Introduction
Angiogenin, also known as ribonuclease 5, is a 14
kDa protein expressed in various tissues that can induce or inhibit
target protein synthesis. Under cellular growth conditions
angiogenin is translocated into the nucleus where it enhances
ribosomal RNA (rRNA) transcription and thus subsequent ribosome
formation and protein synthesis required for cell growth and
proliferation (1,2). Under cellular stress conditions,
angiogenin is sequestered in the cytoplasm, where it cleaves
transfer RNA (tRNA) to produce tRNA-derived, stress-induced small
RNAs (tiRNAs). In turn, tiRNAs repress global translation by
interfering with translation initiation factors, but specifically
facilitate the translation of mRNA that contains internal ribosomal
entry sites (IRES) (3–5). Since numerous IRES-containing mRNAs
encode pro-survival and anti-apoptotic genes, angiogenin protects
cells from apoptosis during cell stress (6,7). In
addition, the direct interaction of angiogenin with p53 (8), and interaction between tiRNAs and
cytochrome c released from the mitochondria (9), have been detected to inhibit cell
apoptosis.
Interestingly, angiogenin may serve a role in the
immune response. Angiogenin is expressed in numerous malignant and
normal cells, including lymphocytes (10). Angiogenin levels are increased in
inflammatory conditions (11,12) and
autoimmune diseases, where it has been detected in the synovial
fluid of patients with rheumatoid arthritis (13). During the adaptive immune response,
the expansion phase (rapid T-cell proliferation) is followed by the
contraction phase (T-cell apoptosis). Insufficient apoptosis of
activated T-cells can lead to autoimmune diseases, whereas
excessive apoptosis can result in immunodeficiency (14). Although angiogenin has been
implicated in cell proliferation and apoptosis, its role in the
T-cell response has not been well studied.
In the present study, the two-way mixed lymphocyte
reaction (MLR) was used as a measure of alloreactivity (15), along with neamine that inhibits the
nuclear translocation of angiogenin. Neamine is a neomycin
derivative with low toxicity, which represses rRNA transcription
and cell proliferation through inhibiting angiogenin nuclear
translocation (2,16–18). In
addition, nuclear translocation of angiogenin may be required for
its ribonuclease activity. In S28N mutant angiogenin that is found
in some patients with amyotrophic lateral sclerosis, the mutation
is near to the nuclear localization sequence and inhibits nuclear
translocation and tiRNA production (19).
Materials and methods
Study participants
Blood samples were collected on the same day in
February 2016 from 5 healthy volunteers (3 men and 2 women; mean
age, 36±8 years) at the University Hospital of Larissa (Larissa,
Greece). Informed consent was obtained from each participant
enrolled in the study and the Ethics Committee of the University
Hospital of Larissa (Larissa, Greece) approved the study
protocol.
Cell culture conditions and two-way
MLRs
Peripheral blood mononuclear cells (PBMC) were
isolated from whole blood samples by Ficoll-Hypaque density
gradient centrifugation using Histopaque-1077 (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany). Briefly, the interface was
collected and washed with Roswell Park Memorial Institute
(RPMI)-1640 medium (Sigma-Aldrich; Merck Millipore). Then, isolated
PBMCs were counted using a Neubauer chamber (Paul Marienfeld GmbH,
Lauda-Königshofen, Germany) on an optical microscope. Cell
viability was assessed using the trypan blue exclusion assay
(Sigma-Aldrich; Merck Millipore). Cells were cultured in RPMI-1640
medium, supplemented with L-glutamine, 10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 10%
fetal bovine serum (Sigma-Aldrich; Merck Millipore) and
antibiotic-antimycotic solution (Sigma-Aldrich; Merck Millipore).
Cultures were incubated at 37°C in an atmosphere of 95% relative
humidity and 5% CO2.
To determine cell proliferation and neamine
cytotoxicity, 10 two-way MLRs were performed in the presence or
absence of 100 µM neamine (angiogenin nuclear translocation
inhibitor; Sigma-Aldrich; Merck Millipore). This concentration was
chosen as it was in line with previous experiments (17,18).
MLRs were performed in 96-well plates over 7 days. The quantity of
PBMCs, from each of the two individuals that contribute to the
formation of an MLR couple, was 5×104 cells measuring
1×105 cells per well. Resting (unstimulated) PMBC
cultures of 1×105 cells per well from each member of the
MLR couple were used as controls.
To assess angiogenin concentration in the
supernatant of the MLRs and expression of specific proteins
(described below) in cluster of differentiation 4 positive
(CD4+) T-cells, 10 two-way MLRs were performed in
12-well plates over 7 days. The number of PBMCs, from each of the
two individuals that contribute to the formation of an MLR couple,
was 5×105 cells, measuring 1×106 PBMCs per
well. Resting PMBC cultures of 1×106 cells per well from
each member of the MLR couple were used as controls. At the end of
the 7-day period, supernatants were collected and stored at −80°C
and CD4+ T-cells were isolated by negative selection
using the Human CD4+ T Cell Isolation Kit (cat. no.
130-096-533; Miltenyi Biotec GmbH, Bergisch Gladbach, Germany).
Cell proliferation in the MLR
To assess cell proliferation the Cell Proliferation
ELISA kit (cat. no. 11647229001; Roche Diagnostics, Indianapolis,
IN, USA) was used, which is based on bromodeoxyuridine labeling and
immunoenzymatic detection. The proliferation index of cells was
calculated as the ratio of the optical density (OD) at 450 nm of
the MLR to the mean OD of the control resting PBMC cultures of the
two subjects that constituted each specific MLR. Experiments were
performed in triplicate and the results are presented as the
mean.
Neamine cytotoxicity
Neamine cytotoxicity was assessed by performing a
lactate dehydrogenase (LDH) release assay using the CytoTox
Non-Radioactive Cytotoxic Assay kit (cat. no. G1780; Promega
Corporation, Madison, WI, USA) in MLRs. To assess cytotoxicity in
resting PBMCs, the mean of measurements from the control resting
PBMC cultures of the two subjects that constituted each MLR couple
were used. Results were expressed as the percentage of dead cells.
Experiments were performed in triplicate and the results are
presented as the mean.
Angiogenin expression in the MLR
Angiogenin expression in the supernatants of MLRs
was assessed by measuring its concentration using an ELISA with a
sensitivity of <6 pg/ml (cat. no. DAN00; Human Angiogenin
Quantikine ELISA; R&D Systems, Inc., Minneapolis, MN, USA). To
assess angiogenin concentration in resting PBMCs, the mean of the
measurements derived from the control resting PBMC cultures of the
two subjects that constituted each specific MLR couple were
used.
Western blotting for expression of
specific proteins in MLR CD4+ T-cells
The expression of B-cell lymphoma 2 (Bcl-2),
Bcl-2-associated X protein (Bax), activated cleaved caspase-3 (CC3;
activated by cleavage at aspartate 175) and β-actin in
CD4+ T-cells isolated from the MLRs was assessed by
western blotting. Isolated CD4+ T-cells were counted
using an optical microscope and Neubauer chamber, and cell
viability was determined by the trypan blue exclusion assay
(Sigma-Aldrich; Merck Millipore). Equal numbers of CD4+
T-cells from each MLR were lysed using the T-PER tissue protein
extraction reagent (Thermo Fisher Scientific, Inc., Rockford, IL,
USA), supplemented with protease inhibitors
[4-(2-aminoethyl)benzenesulfonyl fluoride, E-64, bestatin,
leupeptin, aprotinin, phenylmethanesulfonyl fluoride and
ethylenediaminetetraacetic acid] and phosphatase inhibitors against
acid and alkaline phosphatases (both Sigma-Aldrich; Merck
Millipore), in addition to serine/threonine and tyrosine protein
phosphatases inhibitors (both Roche Diagnostics). Extracted protein
was quantified via the Bradford assay (reagent provided by
Sigma-Aldrich; Merck Millipore) and 10 µg from each sample was used
for SDS-PAGE (4–12% gels) and subsequent western blotting.
Polyvinylidene fluoride (PVDF) blots were incubated at 4°C with a
primary antibody [rabbit monoclonal anti-Bcl-2 (cat. no. 4223;
1:1,000), rabbit monoclonal anti-Bax (cat. no. 5023; 1:1,000),
rabbit monoclonal anti-caspase-3 (activated by cleavage at
aspartate 175; cat. no. 9664; 1:1,000) or rabbit monoclonal
anti-β-actin (cat. no. 4967; 1:2,500) (all from Cell Signaling
Technology, Inc., Danvers, MA, USA)] for 16 h, followed by 3 washes
and incubation with a secondary antibody (anti-rabbit IgG,
HRP-linked antibody; cat. No. 7074; 1:1,000; Cell Signaling
Technology, Inc.) for 30 min at room temperature. The BenchMark
Pre-Stained Protein Ladder (Thermo Fisher Scientific, Inc.) was
used as a marker. Bands were then visualized by enhanced
chemiluminescent detection using the LumiSensor Plus
Chemiluminescent HRP Substrate kit (cat. no. L00225; GenScript,
Piscataway, NJ, USA). When re-probing PVDF blots, the previous
antibodies were removed using the Restore Western Blot Stripping
Buffer (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol and western blotting was resumed as
previously described, using a different primary antibody.
Statistical analysis
SPSS software (version 13.0; SPSS, Inc., Chicago,
IL, USA) was used for statistical analysis. The normality of the
evaluated variables was assessed and confirmed by one-sample
Kolmogorov-Smirnov tests. For statistical comparison of means a
paired-sample t-tests were used. Results are expressed as mean ±
standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Analysis and quantification of the westerns blots
was performed using Image J software (version 1.49; National
Institutes of Health, Bethesda, MD, USA) and results expressed as
arbitrary OD units. Statistical analysis following normalization
for the control OD values was avoided, in order to prevent
disruption of the prerequisite for normal distribution of the
compared variables when applying parametric statistical tests.
However, for the reader's convenience, the results are presented
following normalization to the control group.
Results
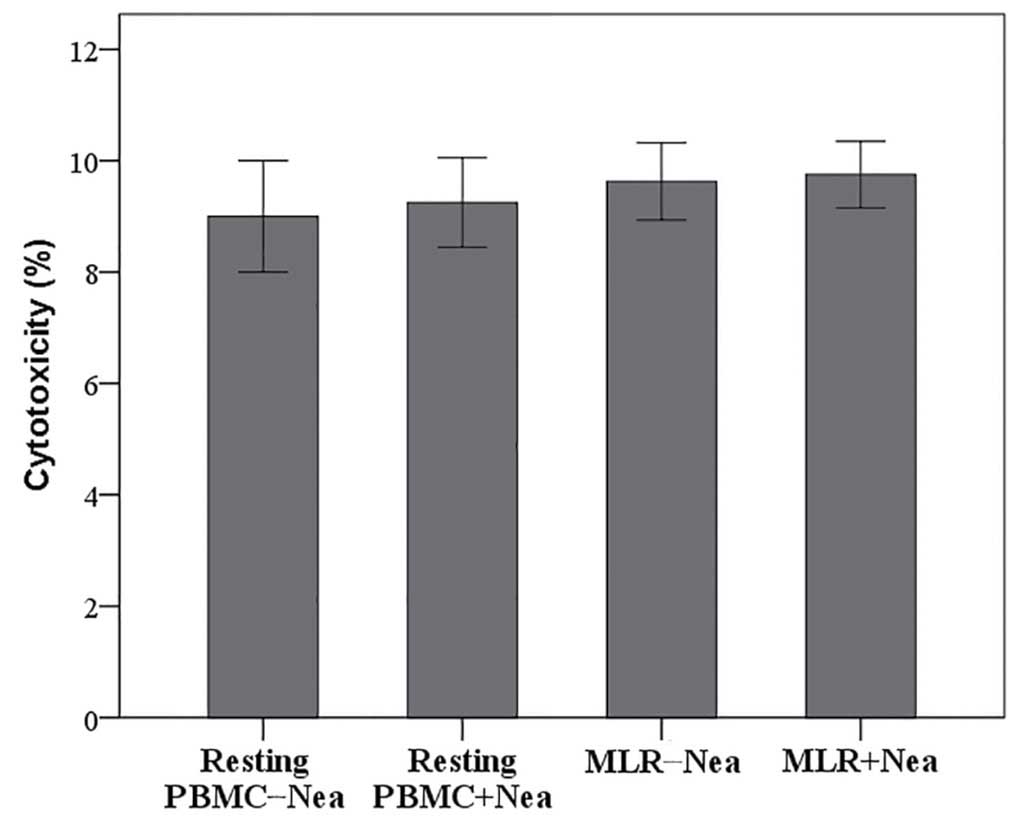
Neamine is not cytotoxic to resting
PBMCs or MLRs
The LDH release assay revealed that 100 µΜ neamine
does not induce cell necrosis in resting PBMCs or MLRs.
Cytotoxicity was 9.00±1.00% in resting PBMCs vs. 9.25±0.80% in
resting PBMCs treated with neamine (P=0.692; Fig. 1). In untreated MLRs cytotoxicity was
10.00±1.00% vs. 10.00±0.75% in neamine-treated MLRs (P=1.000;
Fig. 1). Results are presented as
the percentage of cell death.
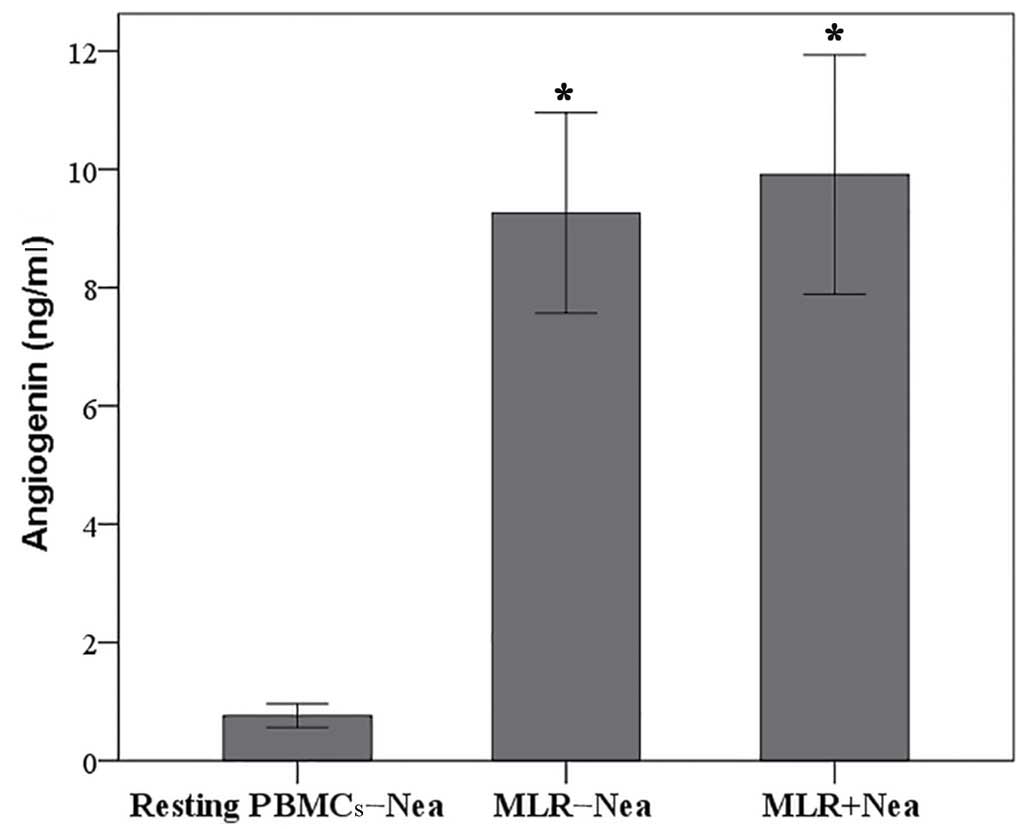
In MLRs angiogenin expression
increased significantly compared with resting PMBCs irrespective of
the presence of neamine
Compared to resting PBMCs, angiogenin concentration
was significantly increased in the supernatants of MLRs (0.76±0.20
vs. 9.26±1.69 ng/ml, respectively; P<0.001; Fig. 2). The concentration of angiogenin in
the supernatant of neamine-treated MLRs was 9.91±2.02 ng/ml,
significantly higher compared with resting PBMCs (0.76±0.20 ng/ml;
P<0.001; Fig. 2), but similar to
the concentration found in untreated MLRs (9.26±1.69 ng/ml;
P=0.070; Fig. 2).
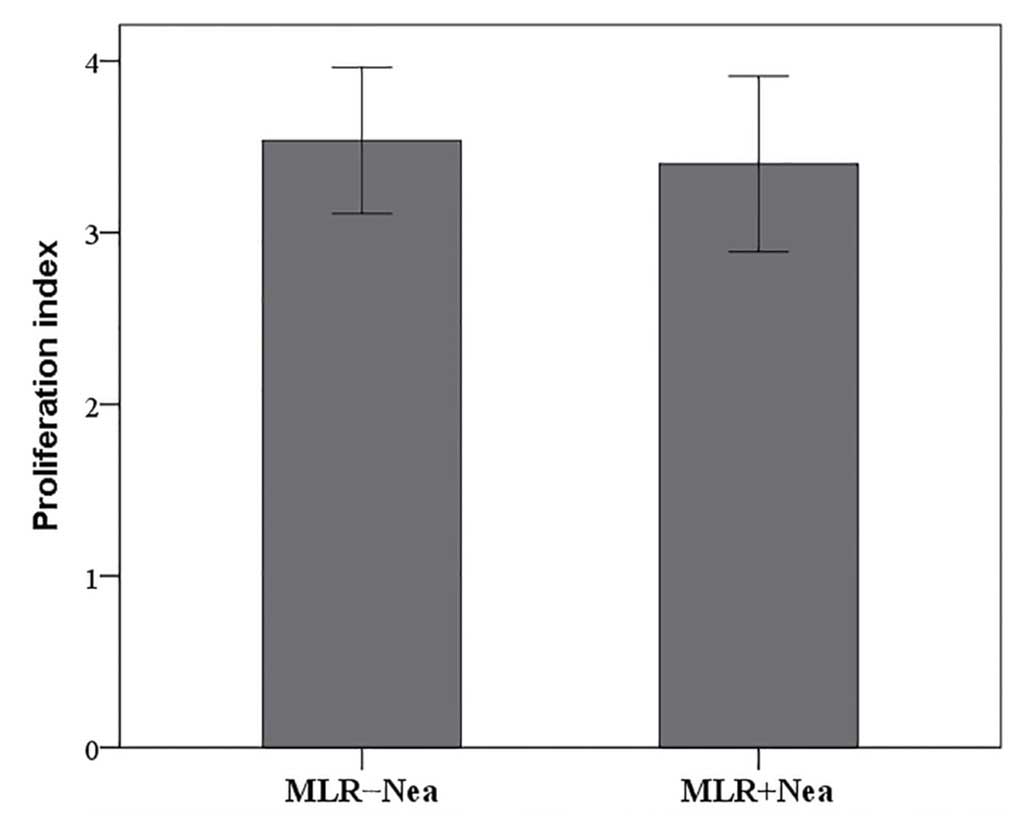
In MLRs neamine does not affect cell
proliferation
In MLRs, neamine did not significantly alter cell
proliferation. The proliferation index was 3.54±0.43 in untreated
MLRs and 3.40±0.51 in neamine-treated MLRs (P=0.097; Fig. 3).
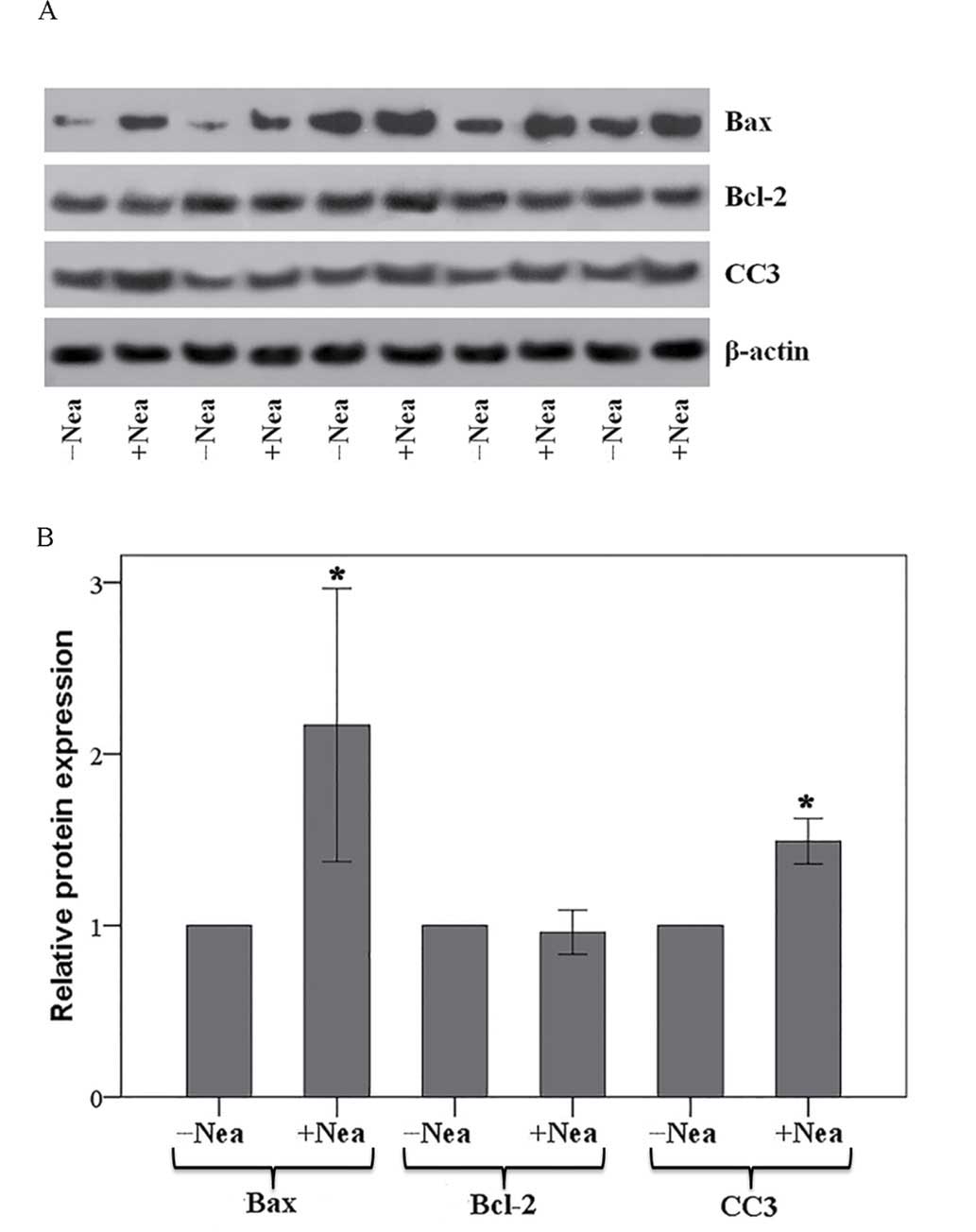
Neamine increases protein levels of
Bax and activated caspase-3 in MLR-derived CD4+
T-cells
The expression of Bcl-2, Bax, CC3 and β-actin in
CD4+ T-cells isolated from the MLRs was assessed by
means of western blotting (Fig. 4A)
and subsequent quantification of relative protein expression levels
(Fig. 4B). In CD4+
T-cells isolated from the MLRs, neamine significantly increased the
expression of pro-apoptotic Bax by a factor of 2.17±0.80
(P<0.001 vs. untreated MLR CD4+ T-cells; Fig. 4B). Conversely, neamine had no effect
on the expression of anti-apoptotic Bcl-2, altering expression by a
factor of 0.96±0.13 (P=0.397 vs. untreated MLR; Fig. 4B). Neamine significantly increased
the protein levels of CC3 by a factor of 1.49±0.13 relative to
untreated MLR CD4+ T-cells (P<0.001; Fig. 4B).
Discussion
In the present study, the effect of angiogenin on
human alloreactive CD4+ T-cell proliferation and
apoptosis was evaluated. For this purpose, the two-way MLR was used
as a measure of alloreactivity (15), along with neamine (an angiogenin
nuclear translocation inhibitor) treatment (2,16–18). At
100 µM neamine did not induce cell necrosis in resting PBMCs or
MLRs.
Interestingly, angiogenin production in the
supernatants of untreated MLRs was increased by ~1,250% compared
with resting PBMCs controls. This indicates that angiogenin
expression increases during the adaptive immune response, which
from a teleological point of view makes its potential role in the
regulation of the immune response plausible. The presence of
neamine did not affect angiogenin production in the MLRs.
Besides the known effect of angiogenin in rRNA
transcription, global protein synthesis and cell proliferation
(1,2), the angiogenin nuclear translocator
inhibitor neamine did not affect cell proliferation in MLRs.
Although suppression of cell proliferation by neamine is typical in
various cancer cell lines tested (1,2,16–18),
this effect is not universal. For instance, neamine inhibits
nuclear translocation of angiogenin in both HSC-2 and SAS cells,
but proliferation is only suppressed in HSC-2 cells lines (17). This has been attributed to the
different expression levels of angiogenin in different cell lines
(17). In addition, previous studies
have shown that the tiRNAs produced by cytoplasmic angiogenin
inhibit global protein synthesis by ~20% (4,5). Such a
modest decrease in global protein synthesis may not be adequate to
suppress cell proliferation in neamine-treated MLRs.
The results of the current study indicate that
angiogenin protects MLR-derived CD4+ T-cells from
apoptosis, since neamine, its nuclear translocation inhibitor,
increased the protein levels of CC3, the terminal caspase that all
apoptotic pathways converge with (20). Preferential translation of
IRES-containing mRNAs is unlikely to be the reason, since Bcl-2
expression remained unaffected by neamine treatment of the
MLR-derived CD4+ T-cells. Bcl-2 is an anti-apoptotic
factor for which translation is mediated via IRES during cell
stress (21). Interestingly, a
previous study identified that angiogenin-induced tiRNAs did not
affect Bcl-2 expression in cells under osmotic stress (9). In that study, tiRNAs protected cells
from apoptosis by binding to the cytochrome c released from
mitochondria, preventing its binding to apoptotic protease
activating factor 1 and apoptosome formation (9), a concept not evaluated in the present
study.
In MLR-derived CD4+ T-cells, neamine
significantly increased the expression of Bax. This suggests
increased apoptosis, since the ratio of pro-apoptotic Bax to
anti-apoptotic Bcl-2 governs mitochondrial membrane permeability
and the release of cytochrome c (20). It is likely that inhibition of
angiogenin nuclear translocation by neamine prevents the direct
interaction of angiogenin with tumor suppressor p53. This
interaction is known to take place in the nucleus and inhibits
phosphorylation of p53, increasing p53 binding with mouse double
minute 2 homolog and p53 degradation (8). In the absence of nuclear angiogenin,
such as when neamine is used, p53 accumulates and increases the
expression of its pro-apoptotic transcriptional targets, including
Bax, leading to apoptosis (8). A
recent study found that in T-cells isolated from MLRs conducted
under the same conditions as the present study, p53 and
phosphorylated p53 were robustly expressed (22).
In conclusion, the results of the present study show
that angiogenin is upregulated during the alloreactive adaptive
immune response. Angiogenin was determined not to effect the T-cell
expansion phase, but did inhibit the T-cell contraction phase
through reducing CD4+ T-cell apoptosis.
References
|
1
|
Tsuji T, Sun Y, Kishimoto K, Olson KA, Liu
S, Hirukawa S and Hu GF: Angiogenin is translocated to the nucleus
of HeLa cells and is involved in ribosomal RNA transcription and
cell proliferation. Cancer Res. 65:1352–1360. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hirukawa S, Olson KA, Tsuji T and Hu GF:
Neamine inhibits xenografic human tumor growth and angiogenesis in
athymic mice. Clin Cancer Res. 11:8745–8752. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fu H, Feng J, Liu Q, Sun F, Tie Y, Zhu J,
Xing R, Sun Z and Zheng X: Stress induces tRNA cleavage by
angiogenin in mammalian cells. FEBS Lett. 583:437–442. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yamasaki S, Ivanov P, Hu GF and Anderson
P: Angiogenin cleaves tRNA and promotes stress-induced
translational repression. J Cell Biol. 185:35–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Emara MM, Ivanov P, Hickman T, Dawra N,
Tisdale S, Kedersha N, Hu GF and Anderson P: Angiogenin-induced
tRNA-derived stress-induced RNAs promote stress-induced stress
granule assembly. J Biol Chem. 285:10959–10968. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li S and Hu GF: Emerging role of
angiogenin in stress response and cell survival under adverse
conditions. J Cell Physiol. 227:2822–2826. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saikia M and Hatzoglou M: The Many Virtues
of tRNA-derived Stress-induced RNAs (tiRNAs): Discovering novel
mechanisms of stress response and effect on human health. J Biol
Chem. 290:29761–29768. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sadagopan S, Veettil MV, Chakraborty S,
Sharma-Walia N, Paudel N, Bottero V and Chandran B: Angiogenin
functionally interacts with p53 and regulates p53-mediated
apoptosis and cell survival. Oncogene. 31:4835–4847. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Saikia M, Jobava R, Parisien M, Putnam A,
Krokowski D, Gao XH, Guan BJ, Yuan Y, Jankowsky E, Feng Z, et al:
Angiogenin-cleaved tRNA halves interact with cytochrome c,
protecting cells from apoptosis during osmotic stress. Mol Cell
Biol. 34:2450–2463. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rybak SM, Fett JW, Yao QZ and Vallee BL:
Angiogenin mRNA in human tumor and normal cells. Biochem Biophys
Res Commun. 146:1240–1248. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Olson KA, Verselis SJ and Fett JW:
Angiogenin is regulated in vivo as an acute phase protein. Biochem
Biophys Res Commun. 242:480–483. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eleftheriadis T, Antoniadi G, Liakopoulos
V, Pissas G, Stefanidis I and Galaktidou G: Plasma angiogenin and
vascular endothelial growth factor a among hemodialysis patients.
Iran J Kidney Dis. 6:209–215. 2012.PubMed/NCBI
|
|
13
|
Lioté F, Champy R, Moenner M,
Boval-Boizard B and Badet J: Elevated angiogenin levels in synovial
fluid from patients with inflammatory arthritis and secretion of
angiogenin by cultured synovial fibroblasts. Clin Exp Immunol.
132:163–168. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brenner D, Krammer PH and Arnold R:
Concepts of activated T cell death. Crit Rev Oncol Hematol.
66:52–64. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sato T, Deiwick A, Raddatz G, Koyama K and
Schlitt HJ: Interactions of allogeneic human mononuclear cells in
the two-way mixed leucocyte culture (MLC): Influence of cell
numbers, subpopulations and cyclosporin. Clin Exp Immunol.
115:301–308. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ibaragi S, Yoshioka N, Li S, Hu MG,
Hirukawa S, Sadow PM and Hu GF: Neamine inhibits prostate cancer
growth by suppressing angiogenin-mediated rRNA transcription. Clin
Cancer Res. 15:1981–1988. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kishimoto K, Yoshida S, Ibaragi S,
Yoshioka N, Hu GF and Sasaki A: Neamine inhibits oral cancer
progression by suppressing angiogenin-mediated angiogenesis and
cancer cell proliferation. Anticancer Res. 34:2113–2121.
2014.PubMed/NCBI
|
|
18
|
Liu YP, Hu GF and Wu YX: Neamine is
preferential as an anti-prostate cancer reagent by inhibiting cell
proliferation and angiogenesis, with lower toxicity than
cis-platinum. Oncol Lett. 10:137–142. 2015.PubMed/NCBI
|
|
19
|
Wu D, Yu W, Kishikawa H, Folkerth RD,
Iafrate AJ, Shen Y, Xin W, Sims K and Hu GF: Angiogenin
loss-of-function mutations in amyotrophic lateral sclerosis. Ann
Neurol. 62:609–617. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fadeel B and Orrenius S: Apoptosis: A
basic biological phenomenon with wide-ranging implications in human
disease. J Intern Med. 258:479–517. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sherrill KW, Byrd MP, Van Eden ME and
Lloyd RE: BCL-2 translation is mediated via internal ribosome entry
during cell stress. J Biol Chem. 279:29066–29074. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eleftheriadis T, Pissas G, Antoniadi G,
Spanoulis A, Liakopoulos V and Stefanidis I: Indoleamine
2,3-dioxygenase increases p53 levels in alloreactive human T cells,
and both indoleamine 2,3-dioxygenase and p53 suppress glucose
uptake, glycolysis and proliferation. Int Immunol. 26:673–684.
2014. View Article : Google Scholar : PubMed/NCBI
|