Introduction
Cancer has become a significant disease for humans.
In the USA alone, there were ~1.6 million new cases and 577,190
predicted mortalities in 2012. Among all types of cancer, prostate
cancer is one of the top causes of male cancer fatalities worldwide
(1). It is predicted that 233,000
new cases of prostate cancer will occur in America during 2014
(2).
Several treatments are available for treatment of
prostate cancer, by overcoming the aggressive tumor. These include
surgery, radiation, radioactive implants and hormonal therapy.
However, the treatment often impacts the quality of life due to
side-effects or complications (3).
Thus, numerous investigators have focused on discovering novel
drugs or treatments. Among all the agents tested, natural products
derived from medicinal plants are among the most favorable.
In our previous study,
kaempferol-3-O-rhamnoside, the major compound found in the
ethyl acetate fractions of the Schima wallichii (S.
wallichii) Korth. leaves, was isolated and its properties were
investigated against breast cancer cell lines. The results
indicated that kaempferol-3-O-rhamnoside was favorable for
further exploration of its anticancer therapeutic potential
(4). Therefore, in the present
study the anticancer properties and mechanism of
kaempferol-3-O-rhamnoside were investigated in prostate
cancer cell lines.
Materials and methods
Plant materials
S. wallichii Korth. leaves were collected
from Lembang, West Java, Indonesia. The plant species was
identified at the Department of Biology, Faculty of Mathematics and
Natural Sciences, University of Padjadjaran (West Java,
Indonesia).
Extraction and isolation
The S. wallichii leaves were dried and
extracted with 70% ethanol at room temperature three times for 24 h
each. A concentrated extract was obtained in vacuo at 50°C.
The ethanol extract was partitioned into n-hexane, ethyl
acetate and aqueous phases. Column chromatography on a Wakogel C
200 (Wako Pure Chemical Industries, Ltd., Osaka, Japan) column was
performed to the ethyl acetate fraction, as it was previously
reported as the most active fraction against cancer cell lines,
using a mixture of n-hexane, ethyl acetate and methanol with
increasing polarity. The major compound observed was purified using
silica G 60 with sulfuric acid ethanol (1:9) and was found to be
the most active fraction of S. wallichii, which was
characterized and analyzed as described previously (4). The isolate was, however, identified by
spectroscopic methods (ultraviolet, infrared and nuclear magnetic
resonance) and liquid chromatography mass spectrometry (5).
Cell culture and treatment
The LNCaP human breast cancer cell line was
purchased from Dainippon Pharmaceutical (Tokyo, Japan). The
non-cancerous esophageal cell line (CHEK-1, an immortalized human
esophageal cell line) was provided by Dr H. Matsubara. CHEK-1 was
established by the transduction of the human papillomavirus type 16
E6/E7 into primary cultures of esophageal keratinocytes (6). The cell lines were cultured in
RPMI-1640 medium (Sigma, St. Louis, MO, USA) supplemented with 10%
fetal bovine serum and antibiotics (100 U/ml penicillin and 100
µg/ml streptomycin). For the cell treatments, various
concentrations of kaempferol-3-O-rhamnoside were added to
the cell culture medium for 24 h, at which time the medium was
replaced. The cells were subsequently collected at the indicated
times.
Drug sensitivity assay
A cell proliferation analysis was performed in the
presence of various concentrations of
kaempferol-3-O-rhamnoside using a colorimetric MTT assay, as
described in a previous study (7).
Briefly, the cells were plated in 96-well plates (2×104
in 50 µl/well). Following the initial cell seeding, the indicated
concentrations of extract were applied and incubated for 24 h.
WST-8 assay cell-counting solution (10 µl) (Dojindo Lab., Tokyo,
Japan) was added to each well and incubated at 37°C for 3 h. After
the addition of 1 M HCl (100 µl/well), the cell proliferation rates
were determined by measuring the absorbance at a wavelength of 450
nm with a reference wavelength of 650 nm using a microtiter plate
reader (Becton-Dickinson, Franklin Lakes, NJ, USA). The results
were derived from triplicate experiments.
Cell extraction and western blot
analysis
Protein concentrations were determined using a
bicinchoninic acid protein assay kit (Pierce, Rockford, IL, USA).
Proteins (40 µg) were electrophoresed on 5–20% Tris-Tricine
ReadyGel (Bio-Rad, Tokyo, Japan) and electro-transferred to a
Hybond-enhanced chemiluminescence membrane (Amersham,
Buckinghamshire, UK). Apoptosis-related proteins were analyzed by
immunoblot analysis using caspase-3 (cat no. 9668), caspase-8 (cat
no. 9746), caspase-9 (cat no. 9508) and poly (ADP-ribose)
polymerase (PARP) antibodies (cat no. 9542) at a 1:1,000 dilution
(Cell Signaling Technology, Beverly, MA, USA). β-actin (cat no.
A1978; Sigma) served as the loading control.
Results
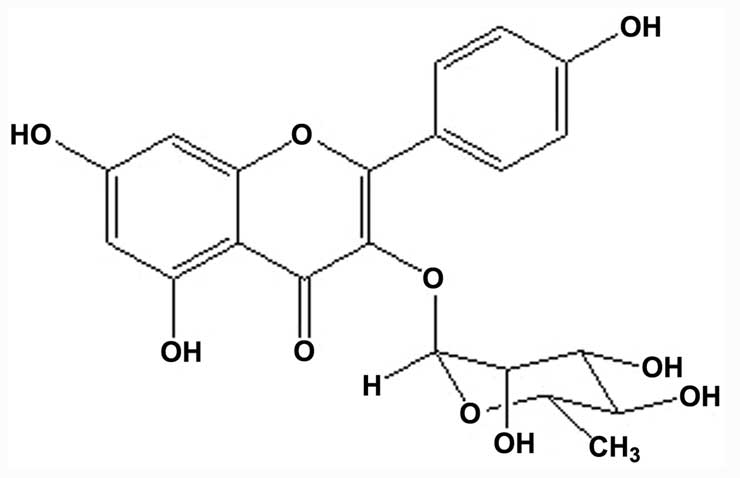
Isolation and identification of
kaempferol-3-O-rhamnoside
Based on a previous study (4), it was found that the major compound of
the ethyl acetate fraction of the S. wallichii extract was
kaempferol-3-O-rhamnoside. The compound was purified,
isolated and identified as kaempferol-3-O-rhamnoside
(C21H20O10 or
3,4′,5,7-tetrahydroxyflavone-3-O-rhamnoside) with a
molecular weight of 432 (Fig.
1).
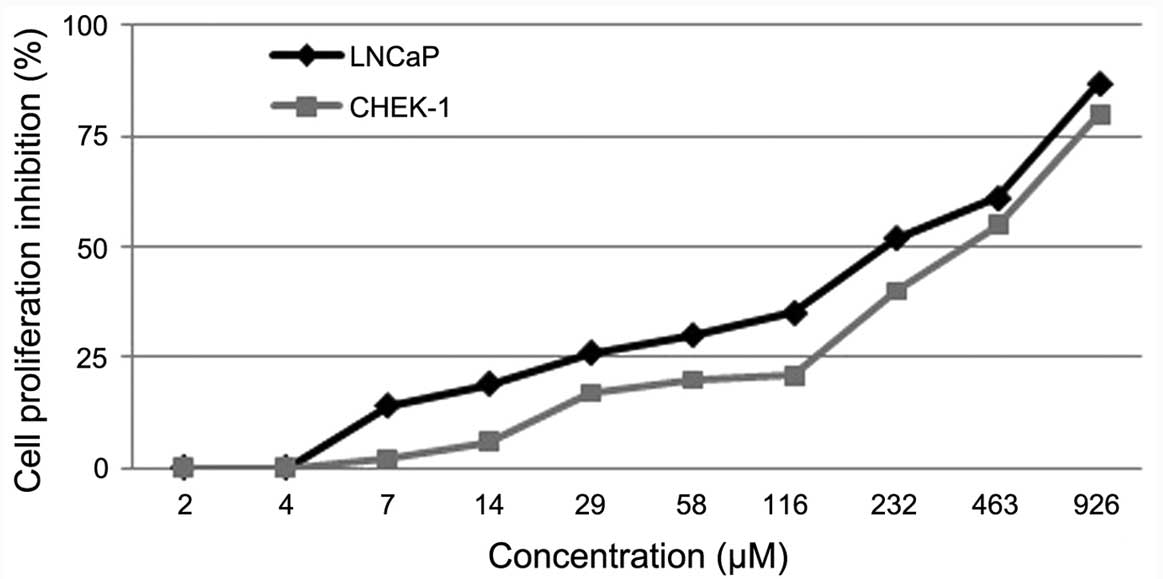
Kaempferol-3-O-rhamnoside inhibits
LNCaP cell proliferation
The effect of kaempferol-3-O-rhamnoside on
the viability of LNCaP and CHEK-1 cells was evaluated. The result
of the MTT assay indicated that the treatment of cancer (LNCaP) and
non-cancer (CHEK-1) cell lines with
kaempferol-3-O-rhamnoside resulted in dose-dependent
inhibition of cell growth (Fig. 2).
Twenty-four hours of treatment with
kaempferol-3-O-rhamnoside inhibited the proliferation of the
LNCaP cells with a half maximal inhibitory concentration
(IC50) value of 218 µM. However, the inhibition of the
CHEK-1 cell proliferation with kaempferol-3-O-rhamnoside
demonstrated an IC50 of 386 µM, which was higher than
that of the LNCaP cells. This may indicate that
kaempferol-3-O-rhamnoside has fewer cytotoxic effects on the
non-cancerous cells.
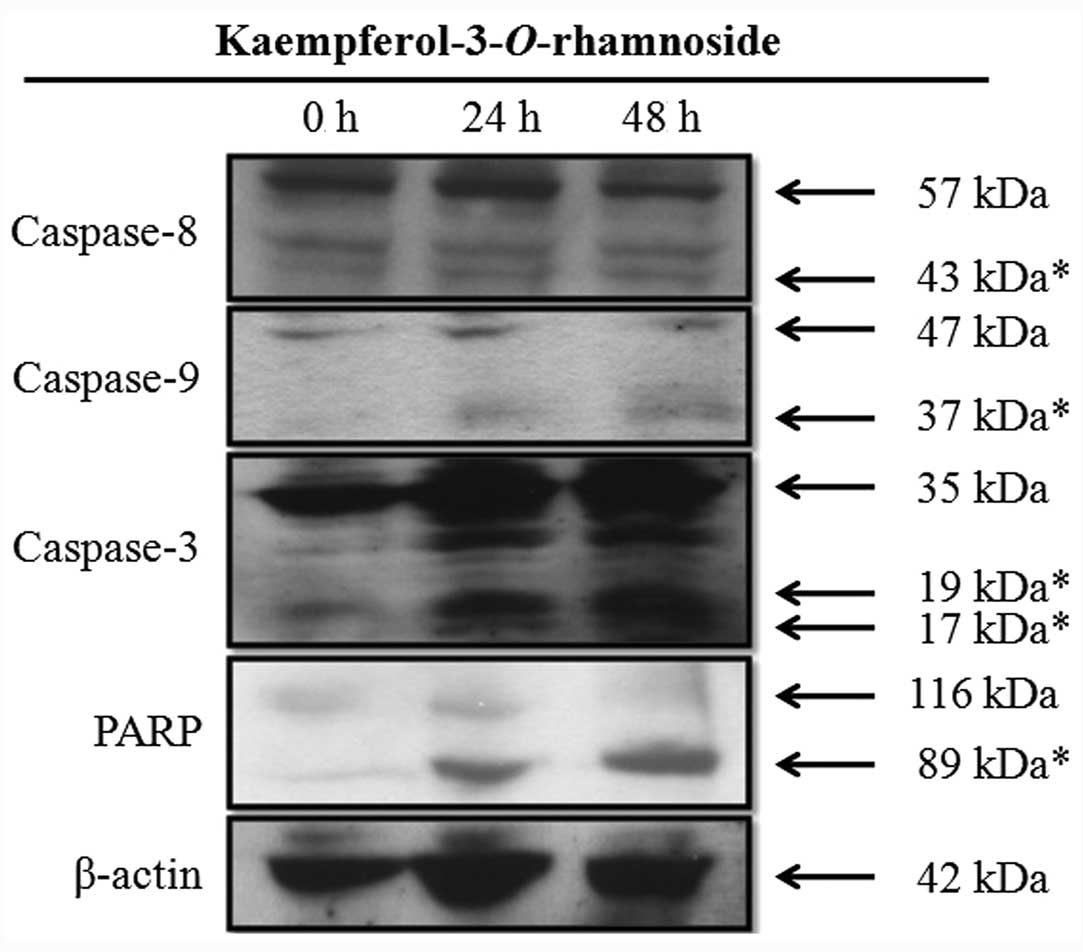
Kaempferol-3-O-rhamnoside induces
caspase cascade protein expression in LNCaP cells
Kaempferol-3-O-rhamnoside treatment of the
LNCaP cells induced the upregulation of caspase-8, caspase-9 and
caspase-3 in a time-dependent manner (Fig. 3). Furthermore, the changes in the
expression levels of PARP, a hallmark biomarker of apoptosis,
suggested that kaempferol-3-O-rhamnoside may induce
apoptosis on LNCaP cells through the activation of
caspase-dependent signaling pathways after 24 h of treatment.
Discussion
S. wallichii plants are found in Asia, from
Indochina to Papua New Guinea. Several of the compounds found in
S. wallichii, including alkaloids and tannins, are
ethnobotanically used for snake and insect bites (8). A previous study reported the
antimicrobial activity of the hydroalcoholic extract from the S.
wallichii bark against Escherichia coli, Pseudomonas
aeruginosa and Shigella species (9). Furthermore, our previous study also
reported the antiplasmodial properties of S. wallichii and
its component, kaempferol-3-O-rhamnoside, against
chloroquine-resistant Plasmodium falciparum (10).
Our previous study also reported the anticancer
properties of kaempferol-3-O-rhamnoside on MCF-7 human
breast cancer cell lines. The results indicated that
kaempferol-3-O-rhamnoside inhibits the proliferation of
MCF-7 cells through the activation of caspase-9 and caspase-3
proteins and that it induced apoptosis (4).
In the present study study,
kaempferol-3-O-rhamnoside inhibited the proliferation of
LNCaP cells in a dose-dependent manner. Notably, the cell
proliferation inhibition by kaempferol-3-O-rhamnoside was
lower on the non-cancerous CHEK-1 cells.
Kaempferol-3-O-rhamnoside may possibly be considered less
harmful to non-cancerous cells. Furthermore, the study also showed
that the anticancer properties of kaempferol-3-O-rhamnoside
occurred via the upregulation of caspase-8, caspase-9, caspase-3
and finally PARP, the marker of apoptosis. Caspases are synthesized
as inactive precursors. There are numerous caspase-activation
pathways that promote apoptosis, such as mitochondrial stress by
apoptosome pathways, death receptor engagement and granzyme
B-induced caspase activation (11).
Caspase-9 is activated by mitochondrial stress. Divergent cellular
stresses, such as DNA damage, heat shock and oxidative stress, may
result in caspase-9 activation through the release of cytochrome
c to the cytoplasm. Efflux of cytochrome c stimulates
several high molecular weight caspase-activating complexes in the
cytoplasm. Cytochrome c is a major component of the
apoptosome and it activates caspase-9. Caspase-8 and caspase-3,
however, are activated by granzyme B. This pathway shows that there
are several cytotoxic granules from cytotoxic T cells, natural
killer cells and granzyme B. Granzyme B will activate caspase-3 and
caspase-8 to facilitate the destruction of the target cells
(11). Therefore, these results
showed that the potential anticancer properties of
kaempferol-3-O-rhamnoside trigger death extrinsically via
caspase-8 activation and intrinsically via caspase-9 activation in
the LNCaP cells.
In conclusion, although further toxicity studies and
activity enhancing structure modifications are required, the
results of the present study indicate the potential application of
kaempferol-3-O-rhamnoside in cancer treatment.
Acknowledgements
The authors would like to thank Dr Yudi
Padmadisastra for his valuable guidance during this research.
References
|
1
|
Siegel R, Naishadam D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
2
|
American Cancer Society, . Cancer Facts
and Figures 2014. American Cancer Society; Atlanta, GA: 2014
|
|
3
|
Stangelberger A, Waldert M and Djavan B:
Prostate cancer in elderly men. Rev Urol. 10:111–119. 2008.
|
|
4
|
Diantini A, Subarnas A, Lestari K, et al:
Kaempferol-3-O-rhamnoside isolated from the leaves of
Schima wallichii Korth. inhibits MCF-7 breast cancer cell
proliferation through activation of the caspase cascade pathway.
Oncol Lett. 3:1069–1072. 2012.
|
|
5
|
Silverstein RM, Webster FX and Kiemle DJ:
Spectrometric Identification of Organic Compounds. 7th. John Wiley
& Sons; New Jersey, NJ: pp. 72–229. 2005
|
|
6
|
Sashiyama H, Shino Y, Sakao S, Shimada H,
Kobayashi S, Ochiai T and Shirasawa H: Alteration of integrin
expression relates to malignant progression of human
papillomavirus-immortalized esophageal keratinocytes. Cancer Lett.
177:21–28. 2002. View Article : Google Scholar
|
|
7
|
Abdulah R, Faried A, Kobayashi K, Yamazaki
C, Suradji EW, Ito K, Suzuki K, Murakami M, Kuwano H and Koyama H:
Selenium enrichment of broccoli sprout extract increases
chemosensitivity and apoptosis of LNCaP prostate cancer cells. BMC
Cancer. 9:4142009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lalfakzuala R, Lalramnghinglova H and
Kayang H: Ethnobotanical usages of plants in western Mizoram.
Indian J Tradit Knowl. 6:486–493. 2007.
|
|
9
|
Dewanjee S, Maiti A, Majumdar R, Majumdar
A and Mandal SC: Evaluation of antimicrobial activity of
hydroalcoholic extract Schima wallichii bark.
Pharmacologyonline. 1:523–528. 2008.
|
|
10
|
Barliana MI, Suradji EW, Abdulah R,
Diantini A, Hatabu T, Shimada JN, Subarnas A and Koyama H:
Antiplasmodial properties of kaempferol-3-O-rhamnoside
isolated from the leaves of Schima wallichii against
chloroquine-resistant Plasmodium falciparum. Biomed Rep.
2:579–583. 2014.
|
|
11
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 37:719–727. 2005. View Article : Google Scholar : PubMed/NCBI
|