Introduction
Breast cancer remains the most commonly diagnosed
cancer and the leading cause of cancer-related mortality among
women worldwide. According to GLOBOCAN 2022 estimates, ~2.3 million
new female breast cancer cases and 666,000-670,000 deaths occurred
globally in 2022, accounting for ~11.6% of all new cancer cases and
6.9% of all cancer deaths (1).
Despite substantial advances in surgery, radiotherapy,
chemotherapy, endocrine therapy and targeted agents, therapeutic
resistance, recurrence and treatment-related toxicities continue to
limit long-term clinical outcomes (2–4). These
challenges underscore the need for mechanistically rational and
clinically translatable therapeutic strategies that improve
efficacy without adding unacceptable systemic toxicity (5,6).
Natural products have historically contributed to
anticancer drug discovery; however, their role in contemporary drug
development has become more limited due to challenges such as
structural complexity, low drug-likeness and difficulties in
optimization and formulation (7).
Nevertheless, bioactive phytochemicals remain valuable as
mechanistic probes or scaffolds for structural modification,
particularly when they exhibit unique biochemical properties not
easily replicated by synthetic molecules. Therefore, natural
products should be evaluated not only for biological activity but
also for pharmacokinetic (PK) feasibility, target selectivity and
safety margins.
Salvia miltiorrhiza (Danshen), a widely used
traditional medicinal herb, contains a series of lipophilic
diterpenoid quinones known as tanshinones, among which tanshinone
IIA (Tan IIA) is the major and most extensively studied constituent
(8–11). Beyond its well-recognized
cardiovascular and anti-inflammatory activities (12–15),
Tan IIA has attracted attention in oncology due to its ability to
interfere with cancer-associated vulnerabilities, including
mitochondrial dysfunction, epigenetic regulation and oncogenic
signaling networks (16,17). However, native Tan IIA has poor
aqueous solubility, extensive first-pass metabolism, low oral
bioavailability of reportedly ~2–6% in rats and a short plasma
half-life after intravenous administration (18). These limitations raise a critical
question as to whether the concentrations required for many in
vitro anticancer mechanisms can be achieved and sustained in
vivo.
Studies have expanded the mechanistic understanding
of Tan IIA in breast cancer, demonstrating effects on apoptosis,
proliferation, angiogenesis, redox homeostasis and hormone- and
growth factor-driven pathways (19,20).
At the same time, substantial efforts have been devoted to
improving its PK profile through structural modification and
advanced delivery technologies, including acetyltanshinone
derivatives (21), nanoscale
formulations (22) and other
rationally designed analogues (23). Nevertheless, most of these findings
remain preclinical and are derived from heterogeneous cell-line
systems, exposure durations and formulations, which limits direct
comparison across mechanisms.
The present review focuses specifically on Tan IIA
research in breast cancer and explicitly links mechanistic insights
to translational constraints. Recent studies are incorporated and
the mechanistic evidence is systematically organized according to
key cancer vulnerabilities and clinically relevant pathway
contexts, including hormone- and growth factor-driven signaling.
Particular emphasis is placed on integrating structure-activity
relationships with mechanistic readouts to clarify how chemical
modifications reshape antitumor efficacy. In parallel, formulation
and delivery strategies are critically assessed in light of current
progress and the practical barriers to clinical development,
including PK limitations and translational feasibility. By aligning
mechanistic claims with druggability realities, this review
delineates what is supported by existing in vitro and in
vivo evidence, what remains uncertain, and what will be
required to advance Tan IIA-derived strategies toward credible and
clinically meaningful applications in breast cancer therapy.
Chemical and pharmacological basis of Tan
IIA
Chemically, Tan IIA is a highly lipophilic diterpene
quinone derived from Salvia miltiorrhiza. It is essentially
insoluble in water and dissolves only in organic solvents (11,24).
These properties contribute to its low systemic availability (oral
bioavailability, ~2–6% in rats). Tan IIA also has a short plasma
half-life (~0.3 h in rats after i.v. administration) (18). Together, its high first-pass
metabolism and poor absorption mean that native Tan IIA requires
special formulation to achieve therapeutic levels. Consequently,
extensive efforts have focused on developing delivery systems
(e.g., nanoparticles, liposomes) and derivatives to overcome these
PK challenges. These PK features have direct implications for
interpreting mechanistic studies. Concentrations that are readily
achieved in vitro may not reflect tumor exposures attainable
in vivo, particularly for native Tan IIA without formulation
support. Therefore, mechanisms reported at high micromolar
concentrations should not be considered equally translatable unless
accompanied by PK/pharmacodynamic (PD) evidence, tumor accumulation
data or in vivo target-engagement validation.
Beyond cancer, Tan IIA has cardiovascular,
anti-inflammatory and metabolic activities (24–33).
These pleiotropic effects may be beneficial in some contexts but
may also complicate anticancer development, because systemic
exposure could affect non-tumor tissues and interact with
cardiovascular or inflammatory homeostasis (34,35).
Therefore, the present review focuses on breast cancer-related
evidence while also considering PK feasibility and potential safety
liabilities.
Mechanisms of Tan IIA in breast cancer
In breast cancer models, Tan IIA and its derivatives
have been associated with several mechanistic effects, including
estrogen receptor α (ERα) modulation, chemosensitization,
apoptosis, breast cancer stem cell (BCSC) inhibition, ferroptosis
and G-quadruplex (G4) stabilization (Table SI). However, these effects should
not be interpreted as parallel mechanisms occurring with equal
relevance. For example, ERα-related effects have been reported at
low micromolar concentrations, whereas ferroptosis and
chemosensitization studies often employed different concentration
ranges and experimental contexts (Table SI), highlighting the heterogeneity
of the available evidence.
The mechanisms summarized below differ in
pharmacological plausibility. ERα degradation has been mainly
characterized in ER-positive models treated with acetyltanshinone
IIA (ATA) or its active metabolite hydroquinone Tan IIA (HTA) at
low micromolar concentrations. BCSC inhibition and
chemosensitization have been reported in defined resistant or
stem-like cell populations over relatively broad but model-specific
concentration ranges. By contrast, reactive oxygen species
(ROS)-dependent apoptosis, ferroptosis and G4 stabilization are
often observed at higher concentrations or with structurally
optimized derivatives. Accordingly, these mechanisms should be
viewed as context-dependent rather than interchangeable. In
particular, high-micromolar in vitro effects require
cautious interpretation in light of the poor bioavailability and
short systemic half-life of native Tan IIA.
Modulation of ER signaling in
ER-positive and ER-context-dependent models
ER signaling plays a pivotal role in the
proliferation and survival of ER+ breast cancers (36). In specific ER+ cell-line models, ATA
showed stronger antiproliferative activity than tamoxifen under the
tested experimental conditions (21).
In vitro studies demonstrate that ATA
inhibits ER+ breast cancer cell growth in a dose-dependent manner
over a concentration range of 1.56–25 µM, with selective
suppression of ER+ cells observable at ~3 µM. The half-maximal
inhibitory concentration (IC50) of ATA against ER+ breast cancer
cell lines is ~1.4–1.5 µM, markedly lower than that of tamoxifen,
while 6 µM has been established as a standard effective
concentration for downstream mechanistic investigations.
Mechanistically, the active ATA metabolite HTA directly binds to
ERα and promotes its nuclear degradation via a
ubiquitin-proteasome-dependent pathway, while simultaneously
downregulating the transcription of estrogen receptor 1 (ESR1), the
gene encoding ERα. This dual mechanism distinguishes ATA from
classical selective ER degraders such as fulvestrant. In addition,
ATA suppresses the transcription of ER-responsive genes, including
growth regulating estrogen receptor binding 1, thereby effectively
inhibiting ER-driven proliferation in breast cancer cells (21). Consistent with these findings,
combination treatment with Tan IIA and fulvestrant produces
synergistic antitumor effects in ER+ breast cancer models. In ER+
ZR-75-1 ×enograft tumors, combined therapy significantly inhibited
tumor growth and elicited earlier therapeutic responses compared
with either agent alone. Importantly,
18F-fluoroestradiol positron emission tomography/CT
imaging enabled real-time monitoring of treatment response, with
quantitative imaging signals closely correlating with intratumoral
ERα expression, further validating the synergistic interaction
between Tan IIA and fulvestrant in vivo (37). Thus, ERα degradation represents one
of the more pharmacologically plausible and subtype-defined
mechanisms, but it is primarily supported by ATA/HTA data in
ER+ models rather than by evidence that native Tan IIA
produces identical ERα target coverage in vivo.
Tan IIA has also shown antiproliferative effects in
ER− models, suggesting that ER-independent mechanisms
may contribute under certain experimental conditions. Both in
vitro and in vivo studies demonstrate that Tan IIA
exhibits greater antitumor efficacy than tamoxifen in
ER− models, potentially through downregulation of p53
and Bcl-2 expression and suppression of genes involved in cell
cycle progression, proliferation, apoptosis and DNA synthesis
(38). Tan IIA displays an IC50
value of ~0.25 µg/ml in both ER+ and ER− breast cancer
cell lines, reflecting potent and relatively non-subtype-restricted
antiproliferative activity. In vitro, Tan IIA inhibits cell
growth in a dose-dependent manner over a concentration range of
0.0625–1.0 µg/ml, with 0.25 µg/ml commonly used for mechanistic
studies. In vivo, subcutaneous administration of Tan IIA at
30 mg/kg significantly suppresses tumor growth and induces
apoptosis, supporting its antitumor efficacy in animal models
(38). Advances in nanomedicine
have further expanded the therapeutic potential of Tan IIA by
addressing its PK limitations and enhancing tumor targeting. Given
that ER is overexpressed in various tumor cells, it has been
explored as a target receptor for nanoparticle-based drug delivery.
Molecular docking studies suggest that Tan IIA, acting as a
phytoestrogen, possesses high ER-binding affinity and can be
effectively incorporated into engineered nanocarriers. A modified
derivative, Tan IIA-NH2, demonstrates favorable biocompatibility,
enhanced tumor-targeting capability, robust antitumor efficacy and
anti-metastatic potential. Furthermore, mesoporous silica
nanoparticles co-loaded with Tan IIA and doxorubicin (Dox)
(Tan-Dox-MSN) exhibit uniform particle size distribution, good
dispersion and high drug-loading capacity. Both in vitro and
in vivo experiments show that Tan-Dox-MSN achieves superior
tumor accumulation and antitumor effects while significantly
reducing systemic toxicity to normal tissues. These findings
highlight the potential of leveraging high-ER-affinity
phytoestrogens such as Tan IIA to improve nanodelivery systems for
breast cancer therapy (22).
However, ER-mediated targeting strategies should be evaluated
carefully because ER expression is not restricted to tumor tissues,
and systemic ER modulation may affect endocrine and inflammatory
homeostasis in normal tissues (Fig.
1).
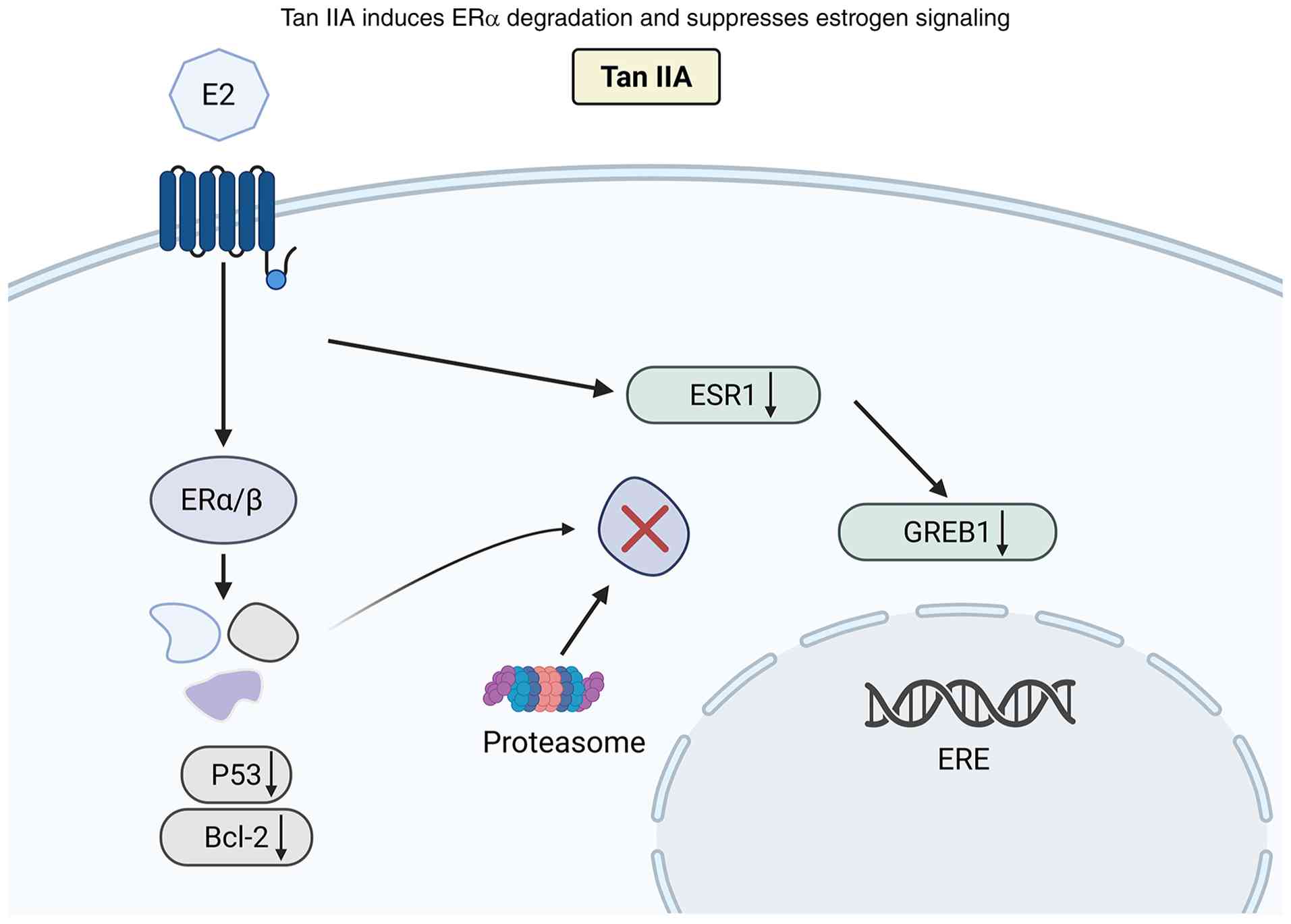 | Figure 1.Tan IIA promotes ubiquitin-mediated
proteasomal degradation of ERα and suppresses ESR1 transcription,
thereby inhibiting the growth of ER+ breast cancer
cells. Tan IIA, Tanshinone IIA; ERα/β, estrogen receptor α/β; ESR1,
estrogen receptor 1; GREB1, growth regulating estrogen receptor
binding 1; E2, estradiol; P53, protein 53; Bcl-2, B-cell lymphoma
2; ERE, estrogen response element; ↑, increase; ↓, decrease. |
Chemosensitization and reversal of
drug resistance
Dox and paclitaxel (taxol) remain frontline
chemotherapeutic agents for breast cancer; however, their long-term
use often results in drug resistance and severe dose-dependent
toxicities, including cardiotoxicity, myelosuppression and weight
loss (39,40). Because BCSCs contribute to drug
resistance and relapse (41), this
section focuses on chemosensitization in resistant models, whereas
BCSC-specific mechanisms are discussed separately below.
Accumulating evidence demonstrates that Tan IIA markedly enhances
the sensitivity of breast cancer cells, including both parental
MCF-7 cells and drug-resistant sublines such as MCF-7/Dox, to Dox
and taxol (42–47). These chemosensitizing effects
involve multiple complementary mechanisms, including inhibition of
drug efflux transporters, modulation of oncogenic signaling
pathways and enhancement of the overall efficacy of combination
chemotherapy.
Mechanistically, Tan IIA downregulates key
ATP-binding cassette transporters, including P-glycoprotein, breast
cancer resistance protein and multidrug resistance-associated
protein 1, thereby increasing intracellular accumulation of Dox and
effectively eliminating both drug-sensitive and drug-resistant
breast cancer cells (42,46). In parallel, Tan IIA suppresses
activation of the PTEN/AKT pathway and inhibits β-catenin nuclear
translocation, further restoring chemosensitivity in resistant
cells (42,43). Consistent with these observations,
Li et al (42) reported that
Tan IIA, at concentrations described as non-toxic in the tested
cell models (≤20 µg/ml), significantly potentiated the cytotoxicity
of Dox, particularly in resistant breast cancer cells, by enhancing
intracellular drug accumulation. However, these concentrations
remain relatively high from a translational PK perspective and
therefore require in vivo exposure confirmation. Combination
treatment markedly reduced the IC50 of Dox, resulting in an
~3.9–4.4-fold increase in chemosensitivity compared with Dox alone
(42).
Tan IIA also enhances the efficacy of taxol in
resistant breast cancer models. In taxol-resistant MCF-7 cells, Tan
IIA suppresses the expression of the microtubule-associated protein
Tau, a known mediator of taxol resistance, thereby potentiating
taxol-induced cytotoxicity (44).
Notably, Tan IIA itself exhibits stronger antiproliferative
activity than taxol in this model, with IC50 values of 8.4 µM for
Tan IIA vs. 23.7 µM for taxol. Combination treatment within
concentration ranges of 1–20 µM (Tan IIA) and 5–100 µM (taxol)
predominantly produced additive effects, indicating that Tau
suppression may contribute to improved taxol responsiveness in this
resistant model. These data support an additive interaction in this
model rather than a universally synergistic effect (44).
Beyond tumor-specific chemosensitization, Tan IIA
exerts dual regulatory effects on ERK1/2 signaling that contribute
to both efficacy enhancement and toxicity mitigation. In breast
cancer cells, Tan IIA suppresses ERK1/2 activation to promote
apoptosis, whereas in cardiomyocytes, it activates ERK1/2
signaling, thereby alleviating Dox-induced cardiotoxicity (45). Another study further revealed that
Tan IIA can reverse hypoxia-induced Dox resistance and suppress
epithelial-mesenchymal transition (EMT). Under hypoxic conditions,
breast cancer cells such as MCF-7 and HCC1937 exhibit reduced
E-cadherin expression and elevated vimentin levels, consistent with
EMT and acquired chemoresistance. Treatment with Tan IIA restores
these markers toward basal levels, significantly reduces cell
viability and proliferation, and likely mediates these effects
through downregulation of hypoxia-inducible factor 1α and TWIST
family basic helix-loop-helix transcription factor-dependent
signaling pathways. Notably, under hypoxic conditions, Tan IIA at
10 µM effectively reverses resistance to Dox (0.2 µg/ml), restoring
drug sensitivity in breast cancer cells (47). The opposite regulation of ERK1/2 in
tumor cells and cardiomyocytes also illustrates the
context-dependent pharmacology of Tan IIA and supports the need to
evaluate tissue-specific responses rather than assuming uniform
pathway inhibition.
Collectively, available preclinical evidence
suggests that Tan IIA can enhance the activity of Dox and taxol in
selected breast cancer models, particularly resistant cell
populations (42–47). However, the magnitude and nature of
the interaction vary across models, ranging from additive to
synergistic effects, and require confirmation at pharmacologically
achievable exposures (Fig. 2).
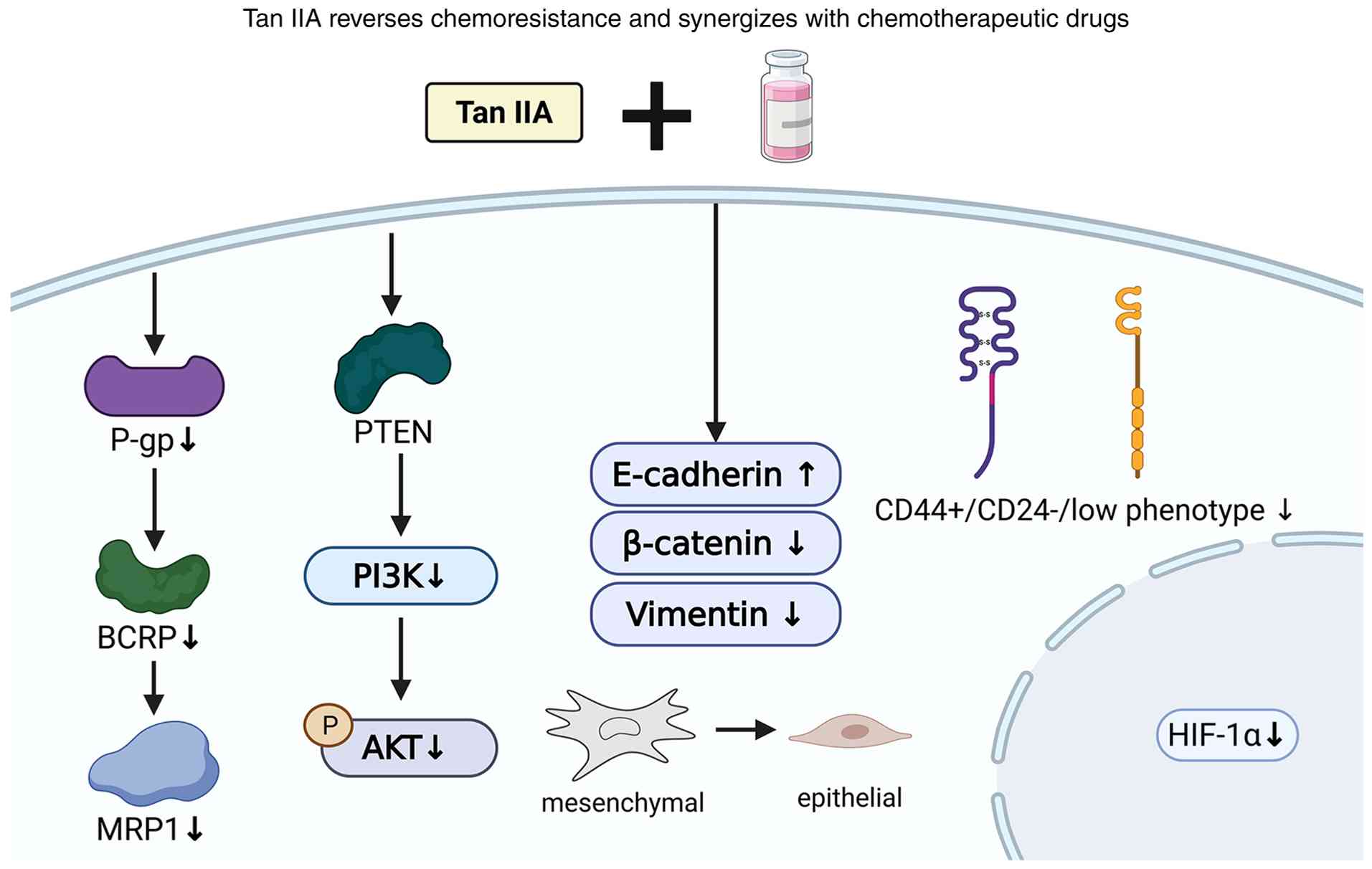 | Figure 2.Tan IIA may attenuate multidrug
resistance by inhibiting drug efflux pumps, suppressing survival
signaling, reversing EMT and targeting BCSCs, thereby improving
chemotherapy responses in selected models. Tan IIA, tanshinone IIA;
AKT, protein kinase B; BCRP, breast cancer resistance protein;
E-cadherin, epithelial cadherin; HIF-1α, hypoxia-inducible factor
1α; MRP1, multidrug resistance-associated protein 1; P-gp,
P-glycoprotein; PI3K, phosphoinositide 3-kinase; PTEN, phosphatase
and tensin homolog; ↑, increase; ↓, decrease. |
Induction of apoptosis and cell cycle
arrest
Apoptosis and cell-cycle arrest are best interpreted
as convergent downstream phenotypes rather than a single uniform
mechanism, because the effective concentrations and upstream
triggers vary considerably among derivatives and cell lines.
Aberrant proliferation and dysregulated apoptosis in breast cancer
cells are major contributors to tumor progression and therapeutic
resistance (48,49). Tan IIA and its derivatives exert
antitumor effects predominantly through the induction of apoptosis
and cell cycle arrest across multiple breast cancer subtypes. Among
these compounds, 1-hydroxy-Tan IIA has been identified as one of
the most active derivatives. These relatively high concentrations
suggest that this derivative may be useful as a mechanistic probe,
but its direct pharmacological relevance requires further
optimization. In MCF-7 and MDA-MB-231 breast cancer cells,
1-hydroxy-Tan IIA promotes DNA fragmentation, upregulates the
pro-apoptotic protein Bax, downregulates the anti-apoptotic protein
Bcl-2 and induces poly(ADP-ribose) polymerase cleavage, thereby
triggering apoptosis (19).
Consistent with these mechanistic findings, tanshinone derivatives
isolated from Stachys parviflora exhibit measurable in
vitro antitumor activity, with 1-hydroxy-Tan IIA showing the
strongest cytotoxicity, characterized by an IC50 of ~22 µg/ml over
a concentration range of 2.5–100 µg/ml (19). In addition to mitochondrial
apoptosis, Tan IIA can induce cell death through ER stress-mediated
pathways. In human breast cancer BT-20 cells, Tan IIA inhibits cell
proliferation in both dose- and time-dependent manners,
significantly increases the proportion of sub-G1 cells and
activates ER stress-related apoptotic signaling, including
caspase-12, growth arrest and DNA damage-inducible protein
153/C/EBP homologous protein, phosphorylated c-Jun N-terminal
kinase (p-JNK) and phosphorylated p38 MAPK. Concurrently, Tan IIA
suppresses the expression of anti-apoptotic Bcl-xL and
phosphorylated ERK, collectively indicating the involvement of ER
stress and MAPK pathway modulation in apoptosis induction (50). Quantitatively, Tan IIA exhibits
time-dependent cytotoxicity in BT-20 cells, with IC50 values of 3.3
µg/ml at 24 h, 1.87 µg/ml at 48 h and 0.67 µg/ml at 72 h, within an
effective concentration range of 0.25–8 µg/ml (50). In TNBC MDA-MB-231 cells, Tan IIA
similarly suppresses proliferation in a dose- and time-dependent
manner, markedly increases the sub-G1 population and activates
apoptotic signaling by upregulating Bax and downregulating Bcl-2
(20).
Additionally, Tan IIA inhibits breast cancer cell
proliferation and migration through modulation of the G
protein-coupled estrogen receptor (GPER)/EGFR/ERK/c-Fos/c-Jun
pathway. Flow cytometry and Transwell assays demonstrate that Tan
IIA induces apoptosis and suppresses migration in MDA-MB-231 cells,
accompanied by reduced expression of c-Fos, c-Jun and cell
cycle-associated proteins. These effects are partially reversed by
GPER inhibitors, implicating GPER as a key mediator. In
vivo, oral administration of Tan IIA significantly reduced
xenograft tumor volume and weight, with concomitant NF-κB
downregulation and caspase-3 upregulation in tumor tissues
(51). In MDA-MB-231 ×enograft
models, long-term oral treatment with Tan IIA (20 or 60 mg/kg, 90
days) inhibited tumor growth, decreased NF-κB p65 and increased
caspase-3 expression, underscoring apoptosis activation as a
central mechanism (52). In the 4T1
breast cancer model, Tan IIA showed antiproliferative and
pro-apoptotic effects associated with changes in key molecules,
including p53, NF-κB, AKT, MYC and BCL-2, as well as enrichment of
the p53, PI3K/AKT, MAPK and mTOR pathways. Molecular docking
further suggested possible interactions between Tan IIA and
proteins such as p53, Bcl-2 and NF-κB; however, these docking
results should be considered hypothesis-generating unless validated
by biochemical binding or in vivo target-engagement assays
(53).
To overcome its poor water solubility and
bioavailability, imidazole groups were introduced to generate a
series of derivatives (TA01-TA12), which significantly inhibited
proliferation, migration and invasion of MDA-MB-231 cells, with
TA12 showing the most potent activity. In zebrafish xenograft
models, TA12 effectively blocked cancer cell metastasis,
potentially through S-phase arrest, ROS generation and DNA damage
induction (54). Further in
vitro and in vivo experiments demonstrate that Tan IIA
inhibits breast cancer cell proliferation in a dose- and
time-dependent manner, markedly reduces colony formation and BrdU
incorporation, and regulates a broad genetic network. Gene
expression profiling revealed that Tan IIA upregulated 41 and
downregulated 24 genes associated with cell cycle regulation,
proliferation, apoptosis, signal transduction, transcriptional
regulation and cell adhesion. In a nude mouse xenograft model of
invasive ductal carcinoma, intraperitoneal administration of Tan
IIA (30 mg/kg) reduced tumor volume by 44.91%, with concomitant
caspase-3 upregulation, indicating that its antitumor effects are
mediated by multi-gene network regulation and apoptosis signaling
activation. These findings collectively support the broad-spectrum
antitumor activity of Tan IIA in both ER-positive and ER-negative
breast cancers (55) (Fig. 3).
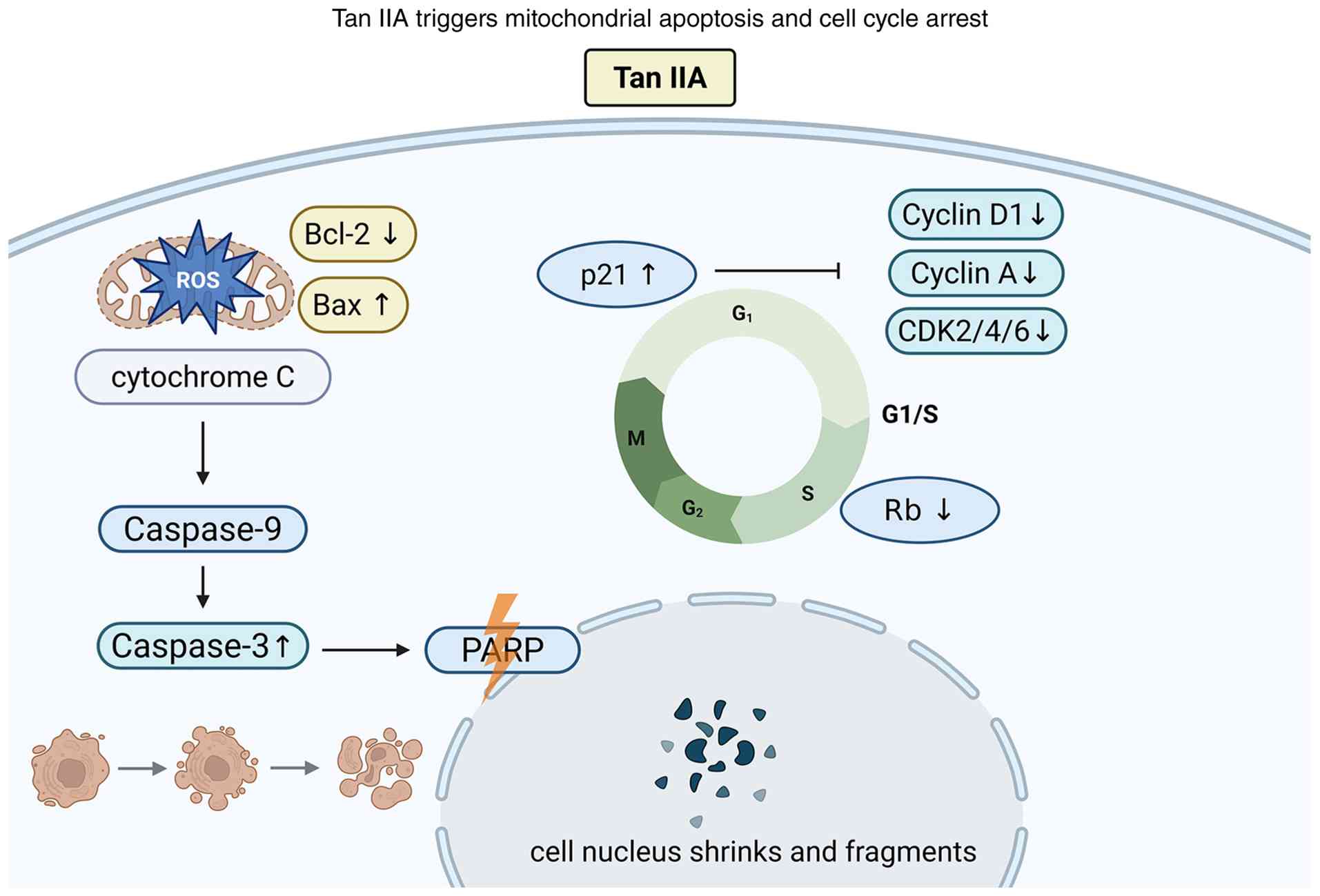 | Figure 3.Tan IIA induces intrinsic apoptosis
via mitochondrial dysfunction and arrests the cell cycle at G1/S
phase by modulating cyclins, CDKs and p21. Tan IIA, tanshinone IIA;
Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma 2;
CDK2/4/6, cyclin-dependent kinase 2/4/6; PARP, poly(ADP-ribose)
polymerase; Rb, retinoblastoma protein; ROS, reactive oxygen
species; ↑, increase; ↓, decrease. |
Targeting BCSCs
BCSCs are critical drivers of drug resistance and
tumor relapse, with inflammatory cytokines and associated signaling
pathways playing key roles in their maintenance (56). The IL-6/STAT3/NF-κB axis is
therefore discussed here as a BCSC-maintenance pathway, while its
contribution to drug resistance is cross-referenced in the
chemosensitization section. Tan IIA, owing to its combined
anticancer and anti-inflammatory activities, has been shown to
markedly inhibit BCSC proliferation and mammosphere formation.
In vitro, Tan IIA treatment significantly reduced BCSC
proliferation and sphere-forming capacity, accompanied by decreased
proportions of CD24−/CD44+ and aldehyde dehydrogenase 1+
subpopulations. Expression levels of inflammatory signaling
proteins, including IL-6, STAT3, phosphorylated STAT3 (Tyr705),
nuclear NF-κB p65 and cyclin D1, were also significantly
suppressed. In vivo, Tan IIA administration inhibited tumor
growth and reduced average tumor weight in xenograft models,
confirming its capacity to effectively target and suppress
BCSC-like populations (57).
Quantitatively, Tan IIA exhibits stronger inhibitory activity
against BCSCs than against parental breast cancer cells, with an
IC50 of ~0.40 µg/ml for BCSCs compared with 0.65 µg/ml for bulk
tumor cells, and an effective in vitro concentration range
of 0.125–2.0 µg/ml (57). In the
study by Li et al (58), Tan
IIA effectively inhibited BCSC-associated traits within a
concentration range of 2.5–20 µM (~0.74–5.92 µg/ml). Mechanistic
analyses further demonstrated that Tan IIA attenuates BCSC stemness
through modulation of the microRNA (miR)-125b/StAR-related lipid
transfer domain protein 13 (STARD13) regulatory axis. Specifically,
Tan IIA downregulates miR-125b while concomitantly upregulating its
target gene STARD13, thereby suppressing miR-125b/STARD13
signaling, reducing stem-like properties and enhancing Dox
sensitivity in breast cancer cells (58). Overexpression of miR-125b or
knockdown of STARD13 reversed the inhibitory effects of Tan IIA on
BCSC stemness, further validating the central role of this
regulatory axis. Collectively, these findings indicate that Tan IIA
suppresses BCSC properties and tumor-initiating capacity through
inhibition of IL-6/STAT3/NF-κB signaling and modulation of the
miR-125b/STARD13 axis, underscoring its therapeutic potential in
overcoming drug resistance and preventing tumor recurrence
(Fig. 4). These findings suggest
that Tan IIA may suppress BCSC-like traits in selected models, but
the durability of this effect and its contribution to relapse
prevention remain to be validated in clinically relevant in
vivo systems.
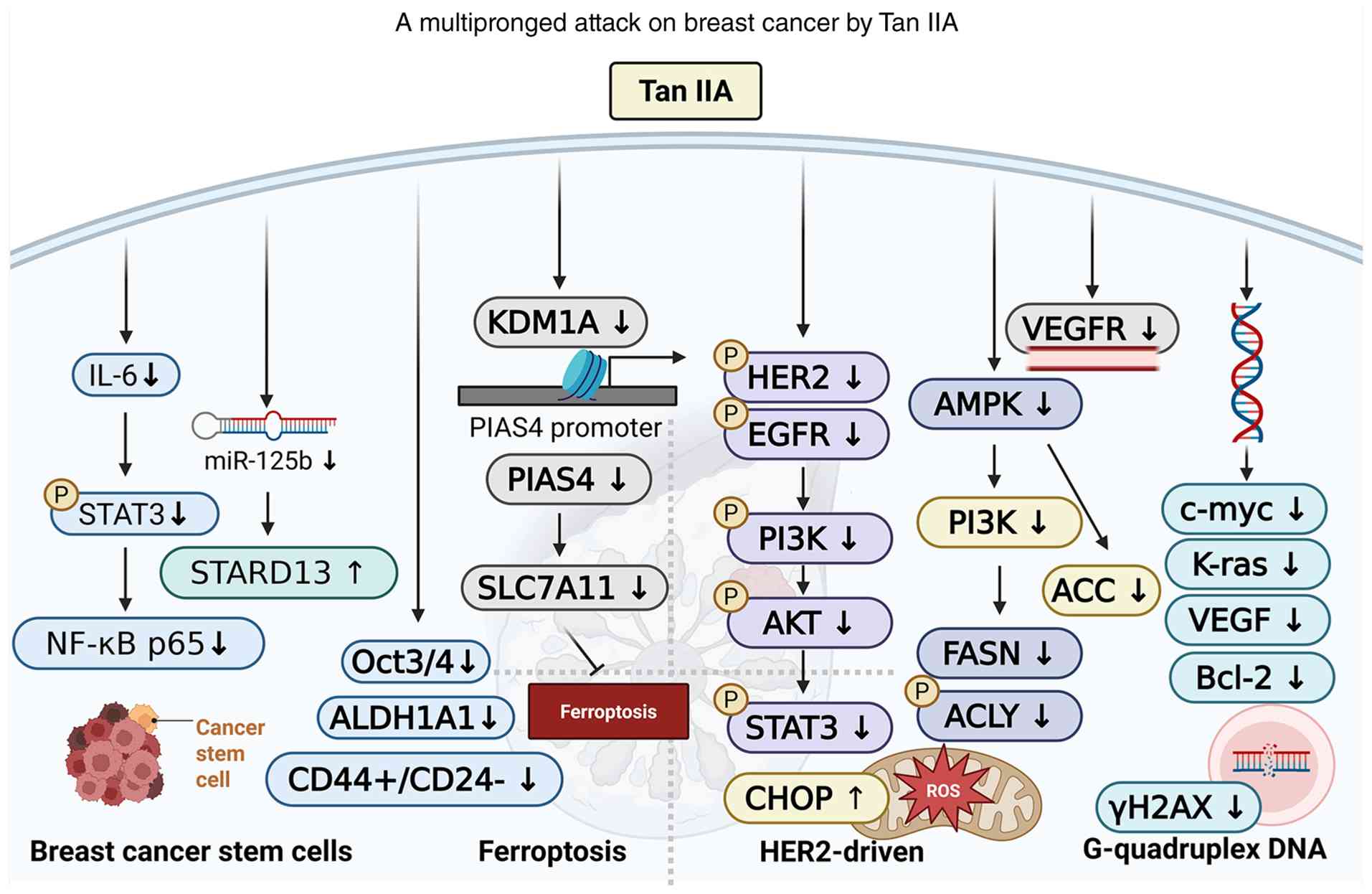 | Figure 4.Tan IIA combats breast cancer through
four reported mechanisms: Targeting breast cancer stem cells,
inhibiting HER2 signaling and metabolic pathways, inducing
ferroptosis and stabilizing G-quadruplex DNA to suppress oncogene
expression. Together, these actions lead to tumor growth
inhibition, enhanced cancer cell death and reduced metastasis.
ACLY, ATP-citrate lyase; AKT, protein kinase B; ALDH1A1, aldehyde
dehydrogenase 1 family member A1; AMPK, AMP-activated protein
kinase; Bcl-2, B-cell lymphoma 2; CHOP, C/EBP homologous protein;
EGFR, epidermal growth factor receptor; FASN, fatty acid synthase;
HER2, human epidermal growth factor receptor 2; IL-6,
interleukin-6; KDM1A, lysine-specific histone demethylase 1A; Kras,
Kirsten rat sarcoma viral oncogene homolog; miR-125b,
microRNA-125b; NF-κB p65, nuclear factor-κB p65 subunit; Oct3/4,
octamer-binding transcription factor 3/4; PI3K, phosphoinositide
3-kinase; PIAS4, protein inhibitor of activated STAT 4; ROS,
reactive oxygen species; SLC7A11, solute carrier family 7 member
11; STAT3, signal transducer and activator of transcription 3;
STARD13, StAR-related lipid transfer domain protein 13; Tan IIA,
tanshinone IIA; VEGFR, vascular endothelial growth factor receptor;
↑, increase; ↓, decrease. |
Induction of ferroptosis
Ferroptosis is an iron-dependent form of programmed
cell death that has emerged as a novel anticancer strategy
(59,60). Tan IIA has recently been linked to
ferroptosis regulation in breast cancer models. Mechanistic studies
showed that Tan IIA downregulated lysine-specific histone
demethylase 1A (KDM1A) expression, thereby reducing the
transcriptional activity of protein inhibitor of activated STAT 4
(PIAS4) and inhibiting PIAS4-mediated SUMOylation of solute carrier
family 7 member 11 (SLC7A11). Destabilization of SLC7A11 impaired
cystine uptake and antioxidant defense, leading to lipid peroxide
accumulation, increased labile iron and ferroptotic cell death in
breast cancer cells (61).
In vitro, Tan IIA treatment suppressed breast
cancer cell proliferation, colony formation and migration, while
in vivo xenograft and lung metastasis models further
supported its tumor-suppressive activity (61). However, ferroptosis-related effects
have been reported over a broad concentration range, including high
micromolar exposures. Therefore, ferroptosis should currently be
regarded as a mechanistically interesting pathway but with
PK-related uncertainty for native Tan IIA, particularly in light of
its poor bioavailability, rapid metabolism and limited evidence of
sustained tumor exposure.
Collectively, these findings identify the
KDM1A/PIAS4/SLC7A11 axis as a potential mechanistic link between
Tan IIA and ferroptotic vulnerability in breast cancer.
Nevertheless, further PK/PD studies are required to determine
whether this pathway can be engaged at achievable tumor
concentrations and whether ferroptosis contributes substantially to
the in vivo antitumor effects of Tan IIA or its optimized
derivatives (Fig. 4).
Suppression of HER2-positive breast
cancer through HER2/EGFR and metabolic signaling
HER2-overexpressing breast cancer cells frequently
depend on HER2/EGFR signaling and downstream anabolic pathways,
including lipid synthesis and protein biosynthesis (62,63).
These metabolic pathways have thus emerged as attractive
therapeutic targets. ATA, a derivative of Tan IIA, demonstrates
superior antitumor activity compared with its parent compound in
HER2-overexpressing breast cancer. In cell lines such as
MDA-MB-453, SK-BR-3 and BT-474, ATA induced G1/S phase arrest and
apoptosis, associated with marked downregulation of EGFR/HER2
receptor tyrosine kinases and inhibition of downstream prosurvival
signaling. Furthermore, ATA triggered oxidative stress and
endoplasmic reticulum stress, while activating the AMP-activated
protein kinase (AMPK) signaling pathway, thereby inactivating key
enzymes involved in lipid and protein biosynthesis and disrupting
metabolic dependencies of tumor cells. In vivo,
intraperitoneal administration of ATA significantly suppressed
xenograft tumor growth in MDA-MB-453 models without inducing body
weight loss or overt toxicity. Functional assays further
demonstrated that ATA inhibited breast cancer cell migration,
invasion and angiogenesis in vitro (64) (Fig.
4). Whether these findings are relevant to HER2-low or
HER2-ultralow breast cancer remains unknown. Given the clinical
expansion of HER2-directed ADCs, future studies should determine
whether Tan IIA derivatives influence ADC sensitivity or resistance
by altering HER2 expression, endocytosis, EMT oxidative stress or
payload susceptibility.
Emerging evidence from non-breast cancer models
suggests that Tan IIA may enhance anti-programmed cell death 1
(PD-1) efficacy by modulating tumor vasculature, endoplasmic
reticulum stress, JNK signaling and immune-cell infiltration
(28,29). However, direct evidence in breast
cancer remains limited, and these findings should be regarded as
hypothesis-generating. Future studies should evaluate whether Tan
IIA-derived compounds can remodel the breast tumor
microenvironment, enhance checkpoint blockade or influence PD-1/PD1
ligand 1 responsiveness in TNBC or other immunologically active
breast cancer subtypes.
Stabilization of G4 DNA
Because of their planar heterocyclic scaffold,
certain Tan IIA derivatives have been investigated as G4
stabilizers. G4 DNA structures represent promising anticancer
targets due to their regulatory roles in oncogene transcription and
translation (65,66). A series of imidazole-modified Tan
IIA derivatives (compounds 1–8), particularly compound 4, exhibited
strong selectivity and binding affinity toward G4 DNA. These
derivatives effectively stabilized multiple G4 structures,
including those in c-Myc, K-ras and VEGF, leading to DNA damage and
suppression of TNBC cell growth, metastasis and angiogenesis.
Molecular docking and interaction studies further confirmed that
these derivatives preferentially bind G4 DNA over duplex DNA,
highlighting their specificity. These results provide a rationale
for developing selected Tan IIA derivatives as G4-targeting leads,
although their pharmacological relevance requires further PK/PD and
target-engagement validation (67)
(Fig. 4). Importantly, G4
stabilization is currently best supported for structurally
optimized derivatives rather than for native Tan IIA itself.
Therefore, this mechanism should be interpreted as a
derivative-specific optimization strategy.
Optimization of derivatives and delivery
systems
Tan IIA and its derivatives have demonstrated
significant pharmacological activity against breast cancer.
However, their poor water solubility, limited bioavailability and
insufficient tumor-targeting capacity remain major obstacles to
clinical application. To address these challenges, researchers have
focused on derivative design and the development of optimized
nanodelivery systems. The objective of these strategies is not
merely to increase cytotoxic potency, but to improve aqueous
stability, systemic exposure, tumor accumulation, target engagement
and therapeutic index. The chemical structures of Tan IIA
derivatives discussed above are summarized in Fig. 5.
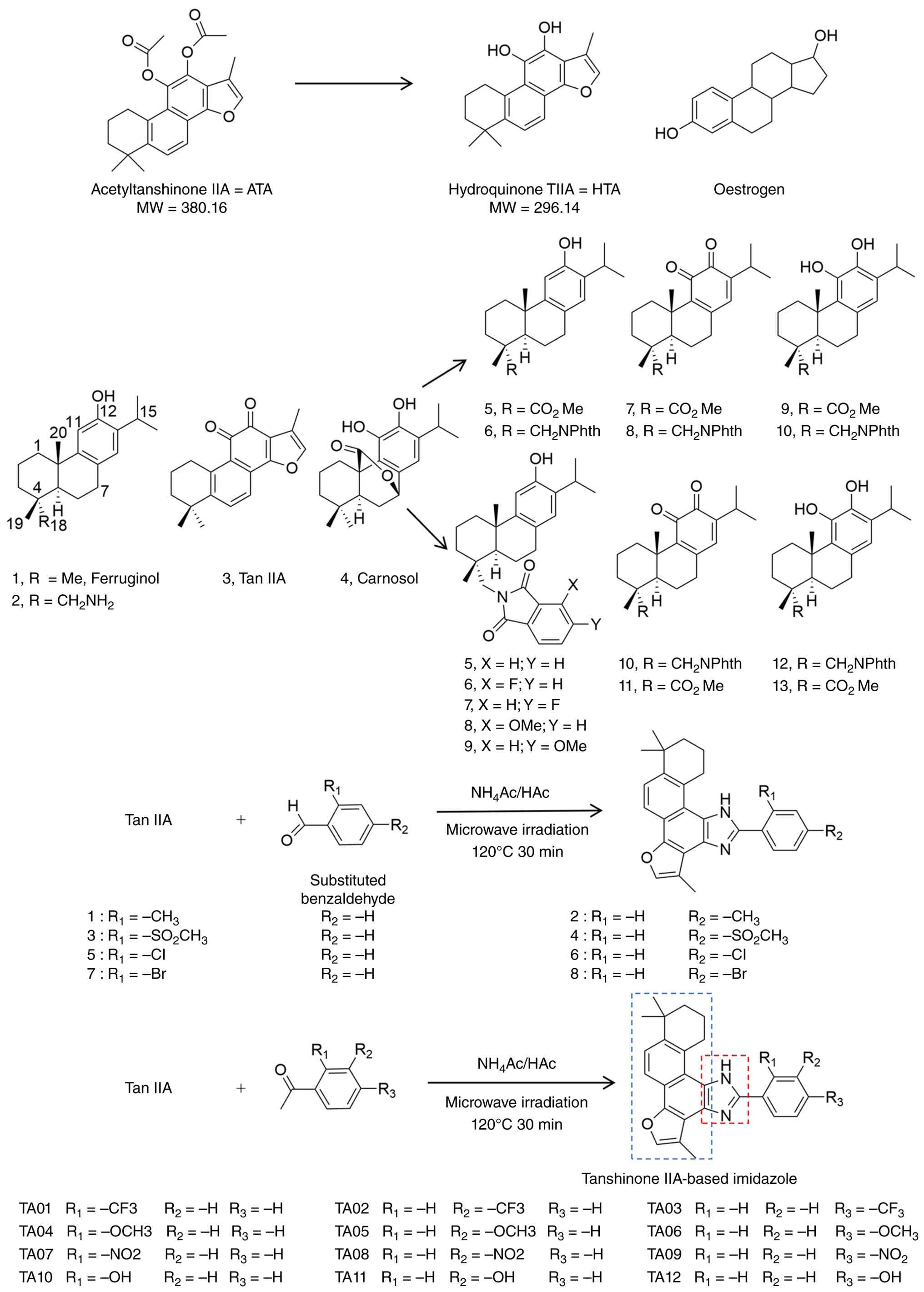 | Figure 5.Chemical structures and
structure-activity relationships of Tan IIA derivatives. Chemical
structures of Tan IIA and its representative derivatives are
discussed in this review, including ATA, its major metabolite HTA
and optimized TA01-TA12. Key modification sites on the Tan IIA
scaffold, such as acetylation, oxidation state changes and
side-chain substitutions, are highlighted to facilitate
structure-activity relationship analysis. In the lower-right
structure, the blue dashed box indicates the Tan IIA-derived core
scaffold and the red dashed box indicates the introduced imidazole
ring. Tan IIA, tanshinone IIA; ATA, acetyltanshinone IIA; HTA,
hydroquinone tanshinone IIA; TA01-TA12, imidazole-substituted Tan
IIA derivatives; MW, molecular weight. |
Regarding derivatives, ATA represents one of the
best-characterized Tan IIA analogues and has shown stronger
inhibitory activity than native Tan IIA in ER-positive breast
cancer models. Its active metabolite, HTA, binds to ERα, promotes
ubiquitin-proteasome-dependent ERα degradation in the nucleus and
downregulates ESR1 transcription, thereby distinguishing this
mechanism from that of classical endocrine agents such as tamoxifen
and fulvestrant (21). ATA has also
demonstrated activity in HER2-overexpressing breast cancer cells,
where it induces G1/S cell-cycle arrest and apoptosis through
downregulation of EGFR/HER2 and downstream survival pathways,
together with ER stress and AMPK activation (64). Other semisynthetic Tan IIA
derivatives have exhibited IC50 values ranging from 1.3
to 18.7 µM across breast cancer cell lines including SUM149,
MDA-MB-231, T47D and MCF-7 cells (68,69).
Imidazole-substituted derivatives, particularly TA12, further
inhibited proliferation, migration and invasion of MDA-MB-231 cells
and suppressed metastasis in zebrafish xenograft models, with
effects associated with S-phase arrest, ROS accumulation and DNA
damage activation (54). In
addition, selected imidazole-modified derivatives have been
reported to stabilize G4 structures within oncogene promoters such
as MYC, KRAS and VEGF, leading to DNA damage and suppression of
TNBC-associated proliferation, migration, invasion and angiogenesis
(67). However, these findings
should be interpreted as derivative-specific optimization
strategies rather than uniform evidence that all Tan IIA analogues
share the same mechanism or translational potential. Table SII summarizes representative Tan
IIA derivatives and delivery systems, showing that ATA exhibits
low-micromolar activity in ER-positive and HER2-overexpressing
breast cancer cells, while optimized derivatives such as TA12 and
compound 4 show selective activity in MDA-MB-231 models (21,54,64,67).
Delivery systems such as T/CM-L and Tan-Dox-MSN further improved
formulation performance and tumor inhibition in vivo,
although further pharmacokinetic, safety and target-engagement
validation remains necessary before clinical translation (22,23).
With respect to delivery systems, several innovative
strategies have been developed to enhance the therapeutic efficacy
of Tan IIA. A multi-component liposomal system co-loaded with
sodium Tan IIA sulfonate (STS) and celastrol (CM) was designed to
achieve sequential release: STS first modulated the tumor
microenvironment, followed by CM-mediated tumor cell killing. This
dual-delivery approach produced synergistic anticancer effects,
significantly reducing tumor volume while minimizing systemic
toxicity (23). In another
approach, a phytoestrogen-modified nanodelivery system
(Tan-Dox-MSN) was constructed to co-deliver Tan IIA derivatives and
Dox. This platform exhibited excellent tumor-targeting properties
and biocompatibility, improved Dox loading efficiency, and markedly
suppressed breast cancer growth and metastasis (22). Although these systems improve tumor
delivery in preclinical models, carrier-related immunogenicity,
reticuloendothelial uptake, organ accumulation and long-term
tolerability remain insufficiently characterized.
Together, these findings suggest that molecular
modification and delivery optimization may improve the therapeutic
feasibility of Tan IIA-based strategies. However, their clinical
relevance remains dependent on quantitative PK/PD validation,
safety assessment and demonstration of target engagement in
vivo.
Structure-activity relationships of Tan IIA
and its derivatives
Systematic structural modification of Tan IIA has
revealed several recurrent structure-activity relationship (SAR)
patterns that critically determine its anticancer potency,
selectivity and mechanistic profile in breast cancer models. At the
core scaffold level, acetylation of Tan IIA to generate ATA
markedly enhances subtype-selective activity, particularly against
ER-positive and HER2-overexpressing breast cancer cells, while
maintaining lower toxicity toward non-malignant mammary epithelial
cells (21,64). This minor modification confers a
dual mechanism involving ERα protein degradation and
transcriptional suppression, as well as inhibition of EGFR/HER2
signaling and downstream metabolic and survival pathways,
highlighting acetylation as an effective strategy to modulate both
target engagement and pharmacological behavior. Beyond core
modifications, the oxidation state at the C11-C12 positions of the
C ring emerges as a dominant determinant of cytotoxic potency.
Conversion of phenolic hydroxyl groups to ortho-quinone structures
consistently enhances antiproliferative activity, particularly in
TNBC cell lines, likely through redox cycling, ROS generation and
mitochondrial dysfunction (68).
Side-chain modifications at the C18 position further fine-tune
activity and selectivity, with bulky substituents such as
phthalimide groups synergizing with ortho-quinone or catechol cores
to improve solubility, therapeutic index and induction of intrinsic
apoptosis (68,69). In parallel, aromatic substitution
patterns in imidazole-fused and phenyl-substituted derivatives
demonstrate that para-oriented substituents-either
electron-withdrawing or electron-donating, depending on the
scaffold-can enhance molecular planarity, optimize electronic
distribution and promote selective targeting of aggressive TNBC
cells through mechanisms such as G4 DNA stabilization or
suppression of migration- and invasion-related signaling axes
(54,67).
Despite these encouraging SAR trends, several
important limitations must be acknowledged when interpreting the
current data and translating them into rational drug design
strategies. Most SAR conclusions are derived from heterogeneous
in vitro systems, with limited consistency in cell lines,
exposure durations and dose reporting, restricting direct
quantitative comparison across studies. Furthermore, while specific
structural motifs-such as ortho-quinone cores, C18 side-chain
extensions and para-substituted aromatic rings-are recurrently
associated with enhanced activity, their precise molecular targets
and off-target liabilities remain insufficiently defined. The
chemical instability of Tan IIA derivatives in aqueous environments
and the lack of comprehensive PK and metabolic profiling further
constrain their clinical translatability. Consequently, existing
SAR insights should be regarded as guiding principles rather than
definitive design rules. Future efforts should integrate rational
structural optimization with systematic PK evaluation,
multi-omics-based target deconvolution and in vivo
validation to balance potency, selectivity and drug-likeness,
thereby enabling the development of next-generation, patentable Tan
IIA-derived candidates for difficult-to-treat breast cancer
subtypes (21,54,64,68,69).
Therefore, future SAR studies should routinely report
IC50 values in malignant and non-malignant cells,
solubility, microsomal stability, plasma exposure, tumor
accumulation and preliminary safety margins.
Conclusion, safety considerations and
perspectives
In conclusion, while Tan IIA displays numerous
anticancer actions in cell and animal experiments, its practical
drug potential is severely limited. Its intrinsic drawbacks,
including poor solubility, rapid metabolism and low
bioavailability, mean that native Tan IIA is unlikely to be
developed as a standalone anticancer drug (70). Instead, the value of Tan IIA lies in
its scaffolding: Derivatives and formulations built on its
structure can achieve potent activity. Furthermore, no
well-controlled clinical studies have validated Tan IIA's efficacy
in cancer patients. Because unmodified Tan IIA is a natural
compound, patent protection for it is weak, further reducing
incentives for costly drug development (71). Going forward, research should
prioritize optimized analogues (with better ADME profiles) and
advanced delivery systems, while rigorously testing these in
preclinical models (72–74).
It must be emphasized that Tan IIA's anticancer data
are entirely preclinical. Most studies have used cell lines or
mouse models (75–77). To move forward, rigorous PK and
toxicology studies in animals are needed, followed by early-phase
human trials to determine whether any benefit translates to
patients. In addition, the multiplicity of Tan IIA's purported
targets (from ERα to KDM1A to G4 DNA) raises the possibility of
off-target pharmacology and unintended pathway perturbation,
particularly at exposures required for anticancer efficacy. Future
development should therefore define a feasible therapeutic window
and systematically evaluate safety liabilities, including potential
endocrine-related effects given its ER-binding and
phytoestrogen-like properties, mitochondrial and oxidative
stress-related toxicity, quinone-associated electrophilic
reactivity, cardiovascular interference and drug-drug interactions
when combined with chemotherapeutic, endocrine, HER2-targeted or
immune checkpoint inhibitors. For advanced delivery systems,
carrier-related immunogenicity, reticuloendothelial uptake, organ
accumulation and long-term tolerability also warrant careful
assessment. Finally, even with derivatives, Tan IIA analogues tend
to remain chemically challenging because of poor solubility and
stability issues. Without highly potent lead compounds or delivery
systems capable of achieving sustained tumor exposure, it may be
difficult to reach therapeutically relevant concentrations in
humans. In short, Tan IIA highlights how a natural scaffold can
inspire drug design, but also why its direct clinical utility
remains constrained (78).
Pathways inhibited in tumor cells may also have
essential physiological functions in normal tissues. ER signaling
contributes to reproductive, skeletal, cardiovascular and
immune-inflammatory homeostasis; therefore, systemic ER-modulating
activity may have consequences in female subjects beyond tumor
inhibition. Similarly, suppression of NF-κB, STAT3, PI3K/AKT or
redox signaling may affect immune competence, hematopoietic
function, tissue repair and inflammatory balance. These risks may
be amplified in combination with chemotherapy, endocrine therapy,
HER2-targeted therapy or immunotherapy.
In summary, Tan IIA serves as a valuable
pharmacological probe but is not itself a drug candidate without
significant improvement. Any clinical application will depend on
developing semisynthetic analogues with truly drug-like properties
(much greater potency and stability) and robust delivery methods.
Given its low patentability in native form, future efforts will
likely focus on novel derivatives. Meanwhile, caution is warranted:
Claims of ‘promising anticancer potential’ should be tempered by
the compound's PK realities and the absence of clinical proof. Only
compounds or formulations that demonstrate achievable tumor
exposure, target engagement, acceptable safety margins and
reproducible efficacy in clinically relevant breast cancer models
should be advanced toward clinical development.
Supplementary Material
Supporting Data
Acknowledgements
The figures were created with Biorender app (web
application; http://www.biorender.com). No AI tools or AI-assisted
technologies were used in the writing of this manuscript.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
XL was responsible for manuscript writing and figure
preparation, and conducted the literature review and data
collection. LF contributed to manuscript revision and proofreading,
supervised the overall study design, provided guidance on the
research direction, oversaw the manuscript drafting and provided
final approval of the submitted version. Data authentication is not
applicable. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ABC
|
ATP-binding cassette
|
|
ADC
|
antibody-drug conjugate
|
|
ADME
|
absorption, distribution, metabolism
and excretion
|
|
AKT
|
protein kinase B
|
|
AMPK
|
AMP-activated protein kinase
|
|
ATA
|
acetyltanshinone IIA
|
|
BCSCs
|
breast cancer stem cells
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
Bcl-xL
|
B-cell lymphoma-extra large
|
|
BCRP
|
breast cancer resistance protein
|
|
BrdU
|
5-bromo-2′-deoxyuridine
|
|
CD44+/CD24-
|
cluster of differentiation 44
positive/cluster of differentiation 24 low or negative
|
|
CHOP
|
C/EBP homologous protein
|
|
CM
|
celastrol
|
|
Dox
|
doxorubicin
|
|
EGFR
|
epidermal growth factor receptor
|
|
EMT
|
epithelial-mesenchymal transition
|
|
ERα
|
estrogen receptor α
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
ESR1
|
estrogen receptor 1
|
|
FES
|
fluoroestradiol
|
|
G4
|
G-quadruplex
|
|
GADD153
|
growth arrest and DNA damage-inducible
protein 153
|
|
GPER
|
G protein-coupled estrogen
receptor
|
|
GREB1
|
growth regulating estrogen receptor
binding 1
|
|
HER2
|
human epidermal growth factor receptor
2
|
|
HIF-1α
|
hypoxia-inducible factor 1α
|
|
HTA
|
hydroquinone tanshinone IIA
|
|
IC50
|
half-maximal inhibitory
concentration
|
|
IL-6
|
interleukin 6
|
|
JNK
|
c-Jun N-terminal kinase
|
|
KDM1A
|
lysine demethylase 1A
|
|
MAPK
|
mitogen-activated protein kinase
|
|
MRP1
|
multidrug resistance-associated
protein 1
|
|
MSN
|
mesoporous silica nanoparticles
|
|
MYC
|
MYC proto-oncogene
|
|
NF-κB
|
nuclear factor κB
|
|
PARP
|
poly(ADP-ribose) polymerase
|
|
PD
|
pharmacodynamic
|
|
PD-1
|
programmed cell death protein 1
|
|
PD-L1
|
programmed death-ligand 1
|
|
PET/CT
|
positron emission tomography/computed
tomography
|
|
P-gp
|
P-glycoprotein
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
PIAS4
|
protein inhibitor of activated STAT
4
|
|
PK
|
pharmacokinetic
|
|
PTEN
|
phosphatase and tensin homolog
|
|
ROS
|
reactive oxygen species
|
|
SAR
|
structure-activity relationship
|
|
SLC7A11
|
solute carrier family 7 member 11
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
STS
|
sodium tanshinone IIA sulfonate
|
|
SUMOylation
|
small ubiquitin-like modifier
conjugation
|
|
TA01-TA12
|
imidazole-substituted tanshinone IIA
derivatives
|
|
Tan IIA
|
tanshinone IIA
|
|
Tan-Dox-MSN
|
tanshinone IIA-doxorubicin-loaded
mesoporous silica nanoparticles
|
|
Tan-NH2
|
amino-modified tanshinone IIA
|
|
Taxol
|
paclitaxel
|
|
TNBC
|
triple-negative breast cancer
|
|
TWIST
|
TWIST family basic helix-loop-helix
transcription factor
|
|
VEGF
|
vascular endothelial growth
factor
|
References
|
1
|
Liu Y, Zong X, Altea-Manzano P and Fu J:
Amino acid metabolism in breast cancer: Pathogenic drivers and
therapeutic opportunities. Protein Cell. 16:506–531. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xiong X, Zheng LW, Ding Y, Chen YF, Cai
YW, Wang LP, Huang L, Liu CC, Shao ZM and Yu KD: Breast cancer:
Pathogenesis and treatments. Signal Transduct Target Ther.
10:492025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barzaman K, Karami J, Zarei Z,
Hosseinzadeh A, Kazemi MH, Moradi-Kalbolandi S, Safari E and
Farahmand L: Breast cancer: Biology, biomarkers, and treatments.
Int Immunopharmacol. 84:1065352020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kawiak A: Molecular research and treatment
of breast cancer. Int J Mol Sci. 23:96172022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Habibi S, Bahramian S, Zare Jalise S,
Mehri S, Ababzadeh S and Kavianpour M: Novel strategies in breast
cancer management: From treatment to long-term remission. Crit Rev
Oncol Hematol. 211:1047152025. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Y and Minden A: Current molecular
combination therapies used for the treatment of breast cancer. Int
J Mol Sci. 23:110462022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Naeem A, Hu P, Yang M, Zhang J, Liu Y, Zhu
W and Zheng Q: Natural products as anticancer agents: Current
status and future perspectives. Molecules. 27:83672022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen W and Chen G: Danshen (Salvia
miltiorrhiza Bunge): A prospective healing sage for
cardiovascular diseases. Curr Pharm Des. 23:5125–5135.
2017.PubMed/NCBI
|
|
9
|
Wei B, Sun C, Wan H, Shou Q, Han B, Sheng
M, Li L and Kai G: Bioactive components and molecular mechanisms of
Salvia miltiorrhiza Bunge in promoting blood circulation to
remove blood stasis. J Ethnopharmacol. 317:1166972023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu T, Zou HX, Le SY, Wang YR, Qiao YM,
Yuan Y, Liu JC, Lai SQ and Huang H: Tanshinone IIA confers
protection against myocardial ischemia/reperfusion injury by
inhibiting ferroptosis and apoptosis via VDAC1. Int J Mol Med.
52:1092023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang X, Deng H, Shen QK and Quan ZS:
Tanshinone IIA: Pharmacology, total synthesis, and progress in
Structure-modifications. Curr Med Chem. 29:1959–1989. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Han D, Chen YB, Zhao K, Li HZ, Chen XY,
Zhu GZ, Tu C, Gao JW, Zhuang JS, Wu ZY and Zhong ZM: Tanshinone IIA
alleviates inflammation-induced skeletal muscle atrophy by
regulating mitochondrial dysfunction. Arch Biochem Biophys.
762:1102152024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma X, Zhang L, Gao F, Jia W and Li C:
Salvia miltiorrhiza and Tanshinone IIA reduce endothelial
inflammation and atherosclerotic plaque formation through
inhibiting COX-2. Biomed Pharmacother. 167:1155012023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shan B, Guo C, Zhou H and Chen J:
Tanshinone IIA alleviates pulmonary fibrosis by modulating
glutamine metabolic reprogramming based on
[U-13C5]-glutamine metabolic flux analysis. J
Adv Res. 70:531–544. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Subedi L and Gaire BP: Tanshinone IIA: A
phytochemical as a promising drug candidate for neurodegenerative
diseases. Pharmacol Res. 169:1056612021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang W, Liu C, Li J, Lu Y, Li H, Zhuang
J, Ren X, Wang M and Sun C: Tanshinone IIA: New perspective on the
anti-tumor mechanism of a traditional natural medicine. Am J Chin
Med. 50:209–239. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Alam SSM, Samanta A, Uddin F, Ali S and
Hoque M: Tanshinone IIA targeting cell signaling pathways: A
plausible paradigm for cancer therapy. Pharmacol Rep. 75:907–922.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu W, Si Y, Wen X, Lin D, Yu Z, Xie X and
Xu J: Pharmacological effects and mechanisms of Tanshinone IIA in
bone injury repair. Pharmaceuticals (Basel). 18:13382025.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shakeri A, Hafezian T, Kúsz N, Hohmann J,
Boozari M, Mottaghipisheh J, Emami SA, Tayarani-Najaran Z and Asili
J: Cytotoxicity, apoptosis inducing activity and Western blot
analysis of tanshinone derivatives from Stachys parviflora
on prostate and breast cancer cells. Mol Biol Rep. 49:8251–8258.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Su CC and Lin YH: Tanshinone IIA inhibits
human breast cancer cells through increased Bax to Bcl-xL ratios.
Int J Mol Med. 22:357–361. 2008.PubMed/NCBI
|
|
21
|
Yu T, Zhou Z, Mu Y, de Lima Lopes G and
Luo KQ: A novel anti-cancer agent, acetyltanshinone IIA, inhibits
oestrogen receptor positive breast cancer cell growth by
down-regulating the oestrogen receptor. Cancer Lett. 346:94–103.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Pan H, Yu C, Liu R, Xing B, Jia
B, He J, Jia X, Feng X, Zhang Q, et al: Phytoestrogen-derived
multifunctional ligands for targeted therapy of breast cancer.
Asian J Pharm Sci. 18:1008272023.PubMed/NCBI
|
|
23
|
Qu D, Wang L, Qin Y, Guo M, Guo J, Huang
M, Liu Y, Liu C, Li H and Chen Y: Non-triggered sequential-release
liposomes enhance anti-breast cancer efficacy of STS and
celastrol-based microemulsion. Biomater Sci. 6:3284–3299. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo R, Li L, Su J, Li S, Duncan SE, Liu Z
and Fan G: Pharmacological activity and mechanism of Tanshinone IIA
in related diseases. Drug Des Devel Ther. 14:4735–4748. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ansari MA, Khan FB, Safdari HA, Almatroudi
A, Alzohairy MA, Safdari M, Amirizadeh M, Rehman S, Equbal MJ and
Hoque M: Prospective therapeutic potential of Tanshinone IIA: An
updated overview. Pharmacol Res. 164:1053642021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang C, Mu Y, Li S, Zhang Y, Liu X and Li
J: Tanshinone IIA: A Chinese herbal ingredient for the treatment of
atherosclerosis. Front Pharmacol. 14:13218802023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao JY, Pu J, Fan J, Feng XY, Xu JW,
Zhang R and Shang Y: Tanshinone IIA prevents acute lung injury by
regulating macrophage polarization. J Integr Med. 20:274–280. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mao D, Wang H, Guo H, Che X, Chen M, Li X,
Liu Y, Huo J and Chen Y: Tanshinone IIA normalized hepatocellular
carcinoma vessels and enhanced PD-1 inhibitor efficacy by
inhibiting ELTD1. Phytomedicine. 123:1551912024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang YZ, Lai HL, Huang C, Jiang ZB, Yan
HX, Wang XR, Xie C, Huang JM, Ren WK, Li JX, et al: Tanshinone IIA
induces ER stress and JNK activation to inhibit tumor growth and
enhance anti-PD-1 immunotherapy in non-small cell lung cancer.
Phytomedicine. 128:1554312024. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bi Z, Wang Y and Zhang W: A comprehensive
review of tanshinone IIA and its derivatives in fibrosis treatment.
Biomed Pharmacother. 137:1114042021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fan Y, Kang S, Shao T, Xu L and Chen J:
Activation of SIRT3 by Tanshinone IIA ameliorates renal fibrosis by
suppressing the TGF-β/TSP-1 pathway and attenuating oxidative
stress. Cell Signal. 122:1113482024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li H, Wu M, Guo C, Zhai R and Chen J:
Tanshinone IIA regulates Keap1/Nrf2 signal pathway by activating
Sestrin2 to restrain pulmonary fibrosis. Am J Chin Med.
50:2125–2151. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chan P, Liu JC, Lin LJ, Chen PY, Cheng TH,
Lin JG and Hong HJ: Tanshinone IIA inhibits angiotensin II-induced
cell proliferation in rat cardiac fibroblasts. Am J Chin Med.
39:381–394. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang P, Liu W and Wang Y: The mechanisms
of tanshinone in the treatment of tumors. Front Pharmacol.
14:12822032023. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fang ZY, Zhang M, Liu JN, Zhao X, Zhang YQ
and Fang L: Tanshinone IIA: A review of its anticancer effects.
Front Pharmacol. 11:6110872020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bardia A, Kaklamani V, Wilks S, Weise A,
Richards D, Harb W, Osborne C, Wesolowski R, Karuturi M, Conkling
P, et al: Phase I study of Elacestrant (RAD1901), a novel selective
estrogen receptor degrader, in ER-positive, HER2-negative advanced
breast cancer. J Clin Oncol. 39:1360–1370. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He S, Wang M and Zhang Y, Luo J and Zhang
Y: Monitoring the early response of fulvestrant plus Tanshinone IIA
combination therapy to estrogen receptor-positive breast cancer by
longitudinal 18F-FES PET/CT. Contrast Media Mol Imaging.
2019:23745652019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lu Q, Zhang P, Zhang X and Chen J:
Experimental study of the anti-cancer mechanism of tanshinone IIA
against human breast cancer. Int J Mol Med. 24:773–780. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bisht A, Avinash D, Sahu KK, Patel P, Das
Gupta G and Kurmi BD: A comprehensive review on doxorubicin:
Mechanisms, toxicity, clinical trials, combination therapies and
nanoformulations in breast cancer. Drug Deliv Transl Res.
15:102–133. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dan VM, Raveendran RS and Baby S:
Resistance to intervention: Paclitaxel in breast cancer. Mini Rev
Med Chem. 21:1237–1268. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lu H, Xie Y, Tran L, Lan J, Yang Y,
Murugan NL, Wang R, Wang YJ and Semenza GL: Chemotherapy-induced
S100A10 recruits KDM6A to facilitate OCT4-mediated breast cancer
stemness. J Clin Invest. 130:4607–4623. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li K, Liu W, Zhao Q, Wu C, Fan C, Lai H
and Li S: Combination of tanshinone IIA and doxorubicin possesses
synergism and attenuation effects on doxorubicin in the treatment
of breast cancer. Phytother Res. 33:1658–1669. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li S, Wu C, Fan C, Zhang P, Yu G and Li K:
Tanshinone II A improves the chemosensitivity of breast cancer
cells to doxorubicin by inhibiting β-catenin nuclear translocation.
J Biochem Mol Toxicol. 35:e226202021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lin H, Zheng L, Li S, Xie B, Cui B, Xia A,
Lin Z and Zhou P: Cytotoxicity of Tanshinone IIA combined with
Taxol on drug-resist breast cancer cells MCF-7 through inhibition
of Tau. Phytother Res. 32:667–671. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li S, FeiyuTeng Zhang J, Zhang P, Li M,
Wang X and Li K: Tanshinone IIA potentiates the chemotherapeutic
effect of doxorubicin against breast cancer cells and attenuates
the cardiotoxicity of doxorubicin by regulating ERK1/2 pathway. J
Biochem Mol Toxicol. 38:e238512024. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li K and Lai H: TanshinoneIIA enhances the
chemosensitivity of breast cancer cells to doxorubicin through
down-regulating the expression of MDR-related ABC transporters.
Biomed Pharmacother. 96:371–377. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fu P, Du F, Chen W, Yao M, Lv K and Liu Y:
Tanshinone IIA blocks epithelial-mesenchymal transition through
HIF-1α downregulation, reversing hypoxia-induced chemotherapy
resistance in breast cancer cell lines. Oncol Rep. 31:2561–2568.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Parton M, Dowsett M and Smith I: Studies
of apoptosis in breast cancer. BMJ. 322:1528–1532. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nagalla S, Chou JW, Willingham MC, Ruiz J,
Vaughn JP, Dubey P, Lash TL, Hamilton-Dutoit SJ, Bergh J, Sotiriou
C, et al: Interactions between immunity, proliferation and
molecular subtype in breast cancer prognosis. Genome Biol.
14:R342013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yan MY, Chien SY, Kuo SJ, Chen DR and Su
CC: Tanshinone IIA inhibits BT-20 human breast cancer cell
proliferation through increasing caspase 12, GADD153 and
phospho-p38 protein expression. Int J Mol Med. 29:855–863.
2012.PubMed/NCBI
|
|
51
|
He Y, Yang K, Liu J, Shi D, Zhang Z, Yang
J, Chen M and Zhao P: Tanshinone IIA inhibits triple-negative
breast cancer cells MDA-MB-231 via G protein-coupled estrogen
receptor-(GPER-) dependent signaling pathway. Dis Markers.
2023:83716232023. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Su CC, Chien SY, Kuo SJ, Chen YL, Cheng CY
and Chen DR: Tanshinone IIA inhibits human breast cancer MDA-MB-231
cells by decreasing LC3-II, Erb-B2 and NF-κBp65. Mol Med Rep.
5:1019–1022. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu J, Zhang C, Liu S, Wang X, Wu X and
Hao J: Tanshinone IIA promotes apoptosis by downregulating BCL2 and
upregulating TP53 in triple-negative breast cancer. Naunyn
Schmiedebergs Arch Pharmacol. 396:365–374. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu Q, Zheng K, Huang X, Li L and Mei W:
Tanshinone-IIA-based analogues of imidazole alkaloid act as potent
inhibitors to block breast cancer invasion and metastasis in vivo.
J Med Chem. 61:10488–10501. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang X, Wei Y, Yuan S, Liu G, Lu Y, Zhang
J and Wang W: Potential anticancer activity of tanshinone IIA
against human breast cancer. Int J Cancer. 116:799–807. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang C, Xu K, Wang R, Han X, Tang J and
Guan X: Heterogeneity of BCSCs contributes to the metastatic
organotropism of breast cancer. J Exp Clin Cancer Res. 40:3702021.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lin C, Wang L, Wang H, Yang L, Guo H and
Wang X: Tanshinone IIA inhibits breast cancer stem cells growth in
vitro and in vivo through attenuation of IL-6/STAT3/NF-kB signaling
pathways. J Cell Biochem. 114:2061–2070. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Li X, Jia Q, Zhou Y, Jiang X, Song L, Wu
Y, Wang A, Chen W, Wang S and Lu Y: Tanshinone IIA attenuates the
stemness of breast cancer cells via targeting the miR-125b/STARD13
axis. Exp Hematol Oncol. 11:22022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Xiang S, Yan W, Ren X, Feng J and Zu X:
Role of ferroptosis and ferroptosis-related long non'coding RNA in
breast cancer. Cell Mol Biol Lett. 29:402024. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang F, Xiao Y, Ding JH, Jin X, Ma D, Li
DQ, Shi JX, Huang W, Wang YP, Jiang YZ and Shao ZM: Ferroptosis
heterogeneity in triple-negative breast cancer reveals an
innovative immunotherapy combination strategy. Cell Metab.
35:84–100.e8. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Luo N, Zhang K, Li X, Hu Y and Guo L:
Tanshinone IIA destabilizes SLC7A11 by regulating PIAS4-mediated
SUMOylation of SLC7A11 through KDM1A, and promotes ferroptosis in
breast cancer. J Adv Res. 69:313–327. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Agostinetto E, Curigliano G and Piccart M:
Emerging treatments in HER2-positive advanced breast cancer: Keep
raising the bar. Cell Rep Med. 5:1015752024. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kang S and Kim SB: HER2-low breast cancer:
Now and in the future. Cancer Res Treat. 56:700–720. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Guerram M, Jiang ZZ, Yousef BA, Hamdi AM,
Hassan HM, Yuan ZQ, Luo HW, Zhu X and Zhang LY: The potential
utility of acetyltanshinone IIA in the treatment of
HER2-overexpressed breast cancer: Induction of cancer cell death by
targeting apoptotic and metabolic signaling pathways. Oncotarget.
6:21865–21877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hänsel-Hertsch R, Simeone A, Shea A, Hui
WWI, Zyner KG, Marsico G, Rueda OM, Bruna A, Martin A, Zhang X, et
al: Landscape of G-quadruplex DNA structural regions in breast
cancer. Nat Genet. 52:878–883. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kosiol N, Juranek S, Brossart P, Heine A
and Paeschke K: G-quadruplexes: A promising target for cancer
therapy. Mol Cancer. 20:402021. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zeng L, Wu Q, Wang T, Li LP, Zhao X, Chen
K, Qian J, Yuan L, Xu H and Mei WJ: Selective stabilization of
multiple promoter G-quadruplex DNA by using
2-phenyl-1H-imidazole-based tanshinone IIA derivatives and their
potential suppressing function in the metastatic breast cancer.
Bioorg Chem. 106:1044332021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gonzalez-Cardenete MA, Gonzalez-Zapata N,
Boyd L and Rivas F: Discovery of novel bioactive tanshinones and
carnosol analogues against breast cancer. Cancers (Basel).
15:13182023. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Gonzalez-Cardenete MA, Mendoza-Hernandez
WE, Lawson SL and Rivas F: In vitro effect of ferruginol,
tanshinone, and carnosol analogues on the proliferation of three
breast cancer cell lines. Molecules. 30:25292025. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Jia Q, Zhu R, Tian Y, Chen B, Li R, Li L,
Wang L, Che Y, Zhao D, Mo F, et al: Salvia miltiorrhiza in
diabetes: A review of its pharmacology, phytochemistry, and safety.
Phytomedicine. 58:1528712019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang Z, Raorao L, Chen Y, Yang H,
Fitzgerald M, Wang Q, Xu Z, Huang N, Lu D and Luo L: Integration of
traditional, complementary, and alternative medicine with modern
biomedicine: The scientization, evidence, and challenges for
integration of traditional Chinese medicine. Acupunct Herbal Med.
4:68–78. 2024. View Article : Google Scholar
|
|
72
|
Zhu YF, He MQ, Lin CS, Ma WH, Ai Y, Wang J
and Liang Q: Multifunctional Nanocarrier drug delivery systems:
From diverse design to precise biomedical applications. Adv Healthc
Mater. 15:e021782025. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Mittal M, Juneja S, Pandey N and Mittal R:
Nanoparticle-based drug delivery systems: Current advances and
future directions. Curr Drug Targets. 27:150–172. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Lv L, Shi Y, Wu J and Li G: Nanosized drug
delivery systems for breast cancer stem cell targeting. Int J
Nanomedicine. 16:1487–1508. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Huang J, Zhang J, Sun C, Yang R, Sheng M,
Hu J, Kai G and Han B: Adjuvant role of Salvia miltiorrhiza
bunge in cancer chemotherapy: A review of its bioactive components,
health-promotion effect and mechanisms. J Ethnopharmacol.
318:1170222024. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Su Z, Li Y, Zhou Z, Feng B, Chen H and
Zheng G: Herbal medicine for colorectal cancer treatment: Molecular
mechanisms and clinical applications. Cell Prolif. 58:e700652025.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Xu S and Liu P: Tanshinone II-A: New
perspectives for old remedies. Expert Opin Ther Pat. 23:149–153.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kannan S and Cheng VWT: Nanoparticle drug
delivery to target breast cancer brain metastasis: Current and
future trends. Int J Cancer. 153:1118–1129. 2023. View Article : Google Scholar : PubMed/NCBI
|














