant effects and its pharmacological
potential as an epigenetic modulator in liver diseases treatment.
Naunyn Schmiedebergs Arch Pharmacol. 398:2365–2386. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cabrera-Serrano AJ, Sánchez-Maldonado JM,
González-Olmedo C, Carretero-Fernández M, Díaz-Beltrán L,
Gutiérrez-Bautista JF, García-Verdejo FJ, Gálvez-Montosa F,
López-López JA, García-Martín P, et al: Crosstalk between autophagy
and oxidative stress in hematological malignancies: Mechanisms,
implications, and therapeutic potential. Antioxidants (Basel).
14:2642025. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Siavoshi A, Taghizadeh M, Dookhe E and
Piran M: Gene expression profiles and pathway enrichment analysis
to identification of differentially expressed gene and signaling
pathways in epithelial ovarian cancer based on high-throughput
RNA-seq data. Genomics. 114:161–170. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang L, Fang T, Hu T, Feng J and Yan P:
Mir-338-3p targeting THBS1 attenuates glioma progression by
inhibiting the PI3K/Akt pathway. Biol Direct. 19:92024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen J, Zhou L, Ye K, Gong J, Wu F, Mo K,
Zhu Y, Chen C and Zhan R: The role of SPI1/VSIG4/THBS1 on
glioblastoma progression through modulation of the PI3K/AKT
pathway. J Adv Res. 71:487–500. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tuoheti K, Bai X, Yang L, Wang X, Cao Y,
Yisha Z, Guo L, Zhan S, Wu Z and Liu T: Forsythiaside A suppresses
renal fibrosis and partial epithelial-mesenchymal transition by
targeting THBS1 through the PI3K/AKT signaling pathway. Int
Immunopharmacol. 129:1116502024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou FL, Hu M, Hu J, Lin T and He YC:
Maslinic acid induces autophagy through PI3K/Akt/mTOR pathway in
human nasopharyngeal carcinoma cells. Chin Tradit Herb Drugs.
51:2481–2485. 2020.
|
|
36
|
Qin P, Li Q, Zu Q, Dong R and Qi Y:
Natural products targeting autophagy and apoptosis in NSCLC: a
novel therapeutic strategy. Front Oncol. 14:13796982024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xin H and Xu W: Effect of licochalcone A
on autophagy in renal cell carcinoma via PI3K/Akt/mTOR signaling
pathway. Zhongguo Zhong Yao Za Zhi. 43:3545–3552. 2018.(In
Chinese). PubMed/NCBI
|
|
38
|
Yang H, Zhou J and Yang X: Effects of
PicrosideII on autophagy of MCF-7 breast cancer cells and its
mechanism. Chin J Clin Pharmacol Ther. 24:535–540. 2019.
|
|
39
|
Malekan M, Dozandeh-Jouybari A, Joudaki N,
Ahangari M, Valadan R, Asgarian-Omran H and Taghiloo S: Inhibition
of PI3K/AKT/mTOR signaling enhances autophagy in HL-60 acute
myeloid leukemia cells: An integrative bioinformatic and in vitro
study. Biochem Biophys Rep. 44:1022202025.PubMed/NCBI
|
|
40
|
Liu Y, Cai X, Liu J, Luo Z, Zhang J, Cao
Z, Ma W, Tang Y, Liu T, Wei H and Yu CY: The role of redox
homeostasis in tumor progression: Implications for cancer therapy.
Acta Biomater. 204:156–186. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ma J, Wu Y, Huang D, Zhou Y, Hou L, Li Y,
Zhong J and Wei X: Averrhoa carambola L. roots DMDD alleviates
myocardial injury in diabetes mellitus mice by regulating
ROS-mediated autophagy pathway. Chin Pharmacol Bull. 37:823–827.
2021.
|
|
42
|
Yu Y, Wang K, Zhu Y, Qi S and Luo L:
Research progress on autophagy regulated tumor radiosensitivity[J].
Journal of Xuzhou Medical University. 42:771–775. 2022.
|















![IVM combined with MET inhibits the
invasive and migratory ability of MCF-7 cells. (A) Representative
Transwell images and (B) quantification of the invasive ability of
MCF-7 cells. (C) Representative scratch images and (D)
quantification of the migration ability in [Ctrl, IVM (6 µM), MET
(6 mM), and IVM + MET] groups of MCF-7 cells. IVM, ivermectin; MET,
metformin, MCF-7, human mammary tumor cell line; Ctrl, control. All
data were measured as mean ± SEM by three independent experiments.
**P<0.01.](/article_images/or/56/1/or-56-01-09136-g01.jpg)