Introduction
Psoriasis vulgaris is a common chronic inflammatory
skin disease marked by the hyperproliferation of keratinocytes and
infiltration of inflammatory cells. Although its pathogenesis has
not been not fully elucidated, the condition is generally
recognized as a polygenic disease resulting from a dysregulated
interplay between hereditary and environmental factors, which gives
rise to the hyperproliferative epidermis and altered
differentiation through an immune-mediated common pathway. Immune
dysfunction plays a vital role in the pathogenic progression of
psoriasis vulgaris (1,2). T cells comprise the majority of the
immune response cells in the lesions. A large number of T-helper
(Th) cells can be found infiltrating the inflammatory reaction
area, and one pathway that is important in psoriasis is the role of
the Th17/interleukin-17 (IL-17) dysregulation (3–5). Th17
cells are a T-cell subset distinct from both Th1 and Th2 cells.
IL-17, which is predominantly produced by Th17 cells, has been
implicated in the pathogenesis of psoriasis vulgaris (6–8).
Receptor-interacting protein 4 (RIP4), as a novel
member of the RIP kinase family, activates mitogen-activated
protein kinases (MAPKs) and nuclear factor κ-light-chain-enhancer
of activated B cells (NF-κB) during the proliferation,
differentiation, apoptosis, inflammation and immune response of
keratinocytes (9,10). The present study aims to investigate
the role of RIP4 in psoriasis vulgaris through the use of
immunohistochemistry to examine the RIP4 expression in the
condition, and by detecting the mRNA and protein levels of RIP4 in
a human immortal keratinocyte cell line, HaCaT, following
stimulation of the cells with graded IL-17 concentrations.
Materials and methods
Patients and skin biopsies
Thirty psoriatic biopsies were obtained from 15 male
and 15 female patients (mean age, 27.96±8.64 years; range, 19–52
years) who had been diagnosed with psoriasis vulgaris in Qilu
Hospital (Jinan, China) and who had not received any local or
systemic treatment since the latest attack of the disease.
Non-lesion biopsies, acquired from 30 age- and gender-matched
healthy individuals, served as controls. All participants were
negative for systemic disease and gave their informed consent prior
to biopsy. The study received approval from the Ethics Committee of
Qilu Hospital (no. QL2009019). The specimens were fixed in 10%
neutral formaldehyde immediately subsequent to their
collection.
Immunohistochemistry
Paraffin-embedded 10% neutral formaldehyde-fixed
tissues were sectioned continuously at 3 µm, deparaffinized with
xylene and rehydrated in a graded alcohol series. For antigen
retrieval, the deparaffinized sections in 1 mM EDTA buffer (pH=9.0)
were heated at 120°C for 5 min and cooled slowly to room
temperature. In order to prevent nonspecific background staining,
the slices were blocked with goat serum following the quenching of
endogenous peroxidase activity with 3% H2O2
solution. Polyclonal rabbit anti-human RIP4 antibody (cat. no.
ZA-0502; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), at a
dilution of 1:100, and a ready-to-use anti-Ki-67 monoclonal
antibody (ZSGB-Bio, Beijing, China) were applied to the sections at
37°C for 1 h, prior to the use of the PV-6000 polymer detection
system (ZSGB-Bio) according to the manufacturer's instructions.
3,3′-Diaminobenzidine (ZSGB-Bio) was applied to visualize the
antibody under a microscope, following which the sections were
counterstained with Harris's hematoxylin. The slices were mounted
with neural gum and images were captured using the ToupCam
microscope camera system (Hangzhou ToupTek Photonics Co., Ltd.,
Hangzhou, China).
Assessment of
immunohistochemistry
Cells were considered to be RIP4- and Ki-67-positive
when yellow to brown granules appeared in the cytoplasm or nucleus
of the keratinocytes, respectively. The proliferation index was
calculated as a percentage by dividing the number of Ki-67-positive
cells by the total number of epidermal cells (basal and
suprabasal). Five high-power fields (x400 magnification) were
counted in each sample.
Cell culture
Immortalized human keratinocyte HaCaT cells were
obtained commercially from the American Type Culture Collection
(Manassas, VA, USA) and frozen in our lab. The HaCaT cells were
thawed using a routine method and grown in Dulbecco's modified
Eagle's medium (Gibco-BRL, Grand Island, NY, USA) supplemented with
10% (vol/vol) fetal bovine serum, 100 U/ml penicillin and 100 µg/ml
streptomycin in a humidified atmosphere containing 5%
CO2 at 37°C. For the subcultures, the cells were
disaggregated with 0.25% trypsin/0.02% EDTA (1:1) solution and
split at a ratio of 1:2 every 2 to 3 days, prior to inoculation
into 12-well plates at a concentration of 1×105
cells/ml. HaCaT cells at 60–70% confluence were stimulated with
IL-17 (50, 70 and 90 ng/ml; ProSpec-Tany TechnoGene, Ltd., Rehovot,
Israel) for RNA isolation or western blotting. The control group
cells were processed in parallel using Dulbecco's modified Eagle's
medium without IL-17.
Cell counting kit-8 (CCK-8) assay
The CCK-8 assay (Dojindo Molecular Technologies,
Inc., Kumamoto, Japan) was adopted to evaluate cell viability.
Cells (5×103/ml) in the logarithmic phase were
inoculated in 96-well plates with 100 µl normal growth medium
containing 10% fetal calf serum. After 24 h, the medium was changed
to serum-free medium. When the cell reached ~70% confluence,
various doses of IL-17 (50, 70 and 90 ng/ml) in serum-free medium
were added. A total of 10 µl CCK-8 was added to each well after 12,
24, 36 and 48 h incubation with IL-17, prior to further incubation
for 2 h at 37°C. A microplate reader (Varioskan™ Flash; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) was applied to measure
the optical density (OD) of each well at 450 nm. The cell viability
(percentage of control) is expressed as the percentage of
(ODtest-ODblank)/(ODcontrol-ODblank).
Reverse-transcription
(RT)-semiquantitative polymerase chain reaction (PCR)
Total RNA was isolated from the cells 48 h after
stimulation with IL-17 using RNAiso Plus [Takara Biotechnology
(Dalian) Co., Ltd., Dalian, China] in accordance with the
manufacturer's instructions and quantified spectrophotometrically
at 260 nm. A total of 1 µg RNA was converted in cDNA using oligo
(dT) primers and then amplified with a Power RT kit (BioTeke,
Beijing, China). The PCR was performed in a total reaction volume
of 50 µl: 25 µl 2X PCR Mix (BioTeke), 2.5 µl forward and reverse
primer, 10 µl templates and 10 µl sterile H2O. The
mixture underwent predenaturation for 5 min at 95°C and was then
subjected to 35 cycles of 95°C for 1 min, 60°C for 45 sec and 72°C
for 45 sec, and finally 72°C for 10 min. The specific primer sets
(Table I) were commercially
synthesized by Shanghai Sangon Biotechnology Co., Ltd. (Shanghai,
China). The amplified DNA fragments were analyzed using 2% agarose
gel electrophoresis. The expected fragment length was 384 bp for
RIP4 and 447 bp for β-actin. Ethidium bromide-stained PCR products
were visualized using an ultraviolet transilluminator (Tanon™
2500R; Tanon Science & Technology Co., Ltd., Shanghai, China),
and the gray value of the DNA bands was simultaneously determined
using an automatic image analyzer with the following formula:
Relative gray value = RIP4/β-actin.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Primer sequence |
|---|
| RIP4 | Upstream:
5′-TGTTAGGTGATTTGGGATAGG-3′ |
|
| Downstream:
5′-AAAGGCACAATGAGGCATA-3′ |
| β-actin | Upstream:
5′-CATTTGCTGCATGGGTTA-3′ |
|
| Downstream:
5′-TCCTACGGCTTGGACTTT-3′ |
| RIP4,
receptor-interacting protein 4. |
|
Western blot analysis
HaCaT cells were dissolved in lysis buffer (cell
lysis buffer for western and immunoprecipitation; Beyotime
Institute of Biotechnology, Haimen, China) containing 1 mM
phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology). The mixture was centrifuged for 5 min at 12,000 × g
and the bicinchoninic acid (BCA) assay method (Enhanced BCA Protein
Assay kit; Beyotime Institute of Biotechnology) was utilized to
quantify the total protein in the supernatant. A total of 30 µg
protein from the cell lysate was solubilized in loading buffer and
subjected to sodium dodecyl sulfate-polyacrylamide gel
electrophoresis on a 10% acrylamide gel, prior to being
electrotransferred onto polyvinylidene difluoride (PVDF) membranes.
Following blocking with 5% skimmed milk at room temperature for 1
h, the PVDF membranes were incubated overnight at 4°C with
polyclonal rabbit anti-human RIP4 antibody at a dilution of 1:400
(sc-83320; Beijing Biosynthesis Biotechnology Co., Ltd., Beijing,
China) or with mouse anti-human anti-β-actin antibody (Santa Cruz
Biotechnology, Inc.) at a dilution of 1:1,000. The membranes were
then thoroughly washed with 0.01 M Tris-buffered saline containing
0.1% Tween 20 and incubated with horseradish peroxidase-labeled
goat-anti-rabbit or goat-anti-mouse immunoglobulin G (heavy- and
light-chain) antibody (EarthOx Life Sciences, Millbrae, CA, USA) at
a dilution of 1:1,000 at room temperature for 1 h. A standard
enhanced chemiluminescence (Shanghai Sangon Biotechnology Co.,
Ltd.) reaction was performed according to the manufacturer's
instructions, and densitometric analysis of the band intensity was
conducted using a chemiluminescence imaging analysis system (Tanon
5000; Tanon Science & Technology Co., Ltd.). Relative
expression was measured by dividing the gray value of RIP4 by that
of β-actin. High gray values represented low expression.
Statistical analysis
All statistical analysis was carried out using the
SPSS 17.0 software package (SPSS, Inc., Chicago, IL, USA), and data
are presented as the mean ± standard error of the mean. The
Student's t-test was performed for the comparison of the results,
and the correlation between RIP4 expression and the proliferation
index was examined using Pearson's correlation analysis.
Results
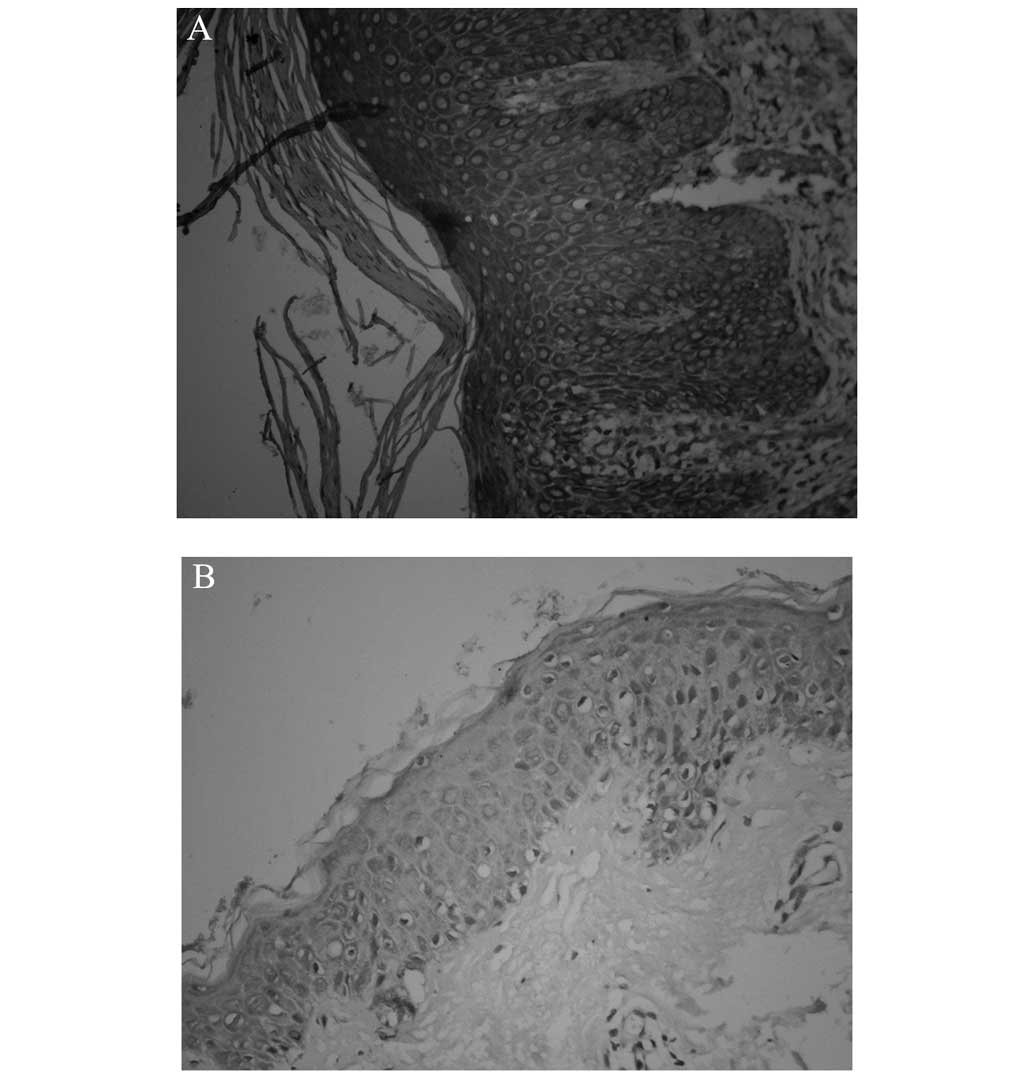
RIP4 is mainly expressed in the
cytosol and nuclei of keratinocytes and shows an evident
upregulation in psoriatic lesions
In healthy skin, RIP4 was found predominantly in the
basal layer of the epidermis; by contrast, the expression in the
psoriatic lesions was concentrated in the basal and spinous layers,
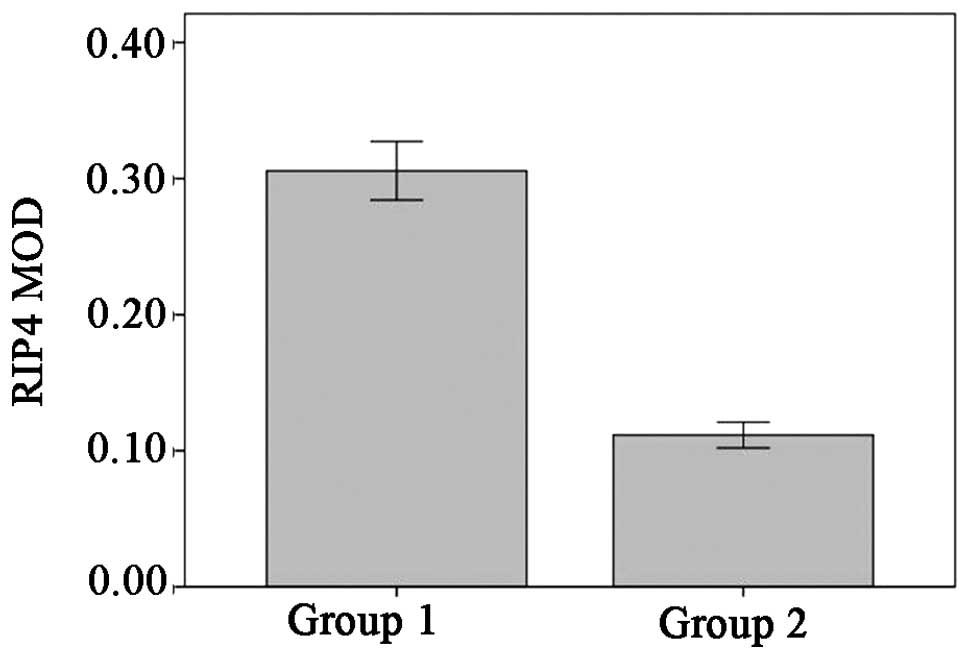
in the cytoplasm and nuclei of the cells. Analysis of the OD showed
the mean OD (MOD) of RIP4 in the healthy controls to be 0.11±0.03,
compared with 0.30±0.05 in the psoriasis vulgaris lesions. This
revealed a statistically significant upregulation in the psoriatic
lesions (t=7.71, P<0.01) (Figs. 1
and 2).
RIP4 expression in psoriatic skin is
associated with keratinocyte proliferation
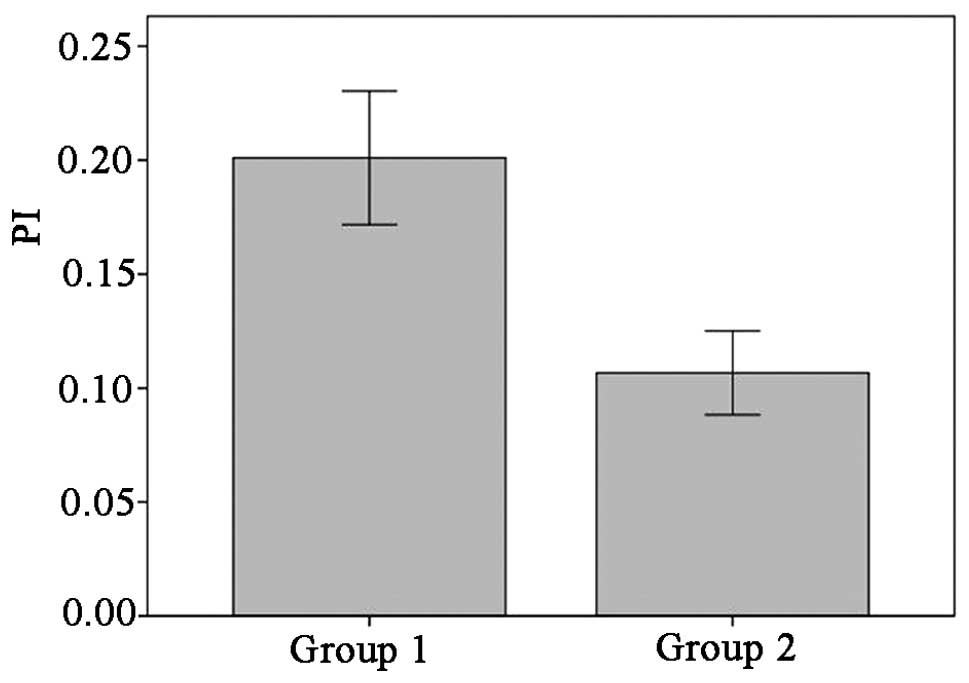
In the healthy control skin, Ki-67-positive cells
only appeared sporadically in the basal layer. By contrast,
Ki-67-positive cells in the psoriatic specimens were distributed in
not only the basal layer but also the middle and lower parts of the
rete ridges of the lesional epidermis. The proliferation index of
the psoriatic lesions was 20.01±7.90%, while that of the healthy
control skin was 10.67±4.92%; this suggested that the Ki-67
expression in the psoriatic lesions was significantly higher than
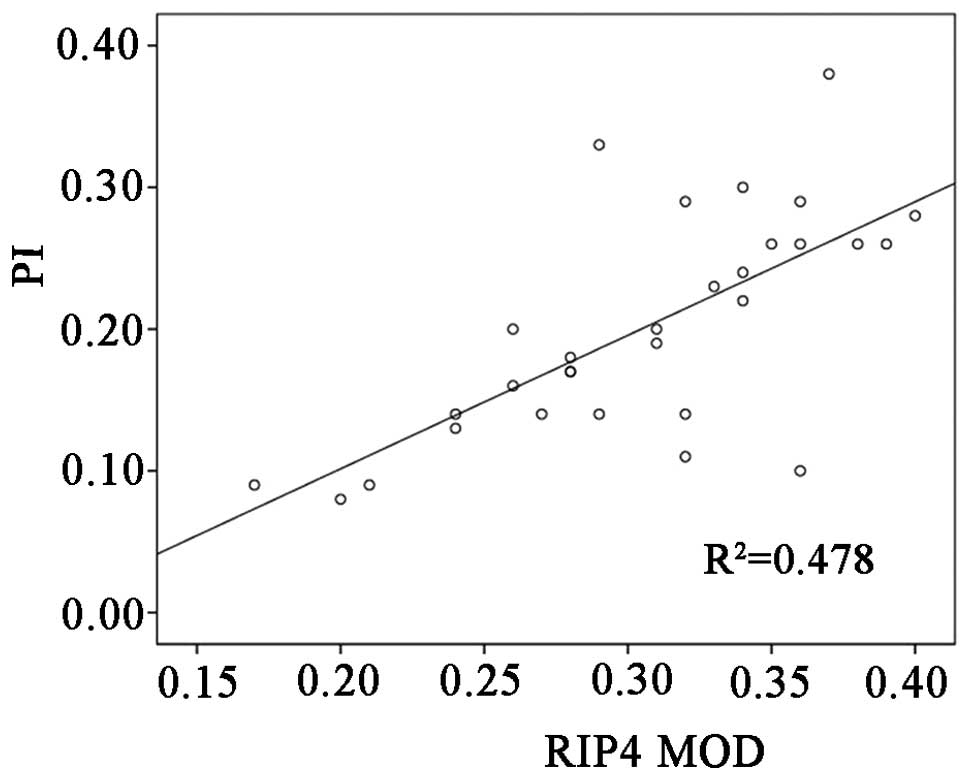
that in the healthy control skin (t=4.535, P<0.05) (Fig 3). In addition, Pearson's correlation
analysis demonstrated a significantly positive correlation between
the RIP4 MOD and the proliferation index, which indicated that a
high level of RIP4 expression led to the active proliferation of
keratinocytes in the epidermis (r=0.692, P<0.01) (Fig. 4).
Promotion of HaCaT cell proliferation
and significant upregulation of RIP4 following IL-17
stimulation
To further investigate the role of RIP4 in
psoriasis, an immortalized line of human epidermal keratinocytes,
HaCaT, which has been extensively adopted as an in vitro
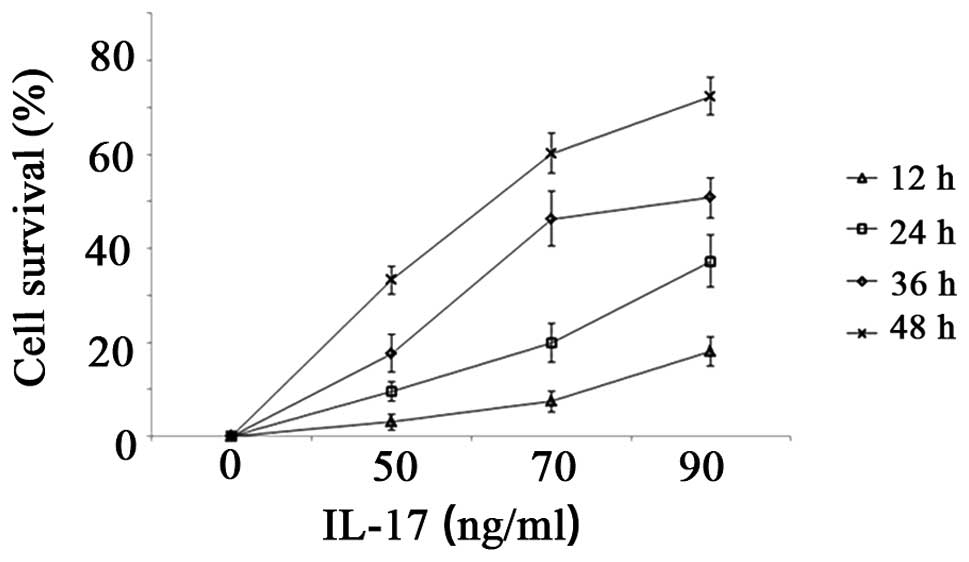
model for studies of psoriasis, was utilized. The CCK-8 assay
performed 12, 24, 36 and 48 h after culture in medium containing
IL-17 revealed that IL-17 at concentrations of between 50 and 90
ng/ml had a marked stimulatory effect on HaCaT cell proliferation,
acting in a dose- and time-dependent manner. Increased
concentrations of IL-17 resulted in higher proliferation rates
(Fig. 5).
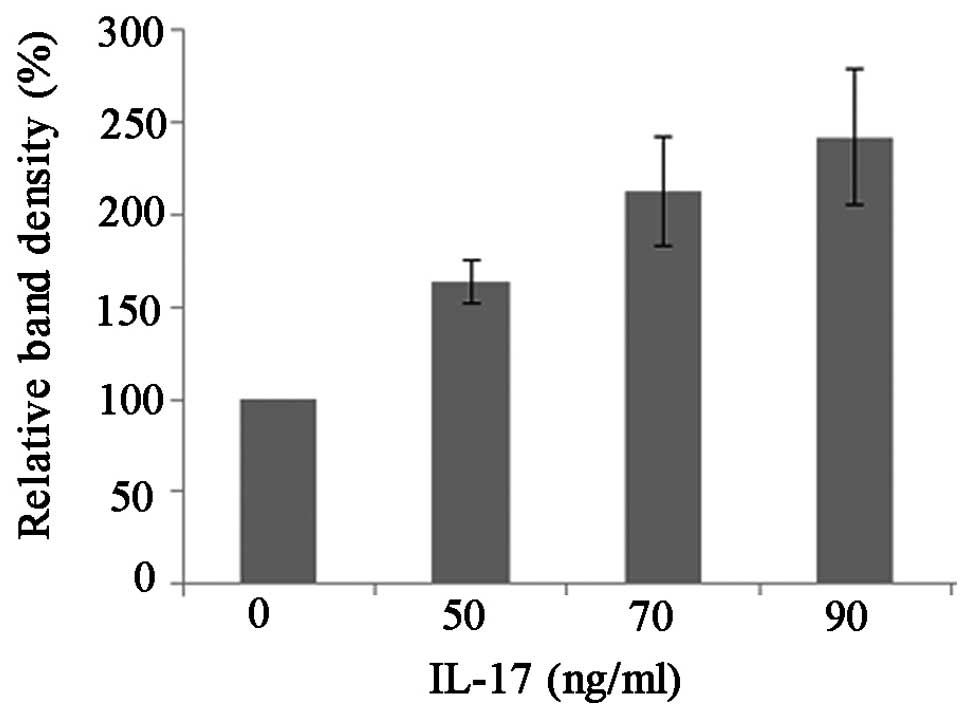
RT-semi-quantitative PCR and western blot analyses
were applied to study the change in RIP4 mRNA and protein
expression, respectively, following exposure to IL-17. The RIP4
mRNA and protein were isolated 48 h after the stimulation of the
cells with 50, 70 and 90 ng/ml IL-17 and subjected to
RT-semi-quantitative PCR or western blot analyses with β-actin as a
housekeeping control. Compared with the control group, it was found
that IL-17 dose-dependently induced a significant upregulation of
RIP4 expression at both the mRNA (t50=5.091,
P50=0.036; t70=7.022, P70=0.020;
t90=18.621, P90=0.003) and protein
(t50=9.076, P50=0.012; t70=6.600,
P70=0.022; t90=6.650, P90=0.022)
levels (Figs. 6–9).
Discussion
Psoriasis vulgaris is a common chronic skin disease
in which a thicker epidermis, excessive keratinocyte proliferation
and parakeratosis, accompanied by extensive inflammatory cell
infiltration, can be observed (9).
It is generally believed that the abnormal keratinocyte
proliferation and differentiation occurs as a result of T cells
recognizing a previously unidentified self-antigen, giving rise to
an immune response (10,11). To date, considerable evidence has
revealed the aggregation of Th17 cells and the upregulation of
IL-17 and IL-17 receptor (IL-17R) in psoriatic lesions (12,13).
Furthermore, the application of agents that act to deplete the
number of T cells in skin lesions and the administration of
monoclonal anti-IL-17 antibodies have been shown to lead to the
clinical and histopathological remission of psoriasis vulgaris,
thereby providing direct evidence that the IL-17-induced changes in
the proliferation and differentiation of keratinocytes play a role
in the pathogenesis of psoriasis vulgaris (12–16).
The RIP kinase family comprises a group of specific
serine/threonine kinases that are crucial mediators of multiple
signal transduction processes, which lead to the activation of
NF-κB and MAPKs (17). RIP4, a novel
member of the RIP kinase family, was initially identified as a
kinase that interacts with an isoform of the protein kinase C (PKC)
family, PKCδ. RIP4 shares a homologous N-terminal kinase domain
with other RIP kinases (~40% identity) and is characterized by a
C-terminal region harboring 11 ankyrin repeats. RIP4 is widespread
in various tissues and cells throughout the body, including the
liver, kidney, brain, heart, lung, skeletal muscles and skin.
Overexpression of RIP4 in 293T cells results in the activation of
NF-κB and the c-Jun N-terminal kinase (JNK) signaling pathway in a
dose-dependent manner (17).
Previous studies (18–20) have demonstrated that RIP4 is required
during epidermal differentiation and cell proliferation in mice,
based on the fact that epidermal dysplasia was observed in
RIP4-deficient mice. The outermost cornified layers were absent in
RIP4−/− skin and were replaced by a thick layer of
flattened, parakeratotic cells with abnormal expression of the
proteins that characteristically mark the specific epidermal
layers. Furthermore, marked hyperplasia of the spinous and granular
layers was observed, indicating that RIP4 is a crucial factor in
the embryonic development of the epidermis (18). The strong downregulation of RIP4
following skin injury has indicated it to be an important regulator
of re-epithelialization following injury (21). In addition, K14-RIP4 mice are
sensitive to cutaneous inflammation with markedly overexpressed
RIP4 (19). Thus, we suggest that
RIP4 could be a key element in the regulation of keratinocyte
proliferation and differentiation and the inflammatory
response.
RIP4 activates NF-κB by inducing inhibitor of κB
phosphorylation, which requires an active kinase domain. Similar to
NF-κB activation, RIP4-mediated JNK signal initiation depends on
the presence of the kinase domain (22,23).
Elevated levels of NF-κB and JNK can be observed in psoriatic
lesions, and drugs aiming to inactivate NF-κB and block the JNK
signaling pathway appear to be of satisfactory efficacy (24–26). It
is speculated that the RIP4-mediated overactivation of NF-κB and
JNK may contribute to such pathological processes as active
keratinocyte proliferation.
IL-17 is one of the most important inflammatory
factors in psoriasis. In the present study, the elevated
proliferation rate of HaCaT cells and the higher levels of RIP4
expression were shown to be enhanced by stimulation with an
increasing concentration of IL-17. Furthermore, the RIP4 protein
level, which was shown to be positively correlated with the
proliferation index, was notably upregulated in the psoriatic
lesions compared with the healthy control skin. It could thus be
inferred that RIP4 is a possible downstream molecule of IL-17.
Interacting with IL-17R on the surface of
keratinocytes, IL-17 produces proinflammatory and
proliferation-regulatory effects by means of activating NF-κB and
the JNK signaling pathway, in which RIP4 is directly involved
(27). In combination, these data
suggest that epidermal hyperproliferation and prosoplasia in
psoriasis vulgaris could be the result of overexpressed RIP4,
induced by the activation of NF-κB and the JNK signaling pathway by
elevated IL-17. In conclusion, IL-17, as an inflammatory factor,
could lead to the upregulation of RIP4 in keratinocytes, and the
IL-17/RIP4 axis may play a role in the epidermal hyperplasia of
psoriasis vulgaris.
Acknowledgements
This study was supported by the 2010 National
Science Foundation (grant no. 81071291).
References
|
1
|
Ariza ME, Williams MV and Wong HK:
Targeting IL-17 in psoriasis: From cutaneous immunobiology to
clinical application. Clin Immunol. 146:131–139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bowcock AM and Krueger JG: Getting under
the skin: The immunogenetics of psoriasis. Nat Rev Immunol.
5:699–711. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gudjonsson JE, Johnston A, Sigmundsdottir
H and Valdimarsson H: Immunopathogenic mechanisms in psoriasis.
Clin Exp Immunol. 135:1–8. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Valdimarsson H, Bake BS, Jónsdótdr I and
Fry L: Psoriasis: A disease of abnormal Keratinocyte proliferation
induced by T lymphocytes. Immunol Today. 7:256–259. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ghoreschi K, Weigert C and Röcken M:
Immunopathogenesis and role of T cells in psoriasis. Clin Dermatol.
25:574–580. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Di Cesare A, Di Meglio P and Nestle FO:
The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J
Invest Dermatol. 129:1339–1350. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Blauvelt A: T-helper 17 cells in psoriatic
plaques and additional genetic links between IL-23 and psoriasis. J
Invest Dermatol. 128:1064–1067. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chiu HY, Cheng YP and Tsai TF: T helper
type 17 in psoriasis: From basic immunology to clinical practice.
Dermatol Sin. 30:136–141. 2012. View Article : Google Scholar
|
|
9
|
Meylan E and Tschopp J: The RIP kinases:
Crucial integrators of cellular stress. Trends Biochem Sci.
30:151–159. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McKay IA and Leigh IM: Altered
keratinocyte growth and differentiation in psoriasis. Clin
Dermatol. 13:105–114. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tschachler E: Psoriasis: The epidermal
component. Clin Dermatol. 25:589–595. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lowes MA, Kikuchi T, Fuentes-Duculan J, et
al: Psoriasis vulgaris lesions contain discrete populations of Th1
and Th17 T cells. J Invest Dermatol. 128:1207–1211. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L, Yang XQ, Cheng J, et al:
Increased Th17 cells are accompanied by FoxP3(+) Treg cell
accumulation and correlated with psoriasis disease severity. Clin
Immunol. 135:108–117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krueger JG, Fretzin S, Suárez-Fariñas M,
et al: IL-17A is essential for cell activation and inflammatory
gene circuits in subjects with psoriasis. J Allergy Clin Immunol.
130:145–154. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leonardi C, Matheson R, Zachariae C, et
al: Anti-interleukin-17 monoclonal antibody ixekizumab in chronic
plaque psoriasis. New Engl J Med. 366:1190–1199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Papp KA, Leonardi C, Menter A, et al:
Brodalumab, an anti-interleukin-17-receptor antibody for psoriasis.
New Engl J Med. 366:1181–1189. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meylan E, Martinon F, Thome M, et al: RIP4
(DIK/PKK), a novel member of the RIP kinase family, activates
NF-kappa B and is processed during apoptosis. EMBO Rep.
3:1201–1208. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Holland PM, Willis CR, Kanaly S, et al:
RIP4 is an ankyrin repeat-containing kinase essential for
keratinocyte differentiation. Curr Biol. 12:1424–1428. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rountree RB, Willis CR, Dinh H, et al:
RIP4 regulates epidermal differentiation and cutaneous
inflammation. J Invest Dermatol. 130:102–112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Adams S and Munz B: RIP4 is a target of
multiple signal transduction pathways in keratinocytes:
Implications for epidermal differentiation and cutaneous wound
repair. Exp Cell Res. 316:126–137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adams S, Pankow S, Werner S and Munz B:
Regulation of NF-kappaB activity and keratinocyte differentiation
by the RIP4 protein: Implications for cutaneous wound repair. J
Invest Dermatol. 127:538–544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bell S, Degitz K, Quirling M, et al:
Involvement of NF-kappaB signalling in skin physiology and disease.
Cell Signal. 15:1–7. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lizzul PF, Aphale A, Malaviya R, et al:
Differential expression of phosphorylated NF-kappaB/RelA in normal
and psoriatic epidermis and downregulation of NF-kappaB in response
to treatment with etanercept. J Invest Dermatol. 124:1275–1283.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gazel A, Banno T, Walsh R and Blumenberg
M: Inhibition of JNK promotes differentiation of epidermal
keratinocytes. J Biol Chem. 281:20530–20541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim HR, Cho ML, Kim KW, et al:
Up-regulation of IL-23p19 expression in rheumatoid arthritis
synovial fibroblasts by IL-17 through PI3-kinase-, NF-kappaB- and
p38 MAPK-dependent signalling pathways. Rheumatology (Oxford).
46:57–64. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Awane M, Andres PG, Li DJ and Reinecker
HC: NF-kappa B-inducing kinase is a common mediator of IL-17-,
TNF-alpha-, and IL-1 beta-induced chemokine promoter activation in
intestinal epithelial cells. J Immunol. 162:5337–5344.
1999.PubMed/NCBI
|
|
27
|
Dong C: Diversification of T-helper-cell
lineages: Finding the family root of IL-17-producing cells. Nat Rev
Immunol. 6:329–333. 2006. View
Article : Google Scholar : PubMed/NCBI
|