Introduction
Pancreatic cancer (PC) is the third leading cause of
cancer-associated mortality in the United States and the sixth in
China, and it is projected to become the second in the United
States by 2030 (1,2). Due to late diagnosis, early metastasis
and poor prognosis, the overall 5-year survival rate for PC is
<13% (3). Surgical resection
combined with systemic chemotherapy remains the only curative
option or possibility with a chance of long-term survival, but most
patients are ineligible due to presenting with locally advanced
disease or distant metastasis at the time of initial diagnosis
(4–6). FOLFIRINOX and gemcitabine-based
therapies have only modestly improved overall survival, and their
efficacy is limited by multidrug resistance and systemic toxicities
(7,8). Thus, new therapeutic strategies are
urgently needed. Recent advances include gene therapy, molecular
targeted therapy, cellular immunotherapy and natural compounds, all
of which highlight the growing interest in innovative PC treatments
(9–13).
Ferroptosis is an iron-dependent form of programmed
cell death (PCD) characterized by distinct morphological,
biochemical and genetic features that differentiate it from other
types of cell death, including apoptosis, pyroptosis, autophagy and
necroptosis (14). The primary
hallmarks of ferroptosis are the accumulation of iron and lipid
peroxides (15). PC cells exhibit
reprogrammed organelle functions, resulting in increased iron
availability and reactive oxygen species (ROS) production compared
with normal cells; this alteration enhances the intrinsic
vulnerability of these cells to ferroptotic cell death (16). Ferroptosis inducers have shown
considerable potential as innovative therapeutic strategies for
inhibiting tumor growth, enhancing the efficacy of conventional
antitumor agents, and overcoming drug resistance to systemic
therapy, radiotherapy and immunotherapy (17–20).
These observations suggest that inducing ferroptosis is a promising
therapeutic strategy for PC.
Schisandra chinensis (Turcz.) Baill., known
as Wu Wei Zi in Chinese, is a medicinal and edible herb, which is
widely used in traditional Chinese formulations documented in the
Chinese Pharmacopoeia (21).
Schisandrin B (Sch B) is one of the most abundant
dibenzocyclooctadiene lignans in S. chinensis fruits, which
exhibits various beneficial properties, including antitumor,
anti-inflammatory, hepatoprotective, cardioprotective and
neuroprotective effects (22–26).
Previous studies have shown that Sch B exerts antitumor effects by
inducing apoptosis, pyroptosis, autophagy and ROS production in
human cancer (27–29). Additionally, Sch B has synergistic
effects with conventional or targeted antitumor agents, enhancing
clinical efficacy and reducing adverse effects (30–33).
These organ-protective and antitumor properties suggest that Sch B
may be a valuable adjunctive treatment for PC; however, whether Sch
B can trigger ferroptosis in PC and its mechanism remain unclear.
The present study aimed to investigate the anti-proliferative
effects and mechanisms of Sch B against PC in vitro and
in vivo.
Materials and methods
Chemicals and antibodies
Sch B (cat. no. B21327-1g) was purchased from
Shanghai Yuanye Bio-Technology Co., Ltd. Sch B was prepared as a
125 mM stock solution in 100% DMSO [the final concentration of DMSO
in culture medium did not exceed 0.1% (v/v)] and stored in the dark
at −20°C in our laboratory. Ferrostatin-1 (Fer-1; cat. no.
HY-100579), deferoxamine mesylate (DFO; cat. no. HY-B1625),
Z-VAD-FMK (cat. no. HY-16658B), N-acetylcysteine (NAC; cat. no.
HY-B0215) and necrostatin-1 (NEC-1; cat. no. HY-15760) were
purchased from MedChemExpress. Anti-acyl-CoA synthetase long-chain
family member 4 (ACSL4; cat. no. 22401-1-AP), anti-GSH synthetase
(GSS; cat. no. 67598-1-Ig) anti-solute carrier family 3 member 2
(SLC3A2; cat. no. 15193-1-AP), anti-transferrin (TF; cat. no.
17435-1-AP), and anti-α-tubulin (cat. no. 66031-1-Ig) were
purchased from Proteintech Group, Inc. Anti-glutathione (GSH)
peroxidase 4 (GPX4; cat. no. ab125066), anti-γ-glutamyl-cysteine
ligase catalytic subunit (GCLC; cat. no. ab207777), anti-solute
carrier family 7 member 11 (SLC7A11; cat. no. ab307601) and
anti-Ki67 (cat. no. ab16667) were purchased from Abcam.
Cell culture
Human pancreatic ductal adenocarcinoma cell lines
MIA PaCa-2 (cat. no. CRL-1420), PANC-1 (cat. no. CRL-1469) and PL45
(cat. no. CRL-2558) were purchased from American Type Culture
Collection. These cell lines were cultured in DMEM (cat. no.
C11995500BT; Gibco; Thermo Fisher Scientific, Inc.) supplemented
with 1% penicillin-streptomycin (PS; cat. no. 15140122; Gibco;
Thermo Fisher Scientific, Inc.) and 10% fetal bovine serum (FBS;
cat. no. FSP500; Shanghai ExCell Biology, Inc.). Human pancreatic
duct epithelial cells (hTERT-HPNE) were purchased from Guangzhou
Jennio Biotechnology Co., Ltd. and cultured in RPMI-1640 medium
(cat. no. C11875500BT; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 1% PS and 10% FBS. All cells were maintained at
37°C with 5% CO2 in a sterile incubator. All cell lines
tested negative for mycoplasma using the Mycoplasma test kit (cat.
no. G1901; Wuhan Servicebio Technology Co., Ltd.).
Cell viability assay
MIA PaCa-2, PANC-1, PL45 and hTERT-HPNE cells were
seeded in 96-well plates at a density of 3×103
cells/well and incubated overnight. The cells were then treated
with Sch B at concentrations of 0, 7.8, 15.6, 31.3, 62.5 or 125 µM
for 24, 48 or 72 h at 37°C. The control groups received an
equivalent concentration of DMSO (≤0.1%). For inhibitor studies,
the cells were pre-treated with Fer-1 (10 µM), DFO (20 µM),
Z-VAD-FMK (50 µM), NEC-1 (50 µM) or NAC (2 mM) for 4 h at 37°C,
followed by treatment with Sch B (20 µM) for 24 h at 37°C.
Subsequently, 10 µl Cell Counting Kit-8 (CCK-8) reagent (cat. no.
CK04; Dojindo Laboratories, Inc.) dissolved in 100 µl fresh medium
was added to each well and incubated for 2 h. Absorbance was
measured at 450 nm using a microplate reader (Tecan Group,
Ltd.).
Colony formation assay
MIA PaCa-2 and PANC-1 cells were seeded in 6-well
plates at a density of 1×103 cells/well and incubated
overnight. The cells were then treated with Sch B (5, 10 or 20 µM)
or vehicle control (<0.1% DMSO) for 24 h at 37°C. Subsequently,
the cells were cultured for 10 days, with the medium changed every
3 days. Colonies were then fixed in 4% paraformaldehyde for 10 min
at room temperature and stained with 0.1% crystal violet for 15 min
at room temperature, after which, images were captured under an
inverted light microscope (Zeiss AG). A colony was defined as a
cluster of ≥50 cells and the number of colonies was semi-quantified
using ImageJ version 1.4.3 software (National Institute of
Health).
EdU assay
MIA PaCa-2 and PANC-1 cells were seeded in 8-well
chamber slides at a density of 9×103 cells/well and were
incubated overnight. The cells were then treated with Sch B (5, 10
or 20 µM) or vehicle control (<0.1% DMSO) for 24 h at 37°C. For
inhibitor experiments, the cells were pre-treated with Fer-1 (10
µM) for 4 h followed by treatment with Sch B (20 µM) for 24 h at
37°C. Subsequently, 500 µl EdU working solution (cat. no. C10310;
Guangzhou RiboBio Co., Ltd.) was added to each well and incubated
for 2 h at 37°C. The cells were then permeabilized with 0.5% Triton
X-100 for 15 min at room temperature and were stained with Apollo
567 for 30 min at room temperature. After washing, the slides were
mounted with DAPI-containing Fluoroshield (cat. no. ab104139;
Abcam). Images of EdU-positive cells were captured using a confocal
microscope (Zeiss AG).
Lactate dehydrogenase (LDH) release
measurement
Cell death was measured using the LDH cytotoxicity
assay kit (cat. no. C0017; Beyotime Biotechnology). MIA PaCa-2 and
PANC-1 cells were seeded in 35-mm dishes at a density of
9×104 cells/dish and incubated overnight. The cells were
then treated with Sch B (5, 10 or 20 µM) or vehicle control
(<0.1% DMSO) for 24 h at 37°C. For inhibitor experiments, the
cells were pre-treated with Fer-1 (10 µM) for 4 h followed by
treatment with Sch B (20 µM) for 24 h at 37°C. After treatment, the
culture medium was collected and centrifuged at 800 × g for 5 min
at 4°C to remove cell debris. Subsequently, 120 µl cell culture
supernatants were transferred to a new 96-well plate and mixed with
60 µl LDH working solution. After incubation for 30 min at 37°C in
the dark, the absorbance was measured at 490 nm using a microplate
reader. The percentage of cell death was calculated as follows:
(treated cell LDH-control cell LDH)/(maximum LDH release-control
cell LDH) ×100.
Trypan blue staining
The Trypan blue assay (cat. no. C0040; Beijing
Solarbio Science & Technology Co., Ltd.) was performed to
confirm the results of the LDH assay. MIA PaCa-2 and PANC-1 cells
were seeded in 35-mm dishes at a density of 9×104
cells/dish and incubated overnight. The cells were then treated
with Sch B (5, 10 or 20 µM) or vehicle control (<0.1% DMSO) for
24 h at 37°C. Subsequently, the cells were stained with 0.04%
Trypan blue solution for 3 min at room temperature. The unstained
(viable) and blue-stained (dead) cells were counted using an
automatic cell counter (Guangzhou BodBoge Technology Co., Ltd.) and
the percentage of dead cells was calculated.
Detection of intracellular iron
Intracellular Fe2+ levels were measured
using the FerroOrange fluorescent probe (cat. no. F374; Dojindo
Laboratories, Inc.). MIA PaCa-2 and PANC-1 cells were seeded in
8-well chamber slides at a density of 9×103 cells/well
and incubated overnight. The cells were then treated with Sch B (5,
10 or 20 µM) or vehicle control (<0.1% DMSO) for 24 h at 37°C.
For inhibitor experiments, the cells were pre-treated with Fer-1
(10 µM) for 4 h followed by treatment with Sch B (20 µM) for 24 h
at 37°C. Subsequently, the cells were stained with 1 µM FerroOrange
in serum-free medium for 30 min at 37°C in the dark. Fluorescence
intensity was acquired using a confocal microscope.
Transmission electronic
microscopy
MIA PaCa-2 and PANC-1 cells were seeded in 150-mm
dishes at a density of 6×105 cells/dish and incubated
overnight. The cells were then treated with Sch B (20 µM) or
vehicle control (<0.1% DMSO) for 24 h at 37°C. Subsequently, the
cells were harvested and fixed in a 2.5% glutaraldehyde solution
(pH 7.4) for 6 h at 4°C. After washing with 0.1 M phosphate buffer
(pH 7.2), the samples were postfixed with 1% OsO4 for 4
h at 4°C. Following dehydration, the samples were embedded in
Epon-Araldite resin and polymerized at 60°C for 48 h. Semithin
sections (1 µm) were stained with toluidine blue for positioning,
after which, ultrathin sections (70 nm) were collected on copper
grids, stained with 2% uranyl acetate for 8 min at room
temperature, followed by 2.7% lead citrate for 8 min at room
temperature, and examined under a transmission electron microscope
(Thermo Fisher Scientific, Inc.).
Measurement of GSH and malondialdehyde
(MDA) content
The levels of GSH and MDA were measured using ELISA
kits (cat. nos. S0052 and S0131S; Beyotime Biotechnology, Inc.)
according to the manufacturer's protocol. MIA PaCa-2 and PANC-1
cells were seeded in 150-mm dishes at a density of 6×105
cells/dish and incubated overnight. The cells were then treated
with Sch B (5, 10 or 20 µM) or vehicle control (<0.1% DMSO) for
24 h at 37°C. For inhibitor experiments, the cells were pre-treated
with Fer-1 (10 µM) for 4 h followed by treatment with Sch B (20 µM)
for 24 h at 37°C. GSH and MDA levels in cell lysates were measured
using a microplate reader.
Determination of lipid
peroxidation
Lipid peroxidation was assessed using the C11-BODIPY
(581/591) assay (cat. no. L267; Dojindo Laboratories, Inc.). MIA
PaCa-2 and PANC-1 cells were seeded in 8-well chamber slides at a
density of 9×103 cells/well and incubated overnight. The
cells were then treated with Sch B (5, 10 or 20 µM) or vehicle
control (<0.1% DMSO) for 24 h at 37°C. For inhibitor
experiments, the cells were pre-treated with Fer-1 (10 µM) for 4 h
followed by treatment with Sch B (20 µM) for 24 h at 37°C.
Subsequently, the cells were incubated with 2 µM C11-BODIPY
(581/591) solution for 30 min at 37°C. Fluorescence intensity was
measured using a confocal microscope.
Drug affinity responsive target
stability (DARTS) analysis
PANC-1 cells were treated with Sch B (20 µM) or
vehicle control (<0.1% DMSO) for 24 h at 37°C and then lysed
using M-PER mammalian protein extraction reagent (cat. no. 78505;
Thermo Fisher Scientific, Inc.) with protease inhibitor cocktail
(cat. no. HY-K0010; MedChemExpress) on ice for 10 min. The lysates
were centrifuged at 12,000 × g for 20 min at 4°C, after which, the
supernatants were collected and mixed with 10X TNC buffer (500 mM
Tris-HCl, pH 8.0; 500 mM CaCl2; 100 mM NaCl). Protein
concentration was then quantified using a BCA assay. The lysates (4
µg/µl) in 1X TNC buffer were digested with pronase (0.25 µg/µl;
cat. no. 63163824; Roche) at a pronase/protein ratio of 1:200 for
30 min at room temperature and the reaction was terminated by
adding 5X SDS-PAGE loading buffer. Subsequently, the proteins were
separated by SDS-PAGE and visualized by Coomassie staining;
specific protein bands were excised for label-free liquid
chromatography-tandem mass spectrometry (LC-MS/MS) analysis.
LC-MS/MS analysis
Nanoflow LC-MS/MS analysis of peptides was performed
using a quadrupole Orbitrap Exploris™ 480 Mass
Spectrometer (Thermo Fisher Scientific, Inc.) equipped with an EASY
nLC 1200 ultra-high pressure system (Thermo Fisher Scientific,
Inc.) via a nano-electrospray ion source. Briefly, peptides (500
ng) were loaded on a 25-cm column (150 µm inner diameter, packed
with ReproSil-Pur C18-AQ 1.9-µm silica beads; Guangzhou Promegene
Biotechnology Co., Ltd.). The peptides were eluted over a 60-min
gradient at a flow rate of 600 nl/min using buffer B (80%
acetonitrile, 0.1% formic acid) as follows: 8–12% B for 5 min,
12–30% B for 30 min, 30–40% B for 9 min, 40–95% B for 1 min,
followed by incubation with 95% B for 15 min.
The mass spectrometer was operated in positive ion
mode with the FAIMS Pro™ Interface (Thermo Fisher
Scientific, Inc.) cycling between compensation voltages of-45 V and
−65 V every 1 sec. Full-scan MS spectra (350–1,200 m/z, 120,000
resolution) were acquired in the Orbitrap analyzer with an
automatic gain control (AGC) target of 300%, and a maximum
injection time of 50 msec. The most intense ions from the full scan
were isolated with an isolation width of 1.6 m/z and fragmented
using higher-energy collisional dissociation with a normalized
collision energy of 33%. MS/MS spectra were collected in the
Orbitrap analyzer (15,000 resolution) with an AGC target of 75% and
a maximum injection time of 22 msec.
The resulting MS/MS data were processed using
Proteome Discoverer version 2.4 (Thermo Fisher Scientific, Inc.).
Tandem mass spectra were searched against the UniProt human protein
database (https://www.uniprot.org/taxonomy/9606). Trypsin/P was
specified as a cleavage enzyme allowing up to two missing
cleavages. The precursor mass tolerance was ±15 ppm, and fragment
mass tolerance was ±0.02 Da. Carbamidomethyl on cysteine was
defined as a fixed modification, and acetylation at the protein
N-terminus and oxidation on methionine were specified as variable
modifications. The peptide ion score threshold was set to >20
and high confidence in peptides was required. Search outcomes
underwent filtering via a linear discriminant algorithm to achieve
an estimated false discovery rate of 1%. Candidate proteins were
selected based on the following criteria: i) Intensity ratio
(compound/control) ≥2.0, ii) a molecular weight of ~70 kDa
(matching the excised band) and iii) protein score ≥10.
Molecular docking
The protein structure of ACSL4 was initially
predicted using AlphaFold3 (https://github.com/deepmind/alphafold) and was then
processed with AutoDockTools 1.5.6 (34) to preserve its original charges and
generate a pdbqt file for docking. The 2D structure of Sch B was
downloaded from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/compound/108130).
Structural optimization was performed using the MOPAC program
(https://openmopac.net/) and PM3 atomic charges
were calculated. The structure of Sch B was also prepared using
AutoDock Tools 1.5.6 to create a pdbqt file. Docking simulations
were carried out using AutoDock 4.2.6 (34). A grid box of 100×100×100 points was
defined with the center at x=71.637, y=70.446, z=104.536. The grid
spacing was 0.375 Å and the exhaustiveness was set to 50. Other
docking parameters were set to default and are therefore not
listed.
RNA sequencing
PANC-1 cells were seeded in 150-mm dishes at a
density of 6×105 cells/dish. The cells were then treated
with Sch B (20 µM) or vehicle control (<0.1% DMSO) for 24 h at
37°C. Total RNA was extracted using RNeasy plus Mini Kit (cat. no.
74134; Qiagen, Inc.) according to the manufacturer's protocol. RNA
quality and RNA integrity number (>8.0) were determined using an
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.) with the
Agilent RNA 6000 Nano Kit (cat. no. 5067-1511; Agilent
Technologies, Inc.). RNA-seq libraries were constructed using the
NEBNext® Ultra™ RNA Library Prep Kit (cat.
no. E7770; New England BioLabs, Inc.). The libraries were then
sequenced on an Illumina NovaSeq 6000 System platform (Illumina,
Inc.) with 150 bp paired-end reads. Prior to sequencing, the RNA
concentration of the library was determined using a
Qubit®2.0 fluorometer (Thermo Fisher Scientific, Inc.)
with a Qubit RNA Assay Kit (cat. no. Q32852; Thermo Fisher
Scientific, Inc.) and was then diluted to 4 nM. Insert size was
assessed using the Agilent Bioanalyzer 2100 system, and qualified
insert size was accurately quantified using StepOnePlus™
Real-Time PCR System (Thermo Fisher Scientific, Inc.; library valid
concentration >10 nM). mRNA libraries were sequenced on the
Illumina sequencing platform by Guangzhou Promegene Biotechnology
Co., Ltd.
For data analysis, differential expression analysis
was performed using DESeq2 package v1.20.0 (https://bioconductor.org/packages/DESeq2) and the
heatmap was generated using the cluster package v2.0.3 (https://cran.r-project.org/web/packages/cluster/)
in R v3.6.2 (https://www.r-project.org/). Kyoto Encyclopedia of
Genes and Genomes (KEGG) enrichment analysis was performed using
the R software clusterProfiler package v3.12.0 (https://bioconductor.org/packages/release/bioc/html/clusterProfiler.html).
The raw sequencing data have been deposited into a publicly
available database.
Western blotting (WB)
MIA PaCa-2 and PANC-1 cells were seeded in 150-mm
dishes at a density of 6×105 cells/dish and incubated
overnight. The cells were then treated with Sch B (5, 10 or 20 µM)
or vehicle control (<0.1% DMSO) for 24 h at 37°C. For inhibitor
experiments, the cells were pre-treated with Fer-1 (10 µM) for 4 h
followed by treatment with Sch B (20 µM) for 24 h at 37°C. Total
cellular protein was extracted using RIPA lysis buffer (cat. no.
R0010; Beijing Solarbio Science & Technology Co., Ltd.) on ice
for 30 min and quantified with a BCA assay kit (cat. no. PC0020;
Beijing Solarbio Science & Technology Co., Ltd.). The lysates
(30 µg) were then separated by SDS-PAGE on 4–15% gels, followed by
transfer of proteins to PVDF membranes. After blocking with 5%
non-fat milk for 1 h at room temperature, the membranes were
incubated with the following primary antibodies overnight at 4°C:
Anti-ACSL4 (1:5,000), anti-GSS (1:5,000), anti-SLC3A2 (1:5,000),
anti-TF (1:1,000), anti-α-tubulin (1:20,000), anti-GPX4 (1:1,000),
anti-SLC7A11 (1:1,000) and anti-GCLC (1:1,000). The membranes were
then washed three times with Tris-buffered saline containing 0.1%
Tween-20 and were subsequently incubated with HRP-conjugated
secondary antibodies (1:10,000; cat. nos. 31430 and 31460; Thermo
Fisher Scientific, Inc.) for 2 h at room temperature. Protein bands
were visualized using an ECL reagent (cat. no. P1010-500; Applygen
Technologies, Inc.). Protein band intensities were semi-quantified
using ImageJ version 1.4.3 software and normalized to the
corresponding α-tubulin loading control.
Cell transfection
MIA PaCa-2 and PANC-1 cells were seeded in 6-well
plates at a density 2×106 cells/well and incubated
overnight. When the cells reached 50–60% confluence, small
interfering (si) RNAs (150 pmol) were transiently introduced into
the cells for 6 h at 37°C using Lipofectamine® RNAiMAX
reagent (cat. no. 13778150; Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. A total of 48 h
post-transfection, the cells were harvested for WB to verify ACSL4
knockdown efficiency. Cell viability assay, EdU assay, LDH assay
and lipid peroxidation assay were carried out as functional
experiments. The target and negative control siRNAs were procured
from Shanghai GenePharma Co., Ltd. The sequences were as follows:
ACSL4 siRNA, 5′-GGGUUGAUAUCUGCAAUAATT-3′ (sense),
5′-UUAUUGCAGAUAUCAACCCTT-3′ (antisense). Negative control siRNA,
5′-UUCUCCGAACGUGUCACGUTT-3′ (sense), 5′-ACGUGACACGUUCGGAGAATT-3′
(antisense).
Experiments in a xenograft mouse
model
Male Balb/c-nude mice (age, 4–5 weeks; weight,
18.5–22.8 g) were purchased from GemPharmatech Co., Ltd. All mice
were housed under specific pathogen-free conditions at the
Laboratory Animal Center of Qingyuan People's Hospital, The Sixth
Affiliated Hospital of Guangzhou Medical University (Qingyuan,
China). The housing conditions were as follows: Temperature,
20–26°C; humidity, 40–70%; 12-h light/dark cycle; standard rodent
chow diet and autoclaved water provided ad libitum. Before
the experiment, the mice were acclimated for 1 week. All animal
procedures were ethically reviewed and approved by the Ethics
Committee on Laboratory Animal Care of The Affiliated Qingyuan
Hospital (Qingyuan People's Hospital) of Guangzhou Medical
University (approval no. LAEC-2023-027). The study was performed in
strict compliance with the UK Animals (Scientific Procedures) Act
1986 (https://www.legislation.gov.uk/ukpga/1986/14/contents)
and its associated guidelines. PANC-1 cells (5×106 cells
in 100 µl saline) were injected subcutaneously into the right flank
of the mice. Mice were monitored by visual observation and
palpation until tumors were detected. On day 10 post-injection, the
mice were randomly divided into three treatment groups (n=5
mice/group): Model group (20% SBE-β-CD), a low-dose Sch B group (50
mg/kg) and a high-dose Sch B group (100 mg/kg). Mice received daily
oral gavage of Sch B or vehicle for 6 weeks. The body weight and
tumor volume were recorded weekly. Tumor volume was calculated
using the formula: V=1/2 × (longest axis) × (shortest
axis)2.
At the end of the study, mice were euthanized by
CO2 exposure at an air displacement rate of 30%/min
following anesthesia with isoflurane (2.5% for induction and 1.5%
for maintenance in oxygen) to ensure humane conditions. Death was
confirmed by cessation of heartbeat and respiration, absence of
reflexes and limb paralysis. Humane endpoint criteria followed the
Laboratory Animal Guidelines for euthanasia in China (GB/T
39760-2021; http://std.samr.gov.cn/gb/search/gbDetailed?id=BD89DE8E080A3D08E05397BE0A0A4FAD),
including a maximum tumor diameter exceeding 15 mm or tumor volume
exceeding 1,500 mm3, weight loss exceeding 20% of
initial body weight, and an inability to access food or water, as
well as difficulties in movement or respiration. No animals
required euthanasia or died before the end of the study.
LSL-KrasG12D/+;
Trp53flox/+; Pdx1-Cre (KPC) mouse model
LSL-KrasG12D/+ (3 male mice; age, 8
weeks; weight, 24.3–26.7 g), Pdx1-Cre (3 male mice; age, 10 weeks;
weight, 24.6–25.8 g) and Tp53flox/flox (2 male mice and
2 female mice; age 6 weeks; weight, 21.6–24.5 g) strains on a
C57BL/6J background were obtained from Shanghai Model Organisms
Center, Inc. The housing conditions were as follows: Temperature,
20–26°C; humidity, 40–70%; 12-h light/dark cycle; standard rodent
chow diet and autoclaved water provided ad libitum. Before
the experiment, the mice were acclimated for 1 week. To generate
KPC mice, LSL-KrasG12D/+ mice were first crossed with
Trp53flox/flox mice to generate
LSL-KrasG12D/+; Trp53flox/+ offspring. These
offspring were then crossed with Pdx1-Cre mice to generate KPC
mice, which were genotyped by PCR. At weaning (21–28 days old), KPC
mice were anesthetized with isoflurane (2.5% for induction and 1.5%
for maintenance in oxygen). A 1–2 mm segment of the tail tip was
excised using sterile scissors, after which, the incision was
pressed against a sterile gauze pad to achieve hemostasis. For
post-operative analgesia, buprenorphine hydrochloride (0.05 mg/kg,
subcutaneous) was administered immediately after the procedure, and
the mice were placed on a heating pad for recovery. Genomic DNA was
extracted from mouse tail samples using a rapid DNA isolation kit
(cat. no. NE0400; Leagene; Beijing Regen Biotechnology Co., Ltd.)
according to the manufacturer's protocol. PCR was performed in a
20-µl reaction mixture containing 10 µl 2X Taq Master Mix (cat. no.
P112-02; Vazyme, Inc.), 0.5 µl each primer (10 µM), 7 µl
ddH2O and 2 µl DNA template. The thermal cycling
conditions comprised an initial denaturation step at 94°C for 3
min; 35 cycles of denaturation at 94°C for 30 sec, annealing at
58°C for 30 sec and extension at 72°C for 30 sec; followed by a
final extension at 72°C for 5 min. The PCR products were
electrophoresed on a 1.5% agarose gel (cat. no. BY-R0100; Biowest,
Inc.) with Goldview nucleic acid stain (cat. no. AG11915; Hunan
Accurate Bio-Medical Technology Co., Ltd.) and visualized under UV
light. The primer sequences are listed in Table SI. All animal procedures were
ethically reviewed and approved by the Ethics Committee on
Laboratory Animal Care of The Affiliated Qingyuan Hospital
(Qingyuan People's Hospital) of Guangzhou Medical University
(approval no. LAEC-2021-024). The study was performed in strict
accordance with the UK Animals (Scientific Procedures) Act 1986 and
its associated guidelines. KPC mice (age, 10 weeks; weight,
20.0–27.8 g; 6 male mice and 4 female mice) were randomly divided
into two treatment groups (n=5 mice/group): A model group (20%
SBE-β-CD) and a Sch B group (100 mg/kg). Mice received daily oral
gavage of Sch B or vehicle for 8 weeks and body weight was
monitored weekly. Tumor volume was calculated using the formula:
V=1/2 × (longest axis) × (shortest axis)2.
At the end of the study, mice were euthanized by
CO2 exposure at an air displacement rate of 30%/min
following anesthesia with isoflurane (2.5% for induction and 1.5%
for maintenance in oxygen) to ensure humane conditions. Death was
confirmed by cessation of heartbeat and respiration, absence of
reflexes and limb paralysis. Humane endpoint criteria included the
emergence of hemorrhagic abdominal ascites, severe cachexia, weight
loss exceeding 20% of initial body weight, and inability to access
food or water, as well as difficulties in movement or respiration.
Mice were observed daily for signs of inactivity and the presence
of abdominal ascites. Before the end of study, two KPC mice in the
model group developed hemorrhagic abdominal ascites and were
euthanized based on the veterinarian's recommendation.
Biochemical analysis
At the end of the study, ~200 µl blood samples were
immediately collected from Balb/c-nude mice via cardiac puncture
following CO2-induced euthanasia. After clotting at room
temperature for 30 min, serum was obtained by centrifuging the
blood at 1,000 × g for 10 min at 4°C and was stored at −80 until
further use. Serum samples were used for biochemical analysis. Six
biochemical indices, including alanine aminotransferase (ALT),
aspartate aminotransferase (AST), creatine kinase (CK), LDH,
creatinine (CREA) and uric acid (UA) (cat. nos. 105-020579,
105-020580, 105-020585, 105-00004A, 105-020587 and 105-00023A;
Shenzhen Mindray Bio-Medical Electronics Co., Ltd.), were
determined according to the manufacturer's protocol using an
automatic biochemical analyzer (Shenzhen Mindray Bio-Medical
Electronics Co., Ltd.) to evaluate the effects of Sch B on liver,
heart and kidney function.
Histological analysis and
immunohistochemistry
Tumor tissues and major organs were fixed in 4%
paraformaldehyde for 24 h at room temperature, embedded in paraffin
and sectioned into 3-µm slices. Following deparaffinization and
rehydration, the sections were stained with hematoxylin for 10 min
and eosin for 10 sec at room temperature, then dehydrated and
mounted for examination. For immunohistochemistry, the
paraffin-embedded sections were deparaffinized and rehydrated,
followed by immersion in citrate buffer (pH 6.0) at 100°C for 2.5
min for antigen retrieval. Endogenous peroxidase activity was
blocked with 3% hydrogen peroxide for 30 min at room temperature.
The sections were then permeabilized with 0.5% Triton X-100
solution for 10 min and blocked with 10% normal goat serum (cat.
no. SL038; Beijing Solarbio Science & Technology Co., Ltd.) for
30 min at room temperature. Subsequently, the sections were
incubated with primary antibodies against ACSL4 (1:500) and Ki67
(1:200; cat. no. ab16667; Abcam) overnight at 4°C. After rinsing,
the sections were incubated with an undiluted HRP-conjugated
secondary antibody (cat. no. PR30009, Proteintech Group, Inc.) for
2 h at room temperature. The reaction was visualized using DAB
(cat. no. PK001; Abcarta, Inc.) for 0.5–1 min followed by
counterstaining with hematoxylin for 5 min at room temperature.
Finally, the sections were sealed using an automatic sealing
machine (Tissue-Tek® Film; Sakura Finetek Japan Co.,
Ltd.), and images were captured using a light microscope (Olympus
Corporation) and semi-quantitatively analyzed with ImageJ version
1.4.3 software.
Statistical analysis
All data are presented as mean ± standard error of
the mean. All in vitro experiments were performed three
times and representative experiments are shown. Comparisons between
two groups were performed by unpaired Student's t-test. Multiple
comparisons were analyzed using one-way analysis of variance
followed by Dunnett's post hoc test. Data were analyzed using
GraphPad Prism 8 (Dotmatics). P<0.05 was considered to indicate
a statistically significant difference.
Results
Sch B inhibited the proliferation of
PC cells and induces cell death
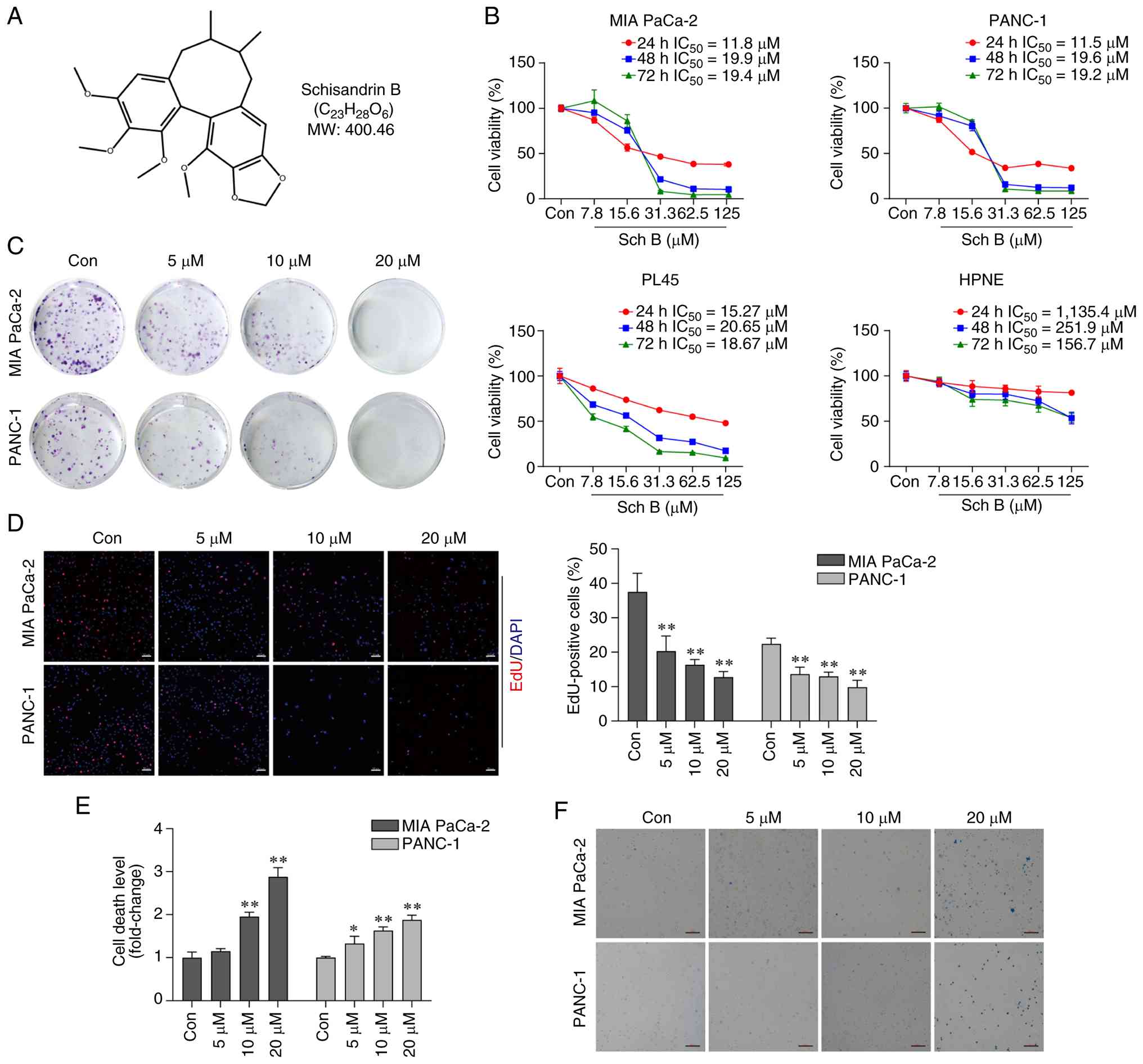
The chemical structure of Sch B is illustrated in
Fig. 1A. PC cell lines (MIA PaCa-2,
PANC-1 and PL45) and a normal pancreatic epithelial cell line
(hTERT-HPNE) were treated with varying concentrations of Sch B for
the indicated times, and cell proliferation was assessed by CCK-8,
colony formation, and EdU assays. The CCK-8 results showed that Sch
B significantly inhibited the viability of all PC cell lines in a
concentration- and time-dependent manner; the half maximal
inhibitory concentration values at 24 h were 11.8 µM for MIA PaCa-2
cells, 11.5 µM for PANC-1 cells and 15.27 µM for PL45 cells
(Fig. 1B). Notably, hTERT-HPNE
cells were substantially less sensitive than the cancer cells.
Because MIA PaCa-2 and PANC-1 exhibited greater sensitivity to Sch
B, they were selected for all subsequent mechanistic and functional
experiments. Colony formation assays (Fig. 1C) and EdU staining (Fig. 1D) confirmed these findings. To
determine whether Sch B induces cell death, LDH release and Trypan
blue staining assays were performed. The LDH assay indicated that
Sch B significantly increased the cell death rate in a
concentration-dependent manner (Fig.
1E). Similar results were obtained from the Trypan blue
staining assay (Fig. 1F). Together,
these findings indicated that Sch B may effectively inhibit PC cell
proliferation and induce cell death.
Cell death induced by Sch B is
associated with ferroptosis
To investigate the mechanisms underlying Sch
B-induced cell death in PC cells, RNA sequencing was performed on
PANC-1 cells treated with 20 µM Sch B. A total of 4,101 significant
differentially expressed genes (DEGs) were identified between the
Sch B group and the control group; among them, 2,525 DEGs were
upregulated and 1,576 DEGs were downregulated (Fig. 2A and B). KEGG pathway enrichment
analysis revealed that the upregulated DEGs were most significantly
enriched in the ‘ferroptosis’ pathway (Fig. 2C). Downregulated DEGs were
predominantly enriched in ‘Oxidative phosphorylation’, ‘Chemical
carcinogenesis-reactive oxygen species’, ‘Alanine, aspartate and
glutamate metabolism’, and ‘Glutathione metabolism’ pathways; these
pathways are closely associated with ferroptosis (Fig. 2D). To further investigate the mode
of cell death induced by Sch B, specific inhibitors of cell death
were utilized in MIA PaCa-2 and PANC-1 cells. As shown in Fig. 2E and F, the ferroptosis inhibitors
Fer-1 and DFO, the apoptosis inhibitor Z-VAD-FMK and the ROS
scavenger NAC effectively reversed the inhibitory effects of Sch B
on cell viability. By contrast, the necroptosis inhibitor NEC-1 did
not affect Sch B-induced cell viability. These findings indicated
that ferroptosis may be partially involved in Sch B-induced PC cell
death.
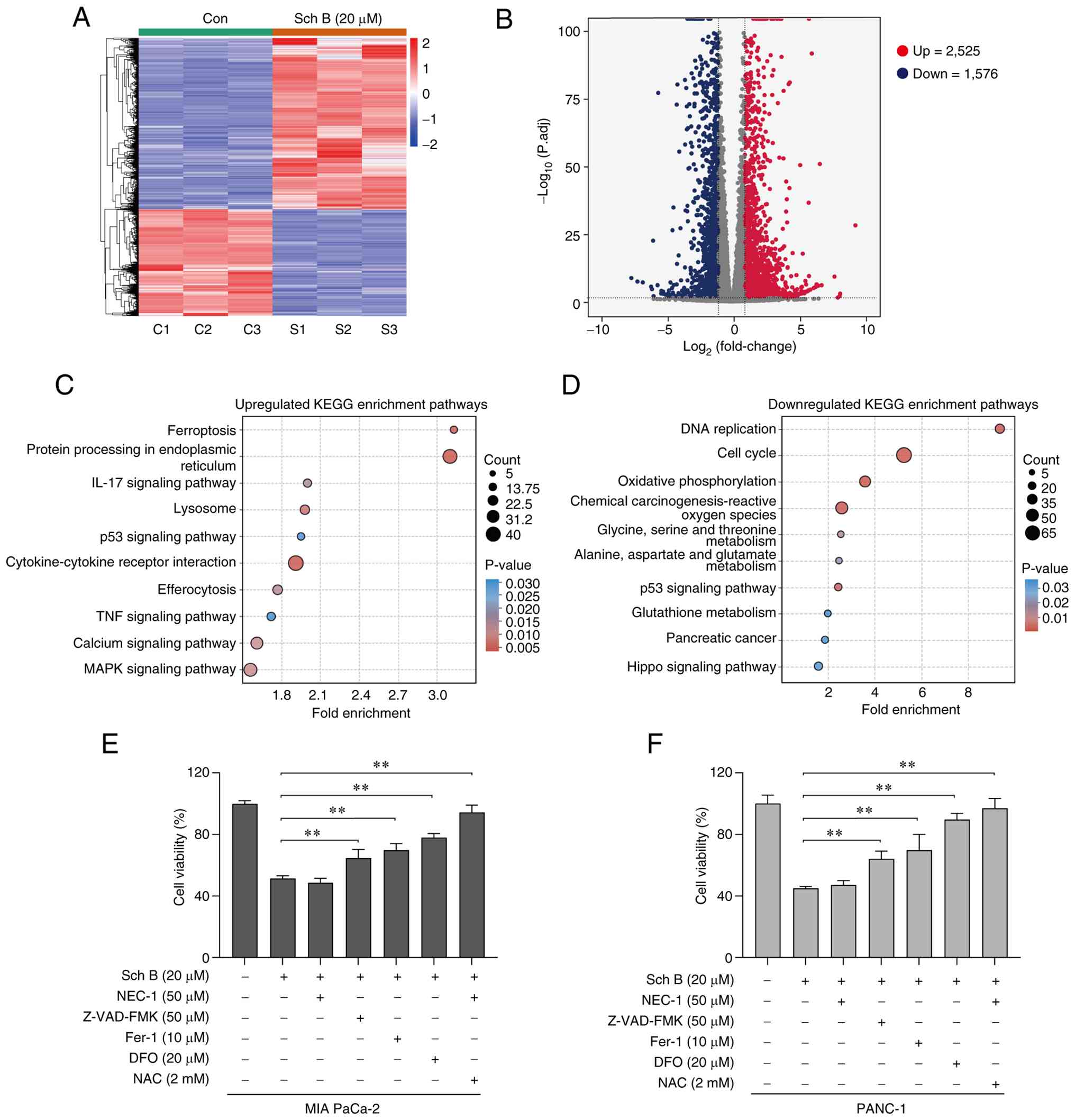 | Figure 2.Cell death induced by Sch B is
associated with ferroptosis. (A) Heat map and (B) volcano plot of
the DEGs in PANC-1 cells between the Sch B group and the control
group. KEGG annotation and enrichment analysis of the (C)
upregulated and (D) downregulated DEGs in PANC-1 cells. Viability
of (E) MIA PaCa-2 and (F) PANC-1 cells treated with Sch B (20 µM)
with or without inhibitors, NEC-1 (50 µM), Z-VAD-FMK (50 µM), Fer-1
(10 µM), DFO (20 µM) and NAC (2 mM), was assessed using Cell
Counting Kit-8 assay. **P<0.01. Con, control; DEG,
differentially expressed gene; DFO, deferoxamine mesylate; Fer-1,
ferrostatin-1; KEGG, Kyoto Encyclopedia of Genes and Genomes; NAC,
N-acetylcysteine; NEC-1, necrostatin-1; Sch B, Schisandrin B. |
Sch B triggers ferroptosis in PC
cells
Intracellular iron overload and lipid peroxidation
of membranes are hallmark features of ferroptosis. The results of
FerroOrange staining and the C11-BODIPY assay showed that Sch B
dose-dependently increased the levels of Fe2+ (Fig. 3A) and lipid peroxidation (Fig. 3B) in both MIA PaCa-2 and PANC-1
cells. Sch B also significantly elevated MDA, a marker of lipid
peroxidation, while concurrently reducing GSH levels (Fig. 3D). Morphologically, ferroptotic
cells exhibit distinct ultrastructural changes in the mitochondria;
therefore, subcellular changes in Sch B-treated cells were
observed. Transmission electron microscopy revealed that
mitochondria in the Sch B group appeared rounder and smaller, with
denser membranes and fewer cristae, which is a typical feature of
ferroptosis (Fig. 3C).
Subsequently, the expression levels of proteins involved in iron
metabolism, GSH metabolism and lipid peroxidation were assessed.
The results showed that Sch B upregulated the expression levels of
ACSL4 and SLC3A2, and downregulated GPX4, GCLC and GSS, whereas TF
and SLC7A11 exhibited no notable changes in PC cells (Fig. 3E). Collectively, these findings
indicated that Sch B may induce ferroptosis in PC cells, at least
in part by increasing iron availability and promoting lipid
peroxidation, while suppressing glutathione synthesis and
antioxidant defenses.
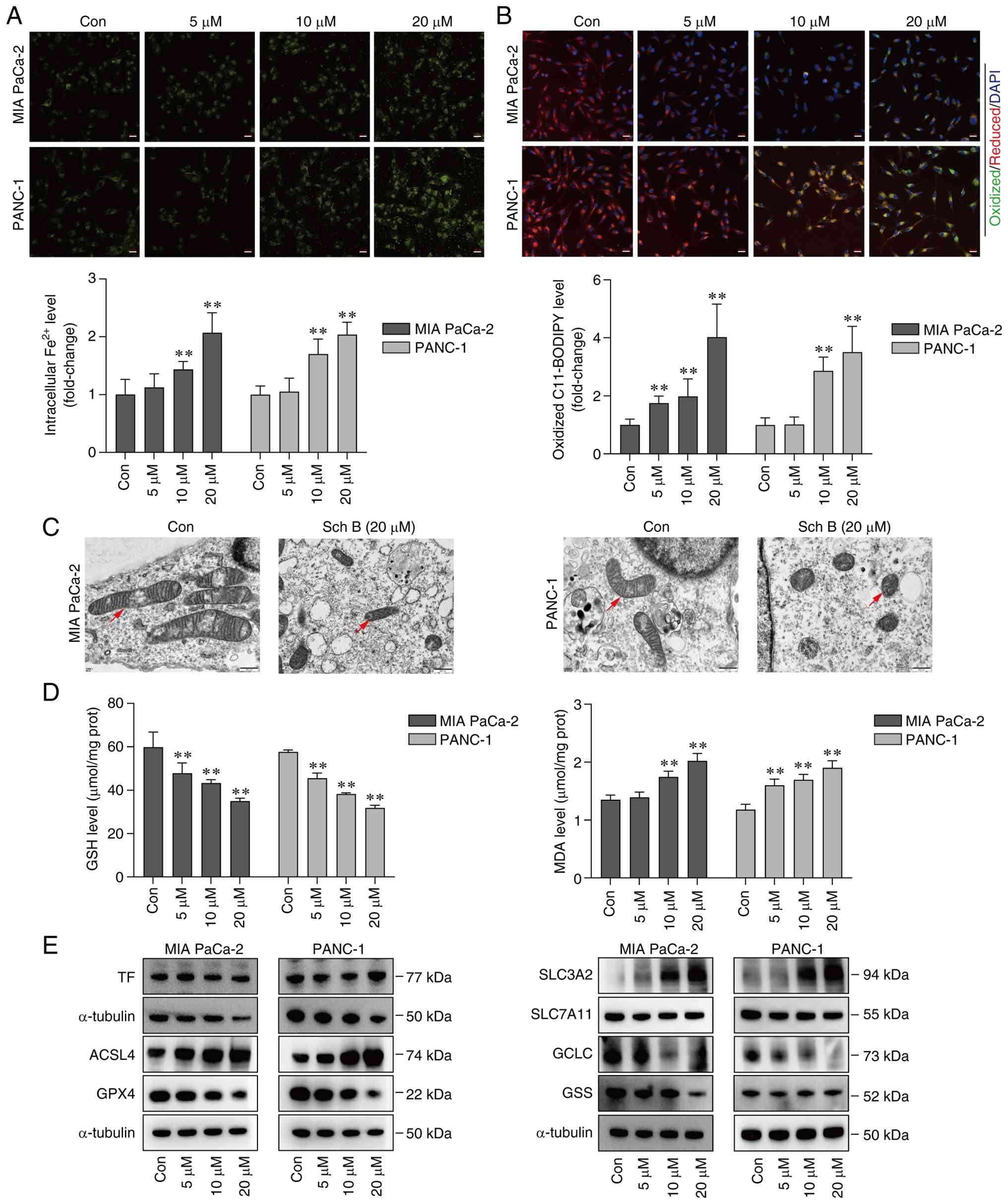 | Figure 3.Sch B triggers ferroptosis in
pancreatic cancer cells. (A) Intracellular Fe2+ levels
in MIA PaCa-2 and PANC-1 cells were detected by FerroOrange probe.
Scale bars, 20 µm. (B) Lipid peroxidation levels of MIA PaCa-2 and
PANC-1 were detected by C11-BODIPY 581/591 fluorescent probe. Scale
bars, 20 µm. (C) Transmission electron microscopy imaging of
mitochondrial ultrastructure in MIA PaCa-2 and PANC-1 cells was
conducted. Scale bars, 500 nm. Red arrowheads indicate
mitochondria. (D) Levels of GSH and MDA in MIA PaCa-2 and PANC-1
cells were measured using ELISA kits. (E) Expression levels of
ferroptosis-related proteins in MIA PaCa-2 and PANC-1 cells were
detected by western blotting. **P<0.01 vs. Con. ACSL4, acyl-CoA
synthetase long-chain family member 4; Con, control; GCLC,
γ-glutamyl-cysteine ligase catalytic subunit; GPX4, glutathione
peroxidase 4; GSH, glutathione; GSS, glutathione synthetase; MDA,
malondialdehyde; Sch B, Schisandrin B; SLC3A2, solute carrier
family 3 member 2; SLC7A11, solute carrier family 7 member 11; TF,
transferrin. |
Blocking ferroptosis reverses Sch
B-induced cell death
To elucidate the involvement of ferroptosis in the
antitumor effects of Sch B, the ferroptosis inhibitor Fer-1 was
used in vitro. Fer-1 pre-treatment effectively prevented Sch
B-induced cell death (Fig. 4A) and
significantly diminished the inhibitory effect of Sch B on cell
proliferation (Fig. 4B). Moreover,
Fer-1 pre-treatment significantly decreased Sch B-induced
Fe2+ levels (Fig. 4C),
lipid peroxidation (Fig. 4D), and
MDA levels (Fig. 4E), whereas it
significantly increased GSH levels (Fig. 4E). WB revealed that Fer-1
pre-treatment also reversed the Sch B-induced upregulation of ACSL4
and downregulation of GPX4 (Fig.
4F). Collectively, these findings suggested that ferroptosis
may be the predominant mechanism underlying Sch B-induced PC cell
death.
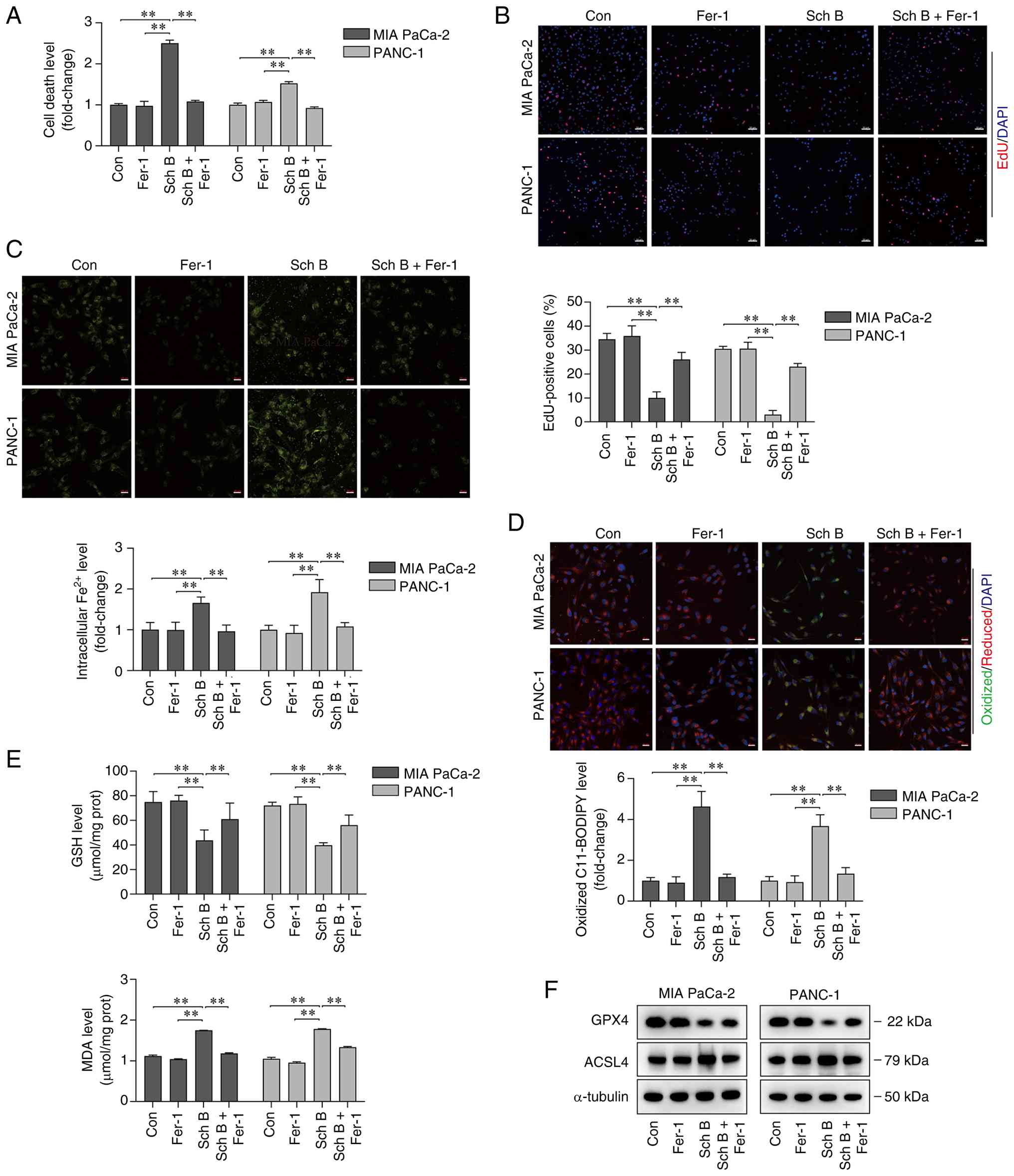 | Figure 4.Fer-1 blocks Sch B-mediated
ferroptosis in pancreatic cancer cells. (A) Cell death of MIA
PaCa-2 and PANC-1 cells treated with Sch B (20 µM) with or without
Fer-1 (10 µM) was measured using lactate dehydrogenase cytotoxicity
assay. (B) Cell proliferation of MIA PaCa-2 and PANC-1 cells
treated with Sch B (20 µM) with or without Fer-1 (10 µM) was
evaluated using EdU staining. Scale bars, 50 µm. (C) Intracellular
Fe2+ levels of MIA PaCa-2 and PANC-1 cells treated with
Sch B (20 µM) with or without Fer-1 (10 µM) were detected using
FerroOrange probe. Scale bars, 20 µm. (D) Lipid peroxidation levels
of MIA PaCa-2 and PANC-1 cells treated with Sch B (20 µM) with or
without Fer-1 (10 µM) were detected using C11-BODIPY 581/591
fluorescent probe. Scale bars, 20 µm. (E) GSH and MDA levels of MIA
PaCa-2 and PANC-1 cells treated with Sch B (20 µM) with or without
Fer-1 (10 µM) were measured using kits. (F) Protein expression
levels of ACSL4 and GPX4 in MIA PaCa-2 and PANC-1 cells treated
with Sch B (20 µM) with or without Fer-1 (10 µM) were detected by
western blotting. **P<0.01. ACSL4, acyl-CoA synthetase
long-chain family member 4; Con, control; Fer-1, ferrostatin-1;
GPX4, glutathione peroxidase 4; GSH, glutathione; MDA,
malondialdehyde; Sch B, Schisandrin B. |
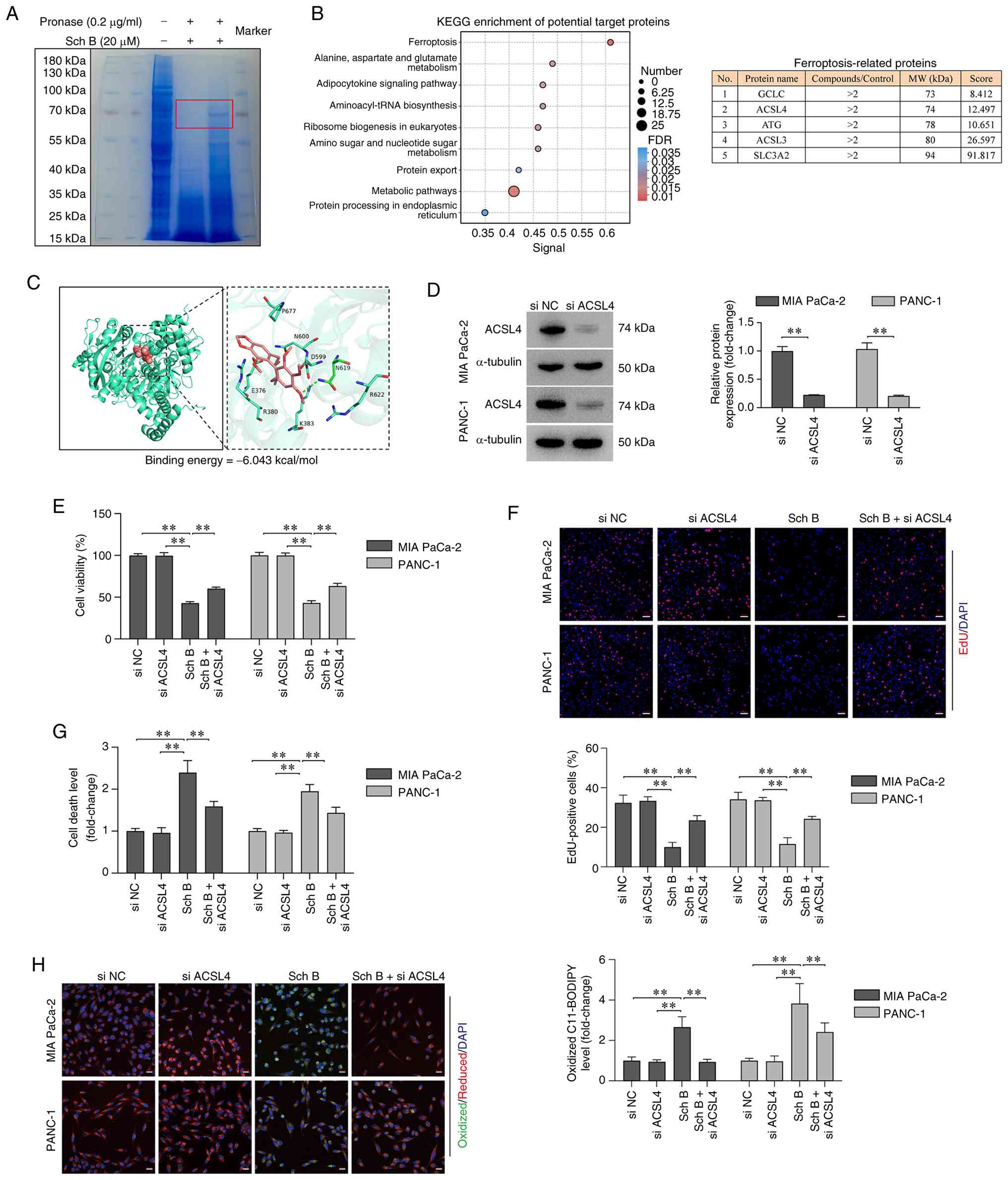
Sch B promotes ACSL4-mediated
ferroptosis in PC cells
To explore the molecular mechanism of Sch B-induced
ferroptosis, DARTS and mass spectrometry analyses conducted in
PANC-1 cells were. The results showed that a protein band with a
molecular weight of ~70 kDa was distinctly protected by Sch B
(Fig. 5A). Mass spectrometry
identified 136 proteins as potential targets with significant
differences (intensity ratio ≥2.0). KEGG pathway enrichment
analysis revealed that the potential target proteins were enriched
in the ‘ferroptosis’ pathway. Based on the combination of intensity
ratio ≥2.0, molecular weight matching the protected band (~70 kDa),
and protein score ≥10.0, ACSL4 was identified as a promising
candidate (Fig. 5B). Molecular
docking was performed between Sch B and the predicted protein
structure of ACSL4; the binding energy was-6.043 kcal/mol,
indicating a potential interaction (Fig. 5C). Based on these results, it was
hypothesized that Sch B may promote ferroptosis by interacting with
ACSL4. To test this hypothesis, ACSL4 was silenced using siRNA in
PANC-1 cells before Sch B treatment (Fig. 5D). ACSL4 knockdown effectively
reduced the sensitivity of PC cells to Sch B, as evidenced by
restored cell viability (Fig. 5E)
and proliferative capacity (Fig.
5F). Additionally, ACSL4 knockdown protected against Sch
B-induced cell death (Fig. 5G) and
reduced lipid peroxidation (Fig.
5H). These findings confirmed that Sch B induces ferroptosis by
activating the ACSL4-regulated ferroptosis pathway in PC cells.
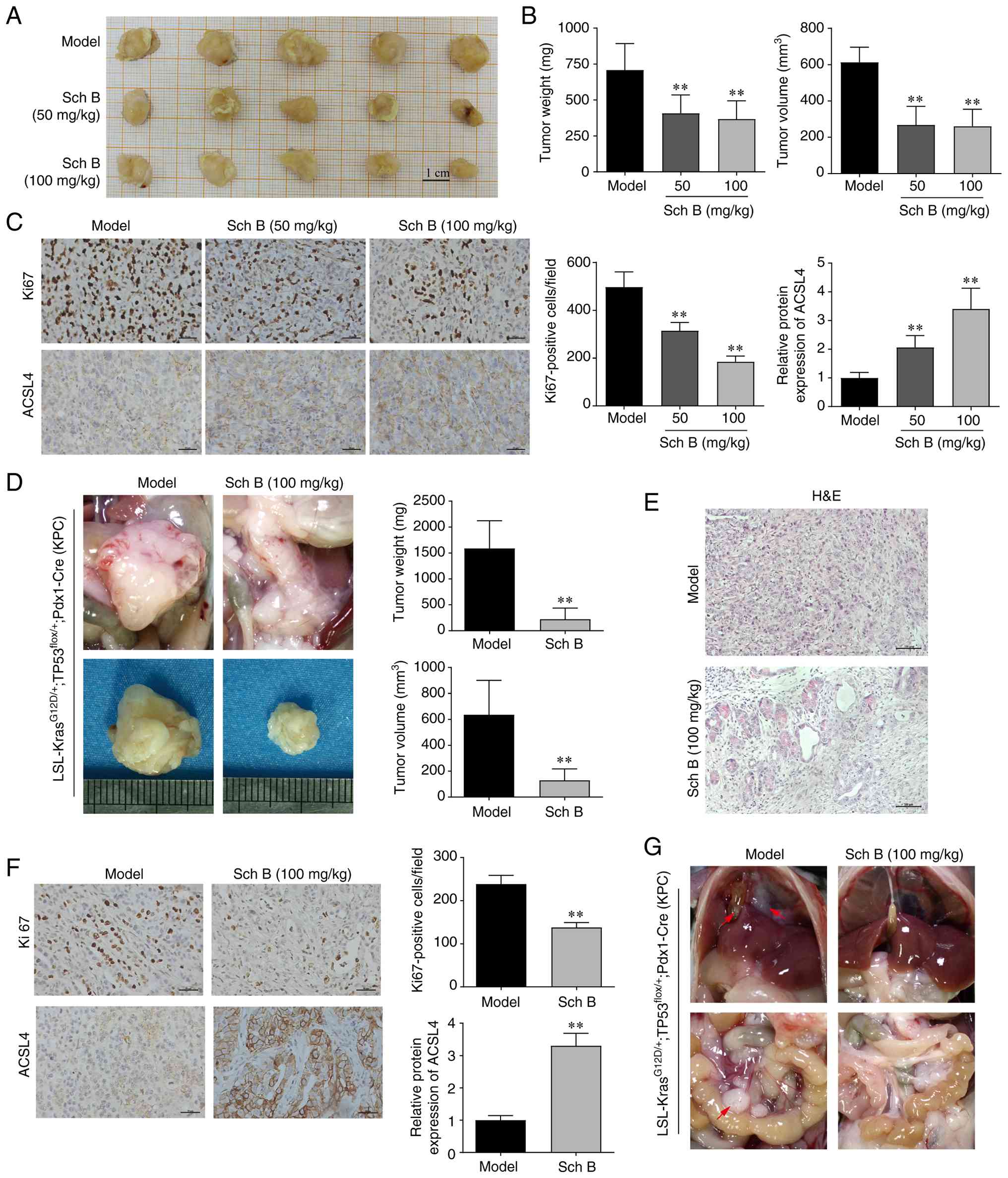
Sch B exerts antitumor efficacy by
inducing ferroptosis in vivo
To assess the therapeutic efficacy of Sch B in
vivo, two murine models were established: A subcutaneous
xenograft tumor model and a genetically engineered orthotopic
spontaneous pancreatic tumor model (KPC). In the xenograft model,
Sch B treatment led to a significant dose-dependent inhibition of
tumor growth; this was evidenced by reduced tumor weight and volume
compared with in the model group (Fig.
6A and B). Immunohistochemical analysis showed a decrease in
Ki67-positive cells and an increase in ACSL4 expression after Sch B
treatment (Fig. 6C). Furthermore,
the role of Sch B in cancer development and metastasis was
investigated using the KPC model. Starting at 10 weeks of age, KPC
mice received daily oral administration of Sch B (100 mg/kg) or
vehicle. Gross pathological examination revealed a substantial
pancreatic tumor burden occupying most of the pancreatic space in
the model group. By contrast, mice treated with Sch B displayed
significantly smaller tumors with preserved pancreatic architecture
(Figs. 6D and S1). Histological analysis indicated that
mice in the model group developed poorly differentiated tumors
characterized by a prominent desmoplastic stroma and complete loss
of normal acinar structure. Conversely, Sch B-treated mice
exhibited moderately differentiated tumors with decreased stromal
density and residual normal pancreatic tissue (Fig. 6E). Immunohistochemical analysis
confirmed that Sch B treatment significantly reduced Ki67-positive
cells and increased ACSL4 expression compared with in the model
group (Fig. 6F). Notably, all model
mice (5/5) developed abdominal metastases (mesenteric: 5/5; liver:
3/5; diaphragmatic: 3/5; bile duct: 2/5; peritoneal: 2/5); by
contrast, only one Sch B-treated mice showed metastatic lesions
(mesenteric: 1/5) (Fig. 6G).
Additionally, toxicological assessments in xenograft mice indicated
that Sch B treatment significantly increased relative liver weight,
but had no effect on body weight or the relative weights of other
organs (Fig. 7A). Histological
examination revealed no observable systemic organ toxicity compared
with in the model group (Fig. 7B).
Serum biochemical analysis revealed no significant differences in
ALT, AST, CK, LDH, CREA and UA levels between the Sch B-treated
group and the model group (Fig.
7C). Overall, Sch B effectively suppressed tumor growth in
vivo by promoting ferroptosis.
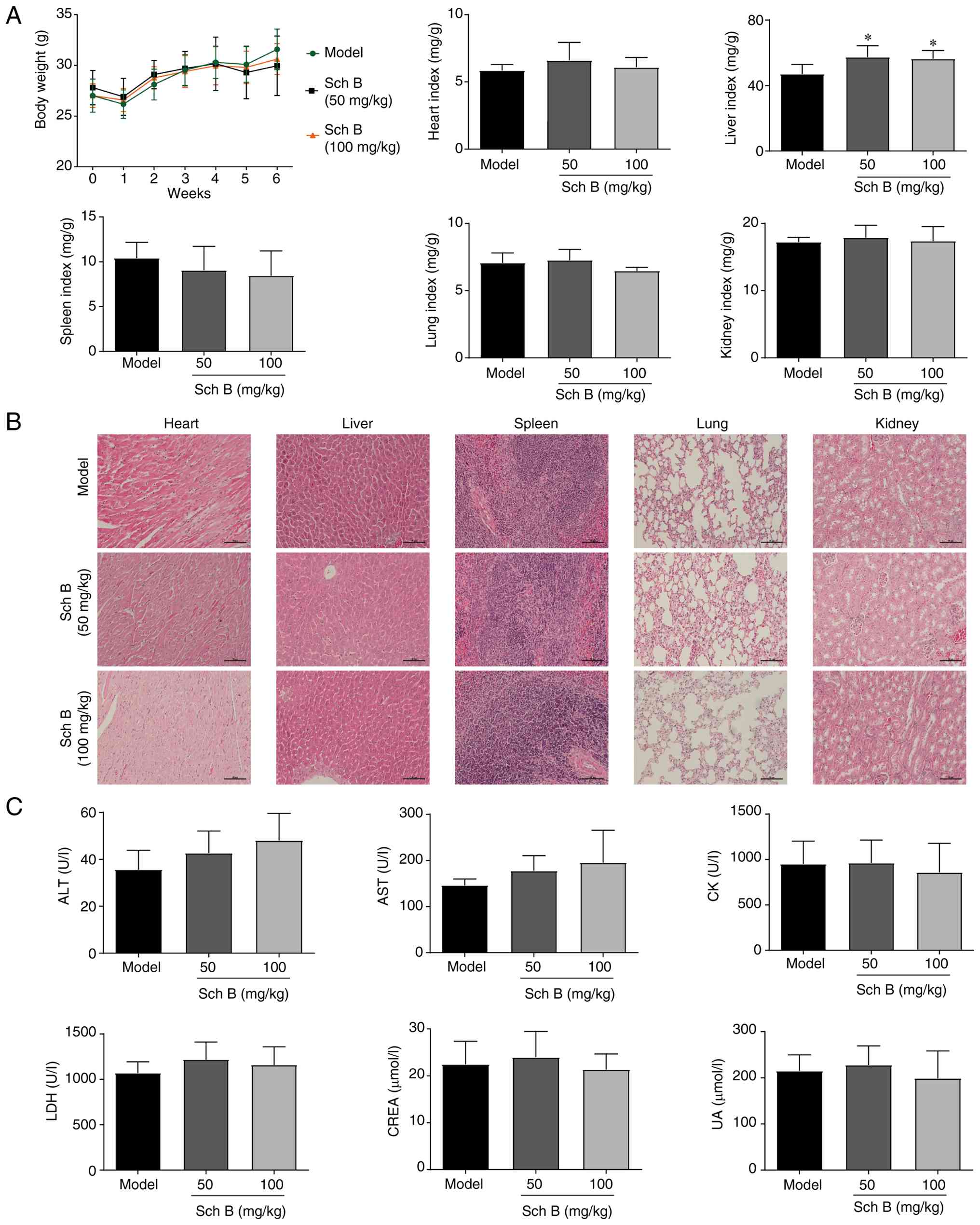 | Figure 7.Evaluation of the potential toxicity
of Sch B in PANC-1 ×enograft mice. (A) Body weight and relative
organ weight of mice in each group. (B) Hematoxylin and eosin
staining of organs in each group. Scale bars, 100 µm. (C) Levels of
ALT, AST, CK, LDH, CREA and UA in serum. *P<0.05 vs. model. ALT,
alanine aminotransferase; AST, aspartate aminotransferase; CK,
creatine kinase; CREA, creatinine; LDH, lactate dehydrogenase; Sch
B, Schisandrin B; UA, uric acid. |
Discussion
PC is one of the most lethal malignant tumors of the
digestive system, with limited treatment options (4). For advanced PC, multiagent
chemotherapy regimens provide only modest survival benefits over
gemcitabine alone (35).
Consequently, more effective treatments are urgently needed.
Natural-product-based herbal medicines represent valuable resources
for drug discovery in PC due to their established efficacy and
safety profiles (36,37). Sch B, the main active compound
derived from Schisandra chinensis (Turcz.) Baill., exhibits
antitumor activity against various types of human cancer. For
example, our recent study revealed that Sch B can inhibit
hepatocellular carcinoma progression by disrupting the crosstalk
between macrophages and hepatoma cells (38). However, to the best of our
knowledge, its therapeutic efficacy in PC has yet to be
investigated. The present findings now confirm that Sch B inhibits
PC cell proliferation and induces cell death, without exhibiting
overt cytotoxic effects on normal pancreatic epithelial cells
(HPNE); therefore, Sch B is a promising potential treatment for
PC.
Ferroptosis was initially characterized as
iron-dependent oxidative cell death occurring in KRAS-mutant tumor
cells (39). Oncogenic KRAS
mutations are present in >90% of patients with PC, and
KRAS-mutant PC cells exhibit heightened susceptibility to
ferroptosis, making it a promising therapeutic strategy (40,41).
Several natural compounds, including Wogonin, Tiliroside and
Solasonine, inhibit PC growth by inducing ferroptosis (42–44).
However, whether Sch B induces ferroptosis in PC remained unclear.
In the current study, transcriptome analysis of Sch B-treated
PANC-1 cells revealed significant enrichment of ferroptosis-related
pathways, such as oxidative phosphorylation, ROS, glutamate and
glutathione metabolism. Pharmacological experiments using
inhibitors indicated that the ferroptosis inhibitor Fer-1 partially
rescued Sch B-induced cell death. Furthermore, transmission
electron microscopy further demonstrated shrunken mitochondria with
increased membrane density and reduced cristae, which are
characteristic features of ferroptosis (45). Additionally, Sch B induced iron
overload, lipid peroxidation, GSH depletion, MDA accumulation and
altered expression of key ferroptosis-related proteins. Notably,
these changes were reversed by Fer-1. Based on these results, it
may be hypothesized that ferroptosis is partially involved in Sch
B-induced cell death in PC.
Lipid peroxidation is the major signal for
ferroptosis and is mediated by both enzymatic and non-enzymatic
Fenton reaction pathways (46).
Polyunsaturated fatty acid (PUFA)-containing phospholipids are the
primary substrates for peroxidation in ferroptosis, which is
regulated by various catalytic enzymes, including ACSL4,
lysophosphatidylcholine acyl-transferase 3 and lipoxygenases. ACSL4
promotes the biosynthesis of lipids with PUFA by catalyzing the
esterification of arachidonic acid and adrenic acid with
phosphatidylethanolamine, which is a vital initiator of ferroptosis
(47). Several studies have
highlighted the pro-ferroptotic role of ACSL4 in PC cells. For
example, dipeptidyl peptidase-4 stabilizes ACSL4 to enhance
ferroptosis sensitivity, and protein tyrosine phosphatase
mitochondrial 1 knockdown upregulates ACSL4 to sensitize PC cells
to erastin-induced ferroptosis (48,49).
Conversely, low ACSL4 expression in PC is associated with poor
prognosis, and ACSL4-mediated suppression of ferroptosis
contributes to resistance against chemotherapy and immunotherapy in
PC cells (50–52). The present study revealed that Sch B
significantly upregulated ACSL4 expression in PC cells. Using DARTS
and molecular docking analysis, ACSL4 was identified as a target
protein for Sch B, and ACSL4 knockdown abolished Sch B-induced
ferroptosis. Complementing these in vitro findings, Sch B
significantly inhibited tumor growth in both xenograft and KPC
models, increased ACSL4 expression and reduced metastasis, with no
overt toxicity. These results suggested that Sch B inhibits PC
through ACSL4-dependent ferroptosis.
The present findings add to the growing body of
evidence supporting the potential of targeting metabolic pathways,
particularly ferroptosis, for treating aggressive gastrointestinal
malignancies, including PC. These data align with the concept that
ferroptosis is a targetable vulnerability in cancer characterized
by high levels of iron and ROS (53). More broadly, PCD, including
pyroptosis, necroptosis, PANoptosis or autophagy, have garnered
increasing attention in neoplastic diseases. These PCD pathways can
either inhibit tumorigenesis or facilitate tumor progression,
immune escape and chemoresistance, depending on the cellular
context (54–59). For example, c-MYC upregulation
drives resistance to anti-EGFR treatment in metastatic colorectal
cancer, exemplifying how oncogenic signaling reprogramming can
undermine targeted treatment efficacy (60). The interplay among these different
modes of cell death is further complicated by microbial factors.
Notably, Fusobacterium nucleatum potentiates gastric
tumorigenesis by altering the tumor microenvironments and enhancing
immune evasion, underscoring the relevance of infectious agents as
modulators of cell death (61).
These observations reveal the complexity of cell death in
gastrointestinal malignancies and suggest that effective
ferroptosis-based therapies must consider this broader network,
which includes potential interactions with microbial and metabolic
factors.
The pharmacokinetic (PK) and pharmacodynamic (PD)
profiles of Sch B are important considerations for its clinical
translation. Although PK/PD parameters were not assessed in the
present study, prior findings have indicated that Sch B is orally
bioavailable and tends to accumulate in tumor tissues. A PK study
in rats reported an absolute oral bioavailability of ~55.0% at a
dose of 10 mg/kg, with linear PK properties observed at 10–40 mg/kg
(62). Oral administration of Sch B
at 150 mg/kg resulted in peak plasma concentrations ranging between
5.9 and 10.2 µM in rats, which aligned well with the effective dose
identified in in vitro assays (63). Additionally, a PD study in mice
revealed that Sch B effectively reached and accumulated at high
levels in tumor tissues (64).
Notably, the orthotopic KPC model presents a substantially greater
drug delivery barrier than xenograft models, owing to the dense
desmoplastic stroma typical of PC. The potent antitumor efficacy
observed in this challenging model, along with existing PK
evidence, strongly indicates that pharmacologically relevant
concentrations of Sch B can be attained in vivo. However,
direct measurement of intra-tumoral concentrations, systemic
half-life and the maximum tolerated dose of Sch B in this model
remains necessary. Future studies should include comprehensive
PK/PD characterization of Sch B in plasma and tumor tissue to
establish a clear dose-exposure-response relationship.
In conclusion, the present study revealed a novel
mechanism by which Sch B inhibits PC at least partially via the
ACSL4-dependent ferroptosis pathway. However, the current study has
certain limitations. The direct binding between ACSL4 and Sch B
requires further verification, and addressing this issue in future
studies will strengthen its potential for clinical application.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the Medical Science and Technology
Foundation of Guangdong Province (grant no. A2024690), the
Scientific Research Project of Guangdong Bureau of Traditional
Chinese Medicine (grant no. 20251478), the Wu JiePing Medical
Foundation (grant no. 320.6750.2024–03-49), the Guangdong Basic and
Applied Basic Research Foundation (grant nos. 2025A1515010072 and
2024A1515220029) and the Research Capability Enhancement Program of
Guangzhou Medical University (grant no. GMUCR2024-02003).
Availability of data and materials
The proteomics data generated in the present study
may be found in the iProX under accession number
IPX0017006000/PXD078099 or at the following URL: https://www.iprox.cn//page/project.html?id=IPX0017006000.
The raw sequencing data generated in the present study may be found
in the Gene Expression Omnibus under accession number GSE315793 or
at the following URL: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE315793.
The other data generated in the present study may be requested from
the corresponding author.
Authors' contributions
FL, LQ and TP designed the study. FL, JF and XL
performed most of the experiments. LY, YLiu and YLi performed some
experiments. FL and JF analyzed and interpreted the data. FL, LQ
and TP wrote and edited the manuscript. All authors read and
approved the final version of the manuscript. FL, JF and LQ confirm
the authenticity of all the raw data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
All animal procedures were ethically reviewed and
approved by the Ethics Committee on Laboratory Animal Care of The
Affiliated Qingyuan Hospital (Qingyuan People's Hospital) of
Guangzhou Medical University (approval nos. LAEC-2021-024 and
LAEC-2023-027).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
PC
|
pancreatic cancer
|
|
Sch B
|
Schisandrin B
|
|
CCK-8
|
Cell Counting Kit-8
|
|
IC50
|
half maximal inhibitory
concentration
|
|
Fer-1
|
ferrostatin-1
|
|
DFO
|
deferoxamine mesylate
|
|
NAC
|
N-acetylcysteine
|
|
NEC-1
|
necrostatin-1
|
|
MDA
|
malondialdehyde
|
|
GSH
|
glutathione
|
|
TF
|
transferrin
|
|
ACSL4
|
acyl-CoA synthetase long-chain family
member 4
|
|
GCLC
|
γ-glutamyl-cysteine ligase catalytic
subunit
|
|
GSS
|
glutathione synthetase
|
|
GPX4
|
glutathione peroxidase 4
|
|
SLC3A2
|
solute carrier family 3 member 2
|
|
SLC7A11
|
solute carrier family 7 member 11
|
|
DEGs
|
differentially expressed genes
|
|
DARTS
|
drug affinity responsive target
stability
|
|
ALT
|
alanine aminotransferase
|
|
AST
|
aspartate aminotransferase
|
|
CK
|
creatine kinase
|
|
LDH
|
lactate dehydrogenase
|
|
CREA
|
creatinine
|
|
UA
|
uric acid
|
References
|
1
|
Siegel RL, Miller KD, Wagle NS and Jemal
A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48.
2023.PubMed/NCBI
|
|
2
|
Xia C, Dong X, Li H, Cao M, Sun D, He S,
Yang F, Yan X, Zhang S, Li N and Chen W: Cancer statistics in China
and United States, 2022: Profiles, trends, and determinants. Chin
Med J (Engl). 135:584–590. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zeng H, Zheng R, Sun K, Zhou M, Wang S, Li
L, Chen R, Han B, Liu M, Zhou J, et al: Cancer survival statistics
in China 2019–2021: A multicenter, population-based study. J Natl
Cancer Cent. 4:203–213. 2024.PubMed/NCBI
|
|
4
|
Park W, Chawla A and O'Reilly EM:
Pancreatic cancer: A review. JAMA. 326:851–862. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Strobel O, Neoptolemos J, Jäger D and
Büchler MW: Optimizing the outcomes of pancreatic cancer surgery.
Nat Rev Clin Oncol. 16:11–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Siegel RL, Miller KD, Fuchs HE and Jemal
A: Cancer statistics, 2021. CA Cancer J Clin. 71:7–33.
2021.PubMed/NCBI
|
|
7
|
Halbrook CJ, Lyssiotis CA, Pasca di
Magliano M and Maitra A: Pancreatic cancer: Advances and
challenges. Cell. 186:1729–1754. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grossberg AJ, Chu LC, Deig CR, Fishman EK,
Hwang WL, Maitra A, Marks DL, Mehta A, Nabavizadeh N, Simeone DM,
et al: Multidisciplinary standards of care and recent progress in
pancreatic ductal adenocarcinoma. CA Cancer J Clin. 70:375–403.
2020.PubMed/NCBI
|
|
9
|
Sharma R, Kumar S, Ghosh R, Komal K and
Kumar M: Gene therapy: Transforming the battle against pancreatic
cancer. Curr Gene Ther. 26:152–159. 2026. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liaki V, Barrambana S, Kostopoulou M,
Lechuga CG, Zamorano-Dominguez E, Acosta D, Morales-Cacho L,
Álvarez R, Sun P, Rosas-Perez B, et al: A targeted combination
therapy achieves effective pancreatic cancer regression and
prevents tumor resistance. Proc Natl Acad Sci USA.
122:e25230391222025. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo X and Gao Z: MARVELD1 promotes the
invasiveness in pancreatic adenocarcinoma through the activation of
epithelial-to-mesenchymal transition. Protein Pept Lett.
32:224–233. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song G, Qi X and Zhao Y: iRGD tumor
penetrating peptide-modified NK cells exhibit enhanced tumor immune
infiltration ability and anti-tumor efficacy. Protein Pept Lett.
32:183–193. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kaviyaprabha R, Miji TV, Sreelakshmi PS,
Muthusami S, Arulselvan P and Bharathi M: Unveiling the potential
role of hesperetin and emodin as a combination therapy to inhibit
the pancreatic cancer progression against the C-met gene. Protein
Pept Lett. 32:280–298. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Cao F, Yin HL, Huang ZJ, Lin ZT, Mao
N, Sun B and Wang G: Ferroptosis: Past, present and future. Cell
Death Dis. 11:882020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng X, Jin X, Ye F, Liu X, Yu B, Li Z,
Zhao T, Chen W, Liu X, Di C and Li Q: Ferroptosis: A novel
regulated cell death participating in cellular stress response,
radiotherapy, and immunotherapy. Exp Hematol Oncol. 12:652023.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Noè R, Inglese N, Romani P, Serafini T,
Paoli C, Calciolari B, Fantuz M, Zamborlin A, Surdo NC, Spada V, et
al: Organic selenium induces ferroptosis in pancreatic cancer
cells. Redox Biol. 68:1029622023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lei G, Zhang Y, Koppula P, Liu X, Zhang J,
Lin SH, Ajani JA, Xiao Q, Liao Z, Wang H and Gan B: The role of
ferroptosis in ionizing radiation-induced cell death and tumor
suppression. Cell Res. 30:146–162. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang C, Dong Q, Bao H, Ge Y, Xu Z, Li J,
Jiang X, Xu Y and Zhong X: Ferroptosis: New strategies and ideas
for the treatment of pancreatic ductal adenocarcinoma. Front Biosci
(Landmark Ed). 29:452024. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang L, Yang L, Du K and Yang Y:
Myoglobin improves doxycycline sensitivity in pancreatic cancer
through promoting heme oxygenase-1-mediated ferroptosis. Environ
Toxicol. 39:2166–2181. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao X and Wang X, Zhang W, Tian T, Zhang
J, Wang J, Wei W, Guo Z, Zhao J and Wang X: A ferroptosis-inducing
arsenene-iridium nanoplatform for synergistic immunotherapy in
pancreatic cancer. Angew Chem Int Ed Engl. 63:e2024008292024.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou Y, Men L, Sun Y, Wei M and Fan X:
Pharmacodynamic effects and molecular mechanisms of lignans from
Schisandra chinensis turcz. (baill.), a current review. Eur
J Pharmacol. 892:1737962021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao Y, Wu S, Cong R, Xiao J and Ma F:
Characterization of lignans in Schisandra chinensis oil with
a single analysis process by UPLC-Q/TOF-MS. Chem Phys Lipids.
218:158–167. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Olas B: Cardioprotective potential of
berries of Schisandra chinensis turcz. (baill.), their
components and food products. Nutrients. 15:5922023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yan LS, Zhang SF, Luo G, Cheng BCY, Zhang
C, Wang YW, Qiu XY, Zhou XH, Wang QG, Song XL, et al: Schisandrin B
mitigates hepatic steatosis and promotes fatty acid oxidation by
inducing autophagy through AMPK/mTOR signaling pathway. Metabolism.
131:1552002022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee TH, Jung CH and Lee DH:
Neuroprotective effects of schisandrin B against transient focal
cerebral ischemia in sprague-dawley rats. Food Chem Toxicol.
50:4239–4245. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li J, Lu Y, Wang D, Quan F, Chen X, Sun R,
Zhao S, Yang Z, Tao W, Ding D, et al: Schisandrin B prevents
ulcerative colitis and colitis-associated-cancer by activating
focal adhesion kinase and influence on gut microbiota in an in vivo
and in vitro model. Eur J Pharmacol. 854:9–21. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nasser MI, Zhu S, Chen C, Zhao M, Huang H
and Zhu P: A comprehensive review on schisandrin B and its
biological properties. Oxid Med Cell Longev. 2020:21727402020.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Song A, Ding T, Wei N, Yang J, Ma M, Zheng
S and Jin H: Schisandrin B induces HepG2 cells pyroptosis by
activating NK cells mediated anti-tumor immunity. Toxicol Appl
Pharmacol. 472:1165742023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tan S, Zheng Z, Liu T, Yao X, Yu M and Ji
Y: Schisandrin B induced ROS-mediated autophagy and Th1/TH2
imbalance via selenoproteins in Hepa1-6 cells. Front Immunol.
13:8570692022. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sana-Eldine AO, Abdelgawad HM, Kotb NS and
Shehata NI: The potential effect of schisandrin-B combination with
panitumumab in wild-type and mutant colorectal cancer cell lines:
Role of apoptosis and autophagy. J Biochem Mol Toxicol.
37:e233242023. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He L, Chen H, Qi Q, Wu N, Wang Y, Chen M,
Feng Q, Dong B, Jin R and Jiang L: Schisandrin B suppresses gastric
cancer cell growth and enhances the efficacy of chemotherapy drug
5-FU in vitro and in vivo. Eur J Pharmacol. 920:1748232022.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yan C, Gao L, Qiu X and Deng C:
Schisandrin B synergizes docetaxel-induced restriction of growth
and invasion of cervical cancer cells in vitro and in vivo. Ann
Transl Med. 8:11572020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan YH, Kong L, Lu YB, Li SY, Yan AW, Song
YW, Huang ZH and Zhu HN: Inhibition of hepatocellular carcinoma
progression by methotrexate-modified pH-sensitive sorafenib and
schisandrin B micelles. Biomed Mater. 20:0150222025. View Article : Google Scholar
|
|
34
|
Morris GM, Huey R, Lindstrom W, Sanner MF,
Belew RK, Goodsell DS and Olson AJ: AutoDock4 and AutoDockTools4:
Automated docking with selective receptor flexibility. J Comput
Chem. 30:2785–2791. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Conroy T, Desseigne F, Ychou M, Bouché O,
Guimbaud R, Bécouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de
la Fouchardière C, et al: FOLFIRINOX versus gemcitabine for
metastatic pancreatic cancer. N Engl J Med. 364:1817–1825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim A, Ha J, Kim J, Cho Y, Ahn J, Cheon C,
Kim SH, Ko SG and Kim B: Natural products for pancreatic cancer
treatment: From traditional medicine to modern drug discovery.
Nutrients. 13:38012021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sahu SK, Prabhakar PK and Vyas M:
Therapeutical potential of natural products in treatment of
pancreatic cancer: A review. Mol Biol Rep. 52:1792025. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang B, Yang J, Huang Q, Li W, Peng Q,
Gan H, Peng T, Yao L and Qi L: Schisandrin B downregulates exosomal
fibronectin 1 expression to inhibit hepatocellular carcinoma
growth. Front Pharmacol. 16:15476852025. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu X, Yang J, Huang S, Hong Y, Zhu Y,
Wang J, Wang Y, Liang T and Bai X: Pancreatic cancer-derived
extracellular vesicles enhance chemoresistance by delivering
KRASG12D protein to cancer-associated fibroblasts. Mol Ther.
33:1134–1153. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhao H, Huang Q, Liu YA and Wu W:
Oncogenic KRAS promotes ferroptosis in pancreatic cancer through
regulation of the Fosl1-tfrc axis. Pancreas. 54:e235–e245. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu X, Peng X, Cen S, Yang C, Ma Z and Shi
X: Wogonin induces ferroptosis in pancreatic cancer cells by
inhibiting the Nrf2/GPX4 axis. Front Pharmacol. 14:11296622023.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu M, Zhong W, Yang C, Liu M, Yuan X, Lu
T, Li D, Zhang G, Liu H, Zeng Y, et al: Tiliroside disrupted iron
homeostasis and induced ferroptosis via directly targeting
calpain-2 in pancreatic cancer cells. Phytomedicine.
127:1553922024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liang X, Hu C, Han M, Liu C, Sun X, Yu K,
Gu H and Zhang J: Solasonine inhibits pancreatic cancer progression
with involvement of ferroptosis induction. Front Oncol.
12:8347292022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li J, Jia YC, Ding YX, Bai J, Cao F and Li
F: The crosstalk between ferroptosis and mitochondrial dynamic
regulatory networks. Int J Biol Sci. 19:2756–2771. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen X, Kang R, Kroemer G and Tang D:
Broadening horizons: The role of ferroptosis in cancer. Nat Rev
Clin Oncol. 18:280–296. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen F, Kang R, Liu J and Tang D: The
ACSL4 network regulates cell death and autophagy in diseases.
Biology (Basel). 12:8642023.PubMed/NCBI
|
|
48
|
Huang XD, Xiao FJ, Guo YT, Sun Y, Zhang YK
and Shi XJ: Protein tyrosine phosphatase 1 protects human
pancreatic cancer from erastin-induced ferroptosis. Asian J Surg.
45:2214–2223. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhou X, Kong L, Zhang B, Xie S, Wang W and
Chen G: DPP4 suppresses pancreatic cancer growth by enhancing
ferroptosis sensitivity through stabilization of ACSL4. Cell
Signal. 143:1124462026. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Uchihara D, Shimajiri S, Harada Y,
Kumamoto K, Oe S, Miyagawa K, Nakamura K, Katafuchi E, Nuratdinova
F, Honma Y, et al: Long-chain fatty acyl CoA synthetase 4
expression in pancreatic cancer: A marker for malignant lesions and
prognostic indicator for recurrence. Diagn Pathol. 20:592025.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Qi R, Bai Y, Li K, Liu N, Xu Y, Dal E,
Wang Y, Lin R, Wang H, Liu Z, et al: Cancer-associated fibroblasts
suppress ferroptosis and induce gemcitabine resistance in
pancreatic cancer cells by secreting exosome-derived
ACSL4-targeting miRNAs. Drug Resist Updat. 68:1009602023.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
He Z, Zheng D, Li F, Chen L, Wu C, Zeng Z
and Yu C: TMOD3 accelerated resistance to immunotherapy in
KRAS-mutated pancreatic cancer through promoting
autophagy-dependent degradation of ASCL4. Drug Resist Updat.
78:1011712025. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lei G, Zhuang L and Gan B: Targeting
ferroptosis as a vulnerability in cancer. Nat Rev Cancer.
22:381–396. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang Y and Kanneganti TD: From pyroptosis,
apoptosis and necroptosis to PANoptosis: A mechanistic compendium
of programmed cell death pathways. Comput Struct Biotechnol J.
19:4641–4657. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang S, He H, Qu L, Shen Q and Dai Y: Dual
roles of inflammatory programmed cell death in cancer: Insights
into pyroptosis and necroptosis. Front Pharmacol. 15:14464862024.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Strippoli A, Cocomazzi A, Basso M, Cenci
T, Ricci R, Pierconti F, Cassano A, Fiorentino V, Barone C, Bria E,
et al: c-MYC expression is a possible keystone in the colorectal
cancer resistance to EGFR inhibitors. Cancers (Basel). 12:6382020.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yun CW and Lee SH: The roles of autophagy
in cancer. Int J Mol Sci. 19:34662018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Santana-Codina N, Mancias JD and Kimmelman
AC: The role of autophagy in cancer. Annu Rev Cancer Biol. 1:19–39.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Pizzimenti C, Fiorentino V, Ruggeri C,
Franchina M, Ercoli A, Tuccari G and Ieni A: Autophagy involvement
in non-neoplastic and neoplastic endometrial pathology: The state
of the art with a focus on carcinoma. Int J Mol Sci. 25:121182024.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Pizzimenti C, Fiorentino V, Franchina M,
Martini M, Giuffrè G, Lentini M, Silvestris N, Di Pietro M, Fadda G
and Tuccari G,and Ieni A: Autophagic-related proteins in brain
gliomas: Role, mechanisms, and targeting agents. Cancers (Basel).
15:26222023. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sorino J, Della Mura M, Ingravallo G,
Cazzato G, Pizzimenti C, Zuccalà V, Pepe L, Germanà E, Martini M,
Ieni A and Fiorentino V: Fusobacterium nucleatum and gastric
cancer: An emerging connection. Int J Mol Sci. 26:79152025.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang Z, You L, Cheng Y, Hu K, Wang Z,
Cheng Y, Yang J, Yang Y and Wang G: Investigation of
pharmacokinetics, tissue distribution and excretion of schisandrin
B in rats by HPLC-MS/MS. Biomed Chromatogr. 32:e40692018.
View Article : Google Scholar
|
|
63
|
Li S, Shao Q and Qiao H: Preclinical
concomitant toxicokinetic study of schisandrin B by HPLC-MS/MS.
Biomed Chromatogr. 39:e700682025. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lee PK, Co VA, Yang Y, Wan MLY, El-Nezami
H and Zhao D: Bioavailability and interactions of schisandrin B
with 5-fluorouracil in a xenograft mouse model of colorectal
cancer. Food Chem. 463:1413712025. View Article : Google Scholar : PubMed/NCBI
|