Introduction
Rheumatoid arthritis (RA) is an inflammatory disease
characterized by chronic inflammation of the synovial joints and
destruction of cartilage and bone that affects ~1% of the
population worldwide (1). Despite
the advent of anticytokine therapies that ameliorate the
inflammatory manifestations of RA, RA remains incurable (2). Proliferation of synovial cells and
infiltration of activated immunoinflammatory cells result in the
progressive destruction of cartilage and bone (3). Fibroblast-like synoviocytes (FLSs) have
a key role in this process through producing cytokines that
perpetuate autoimmune inflammation and proteases that contribute to
cartilage destruction (4). Synovial
hyperplasia in RA is also considered to impair apoptosis of FLSs
(5,6). Proliferation of FLSs in RA occurs as a
result of the imbalance between cell proliferation, survival and
death. The synovial environment in RA is beneficial to FLS survival
and inhibits FLS apoptosis, and is, thus, involved in preventing
their elimination. Novel therapeutic strategies targeting FLSs
should aim to be effective in inflammatory arthritis without
suppressing systemic immunity.
Interleukin-6 (IL-6) is a pleiotropic cytokine that
has multiple biological functions, including involvement in the
development of the nervous and hematopoietic systems, and
acute-phase, inflammatory and immune responses (7). As a proinflammatory cytokine, IL-6 is
produced by a variety of cell types in the inflamed RA synovial
microenvironment, including macrophages, FLSs and chondrocytes
(8). Previous reports have indicated
that synovial fluid levels of IL-6 are increased in patients with
RA and may be associated with progressive joint damage (9,10).
Anti-IL-6 receptor antibody treatment of patients with RA also
revealed a clinically significant therapeutic effect (11,12). In
addition, IL-6 was observed to contribute to various models of
antigen-induced RA, including collagen-induced and adjuvant-induced
arthritis (13,14). gp130 is a receptor subunit and signal
transducer for the cytokines of the IL-6 family, and is involved in
the Janus-activated kinase (JAK)/signal transducer and activator of
transcription 3 (STAT3) signaling pathway. STAT3 has an important
role in cell survival, growth and differentiation, and is also
associated with osteoclastogenesis (15). Hence, modulation of the activation of
the IL-6/gp130/STAT3 signaling pathway is likely to be a potent
therapeutic strategy in the treatment of RA. Nuclear factor-κB
(NF-κB) is also involved in inflammation, cell survival,
proliferation and differentiation. NF-κB regulates the expression
levels of several proinflammatory gene clusters, including
cytokines, chemokines, adhesion molecules and nitric oxide
synthases. In addition, activation of NF-κB is associated with
chronic inflammatory disorders in RA. Furthermore, animal models of
RA, including collagen- and adjuvant-induced arthritis rat models,
indicated its role in synovial inflammation (16,17).
Thus, the NF-κB and JAK/STAT signaling pathways appear to be
involved in the inflammatory proliferation of FLSs induced by
inflammatory cytokines.
Chlorogenic acid (CGA) is one of the most abundant
polyphenols and exists widely in medicinal herbs. Several studies
have shown that CGA exhibits a wide range of biological activities,
including potent immunoprotective, anti-inflammatory,
anti-bacterial and anti-oxidant activities (18–20).
However, there are no reports regarding the inhibitory effects of
CGA on the proliferation of FLSs through inducing apoptosis in RA,
or the associated molecular mechanism.
In the present study, a rat FLS cell line (RSC-364)
stimulated by IL-6 was used to observe the ability of CGA to
suppress the proliferation of FLSs induced by proinflammatory
cytokines by triggering apoptosis. The modulated function of the
aforementioned complex on the activation of JAK/STAT and NF-κB
signaling pathways were also detected. Therefore, the present study
aimed to investigate the effect and mechanism of CGA on inhibiting
IL-6-mediated proliferation of FLSs through inducing apoptosis, and
investigate the potent inhibitory therapeutic function of CGA on
FLS proliferation in inflammatory hyperplasia of the synovium in
RA.
Materials and methods
Preparation of IL-6 and CGA
The purities of CGA (National Institutes for Food
and Drug Control, Beijing, China) were detected using an API 3200
LC/MS/MS System (AB SCIEX, Ontario, Canada). A total of 1 mol/l CGA
was dissolved in dimethylsulfoxide (DMSO; Sigma-Aldrich, St. Louis,
MO, USA) and 100 mg/l IL-6 was dissolved in Dulbecco's modified
Eagle's medium (DMEM; both Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The solutions were diluted to final
concentrations of 0, 5, 10, 25, 50 and 100 µmol/l CGA, and 0, 0.1,
1, 2.5, 5 and 10 µg/l IL-6.
Cell culture
The rat FLS cell line, RSC-364 (21,22),
used in the present study was a generous gift from Dr Jingxiang
Huang (Chinese People's Liberation Army General Hospital, Bejing,
China). Cells were cultured in DMEM containing 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) supplemented
with 100 U/ml penicillin and 100 µg/ml streptomycin (Sigma-Aldrich)
at 37°C in a humidified atmosphere of 5% CO2 in air.
When cells reached 90% confluency, they were harvested with
trypsin/EDTA (both Thermo Fisher Scientific, Inc.) and subcultured
at a split ratio of 1:3 into new flasks.
Cell viability assay
Cell viability assays were detected using the
3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT)
method. Briefly, RSC-364 cells were seeded into a 96-well plate at
a density of 4,000 cells/well. Following treatment with various
concentrations of CGA (5, 10, 25, 50 and 100 µmol/l) and IL-6 (0.1,
1, 2.5, 5 and 10 µg/l) for 24 h at 37°C, cells were added to wells
with 20 µl MTT (5 mg/ml) per well and incubated for an additional 4
h. Subsequently, after discarding the supernatant, cells were lysed
in 100 µl DMSO and absorbance was detected at 492 nm using a
Varioskan Flash microplate reader (Thermo Fisher Scientific,
Inc.).
Flow cytometric analysis
RSC-364 cells were plated at a density of
1×105 cells in 25 cm2 flasks and incubated
for 15–17 h in a tissue culture incubator at 37°C with an
atmosphere of 5% CO2. The media was discarded and cells
were washed twice with phosphate-buffered saline. Cells were
incubated for 24 h in DMEM containing 3% FBS with 2.5 µg/l IL-6
alone, or 2.5 µg/l IL-6 in combination with 25 µmol/l CGA. The
final concentrations of IL-6 and CGA were determined by the results
of the cell viability assay. RSC-364 cells were stained with
fluorescein isothiocyanate (FITC)-conjugated Annexin V and
propidium iodide (PI). Cells were trypsinized and collected by
centrifugation for 5 min at 800 × g for the detection of apoptotic
cells using an Annexin V-FITC Apoptosis Detection kit (eBioscience,
Inc., San Diego, CA, USA). Briefly, RSC-364 cells were washed twice
with cold PBS at 4°C and resuspended in 500 µl binding buffer [10
mmol/l HEPES-NaOH (pH 7.4), 140 mmol/l NaCl, 2.5 mmol/l
CaCl2] at a concentration of 1×106 cells/ml.
After the addition of 5 µl Annexin V-FITC solution and PI (1
µg/ml), cells were incubated for 15 min at room temperature and
then analyzed by flow cytometry (FC 500; Beckman Coulter, Inc.,
Fullerton, CA, USA). All experiments were performed in triplicate
with three replicates each.
Western blot analysis
Cells were harvested and stimulated, as described in
the previous paragraph. Following treatment, whole cell lysates
from 1×106 RSC-364 cells were generated using a Total
Protein Extraction kit (Thermo Fisher Scientific, Inc.)
supplemented with 1X complete protease inhibitor cocktail (Roche
Diagnostics GmbH, Mannheim, Germany). The protein concentration was
determined using a Pierce BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.). Equal quantities of protein samples (40 µg) were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (Sigma-Aldrich). Electrophoresis was initially
performed at 40 V constant voltage. Once the dye had passed through
the stacking gel, the voltage was increased to 80 V until the dye
ran off the bottom of the gel (<2 h). The protein samples were
subsequently transferred to nitrocellulose membranes (EMD
Millipore, Billerica, MA, USA) and the membranes were blocked with
5% skimmed milk powder (Applygen Technologies Inc., Beijing, China)
and incubated with primary antibodies at 4°C for 12 h. Antibodies
involved in i) JAK/STAT signaling, including gp130 (1:1,000; 3732),
JAK1 (1:1,000; 3344), phosphorylated (p)-STAT3 (1:1,000; 4093) and
p-inhibitor of κB kinase subunit α/β (IKKα/β; 1:1,000; 2697)
antibodies; and ii) NF-κB signaling, including NF-κB p50 antibody
(1:1,000; 4717), were used. Membranes were incubated with
glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:1,000; 2118)
antibody as the loading control. All primary rabbit monoclonal
antibodies were purchased from Cell Signaling Technology, Inc.
(Boston, MA, USA). Then, the membranes were incubated with the
horseradish peroxidase-conjugated goat anti-rabbit IgG secondary
antibody (1:5,000; BA1055; Boster Biological Technology, Inc.,
Wuhan, China) for 1 h at room temperature. The immunoreactive
proteins were enhanced by SuperSignal West Pico Chemiluminescent
Substrate (Thermo Fisher Scientific, Inc.). Each experiment was
performed in triplicate with three replicates each. Densitometry
values were quantified for each band using Image-Pro Plus version
4.0 (Media Cybernetics, Rockville, MD, USA).
Statistical analysis
All data are presented as mean ± standard deviation.
The statistical analyses were performed using SPSS version 13.0
(SPSS, Inc., Chicago, IL, USA). One-way analysis of variance
followed by the Tukey-Kramer test for multiple comparisons were
used to compare the treatment groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
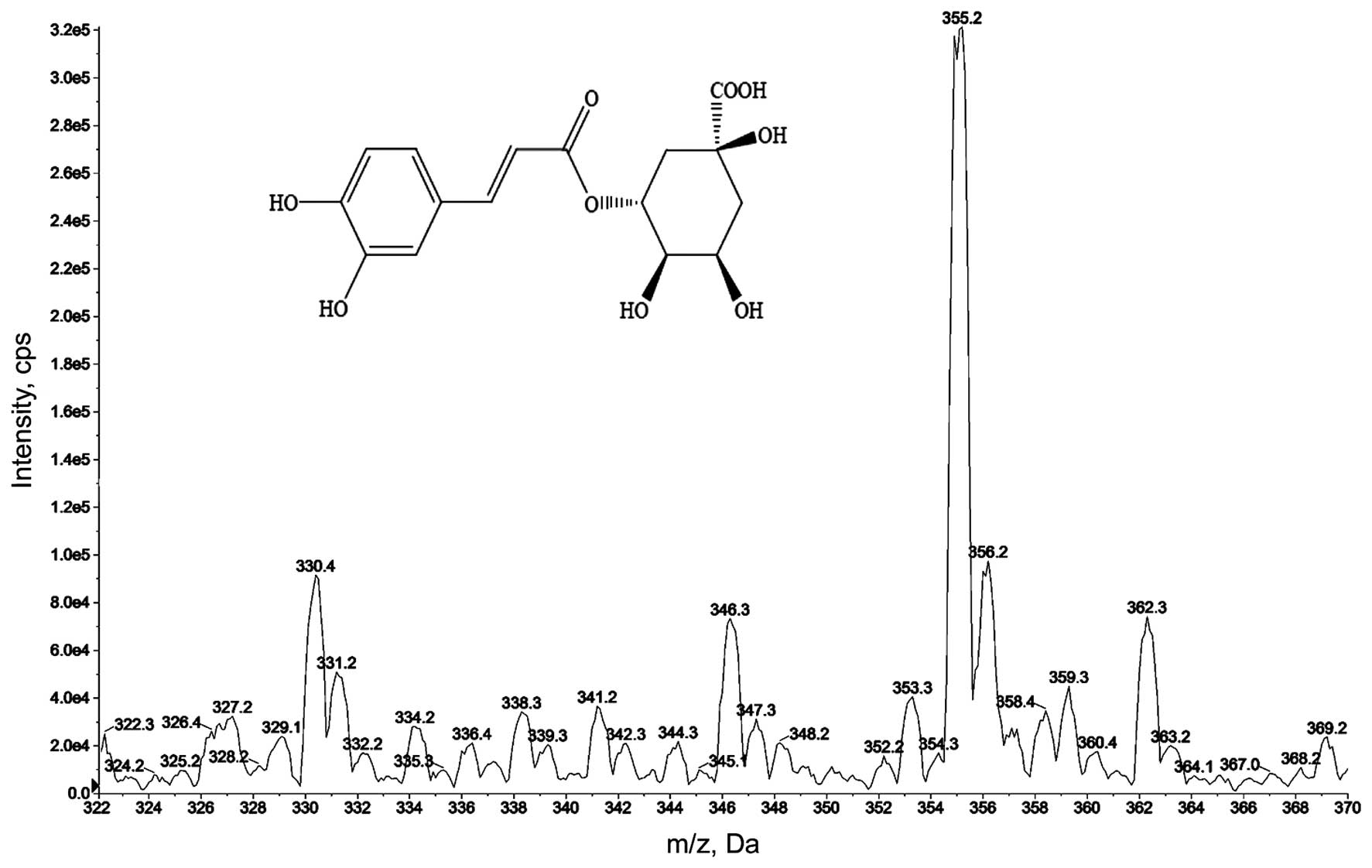
Chemical structures of CGA
CGA is the major component of several medicinal
herbs, such as Caulis Lonicera, which is commonly used in
treatment of RA by traditional Chinese medical practitioners. CGA
(C16H18O9) is one of the most
abundant phenolic acids in nature (23) and its molecular weight is 354.31 Da.
The purity of CGA was detected using an API 3200 LC/MS/MS System.
As revealed in Fig. 1, the greatest
peak is evident at 355.2 Da, providing confirmation that the purity
of CGA used in the present study was suitable for use in
experimental applications.
Selection of the appropriate
stimulating concentrations of IL-6 and CGA
To investigate the effect of IL-6 on the
proliferation of RSC-364 cells and to determine the appropriate
concentration of CGA, an MTT assay was performed to detect the
degree of proliferation of RSC-364 cells stimulated by IL-6 at
different concentrations, in addition to the cytotoxicity
experienced by cells treated with CGA. As displayed in Fig. 2A, RSC-364 cells proliferated
significantly after 24 h of incubation with varying concentrations
of IL-6 compared with those cultured in normal medium. The degree
of proliferation of RSC-364 cells treated with IL-6 (2.5 µg/l)
increased most significantly, as compared with untreated cells
(P=0.003). The toxicity to cells was evident after 24 h of
treatment with CGA at concentrations of 50 and 100 µmol/l (Fig. 2B). The results indicated that
concentrations of 2.5 µg/l IL-6 and 25 µmol/l CGA were most
suitable for stimulating RSC-364 cells in the present study.
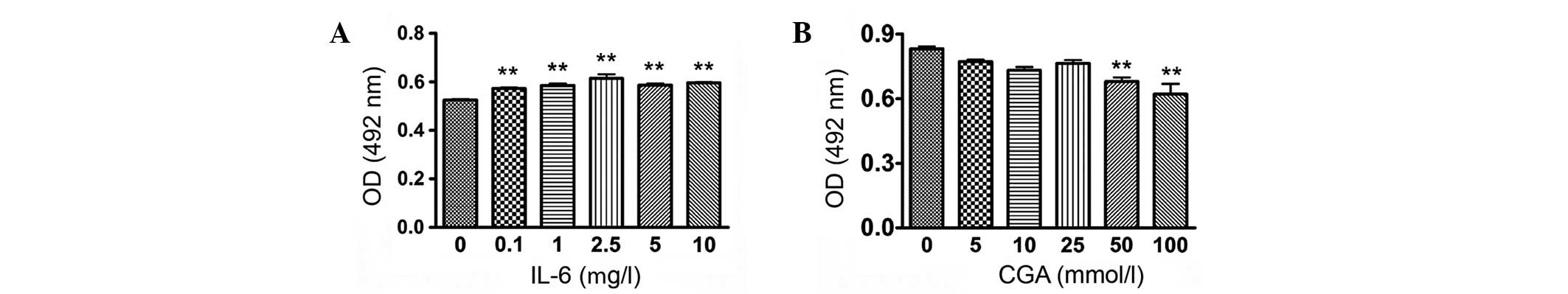 | Figure 2.Determination of appropriate treatment
concentrations of IL-6 and CGA. Viability of RSC-364 cells was
determined following stimulation with (A) IL-6 (0, 0.1, 1, 2.5, 5,
10 µg/l) and (B) CGA (0, 5, 10, 25, 50 and 100 µmol/l) at different
concentrations. Data are expressed as the mean ± standard deviation
of individual groups (n=6 per group). **P<0.01 vs. cells
cultured in normal medium. OD, optical density; IL-6,
interleukin-6; CGA, chlorogenic acid. |
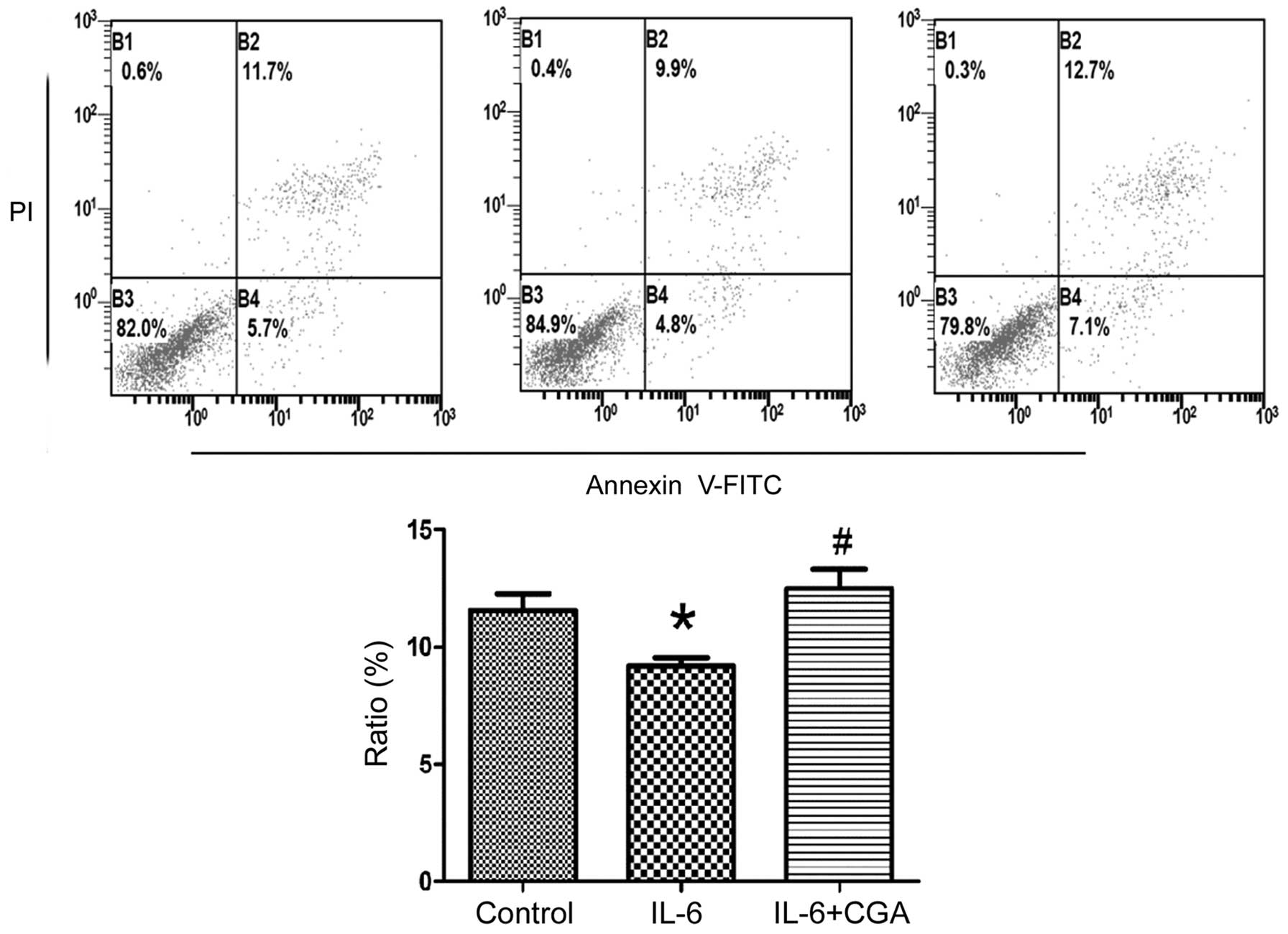
Ability of CGA to induce apoptosis of
RSC-364 cells stimulated by IL-6
RSC-364 cells were treated with IL-6 alone or in
combination with CGA, and apoptosis was detected in the cells by
Annexin V/PI staining. The ratios of apoptosis were observed by
flow cytometry. As displayed in Fig.
3, CGA appeared to significantly induce apoptosis in RSC-364
cells stimulated by IL-6 compared with IL-6 stimulation alone
(P-0.011). By contrast, the ratio of apoptosis in RSC-364 cells was
significantly reduced after 24 h of IL-6 treatment compared with
untreated cells (P=0.037). The results revealed that CGA was able
to increase the ratio of apoptosis significantly in RSC-364 cells
stimulated by IL-6 compared with those incubated with IL-6
alone.
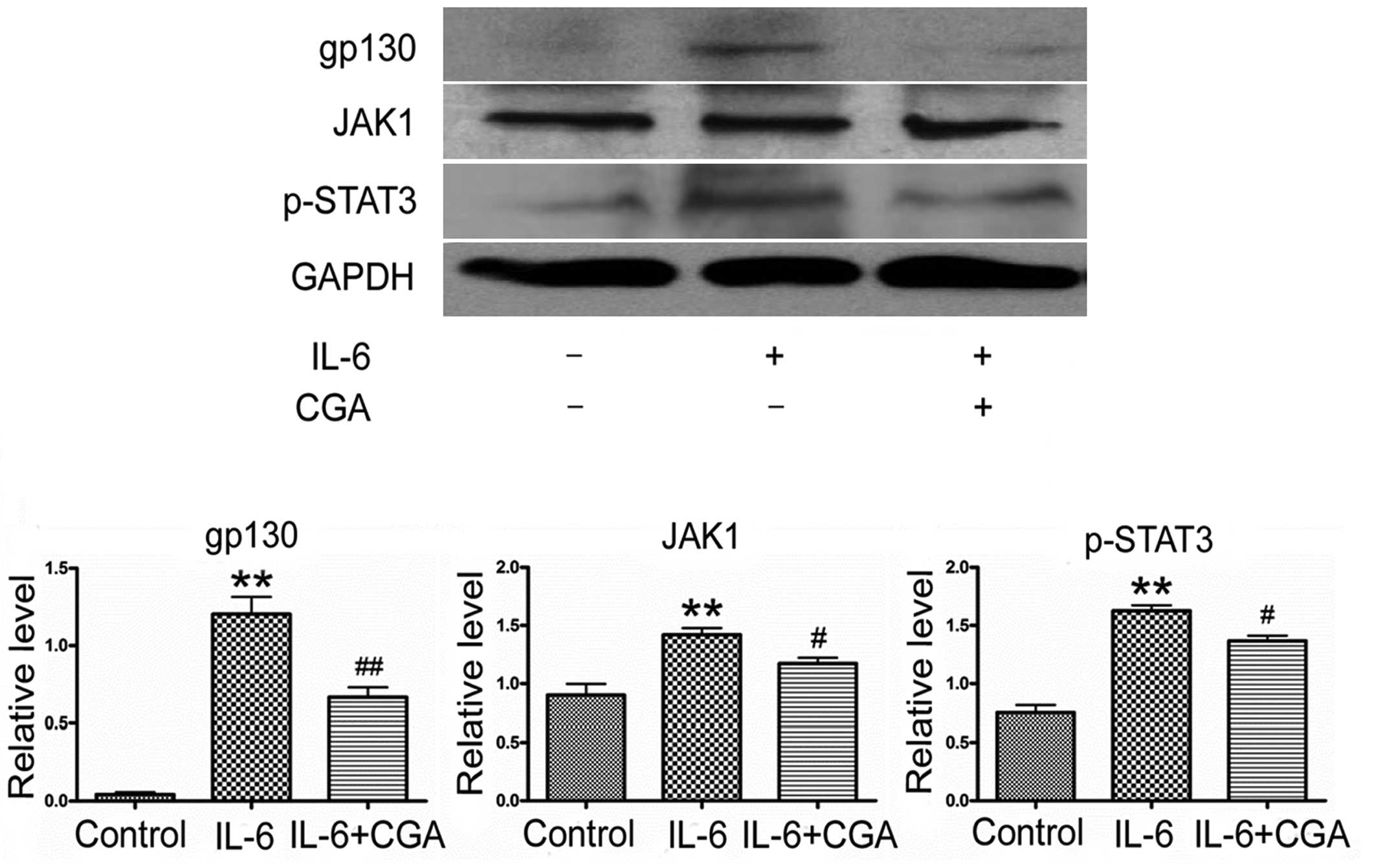
CGA modulates the activation of the
JAK/STAT signaling pathway
Following the observation of the effects of CGA on
the apoptosis of RSC-364 cells, protein expression levels of key
molecules of JAK/STAT signaling were detected in RSC-364 cells
after 24 h of treatment. As is evident in Fig. 4, significantly increased expressions
levels of p-STAT3, JAK1 and gp130 were detected in RSC-364 cells
after 24 h of treatment with IL-6, compared with those cultured in
normal medium (P=0.013, P=0.002 and P=0.001, respectively).
However, the high expression levels of the p-STAT3, JAK1 and gp130
were inhibited significantly by CGA, compared with IL-6 treatment
alone (P=0.002, P=0.048 and P=0.002, respectively).
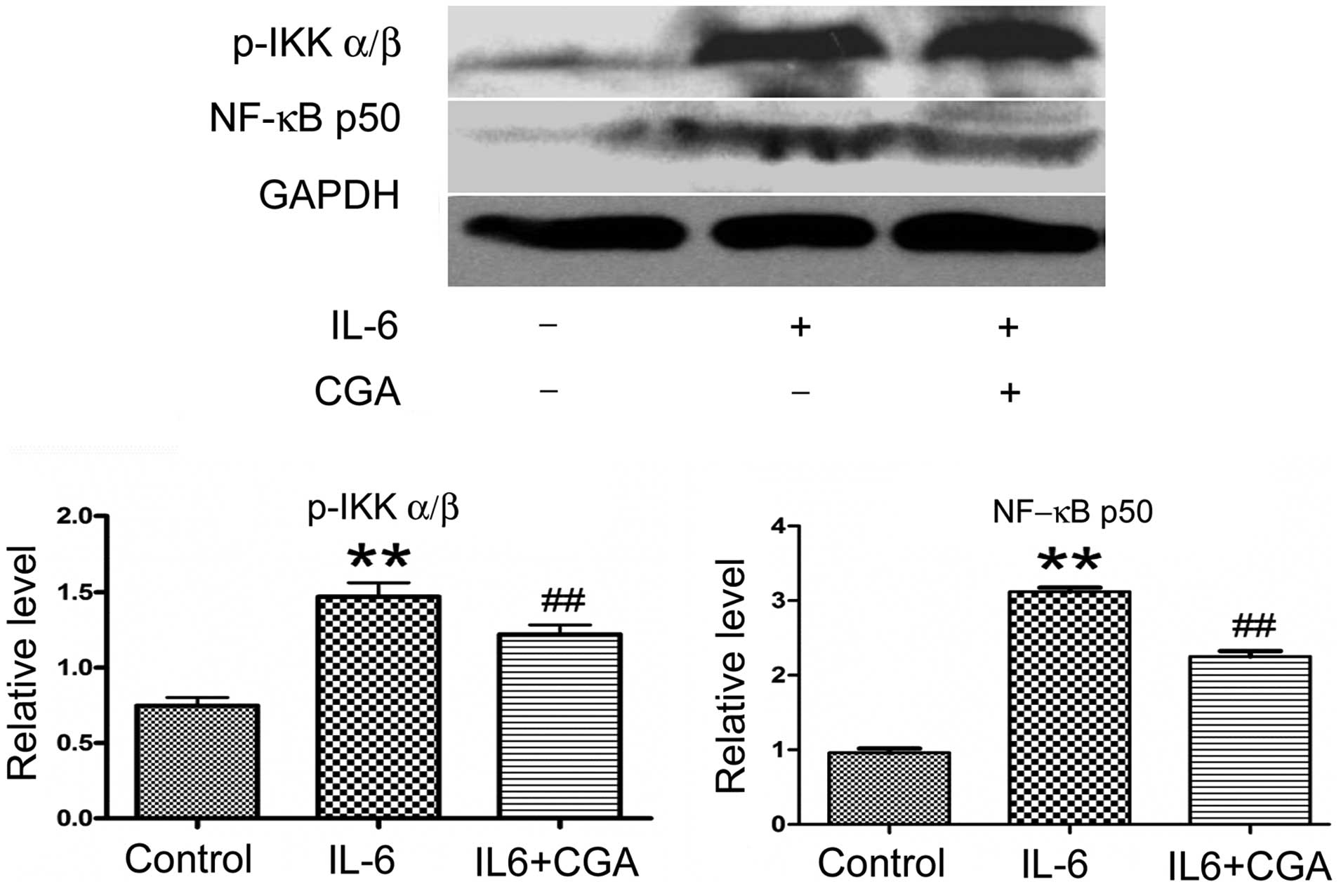
Effect of CGA on suppressing the
activation of the NF-κB signaling pathway
The ability of CGA to modulate the expression levels
of key molecules of the NF-κB signaling pathway in RSC-364 cells
stimulated by IL-6 was also investigated. The expression levels of
NF-κB p50 and p-IKKα/β were increased significantly after 24 h of
IL-6 stimulation (P=0.001 and P=0.003, respectively; Fig. 5) compared with cells cultured in
normal medium. Following incubation with IL-6 and CGA for 24 h, the
expression levels of NF-κB p50 and IKKα/β in RSC-364 cells were
suppressed significantly compared with those cultured in IL-6 alone
(P=0.003 and P=0.026, respectively; Fig.
5).
Discussion
RA is a chronic autoimmune joint disease affecting
~1% of the global population, and involves the small joints of the
hands and feet. The etiology of RA has yet to be elucidated,
however, genetic and environmental influences have been
demonstrated to participate. RA is characterized by persistent
inflammation of the synovial tissues of joints, resulting in loss
of joint function (24). FLSs have
an important role in the initiation and perpetuation of RA, and are
characterized by resistance to apoptosis, consequential
overexpansion and the destruction of articular cartilage. Synovial
tissue is comprised of two layers; the intimal lining and the
sublining. The intimal lining of the synovium displays marked
changes during RA, characterized by an increase in cellularity.
There are two cell types present in the aforementioned structure,
termed type A macrophage-like cells and type B fibroblast-like
cells. Numerous reports indicate that type A cells predominate in
RA by assisting the migration of new cells from the bone marrow
through the circulation, and producing proinflammatory cytokines,
chemokines and growth factors (5,6). The
aforementioned factors activate FLSs in the lining and subsequently
produce array of mediators, including IL-6, prostanoids and matrix
metalloproteinases (25). Therefore,
the pathological reaction is likely to perpetuate synovitis,
recruit new cells to the joint and contribute to the destruction of
the extracellular matrix (26). The
synovial microenvironment in RA supports FLS survival and inhibits
their elimination through apoptosis (27). As a result, proliferation of FLS is
difficult to control, leading to progressive aggravating
inflammation in the lining of synovium, and the destruction of
cartilage and bone.
CGA is the major effective component in Caulis
Lonicera, a Chinese herb commonly used in Chinese medicine
for the treatment of RA. According to several reports, CGA has been
identified to possess pharmacological activity, including
anti-inflammatory, anti-oxidative and anti-carcinogenic effects
(28–30). In the present study, the ability of
CGA to induce apoptosis for the purpose of inhibiting inflammatory
proliferation of IL-6-induced FLS was investigated. The results of
the present study revealed that CGA was able to inhibit
IL-6-mediated proliferation of RSC-364 cells significantly through
promoting cell apoptosis. The aforementioned results indicate that
CGA may be a promising agent for the control of inflammatory
hyperplasia of the synovium in RA.
IL-6 is a pleiotropic proinflammatory cytokine
produced by several cell types in the RA synovial microenvironment,
including macrophages, FLSs and chondrocytes (8). IL-6 exerts its function via the signal
transducer, gp130, leading to the activation of the JAK/STAT
signaling cascade. The complex of IL-6 and its receptor
homodimerizes with gp130 at the cell surface, and subsequently
transduces the signal for the activation of intracytoplasmic JAK
tyrosine kinase. JAK tyrosine kinase is recruited and
preferentially induces tyrosine phosphorylation of STAT3.
Subsequently, p-STAT3 is dimerized. The dimers of p-STAT3
translocate to the nucleus where they bind to the promoter regions
and initiate the transcription of their target genes. Previous
studies have revealed that JAK inhibition has a prominent effect on
autoimmune diseases, including RA (31,32) The
present results revealed that the expression levels of gp130, JAK1
and p-STAT3 in FLS cells induced by IL-6 were suppressed
significantly after 24 h of treatment with CGA. Therefore, the
results of the present study indicate that CGA may be useful for
blocking JAK/STAT-mediated proinflammatory responses through
IL-6-stimulated signaling in RA.
NF-κB is also involved in the pathophysiology of RA
and has an important role in the induction of proinflammatory
cytokines. NF-κB proteins include NF-κB2 p52/p100, NF-κB1 p50/p105,
c-Rel, RelA/p65 and RelB. These proteins function as dimeric
transcription factors that control several genes, regulating a
broad range of biological processes, including innate and adaptive
immunity, in addition to inflammation. NF-κB proteins are bound and
inhibited by inhibitor of κB (IκB) proteins. Proinflammatory
cytokines activate the IKK complex (IKKα and IKKβ), which can
induce the phosphorylation of IκB proteins. The phosphorylation of
IκB results in its ubiquitination and proteasomal degradation,
releasing the NF-κB dimers. Active NF-κB dimers are further
activated by phosphorylation and translocate to the nucleus where
they induce target gene expression (33). Therefore, the present study
determined whether CGA was able to modulate the activation of the
NF-κB signaling pathway in IL-6-stimulated FLS. The results
indicated that CGA was able to downregulate expression levels of
NF-κB p50 and inhibit the phosphorylation of IKKα/β, compared with
those induced by IL-6 alone. Considering the current results, we
hypothesize that CGA has a suppressive function on the activation
of the NF-κB signaling pathway for transmitting proinflammatory
responses in FLSs stimulated by IL-6.
In conclusion, the present study demonstrated that
CGA has an inhibitory function on the proliferation of FLSs in the
inflammatory microenvironment through inducing apoptosis. Thus, it
may be beneficial to inhibit the IL-6-induced proinflammatory
responses meditated by the JAK/STAT and NF-κB signaling cascades.
Therefore, treatment with CGA may be an efficacious therapy for
preventing inflammatory hyperplasia of the synovium in patients
with RA.
Acknowledgements
The present study was jointly supported by the
National Natural Science Foundation of China (grant no. 81173228)
and the Special Fund for the Construction of a Basic R&D
platform of Guangxi Zhuang Autonomous Region (grant no.
KJT13006).
References
|
1
|
Gibofsky A: Overview of epidemiology,
pathophysiology, and diagnosis of rheumatoid arthritis. Am J Manag
Care. 18:S295–S302. 2012.PubMed/NCBI
|
|
2
|
Semerano L, Minichiello E, Bessis N and
Boissier MC: Novel immunotherapeutic avenues for rheumatoid
arthritis. Trends Mol Med. 22:214–229. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Müller-Ladner U, Pap T, Gay RE, Neidhart M
and Gay S: Mechanisms of disease: The molecular and cellular basis
of joint destruction in rheumatoid arthritis. Nat Clin Pract
Rheumatol. 1:102–110. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Firestein GS: Evolving concepts of
rheumatoid arthritis. Nature. 423:356–361. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartok B and Firestein GS: Fibroblast-like
synoviocytes: Key effector cells in rheumatoid arthritis. Immunol
Rev. 233:233–255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huber LC, Distler O, Tarner I, Gay RE, Gay
S and Pap T: Synovial fibroblasts: Key players in rheumatoid
arthritis. Rheumatology (Oxford). 45:669–675. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hirano T: Interleukin 6 and its receptor:
Ten years later. Int Rev Immunol. 16:249–284. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okamoto H, Yamamura M, Morita Y, Harada S,
Makino H and Ota Z: The synovial expression and serum levels of
interleukin-6, interleukin-11, leukemia inhibitory factor, and
oncostatin M in rheumatoid arthritis. Arthritis and Rheum.
40:1096–1105. 1997. View Article : Google Scholar
|
|
9
|
Hirano T, Matsuda T, Turner M, Miyasaka N,
Buchan G, Tang B, Sato K, Shimizu M, Maini R, Feldmann M, et al:
Excessive production of interleukin 6/B cell stimulatory factor-2
in rheumatoid arthritis. Eur J Immunol. 18:1797–1801. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dasgupta B, Corkill M, Kirkham B, Gibson T
and Panayi G: Serial estimation of interleukin 6 as a measure of
systemic disease in rheumatoid arthritis. J Rheumatol. 19:22–25.
1992.PubMed/NCBI
|
|
11
|
Ishihara K and Hirano T: IL-6 in
autoimmune disease and chronic inflammatory proliferative disease.
Cytokine Growth Factor Rev. 13:357–368. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feldmann M, Brennan FM and Maini RN: Role
of cytokines in rheumatoid arthritis. Annu Rev Immunol. 14:397–440.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Alonzi T, Fattori E, Lazzaro D, Costa P,
Probert L, Kollias G, De Benedetti F, Poli V and Ciliberto G:
Interleukin 6 is required for the development of collagen-induced
arthritis. J Exp Med. 187:461–468. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ohshima S, Saeki Y, Mima T, Sasai M,
Nishioka K, Nomura S, Kopf M, Katada Y, Tanaka T, Suemura M and
Kishimoto T: Interleukin 6 plays a key role in the development of
antigen-induced arthritis. Proc Natl Acad Sci USA. 95:8222–8226.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hirano T, Ishihara K and Hibi M: Roles of
STAT3 in mediating the cell growth, differentiation and survival
signals relayed through the IL-6 family of cytokine receptors.
Oncogene. 19:2548–2556. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kong X, Liu C, Zhang C, Zhao J, Wang J,
Wan H, Zhu H, Zhang P, Chen W, Xiao Y and Lin N: The suppressive
effects of Saposhnikovia divaricata (Fangfeng) chromone extract on
rheumatoid arthritis via inhibition of nuclear factor-κB and
mitogen activated proteinkinases activation on collagen-induced
arthritis model. J Ethnopharmacol. 148:842–850. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Andreas K, Lübke C, Häupl T, Dehne T,
Morawietz L, Ringe J, Kaps C and Sittinger M: Key regulatory
molecules of cartilage destruction in rheumatoid arthritis: An in
vitro study. Arthritis Res Ther. 10:R92008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ruan Z, Liu S, Zhou Y, Mi S, Liu G, Wu X,
Yao K, Assaad H, Deng Z, Hou Y, et al: Chlorogenic acid decreases
intestinal permeability and increases expression of intestinal
tight junction proteins in weaned rats challenged with LPS. PLoS
One. 9:e978152014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
dos Santos MD, Almeida MC, Lopes NP and de
Souza GE: Evaluation of the anti-inflammatory, analgesic and
antipyretic activities of the natural polyphenol chlorogenic acid.
Biol Pharm Bull. 29:2236–2240. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tatefuji T, Izumi N, Ohta T, Arai S, Ikeda
M and Kurimoto M: Isolation and identification of compounds from
Brazilian propolis which enhance macrophage spreading and mobility.
Biol Pharm Bull. 19:966–970. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang J, Ma J, Dong L, Hou Y, Jia X, Niu X
and Fan Y: Effect of anatase TiO2 nanoparticles on the growth of
RSC-364 rat synovial cell. J Nanosci Nanotechnol. 13:3874–3879.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guo HF, Liu SX, Zhang YJ, Liu QJ, Hao J
and Gao LX: High mobility group box 1 induces synoviocyte
proliferation in rheumatoid arthritis by activating the signal
transducer and activator transcription signal pathway. Clin Exp
Med. 11:65–74. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kwak SC, Lee C, Kim JY, Oh HM, So HS, Lee
MS, Rho MC and Oh J: Chlorogenic acid inhibits osteoclast
differentiation and bone resorption by down-regulation of receptor
activator of nuclear factor kappa-B ligand-induced nuclear factor
of activated T cells c1 expression. Biol Pharm Bull. 36:1779–1786.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan L, Wang Q, Liu R, Zong M, He D, Zhang
H, Ding Y and Ma J: Citrullinated fibronectin inhibits apoptosis
and promotes the secretion of pro-inflammatory cytokines in
fibroblast-like synoviocytes in rheumatoid arthritis. Arthritis Res
Ther. 14:R2662012. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li G, Liu D, Guo S, Sunagawa M, Hisamitsu
T and Liu Y: Anti-invasive effects of Celastrus Orbiculatus extract
on interleukin-1 beta and tumour necrosis factor-alpha
combination-stimulated fibroblast-like synoviocytes. BMC Complement
Altern Med. 14:622014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marinova-Mutafchieva L, Williams RO, Funa
K, Maini RN and Zvaifler NJ: Inflammation is preceded by tumor
necrosis factor-dependent infiltration of mesenchymal cells in
experimental arthritis. Arthritis and Rheum. 46:507–513. 2002.
View Article : Google Scholar
|
|
27
|
Nakajima T, Aono H, Hasunuma T, Yamamoto
K, Shirai T, Hirohata K and Nishioka K: Apoptosis and functional
Fas antigen in rheumatoid arthritis synoviocytes. Arthritis and
Rheum. 38:485–491. 1995. View Article : Google Scholar
|
|
28
|
Liu YJ, Zhou CY, Qiu CH, Lu XM and Wang
YT: Chlorogenic acid induced apoptosis and inhibition of
proliferation in human acute promyelocytic leukemia HL60 cells. Mol
Med Rep. 8:1106–1110. 2013.PubMed/NCBI
|
|
29
|
Wu C, Luan H, Zhang X, Wang S, Zhang X,
Sun X and Guo P: Chlorogenic acid protects against atherosclerosis
in ApoE-/- mice and promotes cholesterol efflux from RAW264.7
macrophages. PLoS One. 9:e954522014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo YJ, Luo T, Wu F, Mei YW, Peng J, Liu
H, Li HR, Zhang SL, Dong JH, Fang Y and Zhao L: Involvement of TLR2
and TLR9 in the anti-inflammatory effects of chlorogenic acid in
HSV-1-infected microglia. Life Sci. 127:12–18. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Migita K, Komori A, Torigoshi T, Maeda Y,
Izumi Y, Jiuchi Y, Miyashita T, Nakamura M, Motokawa S and
Ishibashi H: CP690,550 inhibits oncostatin M-induced JAK/STAT
signaling pathway in rheumatoid synoviocytes. Arthritis Res Ther.
13:R722011. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Borie DC, O'Shea JJ and Changelian PS:
JAK3 inhibition, a viable new modality of immunosuppression for
solid organ transplants. Trends Mol Med. 10:532–541. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008. View Article : Google Scholar : PubMed/NCBI
|