Introduction
Aging is a complex and varied process that involves
the accumulation of numerous molecular changes and resulting
pathological alterations to normal physiological function (1). These aging-associated changes involve
various tissues, for example skin, bones and muscles, and have
attracted much attention in the investigation of the mechanisms
involved in the pathophysiology of aging-associated disorders,
including age-induced osteoporosis (2-5).
Tendons are fibrous tissues that connect muscle to bone, and are
able to resist high levels of force. Their primary function is to
transfer muscle-generated force to the skeleton, facilitating
movement around a joint. Notably, there are clinical data
demonstrating the association between aging and an increased
incidence of tendon rupture (6);
however, to the best of our knowledge, basic studies, animal models
and clinical studies using patients examining aging-associated
tendon degeneration have not been performed.
A healthy tendon is a fibrous type of tissue with a
highly organized type I collagen-based extracellular matrix, and a
minimal number of cells and neurovascular structures (7). As tendons mature postnatally, tenocytes
slowly proliferate to become largely quiescent, resulting in an
extremely slow tissue turnover rate but an absence of atrophy with
aging (1,2). A previous study suggested that one of
the key characteristics of chronic tendinopathy is the degradation
of the extracellular matrix collagen (8). Other data indicated that a marked loss
of bridging collagen in the extracellular matrix was observed in
repaired tendons in an aged mouse mode (9). These observations suggest that the
decreased proliferation of tenocytes and increased matrix
degeneration lead to impaired tendon healing with aging (9). However, the basic mechanisms underlying
the pathological changes of tendons with aging are not well
understood. The majority of previous studies have investigated the
mechanical features and the healing process of aged tendons; to the
best of our knowledge, only one study has focused on the effect of
aging on the biological features of tendons (10).
In the present study, to evaluate the biological and
pathological changes of tendons with aging, the effect of aging on
the tendon structure, distribution of collagen types I and III and
expression of tendon-associated genes was examined using flexor
tendons in a mouse model.
Materials and methods
Animal model
Female C57BL/6 mice (n=26) were used in the present
study. Mice were obtained from Japan SLC, Inc., (Shizuoka, Japan).
Animals were housed in groups, with up to 5 animals/cage in a
pathogen-free environment at 22±2˚C with 40-60% humidity under a 12
h light-dark cycle with ad libitum access to food and water.
The effect of aging on the structure and gene expression in the
tendons was assessed at 8 weeks (young group) and 78 weeks (aged
group); 5 and 8 mice (young or aged) were for employed for
structural and gene expression analyses, respectively. The present
study was approved by the Animal Care Committee of Juntendo
University, Tokyo, Japan (registration no. 1309; approval no.
300052).
Histological assessment of tendon
structure
Following euthanasia, flexor digitorium longus (FDL)
tendons of the right hind paws (including 5 digital tendons and a
bunched portion of the tendons) were harvested under a light
microscope (Zeiss Axioskop2; magnification, x40) for histological
analysis. Then, tendons were fixed in 4% paraformaldehyde at room
temperature for 72 h, and processed and embedded in paraffin. From
the bunched portion of tendons, 3 µm sagittal sections were
prepared and stained with 0.1% hematoxylin for 5 min and 1% eosin
for 2 min at room temperature (5 sections/mouse). In addition, 3 µm
axial sections were prepared from the bunched portion of tendons,
and stained with 0.1% Picrosirius Red (Waldeck GmbH & Co. KG,
Münster, Germany) for 90 min at room temperature (8
sections/mouse).
Soslowsky score
The Soslowsky score is a histological scoring
system, which is used to investigate tendinopathies and tendon
repair (11). The total score may
range from 0 (normal tendon)-12 (most severe degeneration
detectable). Sections were stained with 0.1% hematoxylin for 5 min
and 1% eosin for 2 min at room temperature to assess cellularity
(score 0-3), fibroblastic changes (score 0-3), and collagen fiber
orientation (score 0-3) and disruption (score 0-3). The analysis
was accomplished using a semiquantitative method by evaluating
scores (0-3) of each component (cellularity, fibroblastic change,
collagen fiber orientation and disruption) (12). The slides were randomly selected and
evaluated by two blinded observers under a light microscope
(magnification, x20). The total score of each mouse (5
sections/mouse) was averaged, and compared between the young and
aged groups (5 mice/group).
Picrosirius red staining for
collagen
Picrosirius red staining is utilized for evaluating
the differential distribution of the structurally distinct collagen
types I and III (13-15).
Picrosirius red-stained sections were illuminated with polarized
light, allowing the visualization of collagen fiber organization;
type I collagen appears red, while type III collagen appears
yellow/green (16). Digital images of
all specimens were captured and analyzed with a custom-built
spectral template (red or yellow/green) using the Nuance
multispectral imaging system (Nuance 3.0.2; PerkinElmer, Inc.,
Waltham, MA, USA). The total number of pixels of red and
yellow/green in the specimen was obtained, and the percentage of
red (type I collagen) or yellow/green (type III collagen) staining
was calculated. This proportion was averaged in each mouse (8
sections/mouse) and compared between young and aged groups (5
mice/group).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
FDL tendons of the right and left hind paws were
harvested from each mice (8 mice/young or aged group) using
microscopy.
Total RNA was isolated from flexor tendons (2
tendons/mouse) with an RNeasy Fibrous Tissue Mini kit (Qiagen,
Inc., Valencia, CA, USA). cDNA was synthesized using qPCR RT Master
Mix (Toyobo Life Science, Osaka, Japan) at 37˚C for 15 min and at
50˚C for 5 min. qPCR was performed using an ABI Prism 7500 sequence
detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) with the SYBR-Green PCR Master Mix (Toyobo
Life Science, Osaka, Japan), according to the manufacturer's
protocol. The detector was programmed with the following PCR
thermocycler conditions: Initial denaturation at 95˚C for 1 min, 40
cycles of 15 sec denaturation at 95˚C and 1 min amplification
(annealing and elongation) at 60˚C. All reactions were performed in
triplicate and normalized to the level of the housekeeping gene
glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The relative
differences in the PCR results were calculated using the
comparative cycle threshold method (17). The following primer sets were used:
Type I collagen forward, 5'-GCTCCTCTTAGGGGC CACT-3'; type I
collagen reverse, 5'-CCACGTCTCACCATTGGGG-3'; type III collagen
forward, 5'-ACGTAGATGAATTGGGATG CAG-3'; type III collagen reverse,
5'-GGGTTGGGGCAGTC TAGTG-3'; tenomodulin forward, 5'-TGTACTGGATCAATC
CCACTCT-3'; tenomodulin reverse, 5'-GCTCATTCTGGTC AATCCCCT-3';
scleraxis BHLH transcription factor forward,
5'-CACCCAGCCCAAACAGATCTG CA-3'; scleraxis BHLH transcription factor
reverse, 5'-AGTGGCATCACCTCTTGG CTGCT-3'; Mohawk homeobox forward,
5'-CACCGTGACA ACCCGTACC-3'; Mohawk homeobox reverse, 5'-GCACTA
GCGTCATCTGCGAG-3'; GAPDH forward, 5'-ATGGCCTT CCGTGTTTCCTAC-3'; and
GAPDH reverse, 5'-TGATGT CATCATACTTGGCAGG-3'.
Statistical analysis
Values are expressed as the means ± standard
deviation. Statistical analysis was performed by comparing values
between the young and aged groups with a Mann-Whitney U test using
GraphPad Prism 7 (GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Histological assessment of tendon
structure
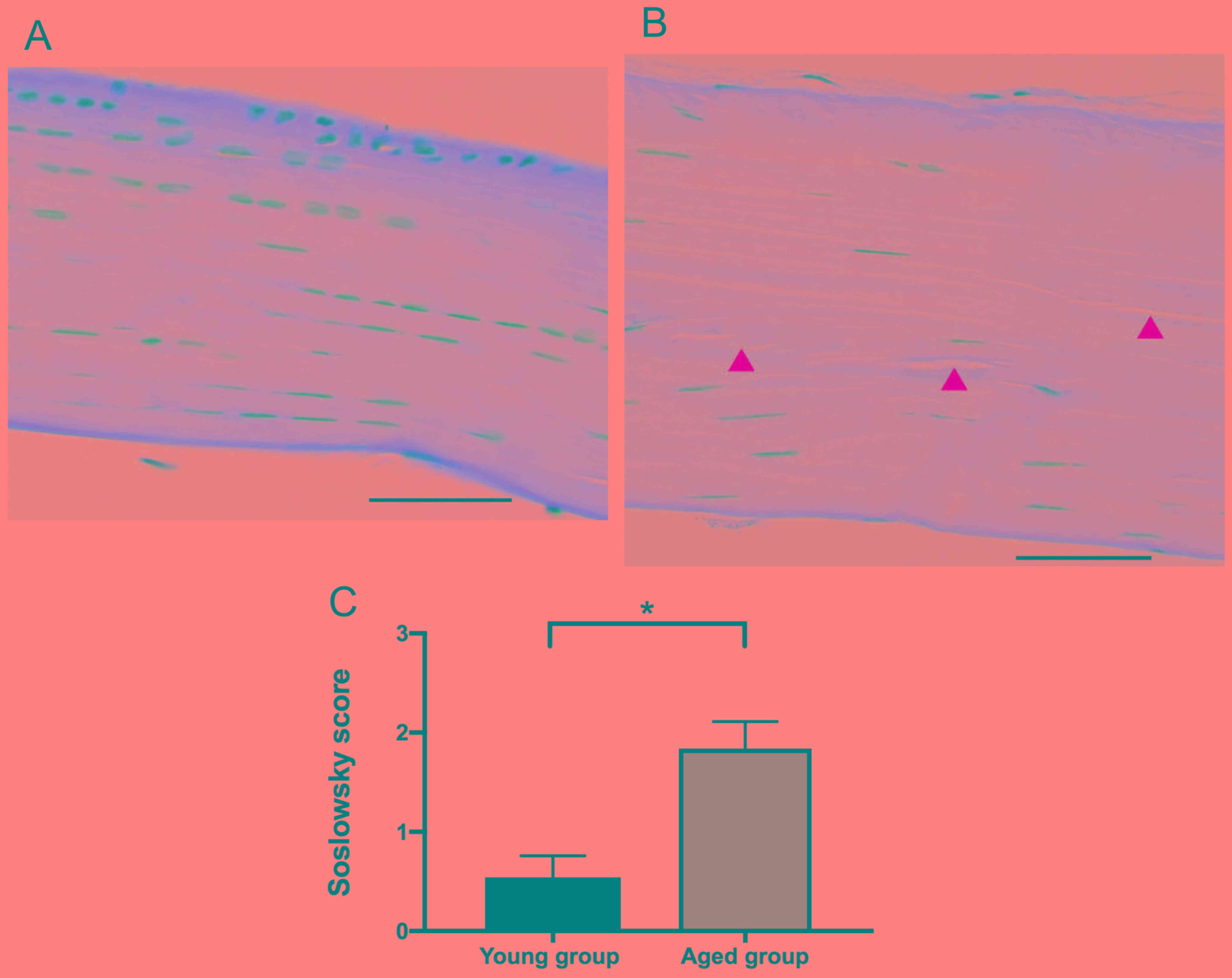
Firstly, the structural changes in the FDL tendons
of the hind paws in the young and aged groups was evaluated by
hematoxylin and eosin staining of tissue sections. Normal collagen
is stained pink-red by hematoxylin and eosin, although the
intensity of the staining is decreased and appears paler with the
degeneration of collagen (18). In
the present study, the collagen fibers in the sections of the young
group were stained pink-red by hematoxylin and eosin (Fig. 1A). However, the staining intensity was
decreased in the aged group (Fig.
1B). Furthermore, the cell number, or cellularity, was observed
to be decreased and the shape of nuclei became elongated,
representing fibroblastic changes, in the aged group compared with
the young group. In addition, the alignment, or orientation, of the
collagen fibers was altered and disrupted in the aged group
(Fig. 1B). Based on the results from
the analysis of cellularity, fibroblastic changes, and collagen
fiber orientation and disruption, the Soslowsky score was
calculated; the score was significantly increased, or worsened, in
the tendons of the aged group compared with that in young group
(P<0.05; Fig. 1C).
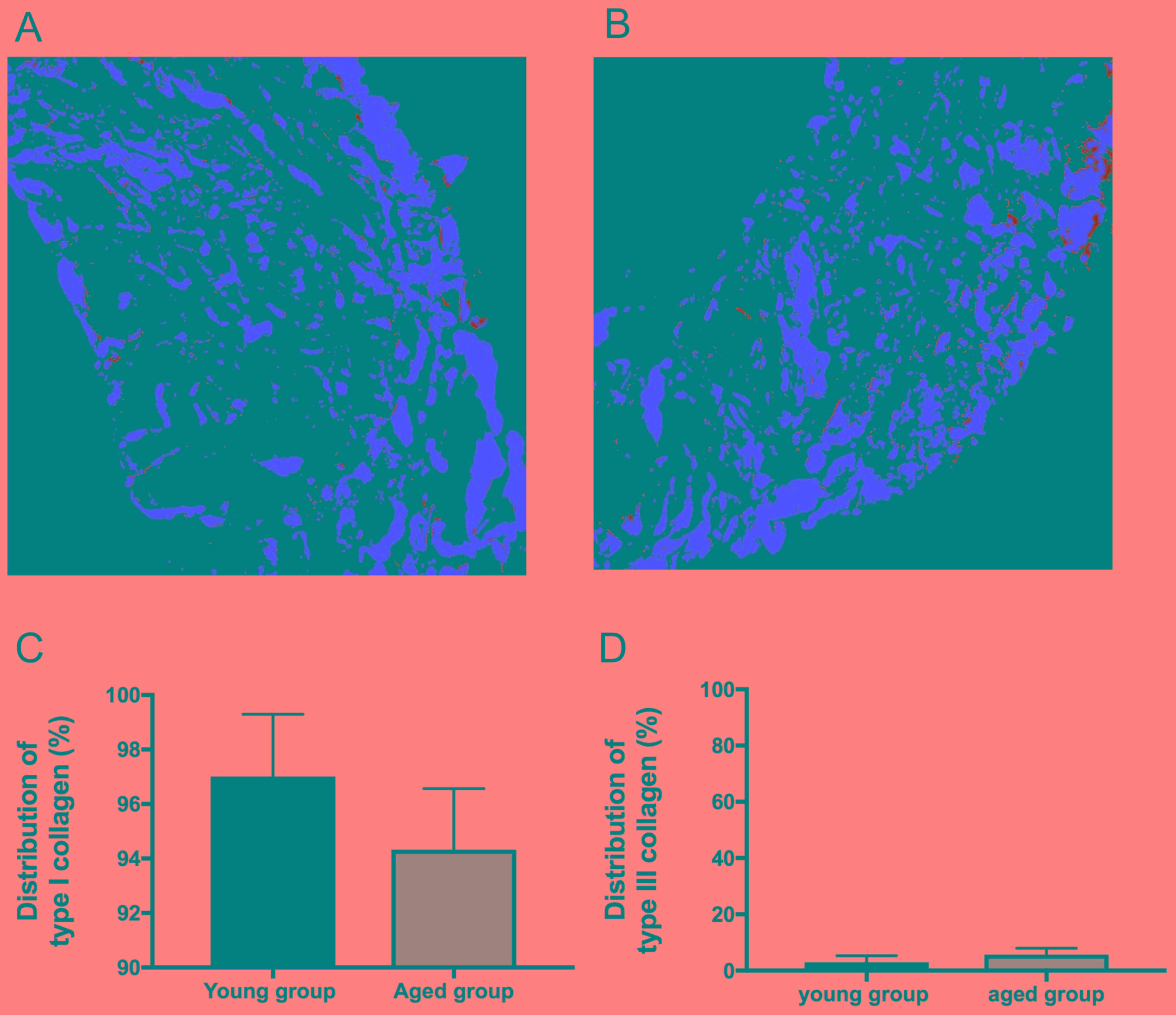
Evaluation of the differential
distribution of collagen types I and III
Next, the differential distribution of collagen
types I and III in the tendons of the young and aged groups was
evaluated. Type I collagen is the predominant collagen in tendons,
comprising 90-95% of the total collagen content, and is localized
in the tendon parenchyma (19,20). By
contrast, type III collagen is the second most abundant type of
collagen in the tendon, comprising up to 5-10% of the collagen
content. In normal tendons, type III collagen is primarily located
in the endotenon, the fine connective tissue between the strands in
a tendon, and the epitenon, which is a sheath around the tendons
(21). Therefore, the present study
assessed the effect of aging on the differential distribution of
collagen type I and III by using picrosirius red staining, which
provides a visualization of type I collagen (red) and type III
collagen (yellow/green) distribution in tissues. In the picrosirius
red-stained sections of the young and aged groups, type I collagen
(red) was localized in the tendon parenchyma, whereas type III
collagen (yellow/green) was localized around the bundle of type I
collagen (red) (Fig. 2A and B). Of
note, relative staining of type I collagen (red) to type III
collagen (yellow/green) was markedly decreased in the aged group
compared with that in the young group (Fig. 2A and B). Furthermore, the total number
of red and yellow/green pixels in the images were obtained and
analyzed with a Nuance multispectral imaging system. As
demonstrated in Fig. 2C and D, the
percentage of type I collagen was decreased (P=0.15) and the
percentage of type III collagen was relatively increased (P=0.15)
in the aged group compared with the young group, although no
statistical significance between the two groups was observed.
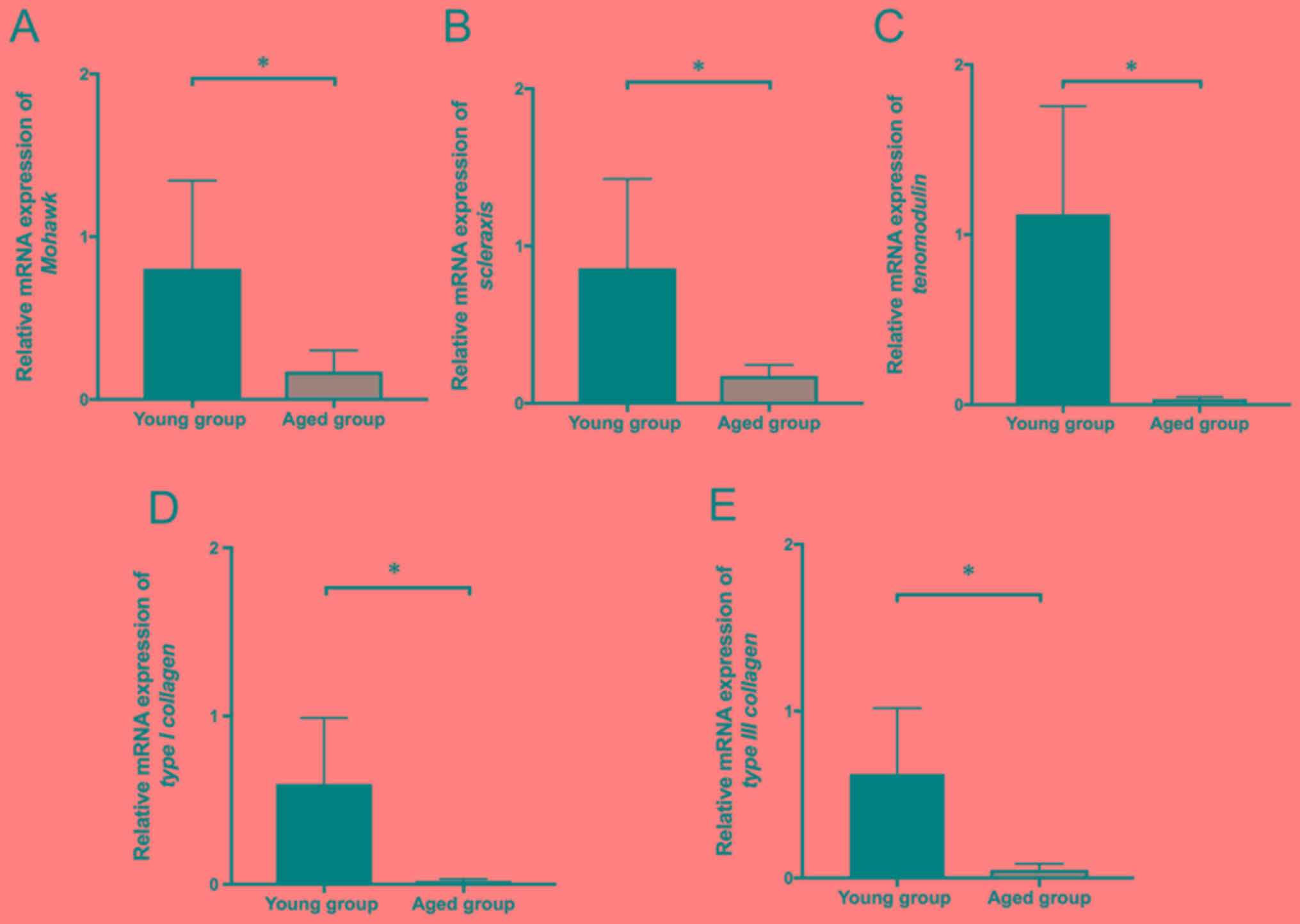
Expression of tendon-associated
genes
Finally, the effect of aging on the mRNA expression
of type I and type III collagen and tendon-associated genes was
evaluated. Mohawk homeobox, a transcription factor, serves a
critical role in tendon differentiation by regulating type I
collagen production in tendon cells (22); scleraxis BHLH transcription factor, a
transcription factor, has been demonstrated to trigger tendon
differentiation in stem cells and improve repair of tendon injury
in animal models (23). Tenomodulin
is a type II transmembrane glycoprotein that is highly expressed in
tendons, and functions as a regulator of tenocyte proliferation and
collagen fibril maturation (24).
Notably, mRNA expression levels of type I and type III collagen and
tenogenic markers (Mohawk homeobox, tenomodulin and scleraxis BHLH
transcription factor) were significantly decreased in the aged
group compared with the young group (P<0.05; Fig. 3).
Discussion
The aim of the present study was to evaluate the
aging-associated changes of tendons using young and aged mice, and
to clarify the basic mechanism for the increased incidence of
tendinopathy associated with aging. Previous studies using a flexor
tendon injury model primarily focused on the morphological changes
at the site of repair and the cellular source contributing to
tendon healing (9,25). However, to the best of our knowledge,
the effects of aging on the histological and biochemical features
of tendons have not been explored thoroughly. In the present study,
histological assessment of tendon structure was performed and the
expression levels of tendon-associated genes in young and aged mice
were examined. The results indicated that the Soslowsky score,
based on the analysis of cellularity, fibroblastic changes, and
collagen fiber orientation and disruption, was significantly
increased, or worsened, in the tendons of the aged group compared
with those in the young group. Furthermore, in the aged group, the
distribution of type I collagen was decreased and the distribution
of type III collagen was relatively increased compared with the
young group. Finally, the mRNA expression levels of collagen (type
I and type III) and tenogenic markers (Mohawk homeobox, tenomodulin
and scleraxis BHLH transcription factor) were significantly
decreased in the aged group compared with the young group.
Therefore, the data from the present study demonstrated that aging
modulates the structure of tendons, distribution of types I and III
collagen and the expression of tendon-associated genes.
It has been demonstrated that the number of
tenocytes decreases during the aging process, and that the
remaining cells exhibit an altered morphology, with rounder cells
in the younger tendons being replaced by a thinner, more elongated
phenotype in aged tenocytes, which is associated with a lower
metabolic activity, including decreased collagen synthesis
(9,26). The decrease in cellularity may be
associated with a decrease in the size of the pool of tendon/stem
progenitor cells (27-29).
Consistent with these data, the present study revealed that the
staining intensity, and potentially the protein content, based on
the result of western blot analysis (as described below), of the
collagen fibers was decreased, the tenocyte number was decreased,
and the shape of nuclei became elongated in the aged group compared
with the young group. Notably, preliminary western blot analysis
experiments in the present study confirmed that the content of type
I collagen, the predominant collagen in tendons, was decreased in
the aged group compared with the young group (data not shown).
However, type III collagen was not detected by western blot
analysis, due to its low expression level. As the total amount of
collagen was observed to be markedly decreased in the tendons of
the aged group by hematoxylin and eosin staining, we hypothesized
that the protein content of type III was also decreased in the aged
tendons, reflecting the decreased mRNA expression of type III
collagen; however, the levels of type III collagen were
demonstrated to be relatively increased from 3.0% in the young
group to 5.6% in the aged group by picrosirius red staining.
Furthermore, it has been suggested that levels of elasticity and
tension decrease in skin when the levels of type III collagen
decrease (30). However, in the
present study, the distribution of type I and type III collagen was
only examined in the tendon, but not in other organs and tissues.
The changes in type I and type III collagen in other organs and
tissues with aging should be evaluated in future studies.
Notably, the histopathological scores between young
and aged tendons were compared using the Soslowsky score system,
which may be used for the evaluation of tendinopathies and tendon
repair, based on the cellularity, fibroblastic changes, and
collagen fiber orientation and disruption. As a result, the score
was significantly increased, or worsened, in the tendons of the
aged group compared with those in the young group. This observation
supports previous data that the cellularity and tendon matrix in
tendons may decrease with age, which results in the increased
incidence of tendinopathy (9).
Furthermore, it has been revealed that mild and
moderate matrix degeneration in tendinopathy characteristically
exhibits a decrease in collagen type I but an increase in collagen
type III (31). Notably, the
picrosirius red staining performed in the present study
demonstrated that in the aged group, the distribution of type I
collagen was decreased but the distribution of type III collagen
was relatively increased compared with the young group. This change
in the distribution of collagen (types I and III) may contribute to
the development of age-associated tendinopathy.
Accumulating evidence has suggested that scleraxis
BHLH transcription factor-expressing tenogenic cells may
participate in the regenerative repair of tendons to promote the
synthesis of collagen fibers with an aligned orientation, which
have improved tensile strength (32).
Tenomodulin is a regulator of tenocyte proliferation and involved
in collagen fibril maturation (24).
In addition, it has been demonstrated clearly that tenomodulin is
necessary for the maintenance and repair of tendons (23). In addition, Mohawk homeobox serves a
critical role in the regulation of type I collagen production in
tendon cells (22). The present study
revealed that the mRNA expression levels of tenogenic markers
Mohawk homeobox, tenomodulin and scleraxis BHLH transcription
factor were significantly decreased in the aged group compared with
those in the young group. Furthermore, mRNA expression of collagen
(types I and III) was significantly decreased in the aged group
compared with the young group. Taken together, these observations
suggest that the synthesis and maturation of collagen and tenocyte
proliferation are suppressed in the flexor tendon with aging, and
these changes are likely to contribute to the degeneration of
tendons in tendinopathy.
Stenosing tenosynovitis is a common tendinopathy in
flexor tendons, and females are 6 times more likely to be affected
compared with males (33,34). In addition, a previous study
demonstrated that estrogen exhibits a protective effect on the
tendon extracellular matrix, in particular collagen synthesis in
females (35). Therefore, in the
present study, to evaluate the aging-associated changes in tendons,
postmenopausal female mice (78 weeks of age) were utilized. It
should be clarified in future studies whether the changes observed
in aged female mice are also present in aged male mice.
Additionally, it has been suggested that treatment with
phytoestrogens and exercise loading are effective for the
maintenance of tendon mass and mechanical properties (35,36).
Therefore, it would be important to determine whether
phytoestrogens and exercise suppress and exhibit protective actions
on the aging-associated decrease in the expression of collagen and
tenogenic markers observed in the present study.
The present study evaluated the changes in tendon
structure and expression of tendon-associated genes accompanied
with aging, using flexor tendons in a mouse model. The results
demonstrated that the structure of the tendons, distribution of
types I and III collagen and the expression of tendon-associated
genes were modulated by aging in the flexor tendon, and these
changes may contribute to the degeneration of tendons in
tendinopathy. We hypothesize that the factors examined in the
present study, including the structure of the tendons, distribution
of types I and III collagen and the expression of tendon-associated
genes, may be utilized in the future for the assessment of the
protective actions of treatment with estrogen-associated substances
(phytoestrogens) and exercise on the tendinopathy, including
tenosynovitis and tendon rupture.
Acknowledgements
The authors would like to thank Dr Takako Ikegami
(Research Support Center, Juntendo University Graduate School of
Medicine, Tokyo, Japan) for their technical expertise in the
RT-qPCR protocols.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YS, KG and KN designed the study; KK, YS, YK, AF, MI
and IN analyzed and interpreted the data regarding histochemical
staining and gene expression; YS and IN were major contributors in
writing the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Care
Committee of Juntendo University (registration no. 1309; approval
no. 300052).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bhatia-Dey N, Kanherkar RR, Stair SE,
Makarev EO and Csoka AB: Cellular Senescence as the Causal Nexus of
Aging. Front Genet. 7(13)2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
López-Otín C, Blasco MA, Partridge L,
Serrano M and Kroemer G: The hallmarks of aging. Cell.
153:1194–1217. 2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Drake MT, Clarke BL and Lewiecki EM: The
Pathophysiology and Treatment of Osteoporosis. Clin Ther.
37:1837–1850. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Carmeli E, Patish H and Coleman R: The
aging hand. J Gerontol A Biol Sci Med Sci. 58:146–152.
2003.PubMed/NCBI
|
|
5
|
Quan T and Fisher GJ: Role of
Age-Associated Alterations of the Dermal Extracellular Matrix
Microenvironment in Human Skin Aging: A Mini-Review. Gerontology.
61:427–434. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Andarawis-Puri N, Flatow EL and Soslowsky
LJ: Tendon basic science: Development, repair, regeneration, and
healing. J Orthop Res. 33:780–784. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Cook JL and Purdam C: Is compressive load
a factor in the development of tendinopathy? Br J Sports Med.
46:163–168. 2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wu YT, Wu PT and Jou IM: Peritendinous
elastase treatment induces tendon degeneration in rats: A potential
model of tendinopathy in vivo. J Orthop Res. 34:471–477.
2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ackerman JE, Bah I, Jonason JH, Buckley MR
and Loiselle AE: Aging does not alter tendon mechanical properties
during homeostasis, but does impair flexor tendon healing. J Orthop
Res. 35:2716–2724. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kostrominova TY and Brooks SV: Age-related
changes in structure and extracellular matrix protein expression
levels in rat tendons. Age (Dordr). 35:2203–2214. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Loppini M, Longo UG, Niccoli G, Khan WS,
Maffulli N and Denaro V: Histopathological scores for
tissue-engineered, repaired and degenerated tendon: A systematic
review of the literature. Curr Stem Cell Res Ther. 10:43–55.
2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Soslowsky LJ, Carpenter JE, DeBano CM,
Banerji I and Moalli MR: Development and use of an animal model for
investigations on rotator cuff disease. J Shoulder Elbow Surg.
5:383–392. 1996.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Borges LF, Gutierrez PS, Marana HR and
Taboga SR: Picrosirius-polarization staining method as an efficient
histopathological tool for collagenolysis detection in vesical
prolapse lesions. Micron. 38:580–583. 2007.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Junqueira LC, Cossermelli W and Brentani
R: Differential staining of collagens type I, II and III by Sirius
Red and polarization microscopy. Arch Histol Jpn. 41:267–274.
1978.PubMed/NCBI
|
|
15
|
Zerbinati N and Calligaro A: Calcium
hydroxylapatite treatment of human skin: Evidence of collagen
turnover through picrosirius red staining and circularly polarized
microscopy. Clin Cosmet Investig Dermatol. 11:29–35.
2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Geary MB, Orner CA, Bawany F, Awad HA,
Hammert WC, O'Keefe RJ and Loiselle AE: Systemic EP4 Inhibition
Increases Adhesion Formation in a Murine Model of Flexor Tendon
Repair. PLoS One. 10(e0136351)2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Δ Δ C(T)) method. Methods. 25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Maffulli N, Barrass V and Ewen SW: Light
microscopic histology of achilles tendon ruptures. A comparison
with unruptured tendons. Am J Sports Med. 28:857–863.
2000.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Thorpe CT, Streeter I, Pinchbeck GL,
Goodship AE, Clegg PD and Birch HL: Aspartic acid racemization and
collagen degradation markers reveal an accumulation of damage in
tendon collagen that is enhanced with aging. J Biol Chem.
285:15674–15681. 2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Wang JH: Mechanobiology of tendon. J
Biomech. 39:1563–1582. 2006.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Duance VC, Restall DJ, Beard H, Bourne FJ
and Bailey AJ: The location of three collagen types in skeletal
muscle. FEBS Lett. 79:248–252. 1977.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Suzuki H, Ito Y, Shinohara M, Yamashita S,
Ichinose S, Kishida A, Oyaizu T, Kayama T, Nakamichi R, Koda N, et
al: Gene targeting of the transcription factor Mohawk in rats
causes heterotopic ossification of Achilles tendon via failed
tenogenesis. Proc Natl Acad Sci USA. 113:7840–7845. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Murchison ND, Price BA, Conner DA, Keene
DR, Olson EN, Tabin CJ and Schweitzer R: Regulation of tendon
differentiation by scleraxis distinguishes force-transmitting
tendons from muscle-anchoring tendons. Development. 134:2697–2708.
2007.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Docheva D, Hunziker EB, Fässler R and
Brandau O: Tenomodulin is necessary for tenocyte proliferation and
tendon maturation. Mol Cell Biol. 25:699–705. 2005.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Oshiro W, Lou J, Xing X, Tu Y and Manske
PR: Flexor tendon healing in the rat: A histologic and gene
expression study. J Hand Surg Am. 28:814–823. 2003.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Kjaer M, Magnusson P, Krogsgaard M, Boysen
Møller J, Olesen J, Heinemeier K, Hansen M, Haraldsson B, Koskinen
S, Esmarck B, et al: Extracellular matrix adaptation of tendon and
skeletal muscle to exercise. J Anat. 208:445–450. 2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Klatte-Schulz F, Pauly S, Scheibel M,
Greiner S, Gerhardt C, Schmidmaier G and Wildemann B: Influence of
age on the cell biological characteristics and the stimulation
potential of male human tenocyte-like cells. Eur Cell Mater.
24:74–89. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Kohler J, Popov C, Klotz B, Alberton P,
Prall WC, Haasters F, Müller-Deubert S, Ebert R, Klein-Hitpass L,
Jakob F, et al: Uncovering the cellular and molecular changes in
tendon stem/progenitor cells attributed to tendon aging and
degeneration. Aging Cell. 12:988–999. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhou Z, Akinbiyi T, Xu L, Ramcharan M,
Leong DJ, Ros SJ, Colvin AC, Schaffler MB, Majeska RJ, Flatow EL,
et al: Tendon-derived stem/progenitor cell aging: Defective
self-renewal and altered fate. Aging Cell. 9:911–915.
2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang C, Rong YH, Ning FG and Zhang GA: The
content and ratio of type I and III collagen in skin differ with
age and injury. Afr J Biotechnol. 10:2524–2529. 2011.
|
|
31
|
Chen J, Wang A, Xu J and Zheng M: In
chronic lateral epicondylitis, apoptosis and autophagic cell death
occur in the extensor carpi radialis brevis tendon. J Shoulder
Elbow Surg. 19:355–362. 2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Tokunaga T, Shukunami C, Okamoto N,
Taniwaki T, Oka K, Sakamoto H, Ide J, Mizuta H and Hiraki Y: FGF-2
Stimulates the Growth of Tenogenic Progenitor Cells to Facilitate
the Generation of Tenomodulin-Positive Tenocytes in a Rat Rotator
Cuff Healing Model. Am J Sports Med. 43:2411–2422. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Adams JE and Habbu R: Tendinopathies of
the Hand and Wrist. J Am Acad Orthop Surg. 23:741–750.
2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Miyamoto H, Miura T, Isayama H, Masuzaki
R, Koike K and Ohe T: Stiffness of the first annular pulley in
normal and trigger fingers. J Hand Surg Am. 36:1486–1491.
2011.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ramos JE, Al-Nakkash L, Peterson A, Gump
BS, Janjulia T, Moore MS, Broderick TL and Carroll CC: The soy
isoflavone genistein inhibits the reduction in Achilles tendon
collagen content induced by ovariectomy in rats. Scand J Med Sci
Sports. 22:e108–e114. 2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Thomopoulos S, Parks WC, Rifkin DB and
Derwin KA: Mechanisms of tendon injury and repair. J Orthop Res.
33:832–839. 2015.PubMed/NCBI View Article : Google Scholar
|