Introduction
Sepsis in defined as a systemic inflammatory
response to infection (1). The
importance of immune function disorder in sepsis and septic shock
has previously been demonstrated (2), and previous studies have demonstrated
that T cells are pivotal in the pathogenesis of septic
complications via suppression of the adaptive immune response
(2–4). The role of T cells in the immune
response of sepsis is well documented (3,4), and it
has previously been demonstrated that septic patients exhibit
markedly decreased numbers of total T cells and CD4+ T
lymphocytes in their peripheral blood (5). Furthermore, previous studies have
demonstrated that the persistent lymphopenia correlates with early
and late mortality in sepsis or patients with septic shock
(6–8). It has also been demonstrated that the
early apoptosis of circulating lymphocytes in septic shock is
associated with poor outcomes (7).
Therefore, lymphopenia may serve as a biomarker for sepsis-induced
immunosuppression and the prevention of lymphopenia may be a
potential therapeutic strategy for the treatment of sepsis
(8).
Glucocorticoids have anti-inflammatory,
anti-allergic and anti-shock properties; therefore, they have been
widely used to treat inflammatory and autoimmune diseases,
including severe sepsis and septic shock (9). Previous studies have demonstrated that
glucocorticoids prevent the release of proinflammatory cytokines
via the regulation of lymphocyte function (9,10);
therefore, glucocorticoids may significantly improve lymphopenia in
septic shock. However, the duration of treatment could
differentially affect the patient response to treatment and the
mechanisms underlying the role of glucocorticoids on lymphopenia in
septic shock remain unclear.
Various studies have investigated the role of
glucocorticoids in micro (mi)RNA regulation and have demonstrated
that glucocorticoids may affect T-cell function or the
apoptosis/proliferation of lymphocytes via miRNA regulation
(11,12). As a class of small non-coding RNA
molecules, miRNA has been shown to have a critical role in cell
proliferation/differentiation, apoptosis, innate immune responses
and inflammation, including the regulation of lymphocyte function
and sepsis (13,14). Increasing evidence has indicated that
miRNA-155 is a critical regulator of inflammation and immune
function (15,16). Previous studies have demonstrated
that miRNA-155 has an important role in the regulation of various
inflammatory and immunological diseases, including Graves'
ophthalmopathy, allergic asthma, atherogenesis, multiple sclerosis
and organ transplantation (17–21).
Moreover, miRNA-155 has been associated with the regulation of
immunity and inflammation in infectious diseases, including sepsis
(21–23). This is supported by a previous study
which demonstrated that glucocorticoids are capable of regulating
miRNA-155 expression levels in the livers of septic mice (23). Although previous studies have
elucidated that miRNA-155 has a critical role in the proliferation
of T cells (24,25), whether glucocorticoids regulate
T-cell proliferation or function via miRNA-155 in patients with
septic shock remains unclear.
The present study focused on the role of
glucocorticoids in the regulation of miRNA-155 and T-cell
proliferation in patients with septic shock.
Materials and methods
Patients
Adult patients were recruited from the Intensive
Care Unit at the First Hospital of Jilin University (Jilin, China)
between October 2012 and May 2014. Ethical approval was obtained
from the Medical Ethics Committee of the First Hospital of Jilin
University and informed consent was obtained from either the
patients or the patients' families. Septic shock was diagnosed
based on criteria outlined in the International Guidelines for
Management of Severe Sepsis and Septic Shock: 2012 (26). The diagnosis of sepsis was made on
the basis of an identifiable or suspected infection site and
evidence of systemic inflammatory response syndrome manifested by
at least two of the following criteria: i) Body temperature, >38
or <36°C; ii) respiratory rate, >20 breaths/min; iii) heart
rate, >90 beats/min; iv) white blood cell count,
>12,000/mm3 or <4,000/mm3. Septic shock
was defined as sepsis-induced hypotension persisting despite
adequate fluid resuscitation. Hypotension was defined as systolic
blood pressure (SBP) <90 mmHg, mean arterial pressure <70
mmHg or a SBP reduction of >40 mmHg or standard deviation below
normal for the patient's age (26).
A total of 21 patients undergoing septic shock were
enrolled in the septic shock group, and 25 healthy volunteers were
included in the control group. Exclusion criteria included: i)
Patients without informed consent; ii) patients <18 years old;
iii) patients undergoing continuous renal replacement therapy prior
to sampling; iv) patients receiving immunosuppressive therapy; and
v) patients infected with viruses, including Mycobacterium
tuberculosis. Blood samples collected by venous puncture were
stored in BD Vacutainer tubes supplemented with lithium heparin (BD
Biosciences, New Jersey, NY, USA) prior to steroid therapy. Blood
samples from patients were collected within 24 h after the onset of
septic shock. Demographic characteristics were also collected, and
are shown in Table I.
 | Table I.Demographic characteristics of all
subjects. |
Table I.
Demographic characteristics of all
subjects.
| Characteristics | Control (n=25) | Septic shock
(n=21) | P-value |
|---|
| Age, years | 65.2±13.4 | 67.2±13.4 | >0.05 |
| Gender,
female/male | 16/9 | 13/8 | >0.05 |
| Leukocytes,
109/l | 5.8±1.4 | 14.6±5.5 | <0.001 |
| Lymphocytes,
109/l | 1.1±0.3 | 0.3±0.1 | <0.001 |
| Hs-CRP, mg/l | 2.3±0.7 | 12.6±3.1 | <0.001 |
| Infection site, n
(%) |
|
|
|
| Lung | – | 9 (42.9) |
|
|
Abdomen | – | 5 (23.8) |
|
|
Blood | – | 3 (14.3) |
|
| Burn | – | 2 (9.5) |
|
|
Others | – | 2 (9.5) |
|
Isolation of T cells
Total T cells were isolated from peripheral blood
samples by negative selection using a Rosette Sep kit (15022; Stem
Cell Technologies, Vancouver, CA, USA). T cells were purified using
the MACS Pan T Cell Isolation kit II (130-092-881; Miltenyi Biotec,
Auburn, CA, USA), according to the manufacturer's protocol.
CD4+ T cell purity was ≥96%, as determined by flow
cytometry (Attune NxT, Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Cells were cultured in RPMI-1640 medium (Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) and antibiotics (100
U/ml penicillin and 0.1 mg/ml streptomycin).
Transfection with anti-miRNA-155
oligonucleotides (ODNs)
Antisense ODNs against human miRNA-155 were used as
the anti-miRNA-155 inhibitor (HmiR-AN0221-SN-10), and negative
control (NC) ODNs (CmiR-AN0001-SN) were also used as for comparison
(both Genecopoeia Inc., Guangzhou, China). Cells at 40–60%
confluence were transfected. Individual ODNs were mixed with
Lipofectamine 2000 (Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol and were subsequently administered to
the cells. The efficiency of ODN transfection was assessed by
monitoring the uptake of siRNA labeled with 6-carboxyfluorescein.
Furthermore, the transfection efficiency for each siRNA was >90%
and no significant difference existed between the two types of
siRNA.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cells were seeded into 6-well plates at a
concentration of 5×104 cells/well. Following treatment,
total RNA was extracted from cells using TRIzol reagent (15596-018;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The RNA sample was treated with 10 U of DNase I in a
volume of 50 ml (04716728001; Roche Diagnostics, Basel,
Switzerland) at 37°C for 20 min. First-strand cDNA was synthesized
using a reverse transcription kit (4368814; Thermo Fisher
Scientific, Inc.). qPCR was performed in order to analyze miRNA-155
mRNA expression levels using a TaqMan Universal Master Mix II kit
(4440040; Thermo Fisher Scientific, Inc.). β-actin was used as the
reference gene. The abundance of miRNA-155 expression in each
sample was determined relative to the abundance of the β-actin
reference gene. The qPCR data were analyzed and expressed as the
relative miRNA levels of the cycle threshold value, which was
subsequently converted to the fold change with the
2−ΔΔCT method (27). The
primer sequences of miR-155 were as follows: Sense, 5′
TTAATGCTAATCGTGATAG,-3′ and antisense 5′-ACCTGAGAGTAGACCAGA-3′.
Immunofluorescence staining
Cells were seeded into 24-well plates at a
concentration of 1×104 cells/well. Following treatment,
cells were fixed with 95% alcohol for 5 min and permeabilized with
0.5% Triton X-100 for 5 min. Subsequently, cells were incubated
with 5% bovine serum albumin for 0.5 h to block non-specific
binding. Then cells were incubated with antibodies against
proliferating cell nuclear antigen (rabbit polyclonal antibody
PCNA; 1:100; sc-25280, Santa Cruz Biotechnology, Inc.) for 2 h at
room temperature. Following washing, cells were incubated with
fluorescein isothiocyanate-conjugated anti-mouse immunoglobulin G
secondary antibodies (sc-65561, Santa Cruz Biotechnology, Inc.,)
prepared in phosphate-buffered saline (PBS) for 1 h at room
temperature. Negative controls were conducted with PBS instead of
the primary antibodies. Sections were examined using a fluorescence
microscope (BX53, Olympus Corporation, Tokyo, Japan), and the
results were expressed as the percentage of positive cells.
Cell proliferation
In order to investigate the rate of proliferation, T
cells from patients and controls were stimulated with 10 µg/ml
phytohemagglutinin (PHA; Sigma-Aldrich, St. Louis, MO, USA), and in
a separate experiment, T cells from patients with septic shock were
stimulated with 10, 50 or 100 nM dexamethasone (DXM; D1756,
Sigma-Aldrich). In both experiments, the duration of stimulation
was 48 h. Cell proliferation was evaluated using cell count and
methyl thiazolyl tetrazolium (MTT) assays (Sigma-Aldrich).
For the cell count assay, cells were plated into
6-well plates at a concentration of 5×104 cells/well.
Following stimulation, cells were collected by trypsin (C0201,
Beyotime Institute of Biotechnology, Haimen, China) digestion and
total cell numbers were calculated using a hemocytometer (Countess
Automated Cell Counter, Thermo Fisher Scientific, Inc.) following
trypan blue (ST798, Beyotime Institute of Biotechnology)
exclusion.
For the MTT assay, cells were seeded into 96-well
plates at a concentration of 5×103 cells/well. Following
treatment, 20 µl MTT (ST316, Beyotime Institute of Biotechnology)
was added to the wells and the plates were incubated for 4 h. The
supernatant was subsequently removed and the plate was incubated
with 150 µl/well dimethyl sulfoxide (DMSO; Sigma-Aldrich) at room
temperature for 10 min on a swing bed. Cells were quantified by
spectrophotometry at 490 nm using an absorbance microplate reader
(ELx800; BioTek Instruments, Inc., Winooski, VT, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation
and were analyzed by t-test or one-way analysis of variance,
followed by q-test using SPSS software, version 12.0 (SPSS, Inc.,
Chicago, IL, USA). For all tests, P<0.05 was considered to
indicate a statistically significant difference.
Results
Demographic characteristics of the
study subjects
The demographic characteristics of subjects in the
two groups are shown in Table I. No
significant differences in gender or age were detected between the
two groups. The levels of white blood cells (leukocytes) and serum
high-sensitivity C-reactive protein were significantly higher in
the septic shock group, as compared with the control group; whereas
the lymphocyte count was significantly lower in the septic shock
group, as compared with the control group. In the present study,
the primary cause of septic shock was severe pneumonia (n=9). Other
causes included peritonitis (n=5), injury (n=2) and burn (n=2).
miRNA-155 levels and T cell
proliferation
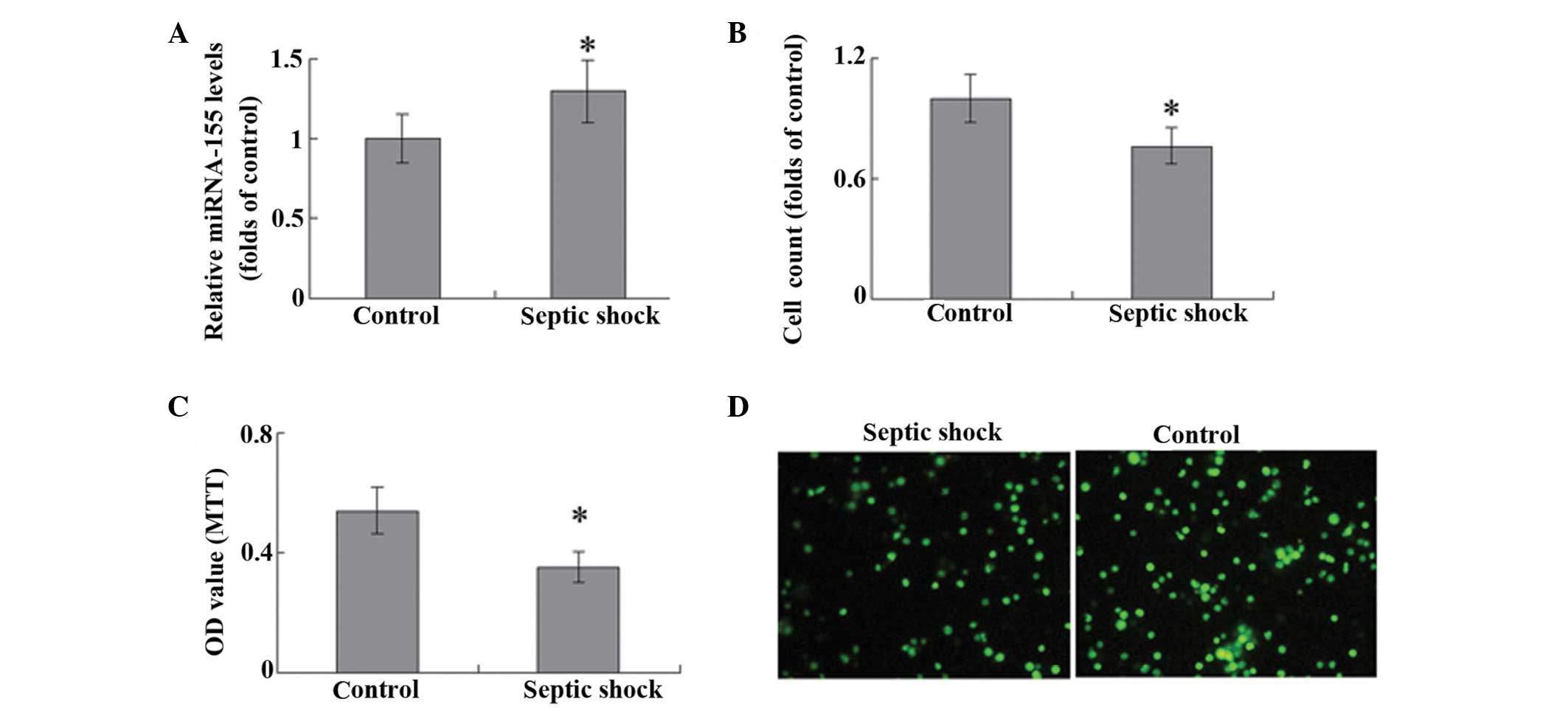
miRNA-155 levels in T cells were significantly
higher in the septic shock group, as compared with the control
group (P<0.05; Fig. 1A). A cells
counting assay was used to evaluate T-cell proliferation, and
stimulation with 10 mg/ml PHA was observed to promote the
proliferation of T lymphocytes in patients with septic shock and
the control group. Notably, the T cells harvested from the
peripheral blood of patients with septic shock exhibited
significantly weaker proliferative ability, as compared with the
controls (P<0.05; Fig. 1B). The
results of MTT assay and PCNA staining were consistent with those
demonstrated by the cells counts (Fig.
1C and D).
DXM regulates T-cell proliferation and
miRNA-155 expression
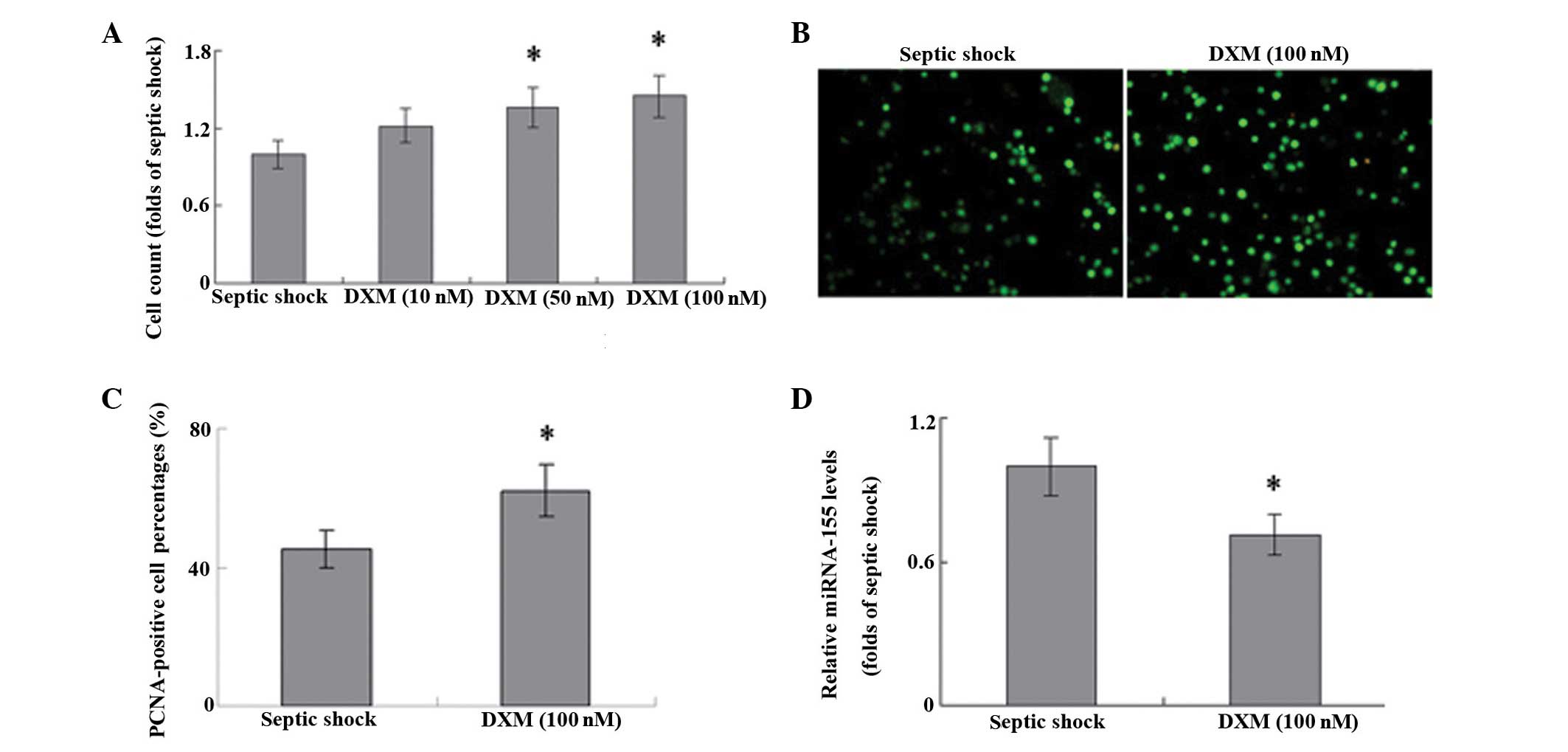
In T cells harvested from patients with septic
shock, treatment with DXM significantly increased the proliferation
of cells in a concentration-dependent manner, as compared with that
in the untreated controls (Fig. 2A;
P<0.05). The results of PCNA staining were consistent with these
results (Fig. 2B and C), which
suggests that DXM treatment may have a role in the proliferation of
T cells during septic shock. Furthermore, stimulation with DXM
markedly inhibited the expression levels of miRNA-155 in the T
cells of patients with septic shock (Fig. 2D).
miRNA-155 is associated with T-cell proliferation.
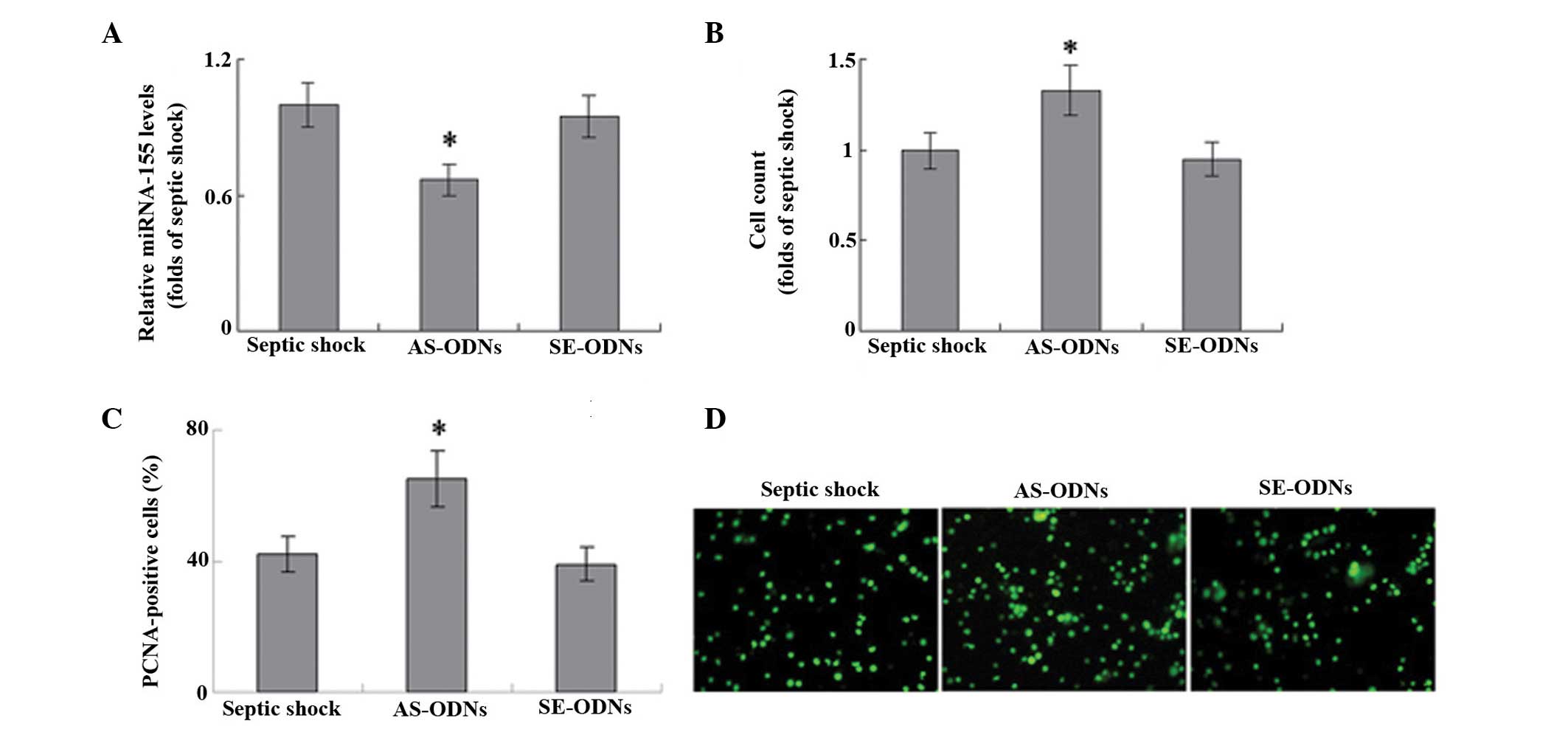
In order to explore the role of miRNA-155 in the proliferation of T
cells, miRNA-155 expression was knocked down using an ODN against
human miRNA-155. The ODN anti-miRNA-155 inhibitor successfully
significantly inhibited miRNA-155 mRNA expression levels in T cells
(Fig. 3A). In a cell count assay,
the anti-miRNA-155 inhibitor significantly increased the
proliferation of T cells induced by 10 mg/ml PHA, as compared with
that of untransfected cells (P<0.05; Fig. 3B). Furthermore, transfection with
this anti-miRNA-155 ODN resulted in a significant increase in the
number of PCNA-positive cells in the T lymphocytes of patients with
septic shock, as compared with the untransfected control (Fig. 3C; P<0.05). These results suggest
that miRNA-155 may be involved in the proliferation of T cells
during septic shock.
Discussion
The results of the present study demonstrate that
the miRNA-155 expression levels were increased in the T cells of
patients with septic shock. Furthermore, DXM administration
successfully inhibited miRNA-155 expression and promoted T cell
proliferation, and a small-interfering RNA against miRNA-155 was
demonstrated to significantly increase the proliferation of T cells
during septic shock. These results suggest that miRNA-155 may be
associated with the glucocorticoid-induced proliferation of T cells
observed in patients with septic shock.
Lymphocytes have a critical role in the immune
response of sepsis (3,4) and it has previously been demonstrated
that patients with septic shock exhibit significantly decreased
levels of total and CD4+ T lymphocytes in peripheral
blood (5). Previous studies have
demonstrated that apoptosis of T cells occurs during septic shock;
however few have focused on the proliferative ability of T cells in
patients with septic shock (5–7). The
present study demonstrated that T cells from patients with septic
shock exhibited weak proliferative ability under the stimulation of
PHA. This suggests that PHA also participates in the persistent
lymphopenia in septic shock; however, the importance and mechanism
of the balance of proliferation/apoptosis of lymphocytes in
patients with sepsis requires further clarification.
miRNAs have been demonstrated to have critical roles
in cell proliferation, apoptosis, inflammation and the regulation
of lymphocyte function (13–15). Previous studies have outlined the
critical role of miRNA-155 in inflammatory and immune diseases
(17–21), and the present study demonstrated the
important role of miRNA-155 in septic shock. The results of the
present study are consistent with previous studies in
lipopolysaccharide (LPS)-induced septic mice. In a mice model of
sepsis, Wang et al (23)
demonstrated that LPS significantly increased miRNA-155 expression
levels in liver tissues, in addition to several inflammatory
factors. Another study demonstrated that LPS was capable of
inducing miRNA-155 expression in the spleens of mice (21). These reports suggest that miRNA-155
has a critical role in sepsis and septic shock. Certain pathways,
including glycogen synthase kinase-3β and arginase-2, have been
found to be associated with the miRNA-155-mediated regulation of
T-cell proliferation in cardiac allograft rejection in a murine
transplantation model (14–25). Although other validated target genes
of miRNA-155 have also been established (28,29),
their role in the proliferation of T lymphocytes during septic
shock is yet to be elucidated.
Previous studies have demonstrated the critical role
of glucocorticoid therapy in the treatment of septic shock; in
particular, glucocorticoids may prevent the release of
proinflammatory cytokines via the regulation of lymphocyte function
(9,10). The results of the present study
suggest that glucocorticoids may promote the proliferation of T
lymphocytes in patients with septic shock. It is well documented
that glucocorticoids are capable of affecting T-cell function and
lymphocyte apoptosis/proliferation via miRNAs, such as miR-98 and
miR-17 (11,12). The present study indicated that
miRNA-155 may be a target for glucocorticoids in T cells. This
result is consistent with a previous study investigating a mouse
model of sepsis, which demonstrated that treatment with DXM
inhibited the expression of miRNA-155 to below baseline levels
(23). These results indicate that
miRNAs may be an important therapeutic target of glucocorticoids in
the regulation of inflammatory and immunological diseases. Notably,
it has been suggested that glucocorticoids may bind directly to the
B-cell integration cluster gene to repress miRNA-155 expression;
however, this mechanism requires further investigation in patients
with septic shock (30).
In conclusion, the present study demonstrates the
key role of miRNA-155 in the proliferation of T cells in patients
with septic shock. Therefore, although the precise molecular
mechanism remains unclear, the regulation of miRNA-155 via
glucocorticoids may be a novel therapeutic mechanism for the
regulation of inflammation and immune response in patients with
septic shock.
References
|
1
|
Annane D, Bellissant E and Cavaillon JM:
Septic shock. Lancet. 365:63–78. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chung CS, Watkins L, Funches A,
Lomas-Neira J, Cioffi WG and Ayala A: Deficiency of gammadelta T
lymphocytes contributes to mortality and immunosuppression in
sepsis. Am J Physiol Regul Integr Comp Physiol. 291:R1338–R1343.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wisnoski N, Chung CS, Chen Y, Huang X and
Ayala A: The contribution of CD4+ CD25+
T-regulatory-cells to immune suppression in sepsis. Shock.
27:251–257. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang LN, Yao YM and Sheng ZY: The role of
regulatory T cells in the pathogenesis of sepsis and its clinical
implication. J Interferon Cytokine Res. 32:341–349. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Inoue S, Suzuki-Utsunomiya K, Okada Y,
Taira T, Iida Y, Miura N, Tsuji T, Yamagiwa T, Morita S, Chiba T,
et al: Reduction of immunocompetent T cells followed by prolonged
lymphopenia in severe sepsis in the elderly. Crit Care Med.
41:810–819. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Drewry AM, Samra N, Skrupky LP, Fuller BM,
Compton SM and Hotchkiss RS: Persistent lymphopenia after diagnosis
of sepsis predicts mortality. Shock. 42:383–391. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Le Tulzo Y, Pangault C, Gacouin A,
Guilloux V, Tribut O, Amiot L, Tattevin P, Thomas R, Fauchet R and
Drénou B: Early circulating lymphocytes apoptosis in septic shock
is associated with poor outcome. Shock. 18:487–494. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Turrel F, Guignant C, Venet F, Lepape A
and Monneret G: Innovative therapeutic strategies for restoring
lymphocyte functions in septic patients. Inflamm Allergy Drug
Targets. 7:181–186. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Annane D, Bellissant E, Bollaert PE,
Briegel J, Confalonieri M, De Gaudio R, Keh D, Kupfer Y, Oppert M
and Meduri GU: Corticosteroids in the treatment of severe sepsis
and septic shock in adults: A systematic review. JAMA.
301:2362–2375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Keh D, Boehnke T, Weber-Cartens S, Schulz
C, Ahlers O, Bercker S, Volk HD, Doecke WD, Falke KJ and Gerlach H:
Immunologic and hemodynamic effects of ‘low-dose’ hydrocortisone in
septic shock: A double-blind, randomized, placebo-controlled,
crossover study. Am J Respir Crit Care Med. 167:512–520. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Davis TE, Kis-Toth K, Szanto A and Tsokos
GC: Glucocorticoids suppress T cell function by up-regulating
microRNA-98. Arthritis Rheum. 65:1882–1890. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Smith LK, Shah RR and Cidlowski JA:
Glucocorticoids modulate microRNA expression and processing during
lymphocyte apoptosis. J Biol Chem. 285:36698–36708. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rodriguez A, Vigorito E, Clare S, Warren
MV, Couttet P, Soond DR, van Dongen S, Grocock RJ, Das PP, Miska
EA, et al: Requirement of bic/microRNA-155 for normal immune
function. Science. 316:608–611. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang HJ, Zhang PJ, Chen WJ, Jie D, Dan F,
Jia YH and Xie LX: Characterization and identification of novel
serum microRNAs in sepsis patients with different outcomes. Shock.
39:480–487. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yao R, Ma YL, Liang W, Li HH, Ma ZJ, Yu X
and Liao YH: MicroRNA-155 modulates Treg and Th17 cells
differentiation and Th17 cell function by targeting SOCS1. PLoS
One. 7:e460822012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu R, Huffaker TB, Kagele DA, Runtsch MC,
Bake E, Chaudhuri AA, Round JL and O'Connell RM: MicroRNA-155
confers encephalogenic potential to Th17 cells by promoting
effectorgene expression. J Immunol. 190:5972–5980. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li K, Du Y, Jiang BL and He JF: Increased
microRNA-155 and decreased microRNA-146a may promote ocular
inflammation and proliferation in Graves' ophthalmopathy. Med Sci
Monit. 20:639–643. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Malmhäll C, Alawieh S, Lu Y, Sjöstrand M,
Bossios A, Eldh M and Rådinger M: MicroRNA-155 is essential for
T(H)2-mediated allergen-induced eosinophilic inflammation in the
lung. J Allergy Clin Immunol. 133:1429–1438. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li J and Gong J, Li P, Li M, Liu Y, Liang
S and Gong J: Knockdown of microRNA-155 in kupffer cells results in
immunosuppressive effects and prolongs survival of mouse liver
allografts. Transplantation. 97:626–635. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang J, Cheng Y, Cui W, Li M, Li B and
Guo L: MicroRNA-155 modulates Th1 and Th17 cell differentiation and
is associated with multiple sclerosis and experimental autoimmune
encephalomyelitis. J Neuroimmunol. 266:56–63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tili E, Michaille JJ, Cimino A, Costinean
S, Dumitru CD, Adair B, Fabbri M, Alder H, Liu CG, Calin GA and
Croce CM: Modulation of miR-155 and miR-125b levels following
lipopolysaccharide/TNF-alpha stimulation and their possible roles
in regulating the response to endotoxin shock. J Immunol.
179:5082–5089. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Piccinini AM and Midwood KS: Endogenous
control of immunity against infection: Tenascin-C regulates
TLR4-mediated inflammation via microRNA-155. Cell Rep. 2:914–926.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang ZH, Liang YB, Tang H, Chen ZB, Li ZY,
Hu XC and Ma ZF: Dexamethasone down-regulates the expression of
microRNA-155 in the livers of septic mice. PLoS One. 8:e805472013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dunand-Sauthier I, Irla M, Carnesecchi S,
Seguín-Estévez Q, Vejnar CE, Zdobnov EM, Santiago-Raber ML and
Reith W: Repression of arginase-2 expression in dendritic cells by
microRNA-155 is critical for promoting T cell proliferation. J
Immunol. 193:1690–1700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Feng Z, Xia Y, Zhang M and Zheng J:
MicroRNA-155 regulates T cell proliferation through targeting GSK3β
in cardiac allograft rejection in a murine transplantation model.
Cell Immunol. 281:141–149. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dellinger RP, Levy MM, Rhodes A, Annane D,
Gerlach H, Opal SM, Sevransky JE, Sprung CL, Douglas IS, Jaeschke
R, et al: Surviving Sepsis Campaign Guidelines Committee including
the Pediatric Subgroup. Surviving sepsis campaign: International
guidelines for management of severe sepsis and septic shock. Crit
Care Med. 41:580–637. 2012. View Article : Google Scholar
|
|
27
|
Liu TE, Wang S, Zhang L, Guo L, Yu Z, Chen
C and Zheng J: Growth hormone treatment of premature ovarian
failure in a mouse model via stimulation of the Notch-1 signaling
pathway. Exp Ther Med. 12:215–221. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ceppi M, Pereira PM, Dunand-Sauthier I,
Barras E, Reith W, Santos MA and Pierre P: MicroRNA-155 modulates
the interleukin-1 signaling pathway in activated human
monocyte-derived dendritic cells. Proc Natl Acad Sci USA.
106:2735–2740. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu F, Weidmer A, Liu CG, Volinia S, Croce
CM and Lieberman PM: Epstein-Barr virus-induced miR-155 attenuates
NF-kappaB signaling and stabilizes latent virus persistence. J
Virol. 82:10436–10443. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yin Q, Wang X, McBride J, Fewell C and
Flemington E: B-cell receptor activation induces BIC/miR-155
expression through a conserved AP-1 element. J Biol Chem.
283:2654–2662. 2008. View Article : Google Scholar : PubMed/NCBI
|