Introduction
Gastric cancer (GC) is associated with high
morbidity and mortality rates. It is fourth most common type of
cancer and one of the leading causes of cancer-associated mortality
around the world (1–3). Based on the GLOBOCAN 2012 data from the
World Health Organization (http://globocan.iarc.fr/Pages/fact_sheets_cancer.aspx),
~1 million new cases of stomach cancer have occurred in 2012
(952,000 cases and 6.8% of the total cancer) with 70% of cases
occurring in developing countries, particularly in Eastern Asia.
Half the world total occurred in Eastern Asia, which made it the
fifth most common malignancy in the world. Stomach cancer is also
the third leading cause of cancer death in both genders worldwide
(723,000 deaths and 8.8% of the total cancer). GC is a
multifactorial disease, with environmental and genetic factors
contributing to its etiology (4,5).
Although advancements have been achieved in the treatment of GC in
recent years, GC is still associated with a poor prognosis,
particularly due to delayed diagnosis (6).
MicroRNAs (miRNAs) are ~22-nucleotide long molecules
that regulate the expression of mRNAs by base pairing to their
3′-untranslated region (3′-UTR), thereby preventing translation
(7,8). A great number of studies have indicated
that miRNAs are critical in numerous biological processes,
including apoptosis, proliferation, insulin secretion,
tumorigenesis and cellular differentiation (7,8). In
humans, >700 miRNAs have been identified, and these miRNAs
regulate the expression of 30% of protein-coding genes (9). In fact, miRNAs target nucleotides in
the ‘seed region’ of the 3′-UTR (2–8 nucleotides in the 5′ end) of
the mRNA. Perfect complementarity between the miRNA and its target
mRNA sequence leads to RNA silencing, and thus, the protein levels
are reduced (10,11). It has been demonstrated that single
nucleotide polymorphisms (SNPs) in the 3′-UTR alter target gene
expression, affecting the risk of cancer development in individuals
(12,13).
A study by Yu et al (14) identified 12 SNPs associated with
cancer development risk, which were located in the miRNA target
site. These SNPs were then genotyped in healthy subjects from the
Hebei area in China, and 6 SNPs were excluded (14) due to presenting a minor allele
frequency of <5%. The SET8 expression have been proven to be
associated with the outcome of gastric cancer (15). The potential value of one of these
SNPs, rs16917496, in the SET8 gene was examined in a preliminary
study to evaluate the correlation between rs16917496 polymorphisms
and SET8 expression in GC patients (unpublished data). Thus, the
present study aimed to investigate the remaining 5 miRNA
binding-site SNPs in the 3′-UTR of RYR3 (rs1044129),
C14orf101 (rs4901706), KIAA0423 (rs1053667),
GOLGA7 (rs11337) and KRT81 (rs3660) in GC patients in
order to assess their association with the risk of cancer
development.
Materials and methods
Tissue specimen collection and DNA
extraction
Blood samples were collected before GC resection
from a total of 153 GC patients (112 males and 41 females) who
underwent GC resection at the Department of General Surgery at the
Fourth Hospital of Hebei Medical University (Shijiazhuang, China)
prior to the operation between February 2008 and November 2010. All
GC patients were diagnosed preoperatively via histopathological
examination (16). The TNM system
was used to determine the different lymph node metastasis and
clinical stages (17). In addition,
blood samples were collected from 233 healthy subjects (169 males,
64 females) with no previous history of cancer. The characteristics
of patients and controls were listed in Table I. Patients provided their informed
consent for participation in the present study. All procedures were
supervised and approved by the Human Tissue Research Committee of
the Fourth Hospital of Hebei Medical University (Shijiazhuang,
China).
 | Table I.Clinical characteristics of gastric
cancer patients and healthy controls. |
Table I.
Clinical characteristics of gastric
cancer patients and healthy controls.
| Characteristic | Cases, n | Controls, n | P-value |
|---|
| Gender |
|
| 0.885 |
| Male | 112 | 169 |
|
|
Female | 41 | 64 |
|
| Age (years) |
|
| 0.209 |
| ≤60 | 61 | 108 |
|
|
>60 | 92 | 125 |
|
| Tumor size
(diameter) |
|
|
|
| ≤6
cm | 76 |
|
|
| >6
cm | 77 |
|
|
| Tumor location |
|
|
|
|
Upper | 55 |
|
|
|
Middle | 47 |
|
|
|
Lower | 51 |
|
|
| Lymph node
metastasis |
|
|
|
| N0 | 43 |
|
|
| N1 | 36 |
|
|
| N2 | 34 |
|
|
| N3 | 40 |
|
|
| Clinical stages |
|
|
|
| I+II | 46 |
|
|
|
III+IV | 90 |
|
|
| Extent of
differentiation |
|
|
|
|
Moderate | 96 |
|
|
|
Poor | 57 |
|
|
| Pathological
subtype |
|
|
|
|
Diffuse | 100 |
|
|
|
Intestinal | 27 |
|
|
|
Mixed | 26 |
|
|
Genomic DNA was extracted using the Wizard Genomic
DNA Purification Kit (A1125; Promega Corp., Madison, WI, USA).
Briefly, 100 µl blood was added to the cell lysis solution for red
blood cells, followed by lysis of the white blood cells and their
nuclei in the Nuclei Lysis Solution with RNase. The cellular
proteins were then removed by a salt-precipitation step. Finally,
the genomic DNA was concentrated and desalted by isopropanol
precipitation.
SNP genotyping of microRNA
binding-site SNPs
The miRNA SNPs located at the binding site,
including RYR3 (rs1044129), C14orf101 (rs4901706),
KIAA0423 (rs1053667), GOLGA7 (rs11337) and
KRT81 (rs3660), were genotyped using the ligation detection
reaction method (18), featuring
forward and reverse primers, in order to amplify DNA fragments
flanking the SNPs. This procedure was performed according to the
SNP database of the National Center for Biotechnology Information
(Bethesda, MD, USA; http://www.ncbi.nlm.nih.gov/snp/).
Polymerase chain reaction (PCR) was performed with
50 ng genomic DNA, 1 µl of 10 nm primer pairs, 12.5 µl of Master
Mix and distilled water to a final volume of 25 µl using a PCR
Master Mix kit (K1081; Promega Corp.), according to the
manufacturer's instructions, and for 35 thermal cycles. The primer
and probe sequences used in PCR are shown in Table II. Ligation was performed with
various probes, which were matched to the SNPs. Subsequently, the
ABI PRISM 3730xl DNA Analyzer (Applied Biosystems; Thermo Fisher
Scientific, Inc., Foster City, CA, USA) was used to separate the
ligated products. SNPs were detected and verified based on
differences in the length of ligated products. The experiment was
performed once if it was successful.
| Table II.Primers and probes used for
genotyping of miRNA SNPs. |
Table II.
Primers and probes used for
genotyping of miRNA SNPs.
| Gene | rs NCBI | Primer
sequence | Probe sequence |
|---|
| RYR3 | rs1044129 | F:
ATGGAGTAATGCTTTATGGTC | S1:
TTTAGGTGAATCTCCTCAAATACAA |
|
| (A/G) | R:
CAGTCACAGAGTGGTTGTAGA | S2:
TTTTTTAGGTGAATCTCCTCAAATACAG |
|
|
|
| S3:
TGAAGTGCCCACTGCAATAAAGTAA |
|
KIAA0423 | rs1053667 | F:
CATGAAATCTGAGTCACATGG | S1:
TTTTAGTAATCATGTTTTAATGTAGAACC |
|
| (C/T) | R:
GCTGAGAAATGAGACATACCA | S2:
TTTTTTTAGTAATCATGTTTTAATGTAGAA |
|
|
|
| S3:
TCAAACAGGATGGAACATCAGTGGATTT |
|
C14orf101 | rs4901706 | F:
AAACTAAGTCATCTCCCAGATA | S1:
TTTTTTTTTTAATGGGGTATTCAGTGACTAAGA |
|
| (A/G) | R:
GTCATCTGGTGAAAGACTGGA | S2:
TTTTTTTTTTTTTAATGGGGTATTCAGTGACTAAGG |
|
|
|
| S3:
TCTGCTATTTATGCAAAATTCTGTTTTTTTT |
| GOLGA7 | rs11337 | F:
CGCTGTATTTGGGAGAGAGTT | S1:
TTTTTTTTTTTTACCATTAAAAGTTTCACTGTCAGAG |
|
| (G/T) | R:
CAGGCTGTAAAGTAACAAATGAG | S2:
TTTTTTTTTTTTTTTACCATTAAAAGTTTCACTGTCAGAT |
|
|
|
| S3:
ATATTGTAGGTGCTAATACTGGATTTTTTTTTTT |
| KRT81 | rs3660 | F:
GTTAGGCACCCCAACTCAAGT | S1:
TTTTTATTTTTGAGAAAAGTCCTGCTCAC |
|
| (G/C) | R:
GCCAGCGGACTTCTTTCTAGG | S2:
TTTTTTTTATTTTTGAGAAAAGTCCTGCTCAT |
|
|
|
| S3:
TTGCACTATTCTATAGAAACTACAATTT |
Renilla/luciferase reporter
assays
A total of 4 oligonucleotides were synthesized based
on dbSNP, the NCBI database of genetic variation, which consisted
of the following parts (from 5′ to 3′): A XhoI sticky end (5
bp), a fragment from the 3′-UTR of the C14orf101 gene
containing the GG or AA genotype (rs4901706; 47 bp), and a
NotI sticky end (2 bp). The following sequences were used:
GG-containing sense,
5′-TCGAGAGTGCTCAGCTACTTCTCCTCCACTTTGAAAGACCCCTCCCAGATCTGC-3′, and
antisense,
5′-GGCCGCAGATCTGGGAGGGGTCTTTCAAAGTGGAGGAGAAGTAGCTGAGCACTC-3′;
AA-containing sense,
5′-TCGAGAGTGCTCAGCTACTTCTCCTGCACTTTGAAAGACCCCTCCCAGATCTGC-3′, and
antisense,
5′-GGCCGCAGATCTGGGAGGGGTCTTTCAAAGTGCAGGAGAAGTAGCTGAGCACTC-3′. The
four oligonucleotides were incubated for 5 min with 1X NEBuffer 2
(New England Biolabs, Ipswich, MA, USA) in a heating block at 95°C.
Next, the temperature was gradually reduced until it reached room
temperature. The psiCheck-2 vector featuring the
Renilla/luciferase and controlled firefly luciferase genes
was linearized by digestion with NotI and XhoI (New
England Biolabs), and the vector was then purified using a 1%
agarose gel electrophoresis. The oligonucleotides were ligated in
the linearized psiCheck-2 vector (Promega Corp.) into the cloning
sites (NotI and XhoI), which were downstream of the
Renilla luciferase reporter gene with T4 DNA ligase (Promega
Corp.). Subsequently, the ligated vectors were transformed in
Escherichia coli-competent cells, as follows: Competent
cells were taken from −80°C freezer and thawed on ice for 20 min. 1
µl of ligated vector was mixed into 100 µl of competent cells in a
microcentrifuge. The competent cell/DNA mixture was then placed on
ice for 20–30 min and each transformation tube was heat shocked by
placing the tube into a 42°C water bath for 90 sec. The tubes were
placed back on the ice for 2 min and 500 µl of Luria-Bertani (LB)
medium was added. The cells were grown at 37°C in a shaking
incubator for 45 min. All of the transformations were plated onto a
10 cm LB agar plate and the plates were incubated at 37°C
overnight. A total of 3 bacterial colonies were selected and the
bacterial culture was incubated at 37°C for 12–18 h in a shaking
incubator. The bacterial fluid samples were sequenced by the
methods of Sanger dideoxy (Sangon Biotech, Shanghai, China), and
sequencing was used to identify and select positive clones.
A human GC cell line (SGC 7901) purchased from the
Shanghai Institute of Biochemistry and Cell Biology (Shanghai,
China) was seeded in 48-well plates and transfected with the
modified psiCheck-2 vector (800 ng) using Lipo 2000 (11668-027;
Invitrogen, CA, USA) at 37°C for 4–6 h with the GG or AA genotype.
At 48 h after transfection, the Renilla luciferase activity
was assessed using the Dual-Lucy Assay kit (Vigorous Instrument
Co., Ltd., Beijing, China) and a luminometer BioFix Lumi-10
(Macherey-Nagel, Dusseldorf, Germany). The transfection efficiency
was normalized against the firefly luciferase activity.
Statistical analysis
The χ2 test was performed to analyze
dichotomous variables, including the presence or absence of an
individual SNP in the patients and healthy controls. The odds ratio
(OR) and 95% confidence interval (CI) were calculated using an
unconditional logistic regression model. In addition, Student's
t-test was used to compare the differential expression levels
between genotypic groups in the Renilla/luciferase reporter
assays. SPSS version 18.0 software (SPSS, Inc., Chicago, IL, USA)
was used to perform statistical analyses. Statistically significant
differences were defined when the P-value was <0.05.
Results
Patient characteristics
A total of 153 patients and 233 healthy controls
were included in the present study. The clinical characteristics of
patients including gender, age, tumor size and location,
metastasis, clinical stage, differentiation status and pathological
types were listed in Table I. The
characteristics of the control, including gender and age were also
listed in Table I. The distribution
frequency was not identified to be different between patients and
controls with respect to their age and gender (P=0.885 and 0.209)
respectively. In addition, the mean age between patients and
controls was not statistically different. An analysis for these
patients and the controls was subsequently performed.
Association of C14orf101 SNP with risk
of GC
In the present study, miRNA binding-site SNPs were
genotyped in the patients and healthy controls, including
C14orf101 (rs4901706), RYR3 (rs1044129),
GOLGA7 (rs11337), KRT81 (rs3660) and KIAA0423
(rs1053667) to evaluate their association with the risk for cancer.
The cancer risk association was evaluated for the distribution
frequency of rs4901706 (GG vs. AG+AA), rs1044129 (AA vs. AG+GG),
rs11337 (GG vs. GT+TT), rs3660 (GG vs. CG+CC) and rs1053667 (TT vs.
CT+CC) between patients and controls using the χ2 test.
As shown in Table III, the
rs1044129 AA carriers were 47 in GC patients and 67 in the controls
whereas the AG+GG carriers were 106 in GC patients and 166 in the
controls; the rs1053667 TT carriers were 113 in GC patients and 161
in the controls whereas the CT+TT carriers were 40 in GC patients
and 72 in the controls. In addition, the rs11337 GG carriers were
92 in GC patients and 141 in the controls while the GT+TT carriers
were 61 in GC patients and 92 in the controls; Finally, the rs3660
GG carriers were 97 in GC patients and 145 in the controls while
CG+CC carriers were 56 in GC patients and 88 in the controls. The
rs 1044129 (relative risk, 1.099; 95% CI, 0.704–1.715; P=0.679),
rs11337 (relative risk, 0.984; 95% CI, 0.649–1.493; P=0.940),
rs3660 (relative risk, 1.023; 95% CI, 0.662–1.583; P=0.917) and
rs1053667 (relative risk, 1.263; 95% CI, 0.801–1.992; P=0.314) were
not associated with cancer risk by the analysis of the present
study. The GG carriers were 100 in GC patients and 125 in the
controls whereas the AG+AA were 53 in GC patients and 108 in the
controls. It was also noted that the GG genotype was susceptible to
GC carcinogenesis (relative risk, 1.630; 95% CI, 1.070–2.483;
P=0.022). The data of the present study demonstrated that the
rs4901706 SNP of C14orf101 was a predictive marker for GC
risk.
| Table III.Distribution frequency of
miRNA-binding SNPs between GC patients and healthy controls. |
Table III.
Distribution frequency of
miRNA-binding SNPs between GC patients and healthy controls.
| Variant | Gene | Genotype | Cases | Controls | P-value | OR | 95% CI |
|---|
| rs1044129 | RYR3 | AA | 47 | 67 | 0.679 | 1.099 | 0.704–1.715 |
|
|
| AG+GG | 106 | 166 |
|
|
|
| rs1053667 |
KIAA0423 | TT | 113 | 161 | 0.314 | 1.263 | 0.801–1.992 |
|
|
| CT+CC | 40 | 72 |
|
|
|
| rs4901706 |
C14orf101 | GG | 100 | 125 | 0.022 | 1.630 | 1.070–2.483 |
|
|
| AG+AA | 53 | 108 |
|
|
|
| rs11337 | GOLGA7 | GG | 92 | 141 | 0.940 | 0.984 | 0.649–1.493 |
|
|
| GT+TT | 61 | 92 |
|
|
|
| rs3660 | KRT81 | GG | 97 | 145 | 0.917 | 1.023 | 0.662–1.583 |
|
|
| CG+CC | 56 | 88 |
|
|
|
rs4901706 SNP in C14orf101 affects
protein translation in GC cells
A vector named psiCheck-2 was constructed containing
the rs4901706 AA or GG genotypes downstream of the
Renilla/luciferase reporter gene in order to assess their
functional effect on C14orf101 expression. The vector was
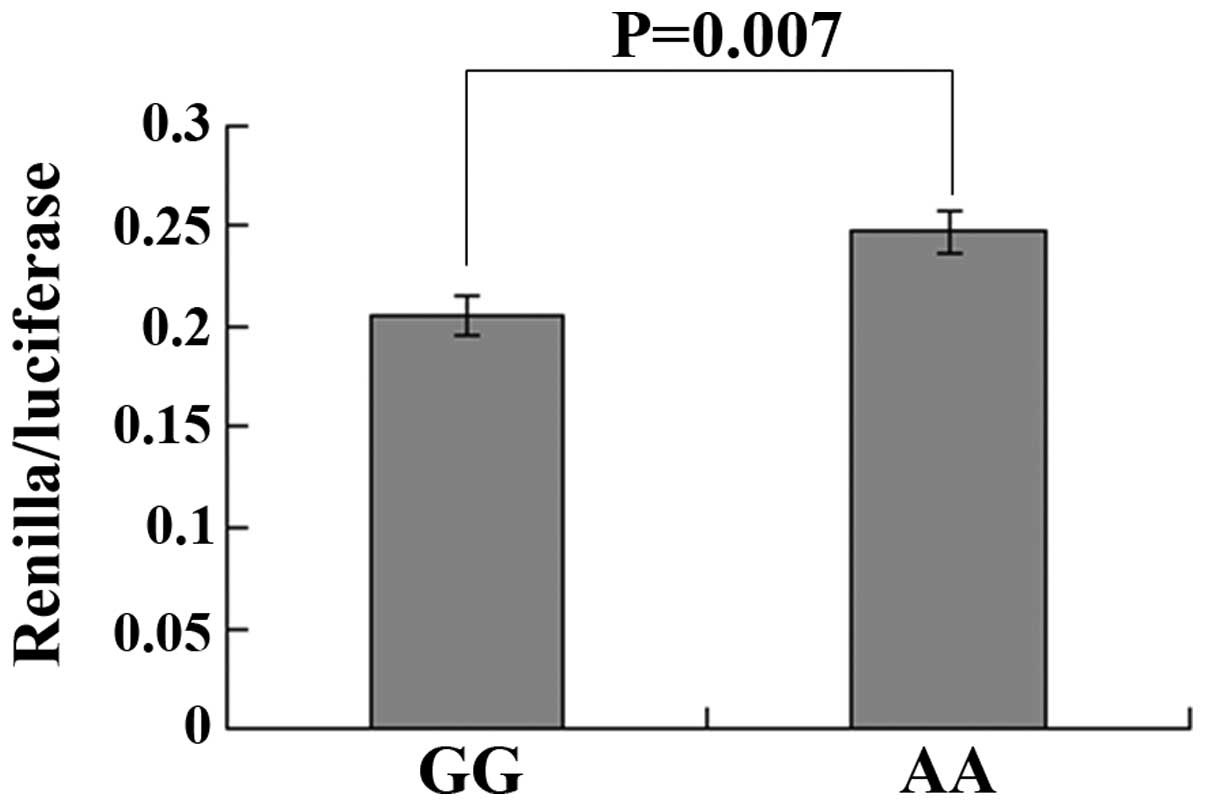
transfected into the GC cell line SGC 7901. As shown in Fig. 1, a significant reduction of
Renilla/luciferase activity was observed in GG genotypes
compared with that of the AA genotypes (P=0.007). These data
demonstrated that rs4901706 SNP may change the binding affinity
between the rs4901706 genotype at the 3′UTR of C14orf101 and
the corresponding miRNA.
Discussion
In the present study, the SNPs of miRNA binding
sites, including RYR3 (rs1044129), C14orf101
(rs4901706), KIAA0423 (rs1053667), GOLGA7 (rs11337)
and KRT81 (rs3660), were examined for their ability to
predict GC cancer risk. The results revealed that the rs4901706 SNP
of C14orf101 gene is a possible risk biomarker for GC. To
the best of our knowledge, the current study is the first to
investigate the miRNA binding-site SNPs at the 3′-UTR of
C14orf101, which exhibit the ability to predict GC risk. The
miR-SNPs defined as the SNP at the miRNA binding site, the miRNAs
and miRNAs processing machinery genes, are critically involved in
disease phenotypes (19,20). We have identified gastric cancer
associated miR-SNP of miRNAs processing machinery genes in previous
study (21). Yu et al
(14) performed a genome-wide
analysis of SNPs located in the miRNA-binding sites of the 3′-UTR
of various human genes and revealed that some miRNA-binding SNP
distribution frequencies between the human cancer EST libraries and
the dbSNP database was significantly different. Furthermore, they
identified that twelve miRNA-binding SNPs displayed an aberrant
allele frequency in human cancers using human cancer specimens
against the dbSNP database for case-control association studies.
Finally, they deduced that SNPs located in miRNA-binding sites was
potentially associated with cancer by affecting miRNA target
expression and function (14).
Consistent with these previous findings, the frequent allele G of
rs4901706 was found to be associated with GC risk in the present
study.
MiRNAs appear to be critical in the response of
patients to various treatments, particularly cancer treatment
(22). In addition, the rs4901706
SNP of the C14orf101 gene has been previously demonstrated
to be associated with cancer risk, since it serves as an miRNA
binding site; however, the miRNA binding to this site that
modulates the C14orf101 expression was not specified
(14,23). In a previous study, we demonstrated
that this SNP was also associated with survival in patients with
non-Hodgkin lymphoma G allele carriers that exhibited a long life
span (24). The data demonstrated
that the GG genotype of C14orf101 would modify both the
carcinogenesis and outcome of the cancer.
C14orf101, also known as transmembrane
protein 260 (TMEM260), is conserved in chimpanzees, rhesus macaque
monkeys, canines, cattle, mice, rats, chickens and zebrafish.
However, a functional study of this gene has not been reported to
date. The results of the current study suggest that the
miRNA-binding SNPs of C14orf101 have an effect on cancer
development. A Renilla/luciferase reporter assay highlighted
the different binding affinity between the SNP of rs4901706 and
miRNA binding to the rs4901706 site. Furthermore, the A to G
transition of rs4901706 in the 3′-UTR of C14orf101 may
result in the destruction of the A:T bond at the miRNA binding
site, so as to alter the affinity of C14orf101 in binding
with miRNAs. This may modulate the expression of the gene, thereby
initiating GC carcinogenesis. However, the current study should be
replicated in other populations and in laboratory-based functional
studies in order to validate the results. The miRNA binding to
C14orf101 should be identified, and the change in the expression of
C14orf101 on proliferation, invasion and apoptosis of gastric
cancer cells should be evaluated in future studies.
In conclusion, SNPs in the C14orf101 miRNA
binding site appeared to represent a biomarker for cancer risk.
rs4901706 SNP was a potential maker for early diagnosis of gastric
cancer and valuable for gastric cancer prevention. Further
analyzing the genetic polymorphisms of miRNA binding sites may
result in better identification of patients susceptible to GC
development. The miRNA binding to C14orf101 should be
identified as well as the role of the expressional change of
C14orf101 on proliferation, invasion and apoptosis of
gastric cancer cells in future studies.
Acknowledgements
This study was supported by a grant from the Key
Basic Research Program of Hebei (grant no. 14967713D).
References
|
1
|
Parkin DM: International variation.
Oncogene. 23:6329–6340. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Bray FI and Devesa SS: Cancer
burden in the year 2000. The global picture. Eur J Cancer. 37(Suppl
8): S4–S66. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stadtländer CT and Waterbor JW: Molecular
epidemiology, pathogenesis and prevention of gastric cancer.
Carcinogenesis. 20:2195–2208. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crew KD and Neugut AI: Epidemiology of
gastric cancer. World J Gastroenterol. 12:354–362. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Forman D and Burley VJ: Gastric cancer:
Global pattern of the disease and an overview of environmental risk
factors. Best Pract Res Clin Gastroenterol. 20:633–649. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xia J, Guo X, Yan J and Deng K: The role
of miR-148a in gastric cancer. J Cancer Res Clin Oncol.
140:1451–1456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicateds
that thousands of human genes are microRNA targets. Cell.
120:15–20. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zeng Y, Yi R and Cullen BR: MicroRNAs and
small interfering RNAs can inhibit mRNA expression by similar
mechanisms. Proc Natl Acad Sci USA. 100:9779–9784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeng Y, Wagner EJ and Cullen BR: Both
natural and designed micro RNAs can inhibit the expression of
cognate mRNAs when expressed in human cells. Mol Cell. 9:1327–1333.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chin LJ, Ratner E, Leng S, Zhai R, Nallur
S, Babar I, Muller RU, Straka E, Su L, Burki EA, et al: A SNP in a
let-7 microRNA complementary site in the KRAS 3′ untranslated
region increases non-small cell lung cancer risk. Cancer Res.
68:8535–8540. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brendle A, Lei H, Brandt A, Johansson R,
Enquist K, Henriksson R, Hemminki K, Lenner P and Försti A:
Polymorphisms in predicted microRNA-binding sites in integrin genes
and breast cancer: ITGB4 as prognostic marker. Carcinogenesis.
29:1394–1399. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu Z, Li Z, Jolicoeur N, Zhang L, Fortin
Y, Wang E, Wu M and Shen SH: Aberrant allele frequencies of the
SNPs located in microRNA target sites are potentially associated
with human cancers. Nucleic Acids Res. 35:4535–4541. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shi XL, Guo ZJ, Wang XL, Liu XL and Shi
GF: SET8 expression is associated with overall survival in gastric
cancer. Genet Mol Res. 14:15609–15615. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kaneko S and Yoshimura T: Time trend
analysis of gastric cancer incidence in Japan by histological
types, 1975–1989. Br J Cancer. 84:400–405. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Washington K: 7th edition of the AJCC
cancer staging manual: Stomach. Ann Surg Oncol. 17:3077–3079. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yi P, Lu W, Guo J, Liu Q, Chen Z, Han J
and Li L: Development of a PCR/ligase detection
reaction/nanogold-based universal array approach for the detection
of low-abundant DNA point mutations. Cell Biochem Biophys.
61:629–636. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Horikawa Y, Wood CG, Yang H, Zhao H, Ye Y,
Gu J, Lin J, Habuchi T and Wu X: Single nucleotide polymorphisms of
microRNA machinery genes modify the risk of renal cell carcinoma.
Clin Cancer Res. 14:7956–7962. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu Z, Chen J, Tian T, Zhou X, Gu H, Xu L,
Zeng Y, Miao R, Jin G, Ma H, et al: Genetic variants of miRNA
sequences and non-small cell lung cancer survival. J Clin Invest.
118:2600–2608. 2008.PubMed/NCBI
|
|
21
|
Xie Y, Wang Y, Zhao Y and Guo Z:
Single-nucleotide polymorphisms of microRNA processing machinery
genes are associated with risk for gastric cancer. Onco Targets
Ther. 8:567–571. 2015.PubMed/NCBI
|
|
22
|
Iorio MV, Ferracin M, Liu CG, Veronese A,
Spizzo R, Sabbioni S, Magri E, Pedriali M, Fabbri M, Campiglio M,
et al: MicroRNA gene expression deregulation in human breast
cancer. Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ryan BM, Robles AI and Harris CC: Genetic
variation in microRNA networks: The implications for cancer
research. Nat Rev Cancer. 10:389–402. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang B, Liu C, Diao L, Wang C and Guo Z: A
polymorphism at the microRNA binding site in the 3′ untranslated
region of C14orf101 is associated with non-Hodgkin lymphoma overall
survival. Cancer Genet. 207:141–146. 2014. View Article : Google Scholar : PubMed/NCBI
|