Introduction
Parkinson's disease (PD) is a long-term degenerative
disorder of the central nervous system, which is reported to be the
second most common neurodegenerative disorder after Alzheimer's
disease (1). PD is an age-related
disease, affecting 1% of the world population aged over 65
(2). PD is a complex syndrome,
clinically characterized by motor symptoms including resting
tremor, rigidity, bradykinesia and abnormal gait, as well as
non-motor symptoms including autonomic insufficiency, sleep
disorders and cognitive impairment (3).
Cognitive disturbances may occur in the early stages
of PD in patients, in some cases occurring prior to diagnosis, and
gradually increase in prevalence with disease duration(3). It has
been reported that 84% of patients with PD exhibit cognitive
decline and approximately 48% meet the diagnostic criteria for
dementia within a 15-year follow-up period (4); furthermore, among patients with PD, the
risk of developing dementia is almost 6 times higher than in the
general population (5). A key issue
is that there is no defined guidelines to clinically define
cognitive deficit in old age, creating difficultly in determining
whether cognitive deficit is prevalent in PD patients. Thus, our
group aims to identify direct evidence for an association between
clinical progression of PD and cognitive function deficit, to
ultimately provide a basis for therapeutic approaches that may
improve patient quality of life and functional status.
Memory, a complex term for cognitive processes and a
set of mind faculties, involved in encoding, storage and retrieval
of true and false information (6),
may influence quality of life at all stages of life; in turn memory
deficit is a source of burden for government and societies. Certain
data indicate memory deficit and impaired memory processes in
patients with PD (7), the extent of
which often depends on age of onset, disease duration and the
progression of clinical symptoms (8).
More notably, although recognition memory deficits may be observed
in PD with or without dementia; however, a greater recognition
memory deficit has been observed in PD with dementia (9). These previous findings indicate a memory
deficit associated with PD progression, though further studies are
still required to specifically investigate the molecular mechanism
and potential therapies for worsening memory deficits.
Melatonin (MT), produced by several tissues besides
the pineal gland in animals (10),
which is established to be an endogenous regulator of the
sleep/wake cycle, may improve sleep quality and memory decline in
both normal aging and dementia (11).
The impacts of MT on intracellular Ca2+ concentration
and calcium channel activities (12)
have been reported as the potential mechanisms underlying the
therapeutic effects of MT on degenerative disease; however, its
specific mechanisms of action remain unclear. As a natural
supplement, MT is non-toxic to the human body, which would
supplement its efficacy as a potential treatment for patients. In
this regard, larger scale and longer term studies are needed to
evaluate the effect and mechanism of MT on intracellular
Ca2+-related neurotransmission.
The fruit fly, Drosophila melanogaster, is
typically used as a model to study neurological diseases due to the
similarity of the nervous systems between fruit flies and humans
(13,14). In a previous study by our group
(15), the role of human leucine-rich
repeat kinase 2 (hLRRK2), the most common genetic cause of
PD (16), was investigated with
regard to sleep problems, and the therapeutic potential of MT in
hLRRK2-associated sleep problems was investigated in
Drosophila. It was identified that 4 mM MT attenuated
hLRRK2-induced sleep disorders and synaptic dysfunction, but
the effects on memory deficits were undetermined. Therefore, in the
current study, as a follow-on study of our previous work, the aim
was to determine the potential molecular mechanism of MT with
regard to Drosophila memory deficits by using a series of
assays, including behavior tests and whole cell patch-clamp
recordings in vitro, to thus elucidate the potential role of
MT in hLRRK2-induced memory defects.
Materials and methods
Generation of hLRRK2 transgenic
flies
The transgenic flies were constructed to bear
different hLRRK2 constructs, Wild-type (WT) and G2019S
[causing loss of dopamine cells to result in more severe symptoms
of Parkinson's disease (17)], under
the control of a yeast upstream activating sequence (UAS). A green
fluorescent protein (GFP) XbaI-EcoRI fragment, gifted
from Dr Pei Zhong (Department of Neurology, The First Affiliated
Hospital, Sun Yat-sen University, Guangzhou, China), was first
ligated into a pUAST vector to generate a UAS-GFP construct.
Flag-tagged hLRRK2 (amino acids 1,333–1,516; gifted from Dr
Pei Zhong) was then inserted between the KpnI and
BglII sites of the UAS-GFP vector. The constructs were
injected into w1118 embryos (Stock no. 3605,
Bloomington Drosophila Stock Center) to obtain transformant
lines. Two transgenic lines each of UAS-GFP-hLRRK2 and
UAS-hLRRK2-GFP-G2019 were generated. The levels of
hLRRK2 expression were examined by western blot analysis
(15) using anti-Flag staining.
OK107-GAL4 was used to selectively express UAS-GFP-hLRRK2
and UAS-GFP-hLRRK2-G2019S in the mushroom bodies (MBs) of
Drosophila. The phenotypic characterization was conducted on
hLRRK2-WT and hLRRK2-G2019S lines. w1118
served as a negative control. Drosophila were grown on
standard cornmeal medium at 25°C under a 12-h light/dark cycle. For
MT treatment, flies were transferred to regular fly food containing
4 mM MT [groups denoted as (M)]. All control groups (w1118,
OK107, WT-LRRK2 and G1029S) and corresponding results are reported
in our previous work (15).
Electrophysiological recordings from
Kenyon cells (KCs) in isolated whole brains
All brains were obtained from adult flies 1–3 days
in age. The entire brains including optic lobes were removed from
the heads. The dissected brains were then mounted in an RC-26
perfusion chamber (Warner Instruments, LLC, Hamden, CT, USA)
containing the recording solution bubbled with 95% O2
and 5% CO2 (2 ml/min) throughout the experiments with
the ventral brain surface facing upward. The standard external
solution contained (in mM): 101 NaCl, 1 CaCl2, 4
MgCl2, 3 KCl, 5 glucose, 1.25
NaH2PO4 and 20.7 NaHCO3, at pH
7.2, osmolarity 250 Osm. Whole-cell recordings were performed with
pipettes (12–20 MΩ) filled with intracellular solution of the
following composition (in mM): 102 D-gluconic acid, 102 CsOH, 0.085
CaCl2, 1.7 MgCl2, 17 NaCl, 0.94 EGTA and 8.5 HEPES, PH
7.2, 235 Osm. For calcium current, tetrodotoxin (TTX; 1 µM),
tetraethylammonium (TEA; 10 mM) and 4 aminopyridine (4-AP; 1 mM)
were added to the external solution and cesium (Cs+; 102
mM) to the internal solution. Giga-ohm seals were achieved prior to
recording in an on-cell configuration, followed by whole-cell
configuration while in voltage-clamp mode. Recordings were made at
room temperature, and a single KC neuron was examined in each brain
(n=6 per group). Voltage-clamp recordings were performed using
patch-clamp electrodes. In initial experiments, KCs were held at a
potential of −70 mV. The voltage pulse applied consisted of a 100
ms step to 30 mV from −70 mV (in 10 mV intervals). Calcium current
recordings were performed using a BX51WI upright microscope
(Olympus Corporation, Tokyo, Japan). Data were acquired by a
MultiClamp 700B amplifier and an Axon Digidata1440A (Molecular
Devices, LLC, Sunnyvale, CA, USA), and were filtered at 5 kHz using
a built-in filter and digitized at 5 kHz. Amplitude and rise time
of the calcium current were analyzed offline using Clampfit 10.2
(Molecular Devices, LLC).
Behavioral experiment
Classical olfactory conditioning in a T-maze for
Drosophila is a well-documented paradigm that leads to
conditioned odor-avoidance behavior and promotes memory formation
(18). In the T-maze training tube,
an electric flexible stimulus isolator (ISO-Flex; A.M.P.I.,
Jerusalem, Israel) was linked to an electrifiable copper grid. The
training tube was controlled by a programmable pulse stimulator
(Master-8 Channel Programmable Pulse Generator; A.M.P.I.). A vacuum
pump (Tianjin Jinteng Experiment Equipment Co., Ltd., Tianjin,
China) was used to maintain airflow at 800 ml/min in the training
and collection tubes, and deliver odors to each tube.
3-octanol (3-OCT; purity 99%; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and benzaldehyde (BEN; purity 99%;
Sigma-Aldrich; Merck KGaA) were used, since both wild-type and
transgenic strains can learn 3-OCT/BEN combinations (19). The odors were dissolved in heavy
mineral oil (Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
delivered to the training tube or to the two collection tubes with
a bubbler. The concentration of 3-OCT was 1.0×10−3 [in
mineral oil (v/v)] and of BEN was 1.3×10−3 [in mineral
oil (v/v)]. The exposure time to each odor and/or electric shock
(ES) in the training sessions and the exposure time to odors in the
testing sessions were 1 and 2 min respectively, as previously
described (20). The ES mode was a
series of 12×1.5-sec 70–V pulses in 5-sec intervals, yielding a
total stimulus duration of 1 min during a single training
cycle.
Behavioral analyses
A single conditioning performance was assayed using
a standard protocol (18). Two groups
were tested in one complete run to produce one score. The first
group, with approximately 100 flies, was conditioned by exposure to
3-OCT (odor A) paired with an ES (conditioned stimulus,
CS+) for 1 min, followed by exposure to BEN (odor B)
without ES (CS−) for another minute. The flies were then
forced into new feeding tubes and remained in the tube for 60 min.
Then, flies were forced to choose between two odors during a 2 min
testing period. Following the testing period, flies would be
gathered from each T-maze collection tube, anesthetized by
CO2, and counted. A second reciprocal group of flies was
trained with odor B as the CS+ group and odor A as the
CS− group. The memory performance index (PI) =
NCS−-NCS+⁄NCS− +NCS+,
where NCS− represents the number of flies approaching
the CS− odor and NCS+ denotes the number of
flies approaching the CS+ odor. The average of the two
PIs from the reciprocal experiments was taken collectively as one
complete PI. To avoid the possibility of odor bias, the two odors
used for the tests were exchanged in the training sessions of the
reciprocal experiments to generate one complete PI.
Data analysis
Statistical analyses were performed using SPSS 17.0
(SPSS Inc., Chicago, IL, USA). Data were analyzed by Student's
t-tests, one-way analysis of variance (ANOVA) or χ2
tests. When appropriate, ANOVAs were followed by planned
comparisons among the relevant groups with a Tukey's Honest
Significant Difference test. All data were presented as mean ±
standard error of the mean. All P-values were two-tailed and
P<0.05 was considered to indicate statistical significance.
Results
MT rescues long-term memory deficits
in hLRRK2 flies
hLRRK2 was selectively expressed in KCs using
the OK107 promoter to target GAL4 expression in all MBs lobes, and
structurally, MB lobes remained intact in flies expressing either
hLRRK2-WT or hLRRK2-G2019S (15).
In the current study, the olfactory conditioning
paradigm of Tully and Quinn (18) was
used since it has been demonstrated to produce a robust memory for
detailed analysis of a specific memory phase. In this study, to
detect the cognitive impairment caused by hLRRK2 and
investigate whether MT could improve the learning and memory
deficits of flies expressing hLRRK2, the short- and
long-term memory phases were measured (Table I). Our data indicated that
hLRRK2 expressed in MBs could affect the long-term memory PI
of Drosophila but not the short-term memory PI (Table I). The long-term memory PIs of
hLRRK2-WT (0.554±0.036) and hLRRK2-G2019S
(0.502±0.028) flies were significantly reduced compared with that
of the w1118 control group (0.775±0.013; P<0.01 and
P<0.001, respectively). Meanwhile, the long-term memory PI of
the hLRRK2-G2019S(M) group (0.695±0.074; P<0.01) was
significantly increased compared with that of hLRRK2-G2019S
flies without MT, while being restored almost to that of the
control group (P>0.05) following MT treatment; in contrast to
the hLRRK2-G2019S group without MT treatment. The long-term
memory PI of the hLRRK2-WT(M) group (0.593±0.065;) remained
significantly reduced compared with that of the control group
(P<0.05). Collectively, the data suggest that MB expression of
hLRRK2 may induce the long-term but not short-term memory
impairment, and the MT treatment could improve this memory
deficit.
 | Table I.Short- and long-term memory of flies
expressing hLRRK2-WT and hLRRK2-G2019S (with and
without melatonin treatment), driven by OK107-GAL4-mediated
expression of upstream activating sequence transgenes. |
Table I.
Short- and long-term memory of flies
expressing hLRRK2-WT and hLRRK2-G2019S (with and
without melatonin treatment), driven by OK107-GAL4-mediated
expression of upstream activating sequence transgenes.
| Group | Memory |
w1118 | WT | G2019S | WT(M) | G2019S(M) |
|---|
| Performance | Short term | 0.675±0.019 | 0.655±0.021 | 0.064±0.055 | 0.666±0.015 | 0.670±0.019 |
| index | Long-term | 0.775±0.013 |
0.554±0.0356a |
0.502±0.028b |
0.593±0.065c |
0.695±0.074d |
MT improved the calcium channel
activity of KCs in the hLRRK2-G2019S mutants
To test the hypothesis that MT may improve
hLRRK2-induced learning and memory impairment by regulating
calcium channel activity, Ca2+ currents were isolated by
blocking NaC conductances with TTX in the external solution, and KC
channels with a mixture of TEA and 4-AP in the external solution,
and Cs+ in the internal solution. Fig. 1A depicts the current traces recorded
in KCs of the w1118 group and hLRRK2-G2019 and
hLRRK2-G2019 groups treated with MT, along with the
excitation increments; figure 1B
presents a diagram of calcium channel current, indicating the rise
time (ms) and amplitude (pA) of calcium channel activity. For all
groups, the current density (peak amplitude/capacitance, pA/pF) and
rise time of the calcium current were detected (Fig. 1C and D). Current density in the
hLRRK2-WT (2.399±0.208 pA/pF) and hLRRK2-G2019S
(2.565±0.242 pA/pF) groups was significantly increased compared
with in the w1118 group (1.653±0.319 pA/pF; P<0.05) due
to hLRRK2-expression. Current density in the
hLRRK2-WT(M) (1.403±0.138 pA/pF) and hLRRK2-G2019S(M)
(1.620±0.286 pA/pF) groups was decreased compared with in the
hLRRK2-WT and hLRRK2-G2019S groups (P<0.05), thus
the excitation effect of hLRRK2 appeared inhibited by MT.
Additionally, the mean open time of hLRRK2-WT (8.552±0.497
ms) and hLRRK2-G2019S (8.833±0.688 ms) channels was
significantly increased compared with in the w1118 group
(5.624±0.328 ms; P<0.01) due to hLRRK2-expression. The
mean open time of hLRRK2-WT channels with MT treatment
(6.966±0.451 ms) was significantly decreased compared with
hLRRK2-WT channels without MT treatment (8.552±0.497 ms;
P<0.05); and of hLRRK2-G2019S channels with MT treatment
(6.666±0.557 ms) was significantly decreased compared with
hLRRK2-G2019S channels without MT treatment (8.833±0.688 ms;
P<0.05). These data indicated that opening of the calcium
channel may reduce influx of calcium into the cell, resulting in
inhibition of KCs.
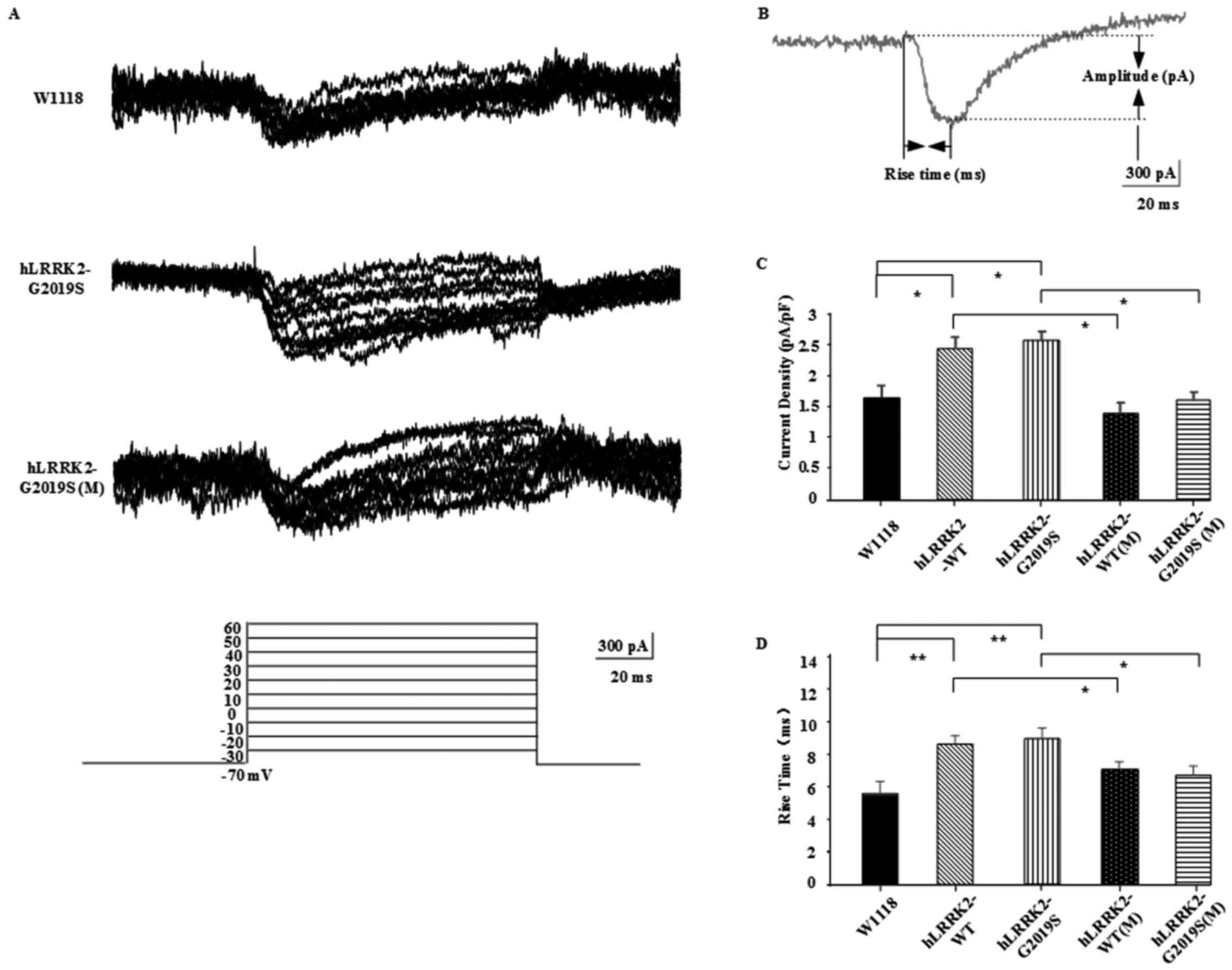 | Figure 1.Calcium channel activity was improved
in hLRRK2-G2019S mutants following treatment with MT. (A)
Representative current traces recorded from KCs of w1118,
hLRRK2-G2019, hLRRK2-G2019(M), hLRRK2-WT,
hLRRK2-WT(M) (data not shown) in whole-cell current-clamp
configuration. The graph (bottom) depicts the series of
depolarizing voltage steps from a holding potential of −70 to 30 mV
elicited by rapid activation of inward calcium currents in KCs. (B)
Diagram of calcium channel current, showing the Rise time (ms),
Delay time (ms) and Amplitude (pA). (C and D) MT decreased the
hLRRK2-induced calcium channel current density and the mean
rise time. Data are presented as the mean ± standard error of the
mean (n=6; three independent experiments performed). Differences
between means were evaluated by one-way analysis of variance
followed by the Tukey's Honest Significant Difference test.
hLRRK2, human leucine-rich repeat kinase 2; WT, wild-type;
G2019S, glycine 2019 serine; KC, Kenyon cell; MT, melatonin; (M),
melatonin-treated. |
Discussion
Increasing clinical and animal studies have revealed
dysregulation of the circadian rhythm in a variety of
neurodegenerative diseases (21). MT,
a hormone that regulates sleep and wakefulness (22), is proposed to have a physiological
role in the aging process (23),
since its secretion decreases with aging (24). It has been reported that there are
greater reductions in MT secretion in populations with dementia
than in those without dementia (25).
More notably, it has been revealed that the level of MT may be
significantly associated with degenerative changes and disease
severity in patients with PD (26).
To detect the association between MT and PD, our
previous research focused on the effect of MT on
hLRRK2-induced sleep problems and hLRRK2-induced
synaptic dysfunction (15). In
addition to sleep problems, it was identified that the expression
of hLRRK2 in MB did not result in any gross morphological
damage; however, could significantly reduce the frequencies of
cholinergic synaptic miniature excitatory postsynaptic current
(mEPSC) and excitatory postsynaptic potential (EPSP), and increase
the synaptic bouton density in transgenic flies. The frequency
changes of cholinergic synaptic mEPSC and EPSP, and the increase of
synoptic bouton, suggested a negative modulatory effect of
hLRRK2 on presynaptic properties; meanwhile, a beneficial
effect of MT on the promotion of presynaptic transmission in KCs
was indicated. Therefore, our previous study suggested a potential
clinical application of MT in patients with PD carrying LRRK2
mutations.
However, to the best of our knowledge, no previous
study has detected the effect of MT on the learning and memory
changes in PD, and its detailed mechanism particularly in relation
to neurotransmitter release and synaptic transmission remains
unclear. Our previous findings appeared associated with learning
and memory abilities (15) and
motivated further study into the effect of MT on learning and
memory. Here, the present research primarily revealed that MT
treatment could also attenuate hLRRK2-induced learning and
memory impairments by regulating the presynaptic membrane calcium
channel activity of KCs. Previous genetic study in
Drosophila has identified distinct temporal memory phases,
including short-term memory, middle-term memory, long-term memory
and so-called anesthesia-resistant memory (27). In the Drosophila brain, the MBs
are key components involved in olfactory learning and memory and
are required during the different phases of memory processing
(28). Here, the present study
detected the memory traces of adult Drosophila, and the data
indicated that the long-term but not the short-term memory of
hLRRK2 flies was significantly decreased, suggesting that
the expression of hLRRK2 could cause learning and memory
deficit; furthermore. G2019S, which is reported to be the most
common mutation in LRRK2-associated PD, leading to the loss of
dopaminergic neurons, retinal degeneration, decreased climbing
activity and early mortality (29,30), may
cause a more severe phenotype than WT (17). As mentioned previously, the present
data demonstrated that the long-term memory of hLRRK2-G2019S
mutants appeared to be impacted to a greater extent than that of
the hLRRK2-WT flies. With the treatment of 4 mM MT, the
long-term memory of Drosophila expressing hLRRK2 was
significantly increased compared with the groups without MT
treatment. Notably, the hLRRK2-G2019S(M) flies exhibited a
somewhat greater beneficial effect than the hLRRK2-WT(M)
flies. In a word, our study was consistent with the hypothesis that
MT should rescue the hLRRK2-induced learning and memory
deficit.
Our previous study identified that MT could
attenuate hLRRK2-induced dysfunction (15), and in the current study, the calcium
channel activity, related to the neurotransmission function of the
Drosophila brain, was detected by patch-clamp recordings. It
was observed that MT inhibited calcium channel activity to reduce
the intracellular Ca2+ levels of KCs in the
hLRRK2 mutants, and decrease both calcium channel current
density and mean rise time compared with in the groups lacking MT
treatment. It is established that KCs are a key cell type
responsible for learning and memory processes (31), and Ca2+ channels
participate in dendritic integration and neurotransmission
(32); therefore, the activation of
presynaptic Ca2+ channels on KCs may influence
postsynaptic output to restore the synaptic transmission and
learning and memory dysfunctions caused by hLRRK2.
In conclusion, the present study demonstrated that
MT could rescue long-term memory deficit by regulating the
presynaptic membrane calcium activity of hLRRK2 mutants,
suggesting that MT may be a potential novel therapy for the
treatment of memory impairment in LRRK2-associated PD. However, the
precise mechanisms underlying the beneficial effects of MT require
further study.
Acknowledgements
The authors are thankful to Dr Pei Zhong at the
Department of Neurology of the First Affiliated Hospital of Sun
Yat-sen University (Guangzhou, China) for providing constructs and
fly stocks.
Funding
The current study was supported by the New Faculty
Program of Pharmacy College of Chonqing Medical University (grant
no. YXY2016×SZ03).
Authors' contributions
JY and HG conceived and designed the study. DR, ZG
and XS performed the experiments. BX revised the manuscript
critically. All authors contributed to the data analysis,
interpretation and approved the final manuscript.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
de Rijk MC, Launer LJ, Berger K, Breteler
MM, Dartigues JF, Baldereschi M, Fratiglioni L, Lobo A,
Martinez-Lage J, Trenkwalder C, et al: Neurologic Diseases in the
Elderly Research Group: Prevalence of Parkinson's disease in
Europe: A collaborative study of population-based cohorts.
Neurology. 54 Suppl 5:S21–S23. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Farlow J, Pankratz ND, Wojcieszek J and
Foroud T: Parkinson Disease OverviewAdam MP, Ardinger HH and Pagon
RA: NCBI Bookshelf. Gene, GeneReviews® (Internet).
Seattle (WA): University of Washington, Seattle; 1993–2018
|
|
3
|
Tolosa E, Gaig C, Santamaria J and Compta
Y: Diagnosis and the premotor phase of Parkinson disease.
Neurology. 72:S12–S20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hely MA, Morris JG, Reid WG and
Trafficante R: Sydney Multicenter Study of Parkinson's disease:
non-L-dopa-responsive problems dominate at 15 years. Mov Disord.
20:190–199. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Aarsland D, Andersen K, Larsen JP, Lolk A,
Nielsen H and Kragh-Sørensen P: Risk of dementia in Parkinson's
disease: A community-based, prospective study. Neurology.
56:730–736. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Melton AW: Implications of short-term
memory for a general theory of memory 1. J Verbal Learn Verbal
Behav. 2:1–21. 1963. View Article : Google Scholar
|
|
7
|
Sagar HJ, Sullivan EV, Gabrieli JDE,
Corkin S and Growdon JH: Temporal ordering and short-term memory
deficits in Parkinson's disease. Brain. 111:525–539. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sullivan EV and Sagar HJ: Double
dissociation of short-term and long-term memory for nonverbal
material in Parkinson's disease and global amnesia. A further
analysis. Brain. 114:893–906. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Whittington CJ, Podd J and Kan MM:
Recognition memory impairment in Parkinson's disease: Power and
meta-analyses. Neuropsychology. 14:233–246. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hardeland R, Pandi-Perumal SR and
Cardinali DP: Melatonin. Int J Biochem Cell Biol. 38:313–316. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Altun A and Ugur-Altun B: Melatonin:
Therapeutic and clinical utilization. Int J Clin Pract. 61:835–845.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pappolla MA, Sos M, Omar RA, Bick RJ,
Hickson-Bick DL, Reiter RJ, Efthimiopoulos S and Robakis NK:
Melatonin prevents death of neuroblastoma cells exposed to the
Alzheimer amyloid peptide. J Neurosci. 17:1683–1690. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mackay TF and Anholt RR: Of flies and man:
Drosophila as a model for human complex traits. Annu Rev Genomics
Hum Genet. 7:339–367. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Koh K, Evans JM, Hendricks JC and Sehgal
A: A Drosophila model for age-associated changes in
sleep:wake cycles. Proc Natl Acad Sci USA. 103:13843–13847. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun X, Ran D, Zhao X, Huang Y, Long S,
Liang F, Guo W, Nucifora FC Jr, Gu H, Lu X, et al: Melatonin
attenuates hLRRK2-induced sleep disturbances and synaptic
dysfunction in a Drosophila model of Parkinson's disease. Mol Med
Rep. 13:3936–3944. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Greggio E and Cookson MR: Leucine-Rich
Repeat Kinase 2 Mutations and Parkinson's Disease: Three Questions.
Asn Neuro. 1:e000022009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Z, Wang X, Yu Y, Li X, Wang T, Jiang
H, Ren Q, Jiao Y, Sawa A, Moran T, et al: A Drosophila model
for LRRK2-linked parkinsonism. Proc Natl Acad Sci USA.
105:2693–2698. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tully T and Quinn WG: Classical
conditioning and retention in normal and mutant Drosophila
melanogaster. J Comp Physiol A Neuroethol Sens Neural Behav
Physiol. 157:263–277. 1985. View Article : Google Scholar
|
|
19
|
Akalal DB, Wilson CF, Zong L, Tanaka NK,
Ito K and Davis RL: Roles for Drosophila mushroom body
neurons in olfactory learning and memory. Learn Mem. 13:659–668.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pinton P, de Virgilio M, Fogarty KE and
Rizzuto R: Behaviour, learning and memoryDrosophila: A Practical
Approach. Connolly JB and Tully T: IRL. Oxford University Press;
Oxford: pp. 265–317. 1998
|
|
21
|
Videnovic A, Lazar AS, Barker RA and
Overeem S: ‘The clocks that time us’ - circadian rhythms in
neurodegenerative disorders. Nat Rev Neurol. 10:683–693. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pandi-Perumal SR, Srinivasan V, Maestroni
GJM, Cardinali DP, Poeggeler B and Hardeland R: Melatonin: Nature's
most versatile biological signal? FEBS J. 273:2813–2838. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pierpaoli W, Dall'Ara A, Pedrinis E and
Regelson W: The pineal control of aging. The effects of melatonin
and pineal grafting on the survival of older mice. Ann N Y Acad
Sci. 621:(1 Physiological). 291–313. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dori D, Casale G, Solerte SB, Fioravanti
M, Migliorati G, Cuzzoni G and Ferrari E:
Chrono-neuroendocrinological aspects of physiological aging and
senile dementia. Chronobiologia. 21:121–126. 1994.PubMed/NCBI
|
|
25
|
Mishima K, Okawa M, Hishikawa Y, Hozumi S,
Hori H and Takahashi K: Morning bright light therapy for sleep and
behavior disorders in elderly patients with dementia. Acta
Psychiatr Scand. 89:1–7. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Breen DP, Nombela C, Vuono R, Jones PS,
Fisher K, Burn DJ, Brooks DJ, Reddy AB, Rowe JB and Barker RA:
Hypothalamic volume loss is associated with reduced melatonin
output in Parkinson's disease. Mov Disord. 31:1062–1066. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tully T, Preat T, Boynton SC and Del
Vecchio M: Genetic dissection of consolidated memory in Drosophila.
Cell. 79:35–47. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
McGuire SE, Le PT and Davis RL: The role
of Drosophila mushroom body signaling in olfactory memory.
Science. 293:1330–1333. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nichols WC, Pankratz N, Hernandez D,
Paisán-Ruíz C, Jain S, Halter CA, Michaels VE, Reed T, Rudolph A,
Shults CW, et al: Parkinson Study Group-PROGENI investigators:
Genetic screening for a single common LRRK2 mutation in familial
Parkinson's disease. Lancet. 365:410–412. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Di Fonzo A, Rohé CF, Ferreira J, Chien HF,
Vacca L, Stocchi F, Guedes L, Fabrizio E, Manfredi M, Vanacore N,
et al: Italian Parkinson Genetics Network: A frequent LRRK2 gene
mutation associated with autosomal dominant Parkinson's disease.
Lancet. 365:412–415. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Akalal DB, Yu D and Davis RL: A
late-phase, long-term memory trace forms in the γ neurons of
Drosophila mushroom bodies after olfactory classical conditioning.
J Neurosci. 30:16699–16708. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guadalupe A, Steffen H, Magdalena S and
Juan B: TRP, TRPL and Cacophony Channels Mediate Ca2+
Influx and Exocytosis in Photoreceptors Axons in Drosophila.
PLoS One. 7:1036. 2012.
|














