Introduction
Depression is a common complication of stroke,
occurring in ~33% of patients (1,2); the
clinical symptoms of this include feelings of guilt, low
self-esteem, reduction in food intake, sleep disorders and fatigue.
There are between 1.6–2.0 million new stroke patients in China
every year and post-stroke depression (PSD) is closely associated
with the increased risk of mortality following a stroke (3). PSD negatively impacts upon subsequent
rehabilitation; in a previous study by Sinyor et al, PSD
patients demonstrated greater functional impairment than patients
without depression, also scoring lower on behavioral action and
functional status (4). The severity
of depression is closely associated with health-associated quality
of life and functional recovery in stroke survivors (5). PSD patients demonstrate a negative mood
during rehabilitation and achieve poorer outcomes during functional
recovery therapy. These patients also have greater difficulty in
restoring social activities (6).
Several studies have described a higher mortality rate in stroke
patients with depression (7), and
certain studies have reported that PSD patients using
antidepressant drugs demonstrate improved ability to function than
those without antidepressant therapy. The treatment of depression
has been demonstrated to aid functional recovery (8), meaning that the early diagnosis and
effective treatment of PSD are crucial to recovery from a
stroke.
Brain-derived neurotrophic factor (BDNF) is a member
of the neurotrophin family, which regulates neurogenesis (9), apoptosis (10), the expression level of monoamine
transmitters (11), and the function
and plasticity of synapses (12).
Neurotrophins may be key in the development of depression (13); BDNF, for instance, can activate
intracellular mitogen-activated protein kinase/extracellular
signal-regulated kinase cascade signaling pathways (14), affecting synaptic plasticity
(12) and alleviating the symptoms
of depression (15), and BDNF may
thus be important in the maintenance of emotional stability.
Deletion of the BDNF receptor induces a reduction in neurogenesis
and increases anxious behavior (16). Additionally, BDNF expression is
associated with 5-hydroxytryptamine receptor expression in the
brain, particularly the hippocampus (11), and is hypothesized to treat
depression in animals by improving the activity of monoamine
transmitters in response to antidepressant drugs (17). BDNF is therefore considered to be a
key neurotrophic factor that modulates a depressive mood through
its activity in the hippocampus. In clinical practice, serum BDNF
is lower in patients with depression, but antidepressant treatment
can induce an increase in BDNF levels (18), making serum BDNF a sensitive
peripheral marker that is predictive of the severity of depression
(19) and of the treatment outcome
(20). Concordantly, previous
studies have indicated that the concentration of BDNF is lower in
PSD patients than in non-PSD patients (21). Prior studies predominantly focused on
the association between serum BDNF levels and depression severity.
BDNF is expressed in multiple regions of the brain, including the
hippocampus, and is involved in brain functionality (22), making BDNF levels a useful metric to
predict the severity of depression in PSD patients.
Estrogen is an effective mood regulator and causes
antidepressant-like effects on a depression model in rats (23). Estrogen acts on the central nervous
system through its nuclear receptors. There are two well-known
types of estrogen receptors (ER): ERα and ERβ. ERs are widely
distributed in the brain and ERβ is abundantly expressed in the rat
hippocampus (24). Estrogen
implements its antidepressive effects by activating ERβ (25). Numerous previous studies have
indicated that estrogen and BDNF stimulate neurogenesis in the
hippocampus (9,26) and are involved in synaptic
modification (27,28) to enhance learning and memory
(28–30). Resultantly, the present study
investigated the association between estrogen and BDNF in the
hippocampus and the hypothesis that BDNF is involved in
estrogen-mediated antidepressant effects upon PSD.
Materials and methods
Animals
Female two-month-old Sprague-Dawley rats, each
weighing 230–250 g, were used in all experiments (Wenzhou medical
University, Wenzhou, China). The rats were housed under the
following conditions: A 12:12-h light-dark cycle, with lights on at
7:00 a.m.; temperature maintained at 21±2°C; and the provision of
food and water ad libitum. All animal procedures were
performed in accordance with the guidelines of the Animal Care and
Use Committee of Wenzhou Medical University. All surgical
procedures were performed under chloral hydrate anesthesia
(21). In order to develop a PSD
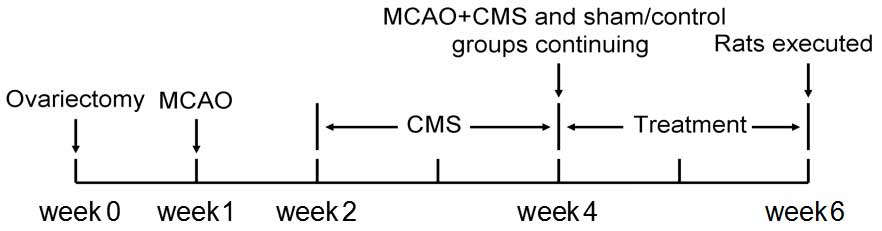
model, the rats were randomly divided into four groups as follows:
The sham/control group, the middle cerebral artery occlusion
(MCAO)/control group, the sham/chronic mild stress (CMS) group and
the MCAO+CMS group (n=20). Comparison of these groups was used to
validate the animal models. The sham/control (control) group and
the MCAO+CMS (PSD) group were then randomly divided into matched
subgroups for subsequent estrogen treatment, as follows: The
vehicle-treated control group, the estradiol (E2)-treated control
group, the vehicle-treated PSD group and the E2-treated PSD group
(n=10). Comparison of the four groups was used to evaluate the
effect of estrogen on PSD. The entire experiment lasted for six
weeks (Fig. 1).
Ovariectomy
All rats were subjected to an ovariectomy prior to
grouping. All experimental procedures were approved by the ethics
committee of Wenzhou Medical University. Under chloral hydrate
anesthesia (3.2 mg/kg), rats were kept in a supine position on the
operating table. Surgery was performed through a median incision on
the back, under chloral hydrate anesthesia. Two small cuts were
made on the muscle 1–2 cm away from lumbar vertebrae 1 and 2. The
arteries beside the ovaries and the ovaries themselves were ligated
and removed.
MCAO
A week after ovariectomy, the MCAO/control and
MCAO+CMS group rats were anesthetized with chloral hydrate (3.2
mg/kg). MCAO was induced using the intraluminal suture occlusion
technique (31) and rats were placed
on a temperature-controlled heating pad throughout surgery. The
left common, external and internal carotid arteries were exposed.
The external carotid artery was ligated and cut off, and the middle
cerebral artery was occluded by inserting a 3-0 suture from the
basal section of the external carotid artery and advanced cranially
into the internal carotid artery. The suture was inserted ~18 mm
into the internal carotid artery and was carefully withdrawn 1.5 h
after MCAO onset. The neurological deficits were evaluated by
Bederson test (32). All scores of
rats used in the present study were >1. Rats included in the
sham group underwent sham surgery, which involved the same
procedure, but without inserting the suture.
CMS procedure
The rats from the sham/CMS and MCAO+CMS groups were
exposed to CMS procedures for a period of two consecutive weeks,
beginning one week after MCAO. The weekly stress procedure
consisted of i) food and water deprivation for 24 h; ii) cage tilt
(40°), in which the rats' cages were tilted for 24 h; iii) soiled
cages, in which the sawdust in the cages was kept wet and dirty
with 200 ml water in the sawdust and bedding to maintain a poor
living environment for 24 h; iv) day and night reversal, involving
turning on the light at 8:00 p.m. and off at 8:00 a.m.; v)
restraint stress, which involved binding the rats to a fixator for
2 h so that the rats could not run or turn their bodies over; vi)
cage tremor, in which the cages were put on a horizontal agitator
that tremored continuously for 40 min; and vii) clamping of the
rats' tails (3 cm from the end) using 3-cm clips. Each rat was
isolated throughout the CMS procedure. The control groups were
maintained in suitable living conditions.
Drug administration
Following the CMS procedure, a MCAO/CMS (PSD) group
and a ham/control (control) group of rats were injected
subcutaneously with 10 µg 17β-estradiol (E2758; Sigma-Aldrich, St.
Louis, MO, USA) in 0.1 ml soybean oil between 9:00 and 10:00 a.m.
every day for two weeks. In another PSD group and control group,
the vehicle was administered in the same manner.
Behavioral tests
Sucrose preference test
Each group of rats had free access to two bottles
containing 1% sucrose solution on the first day and two bottles,
one containing 1% sucrose solution and the other containing water,
on the second day in order to adjust to this test. The rats were
deprived of food and water for 23 h prior to the sucrose preference
test, which was performed as follows: Rats were provided with two
bottles, one containing 1% sucrose solution and the other
containing water, beginning between 9:00 and 10:00 a.m. Data were
collected by weighing the two bottles after 1 h, in order for the
comparative consumption of sucrose solution and water to be
analyzed and a preference calculated using the formula: Preference
(%) = Sucrose solution intake / total intake × 100. The baseline
sucrose preference test was performed prior to MCAO, and the
sucrose preference was monitored every week during periods of
establishment of the model and treatment.
Open field test
A quadrate box was used for this experiment. The box
was 40 cm in height, 100 cm in length and 100 cm in width. The wall
was black and the floor was divided into 25 squares. Rats were
placed on the floor and the scores of the horizontal movement and
vertical movement were recorded for 3 min). The baseline open field
test performance was recorded prior to MCAO, and locomotory
activity during the open field test was monitored every week whilst
establishing the model.
Forced swimming test
The forced swimming test is a standard test used to
screen compounds for antidepressant-like activity. Swim sessions
were conducted by placing rats in glass cylinders (45 cm tall × 18
cm in diameter) containing 23–25°C water at a 30-cm depth. All rats
were placed in this cylinder individually, with no escape platform,
for 15 min as an initial pre-test. The following day, each rat was
placed in the cylinders for 5 min. Rats were then removed from the
cylinders and dried with paper towels. During the test period, the
time of 2 behaviors were recorded: i) Immobility: A rat was judged
to be immobile when it remained floating in the water without
struggling and was only making slight movements necessary to keep
its head above water; and ii) struggling: A rat was judged to be
struggling when it was making active attempts to escape from the
cylinder, including searching for the escape routes and diving.
This test was carried out every week during the period of
treatment.
Western blot analysis
Animals were sacrificed after 2 weeks of treatment.
Rats were anesthetized and decapitated, and their brains were
rapidly removed and placed on ice for immediate dissection of the
required brain regions. Dissected tissue was frozen and stored at
−80°C until the next experiment.
The hippocampus was isolated and homogenized in cell
lysis buffer containing phenylmethanesulfonyl fluoride (Beyotime,
Shanghai, China). Proteins (100 µg total protein) from hippocampus
extracts were separated on 12% sodium dodecyl
sulfate-polyacrylamide gels (Beyotime); following this, proteins
were transferred to a polyvinylidene difluoride membrane (Beyotime)
using electroblotting. The membranes were incubated with a blocking
solution comprising Tris-buffered saline (Beyotime) and 0.05%
Tween-20 (Beyotime) containing 5% skimmed milk for 2 h. Following
blocking, the membranes were incubated with rabbit anti-BDNF
polyclonal antibody (1:200; ab6201; Abcam, Cambridge, MA, USA) at
4°C for 18 h, then the membranes were washed and incubated with
HRP-labeled goat anti-rabbit secondary antibody for 1 h at room
temperature and developed using enhanced chemiluminescence. The
X-OMAT BT films (Kodak, Rochester, NY, USA) were scanned and
densitometric analyses were performed using Quantity One (Bio-Rad,
Berkeley, CA, USA). β-actin was used as an endogenous loading
control.
Statistics
A repeated measures analysis of variance (ANOVA),
with treatment and time as the 2 variables, was used to analyze the
data arising from behavioral tests. Post hoc analyses for multiple
comparisons were made using a least significant difference (LSD)
test. Group differences were considered statistically significant
at P<0.05. The BDNF protein level was analyzed by one-way ANOVA.
Furthermore, results from post hoc tests using LSD measures with
P<0.05 were considered to be statistically significant.
Results
Establishment of the PSD animal
model
Sucrose preference test
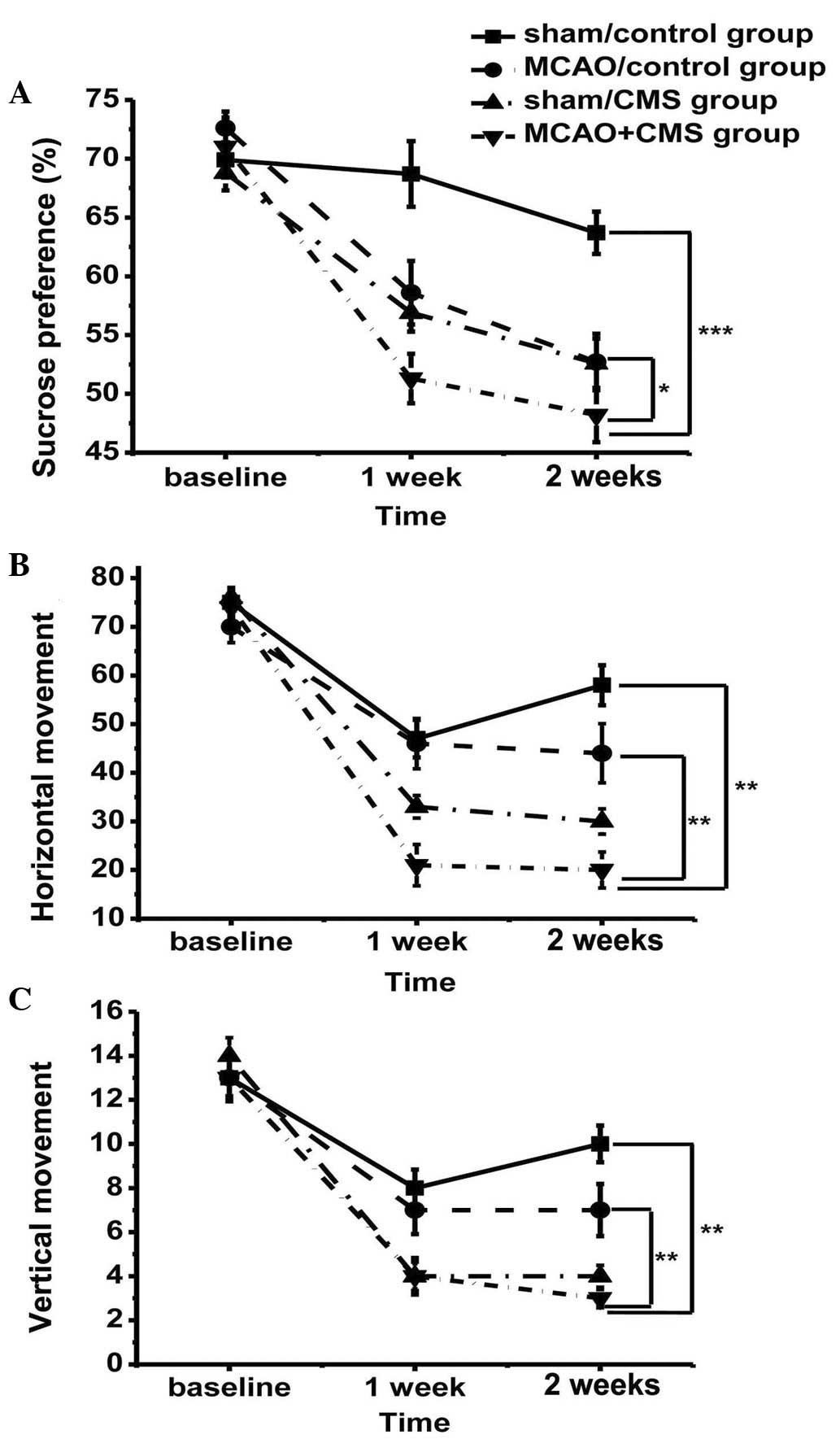
A repeated measures ANOVA of sucrose preference
level between groups during model establishment determined that
consumption of sucrose decreased during the CMS procedure. Variance
(group vs. time) was analyzed by repeated measures, revealing no
identifiable significant effects with regard to time (F=3.015;
dF=2; P=0.055), but a significant difference among groups in terms
of sucrose preference (F(3,76)=12.85; P<0.05).
Sucrose preference was considerably reduced in the MCAO+CMS group
compared with the other groups, arising after only one week of
stress and becoming markedly different on the second week (Fig. 2A); post hoc analysis revealed that
the difference between the MCAO+CMS and sham/control groups was
statistically significant (P<0.05). The sucrose preference of
the MCAO+CMS group was lower than that of the sham/CMS group
(Fig. 2A), but there was no
statistical difference between the two groups. This result
demonstrated severe depression in the MCAO+CMS group, meaning that
MCAO+CMS treatment was successful in establishing a PSD model. Due
to the destruction of brain tissue, the sucrose preference of the
MCAO/control group declined, but the difference between the
MCAO+CMS and MCAO/control groups was statistically significant
(P<0.05) (Fig. 2A).
Open field test
In order to further demonstrate the severity of
depression, rats were also assessed for changes in activity.
Variance (group vs. time) analyses demonstrated a significant
time-by-treatment interaction (F=2.322; dF=6; P=0.042). Rats
gradually adapted to the open field box and groups undergoing CMS
demonstrated significantly reduced horizontal (P<0.05; Fig. 2B) and vertical activity (P<0.05;
Fig. 2C). An acute reduction in
movement occurred in the first week of model establishment. A
significant difference was reported between the groups after two
weeks in terms of horizontal (F(3,76)=9.586; P<0.05;
Fig. 2B) and vertical
(F(3,76)=7.929; P<0.05; Fig. 2C) movement. Post hoc analyses
demonstrated a significant difference between the MCAO+CMS group
and the other groups in terms of horizontal and vertical movement
(P<0.05). The MCAO+CMS rats were less curious to explore their
surroundings and exhibited depression.
Evaluation of the antidepressive
effects of estrogen
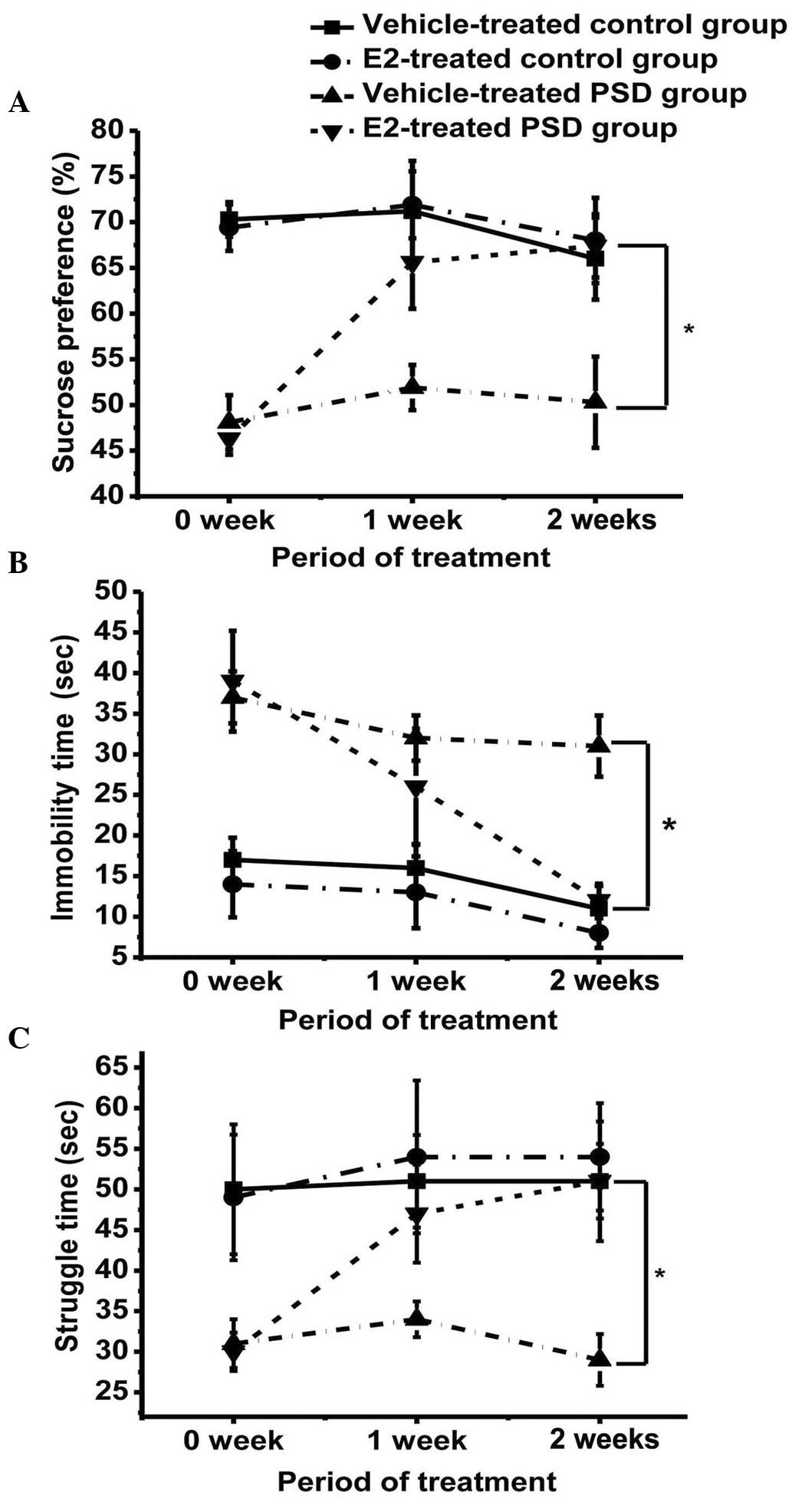
Sucrose preference test
The sucrose preference of the vehicle-treated and
estrogen-treated groups was compared; two weeks of estrogen
treatment led to a significant increase in sucrose preference in
the E2-treated PSD groups (P<0.05), but no significant change in
the vehicle-treated groups (Fig.
3A). The sucrose preference of the E2-treated PSD group was
similar to the control groups, although a statistically significant
difference remained.
Forced swimming test
Immobility time was reduced and duration of
struggling increased in the E2-treated PSD group over the treatment
period, when compared with the vehicle-treated PSD group
(P<0.05), approaching the mean levels of the control groups.
However, the difference in immobility between the E2-treated PSD
group and the two control groups remained statistically significant
(Fig. 3B). The struggle time of the
E2-treated PSD rats significantly increased following E2 treatment
(P<0.05; Fig. 3C); however, no
significant difference was identified between the E2-treated PSD
group and the two control groups (P>0.05), indicating a rescue
of mood in the PSD group following estrogen treatment
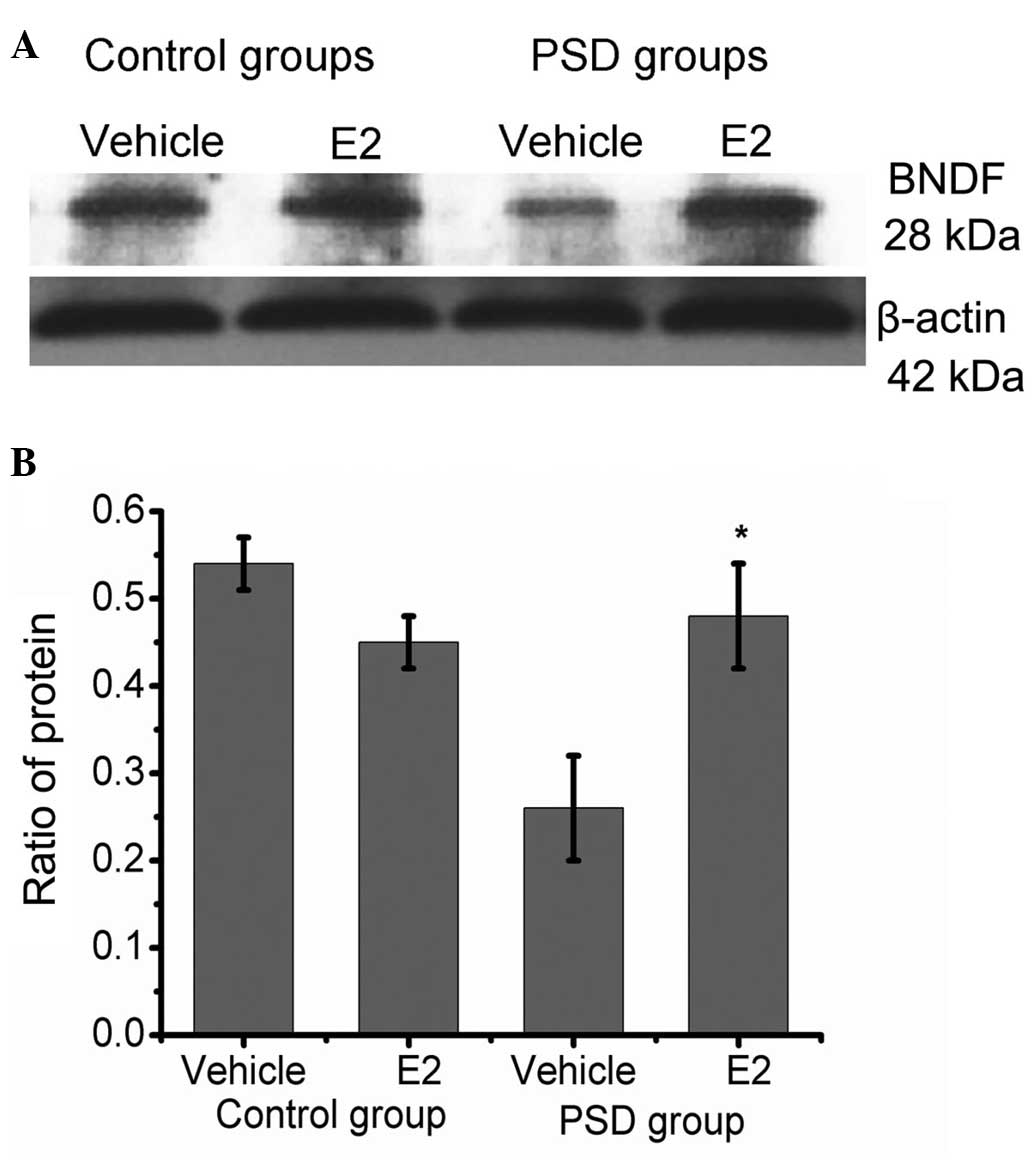
Expression of BDNF in the hippocampus
Protein level was measured by western blotting after
two weeks of treatment. The BDNF band is indicated at ~28 kDa. The
ratio of BDNF:β-actin was used for statistical analysis. This ratio
was different between the PSD groups (P=0.014), suggesting that the
BDNF expression of the E2-treated PSD group was significantly
increased compared with the vehicle-treated PSD group (P<0.05).
No significant difference between the E2-treated PSD and control
groups was identified (P>0.05; Fig.
4).
Discussion
PSD occurs at high incidence rates in male and
female stroke patients (1), but
numerous previous studies have also revealed gender differences in
PSD occurrence; these studies have predominantly reported a higher
prevalence of depression among women than men (33). The higher frequency of PSD in female
patients may be associated with medical conditions, likelihood of
rehabilitation and hormonal changes, amongst other causes, but
hormonal level is considered to be a particularly noteworthy
contributory factor. Age is similarly correlated with the frequency
of PSD (34); female stroke patients
are typically of an advanced age (34) and commonly undergoing menopause. This
indicates that changes to estrogen levels may be a key contributory
factor to the high prevalence of PSD in female patients. As a
consequence, ovariectomy-treated rats were selected for use in the
present study.
In the current study, a PSD model was established
through MCAO, followed by CMS, to imitate the pathogenesis of PSD.
PSD rats demonstrated severe depressive behaviors compared with
MCAO and CMS rats, based on sucrose preference and open field
tests. Brain injury and poor living conditions in the present study
may have contributed to the observed mood disorders; these were
introduced to recapitulate clinical conditions, in which depression
is caused by pathological changes to the brain and changes to
lifestyle following a stroke. In the present study, MCAO was used
to mimic trauma to the brain, specifically in the region of the
brain controlling emotion, and CMS conditions were used to imitate
the difficult life changes encountered following a stroke. Stroke
patients often feel useless when they are unable to perform tasks
that were previously easy and living conditions become
uncomfortable due to their disability, so the present study aimed
to replicate this.
The effects of estrogen on BDNF expression in the
hippocampus of the PSD rats and on animal behavior, reflective of
mood, were analyzed. Estrogen replacement rescued BDNF protein
expression in the hippocampus and improved depression following a
stroke. Preference of sucrose and performance in a forced swimming
test were regarded as key indicators to evaluate the antidepressant
effect during treatment. The key finding of the present study was
that estrogen could significantly improve depression in PSD rats,
according to these metrics, through regulation of BDNF.
Estrogen has been revealed to improve depression and
to have neuroprotective effects on stroke patients (35). The present study demonstrated the
therapeutic action of estrogen on the bio-psycho-social onset of
PSD. Previous studies have reported that estrogen regulates the
cell cycle at the G1/S transition, boosts neurogenesis following ER
activation (36), and promotes
B-cell lymphoma 2 expression and a decrease in apoptosis (37,38). The
inflammatory reaction following stroke causes expansive damage in
the brain (39), but this secondary
injury following stroke can be blocked by estrogen (35). Consequently, neurological function is
significantly improved following estrogen treatment due to
increased neurogenesis and decreased ischemia-associated apoptosis
(35). Estrogen may thus be an
appropriate treatment for PSD, as confirmed by the present
study.
BDNF has an important function in emotional
stability; it has been associated with the development of
depression in clinical and experimental studies (40), and is an important marker to evaluate
the severity of depression. In a previous study, the BDNF level was
low in PSD patients (21), and the
present study reiterated this finding, demonstrating that BDNF
expression in the PSD rats was substantially downregulated. In the
current study, BDNF expression was significantly increased in the
PSD group treated with E2, approaching the BDNF level of the
control group.
There are a number of similarities between estrogen
and BDNF, which interact to exert their effects (41,42). A
possible hypothesis is that estrogen can affect BDNF expression and
that estrogen can promote neurogenesis. ER activation enhances cell
proliferation (36), which increased
BDNF expression following estrogen treatment in the present study.
Taken together, it appears plausible that estrogen promotes the
release of BDNF, subsequently regulating the depressive mood
following a stroke.
The estrogen-BDNF interaction begins an important
molecular cascade to modulate the function of the hippocampus,
which may be responsible for the antidepressant effects associated
with estrogen treatment.
In summary, estrogen replacement therapy is suitable
for the treatment of the PSD observed in ovariectomized rats and
BDNF signaling may be the critical pathway by which estrogen is
able to treat PSD; these findings may contribute to the treatment
of PSD patients.
Acknowledgements
The present study was supported by grants from
Wenzhou Technology Division (no. H20090067) and the National
Institute of Health (nos. AG21980 and NS057186).
References
|
1
|
Hackett ML, Yapa C, Parag V and Anderson
CS: Frequency of depression after stroke: A systematic review of
observational studies. Stroke. 36:1330–1340. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weimar C, Kurth T, Kraywinkel K, Wagner M,
Busse O, Haberl RL and Diener HC: German Stroke Data Bank
Collaborators: Assessment of functioning and disability after
ischemic stroke. Stroke. 33:2053–2059. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Williams LS, Ghose SS and Swindle RW:
Depression and other mental health diagnoses increase mortality
risk after ischemic stroke. Am J Psychiatry. 161:1090–1095. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sinyor D, Amato P, Kaloupek DG, Becker R,
Goldenberg M and Coopersmith H: Post-stroke depression:
Relationships to functional impairment, coping strategies, and
rehabilitation outcome. Stroke. 17:1102–1107. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
CarodArtal FJ and Egido JA: Quality of
life after stroke: The importance of a good recovery. Cerebrovasc
Dis. 27(Suppl 1): 204–214. 2009. View Article : Google Scholar
|
|
6
|
Feibel JH and Springer CJ: Depression and
failure to resume social activities after stroke. Arch Phys Med
Rehabil. 63:276–277. 1982.PubMed/NCBI
|
|
7
|
Janzing JG, Bouwens JM, Teunisse RJ, Van't
Hof MA and Zitman FG: The relationship between depression and
mortality in elderly subjects with less severe dementia. Psychol
Med. 29:979–983. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bilge C, Koçer E, Koçer A and Türk Börü U:
Depression and functional outcome after stroke: The effect of
antidepressant therapy on functional recovery. Eur J Phys Rehabil
Med. 44:13–18. 2008.PubMed/NCBI
|
|
9
|
Rossi C, Angelucci A, Costantin L, Braschi
C, Mazzantini M, Babbini F, Fabbri ME, Tessarollo L, Maffei L,
Berardi N and Caleo M: Brain-derived neurotrophic factor (BDNF) is
required for the enhancement of hippocampal neurogenesis following
environmental enrichment. Eur J Neurosci. 24:1850–1856. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Peng CH, Chiou SH, Chen SJ, Chou YC, Ku
HH, Cheng CK, Yen CJ, Tsai TH, Chang YL and Kao CL: Neuroprotection
by Imipramine against lipopolysaccharide-induced apoptosis in
hippocampus-derived neural stem cells mediated by activation of
BDNF and the MAPK pathway. Eur Neuropsychopharmacol. 18:128–140.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luellen BA, Bianco LE, Schneider LM and
Andrews AM: Reduced brain-derived neurotrophic factor is associated
with a loss of serotonergic innervation in the hippocampus of aging
mice. Genes Brain Behav. 6:482–490. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hall J, Thomas KL and Everitt BJ: Rapid
and selective induction of BDNF expression in the hippocampus
during contextual learning. Nat Neurosci. 3:533–535. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Duman RS and Monteggia LM: A neurotrophic
model for stress-related mood disorders. Biol Psychiatry.
59:1116–1127. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mohajerani MH, Sivakumaran S, Zacchi P,
Aguilera P and Cherubini E: Correlated network activity enhances
synaptic efficacy via BDNF and the ERK pathway at immature CA3 CA1
connections in the hippocampus. Proc Natl Acad Sci USA.
104:13176–13181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Luikart BW, Birnbaum S, Chen J, Kwon
CH, Kernie SG, BasselDuby R and Parada LF: TrkB regulates
hippocampal neurogenesis and governs sensitivity to antidepressive
treatment. Neuron. 59:399–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bergami M, Rimondini R, Santi S, Blum R,
Götz M and Canossa M: Deletion of TrkB in adult progenitors alters
newborn neuron integration into hippocampal circuits and increases
anxiety-like behavior. Proc Natl Acad Sci USA. 105:15570–15575.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Siuciak JA, Lewis DR, Wiegand SJ and
Lindsay RM: Antidepressant-like effect of brain-derived
neurotrophic factor (BDNF). Pharmacol Biochem Behav. 56:131–137.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sen S, Duman R and Sanacora G: Serum
brain-derived neurotrophic factor, depression, and antidepressant
medications: Meta-analyses and implications. Biol Psychiatry.
64:527–532. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Satomura E, Baba H, Nakano Y, Maeshima H,
Suzuki T and Arai H: Correlations between brain-derived
neurotrophic factor and clinical symptoms in medicated patients
with major depression. J Affect Disord. 135:332–335. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dreimüller N, Schlicht KF, Wagner S, Peetz
D, Borysenko L, Hiemke C, Lieb K and Tadić A: Early reactions of
brain-derived neurotrophic factor in plasma (pBDNF) and outcome to
acute antidepressant treatment in patients with Major Depression.
Neuropharmacology. 62:264–269. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang L, Zhang Z, Sun D, Xu Z, Yuan Y,
Zhang X and Li L: Low serum BDNF may indicate the development of
PSD in patients with acute ischemic stroke. Int J Geriatr
Psychiatry. 26:495–502. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Allaman I, Papp M, Kraftsik R, Fiumelli H,
Magistretti PJ and Martin JL: Expression of brain-derived
neurotrophic factor is not modulated by chronic mild stress in the
rat hippocampus and amygdala. Pharmacol Rep. 60:1001–1007.
2008.PubMed/NCBI
|
|
23
|
RomanoTorres M and Fernández-Guasti A:
Estradiol valerate elicits antidepressant-like effects in
middle-aged female rats under chronic mild stress. Behav Pharmacol.
21:104–111. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Herrick SP, Waters EM, Drake CT, McEwen BS
and Milner TA: Extranuclear estrogen receptor beta immunoreactivity
is on doublecortin-containing cells in the adult and neonatal rat
dentate gyrus. Brain Res. 1121:46–58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Walf AA, Rhodes ME and Frye CA:
Antidepressant effects of ERbeta-selective estrogen receptor
modulators in the forced swim test. Pharmacol Biochem Behav.
78:523–529. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tanapat P, Hastings NB, Reeves AJ and
Gould E: Estrogen stimulates a transient increase in the number of
new neurons in the dentate gyrus of the adult female rat. J
Neurosci. 19:5792–5801. 1999.PubMed/NCBI
|
|
27
|
Crispino M, Stone DJ, Wei M, Anderson CP,
Tocco G, Finch CE and Baudry M: Variations of synaptotagmin I,
synaptotagmin IV, and synaptophysin mRNA levels in rat hippocampus
during the estrous cycle. Exp Neurol. 159:574–583. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li XH, Liu NB, Zhang MH, Zhou YL, Liao JW,
Liu XQ and Chen HW: Effects of chronic multiple stress on learning
and memory and the expression of Fyn, BDNF, TrkB in the hippocampus
of rats. Chin Med J (Engl). 120:669–674. 2007.PubMed/NCBI
|
|
29
|
Sandstrom NJ and Williams CL: Spatial
memory retention is enhanced by acute and continuous estradiol
replacement. Horm Behav. 45:128–135. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu F, Day M, Muñiz LC, Bitran D, Arias R,
Revilla-Sanchez R, Grauer S, Zhang G, Kelley C, Pulito V, et al:
Activation of estrogen receptor-beta regulates hippocampal synaptic
plasticity and improves memory. Nat Neurosci. 11:334–343. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao H, Mayhan WG and Sun H: A modified
suture technique produces consistent cerebral infarction in rats.
Brain Res. 1246:158–166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bederson JB, Pitts LH, Tsuji M, Nishimura
MC, Davis RL and Bartkowski H: Rat middle cerebral artery
occlusion: Evaluation of the model and development of a neurologic
examination. Stroke. 17:472–476. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Herrmann N, Black SE, Lawrence J, Szekely
C and Szalai JP: The Sunnybrook Stroke Study: A prospective study
of depressive symptoms and functional outcome. Stroke. 29:618–624.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Berg A, Palomäki H, Lehtihalmes M,
Lönnqvist J and Kaste M: Poststroke depression: An 18-month
follow-up. Stroke. 34:138–143. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Suzuki S, Brown CM and Wise PM:
Neuroprotective effects of estrogens following ischemic stroke.
Front Neuroendocrinol. 30:201–211. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mazzucco CA, Lieblich SE, Bingham BI,
Williamson MA, Viau V and Galea LA: Both estrogen receptor alpha
and estrogen receptor beta agonists enhance cell proliferation in
the dentate gyrus of adult female rats. Neuroscience.
141:1793–1800. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Alkayed NJ, McCune SK, Crain BJ, Traystman
RJ and Hurn PD: Estrogen-enhanced expression of Bcl-2 mRNA in rat
brain after experimental stroke. FASEB J. 12:A9541998.
|
|
38
|
Bagetta G, Chiappetta O, Amantea D,
Iannone M, Rotiroti D, Costa A, Nappi G and Corasaniti MT:
Estradiol reduces cytochrome c translocation and minimizes
hippocampal damage caused by transient global ischemia in rat.
Neurosci Lett. 368:87–91. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Danton GH and Dietrich WD: Inflammatory
mechanisms after ischemia and stroke. J Neuropathol Exp Neurol.
62:127–136. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sairanen M, Lucas G, Ernfors P, Castrén M
and Castrén E: Brain-derived neurotrophic factor and antidepressant
drugs have different but coordinated effects on neuronal turnover,
proliferation, and survival in the adult dentate gyrus. J Neurosci.
25:1089–1094. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhou J, Zhang H, Cohen RS and Pandey SC:
Effects of estrogen treatment on expression of brain-derived
neurotrophic factor and cAMP response element-binding protein
expression and phosphorylation in rat amygdaloid and hippocampal
structures. Neuroendocrinology. 81:294–310. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang LC, Zhang QG, Zhou CF, Yang F, Zhang
YD, Wang RM and Brann DW: Extranuclear estrogen receptors mediate
the neuroprotective effects of estrogen in the rat hippocampus.
PLoS One. 5:e98512010. View Article : Google Scholar : PubMed/NCBI
|