Introduction
Peri-implantitis is a destructive inflammatory
disease that affects the tissues surrounding dental implants
(1,2). It has been demonstrated that
peri-implantitis is a crucial element in implant failure. In total,
~30% of patients that receive dental implants develop
peri-implantitis (3). However, there
are currently no effective therapeutic strategies against
peri-implantitis. Moreover, the use of dental implants is
constantly increasing, therefore an effective therapy to treat
peri-implantitis is required. Thus, further investigations into the
molecular pathophysiology of peri-implantitis are necessary in
order to provide novel options for effective treatment (1).
Recently, a number of studies have investigated the
pathological mechanisms underlying peri-implantitis progression.
Becker et al (4) indicated
that serglycin (SRGN) expression was significantly
upregulated in peri-implantitis when compared with healthy
individuals. It has been suggested that this gene may inhibit bone
mineralization in vitro (5).
Another study indicated that concentrations of the nuclear
factor-κB (NF-B), soluble RANK ligand (sRANKL), osteoprotegerin
(OPG) and sclerostin are significantly increased in patients with
peri-implantitis (6). A number of
typical bone matrix molecules, including collagen, type IX, α 1
(COL9A1), bone gamma-carboxyglutamate (Gla) protein (BGLAP) and
secreted phosphoprotein 1 (SPP1) are decreased in the
peri-implantitis tissues (1).
Furthermore, it has been identified that fibroblasts are involved
in the pathogenesis of peri-implantitis (7). The regulation of inflammatory mediators
and matrix metalloproteinases (MMPs) in peri-implantitis
fibroblasts function in the pathogenesis of the disease (8), and levels of the anti-inflammatory
cytokine interleukin (IL)-10 are decreased in peri-implantitis
(9). Furthermore, peroxisome
proliferator-activated receptor γ (PPARγ) that can inhibit
inflammation and promote osteoblast function is downregulated in
the peri-implantitis tissues (10).
However, other mechanisms associated with peri-implantitis have not
been identified. Therefore, further research should focus on
elucidating other potential mechanisms and investigate target genes
for the treatment of peri-implantitis.
The microarray data of GSE57631 was used to confirm
the similarities and differences of inflamed peri-implantitis
tissues vs. normal peri-implantitis tissues at the mRNA level
(1). In contrast to results from a
previous study (1), the array data
of GSE57631 was downloaded and the differentially expressed genes
(DEGs) associated with peri-implantitis were analyzed using a
biological informatics approach. In addition, functional enrichment
analyses were performed for DEGs. In addition, a protein-protein
interaction (PPI) network was established and four significant
modules were analyzed. The present study aimed to identify the key
genes and pathways of peri-implantitis, and identify possible
significant mechanisms associated with it.
Materials and methods
Affymetrix microarray data
The array data of GSE57631 was downloaded from the
Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) database, which was
deposited by Schminke et al (1). Six samples of peri-implantitis tissues
and two samples of normal tissues were included in the present
study. The raw data and annotation files were downloaded for
subsequent analysis, based on the GPL15034 platform (Affymetrix
Human Gene 1.0 ST Array [HuGene10stv1_Hs_ENTREZ, Brainarray v14];
Affymetrix, Inc., Santa Clara, CA, USA).
Data preprocessing
The raw expression data was preprocessed using the
robust multiarray average (11)
algorithm by applying an oligo (12)
in the R statistical software program in Bioconductor (http://www.bioconductor.org/). Background correction,
normalization and a calculating expression were included in the
process of preprocessing. A total of 18,977 gene expression values
were obtained.
DEG analysis
The limma package (13) in Bioconductor was used to analyze
DEGs in peri-implantitis samples compared with controls. In the
process of the analysis, the P-values of DEGs were calculated using
a t-test in the limma package. |log2FC|≥1 and P<0.05 were used
as a cut-off criteria.
Gene Ontology (GO) and pathway
enrichment analyses
GO is a tool that is used for gene annotation by
collecting defined, structured, controlled vocabulary, which
includes three main categories: Molecular function (MF), biological
process (BP) and cellular component (CC) (14). Kyoto Encyclopedia of Genes and
Genomes (KEGG) is a database used for associating related gene sets
with their pathways (15). Moreover,
DAVID, an integrated data-mining environment, is used to analyze
gene lists (16).
GO annotation and KEGG pathway enrichment analyses
were conducted for upregulated and downregulated genes by DAVID.
Moreover, EASE ≤0.05 and gene counts ≥ were set as the threshold
value.
PPI network analysis
The Search Tool for the Retrieval of Interacting
Genes (STRING) (17) database
provides information regarding the predicted and experimental
interactions of proteins. The prediction method of this database
came from neighborhood, gene fusion, co-occurrence, co-expression
experiments, databases and text mining. Moreover, the interactions
of protein pairs in the database are presented with a combined
score. In the present study, DEGs were mapped into PPIs and a
combined score of >0.9 was used as the cut-off value. In
addition, PPI networks were constructed using Cytoscape software
version 3.2.1 (18).
Topological properties of the PPI network, including
degree (19), subgraph (20) and betweenness (21) centralities were determined using the
R software package igraph (22), in
order to analyze key genes in the network. A degree was used for
describing the importance of protein nodes in the network. Subgraph
centrality based on combining network topology and information of
protein complexes was used to measure the importance of nodes in
the network. The higher the degree and subgraph values were, the
more important the nodes were in the network. Moreover, the
betweenness centrality is an index describing the global
topological properties of the network and could be used to describe
how the nodes affect the connectivity between two nodes.
Betweenness was defined as the ratio of the number of every path
passed per node and the number of the shortest paths. The higher
the betweenness values are, the greater the impact of the node in
the network is. In addition, R software package igraph version
1.0.1 (22) was used for these three
methods.
Module analysis
Network module was one of the characteristics of the
protein network and may contain specific biological significance.
The Cytoscape software package Molecular Complex Detection (MCODE)
(23) was used to analyze the most
notable clustering module. Next, the KEGG pathway enriched by DEGs
in different modules was analyzed using DAVID online tool. EASE
≤0.05 and count ≥2 were set as the cutoff values.
Results
Data processing and DEG analysis
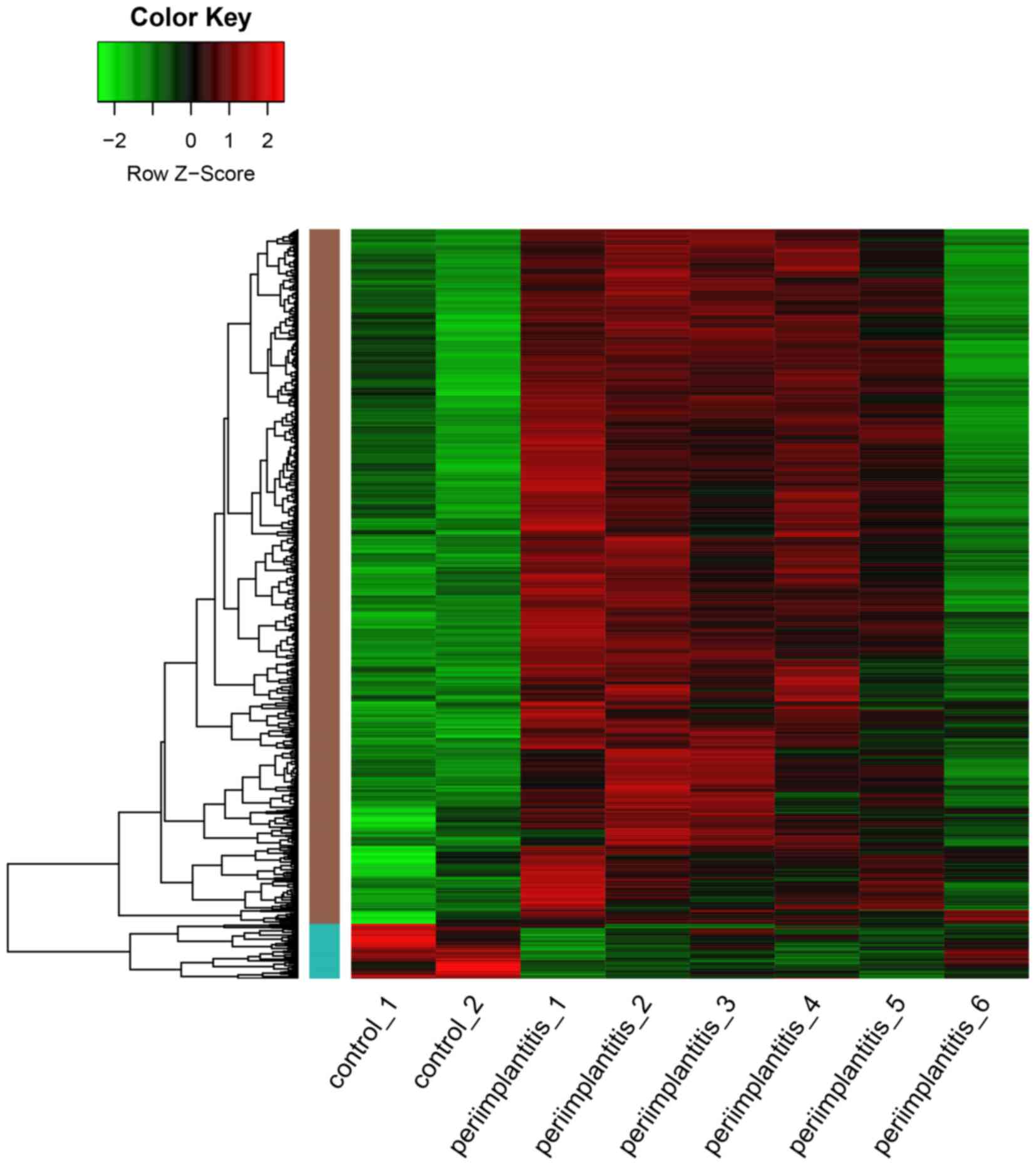
As shown in Fig. 1, a
total of 819 DEGs including 759 upregulated and 60 downregulated
genes, were identified in the peri-implantitis samples compared
with the control ones. As a result, the number of upregulated genes
was found to be significantly higher than that of the downregulated
ones.
GO and pathway enrichment
analyses
GO and KEGG pathway analyses were performed for
upregulated and downregulated DEGs, respectively. The GO terms of
upregulated DEGs were mainly associated with the proteasomal
protein catabolic process, endoplasmic reticulum, melanosome and
structural constituent of the cytoskeleton (Table IA). The downregulated DEGs were
mainly enriched in the epidermis and ectoderm development, and the
extracellular region part and cadmium ion binding (Table IB).
 | Table I.GO for differentially expressed
genes. |
Table I.
GO for differentially expressed
genes.
| A, Upregulated |
|---|
|
|---|
| Terms | Description | Counts | P-value |
|---|
| GO-BP |
|
|
|
| GO:
0010498 | Proteasomal protein
catabolic process | 24 |
5.32×10−11 |
| GO:
0043161 | Proteasomal
ubiquitin-dependent protein catabolic process | 24 |
5.32×10−11 |
| GO:
0032269 | Negative regulation
of cellular protein metabolic process | 30 |
9.32×10−10 |
| GO-CC |
|
|
|
| GO:
0005783 | Endoplasmic
reticulum | 115 |
1.37×10−21 |
| GO:
0042470 | Melanosome | 30 |
1.77×10−17 |
| GO:
0048770 | Pigment
granule | 30 |
1.77×10−17 |
| GO-MF |
|
|
|
| GO:
0005200 | Structural
constituent of cytoskeleton | 15 |
2.22×10−6 |
| GO:
0070003 | Threonine-type
peptidase activity | 8 |
1.13×10−5 |
|
| B,
Downregulated |
|
| Terms | Description | Counts | P-value |
|
| GO-BP |
|
|
|
|
GO:0008544 | Epidermis
development | 20 |
2.57×10−10 |
|
GO:0007398 | Ectoderm
development | 20 |
9.84×10−10 |
|
GO:0006954 | Inflammatory
response | 19 |
9.40×10−6 |
| GO-CC |
|
|
|
|
GO:0044421 | Extracellular
region part | 35 |
1.05×10−5 |
|
GO:0005576 | Extracellular
region | 54 |
1.33×10−4 |
|
GO:0005615 | Extracellular
space | 25 |
2.79×10−4 |
| GO-MF |
|
|
|
|
GO:0046870 | Cadmium ion
binding | 4 |
5.27×10−4 |
|
GO:0005507 | Copper ion
binding | 6 |
6.22×10−3 |
Two pathways that were significantly enriched by
upregulated DEGs were Alzheimer's disease and the proteasome
(Table II). Moreover, an important
gene HSP90AA1 was significantly enriched in the antigen
processing and presentation pathway and the nucleotide-binding
oligomerization domain (NOD)-like receptor signaling pathway, and
NFKB1 was enriched in NOD-like receptor signaling pathway in
KEGG pathway enrichment analysis for upregulated genes. However,
downregulated DEGs did not significantly enriched any pathways.
| Table II.KEGG pathway enrichment analysis for
upregulated differentially expressed genes. |
Table II.
KEGG pathway enrichment analysis for
upregulated differentially expressed genes.
| Term | Description | Counts | P-value |
|---|
| hsa05010 | Alzheimer's
disease | 25 |
1.69×10−5 |
| hsa03050 | Proteasome | 12 |
5.36×10−5 |
| hsa00190 | Oxidative
phosphorylation | 18 |
1.19×10−3 |
| hsa05012 | Parkinson's
disease | 17 |
2.65×10−3 |
| hsa05016 | Huntington's
disease | 21 |
3.41×10−3 |
| hsa00510 | N-Glycan
biosynthesis | 9 |
4.31×10−3 |
| hsa04142 | Lysosome | 15 |
7.11×10−3 |
| hsa00480 | Glutathione
metabolism | 9 |
7.27×10−3 |
| hsa04612 | Antigen processing
and presentation | 12 |
7.74×10−3 |
| hsa04621 | NOD-like receptor
signaling pathway | 10 |
8.63×10−3 |
| hsa00970 | Aminoacyl-tRNA
biosynthesis | 7 |
2.89×10−2 |
| hsa00020 | Citrate cycle (TCA
cycle) | 6 |
3.07×10−2 |
| hsa01040 | Biosynthesis of
unsaturated fatty acids | 5 |
3.51×10−2 |
| hsa05130 | Pathogenic
Escherichia coli infection | 8 |
4.45×10−2 |
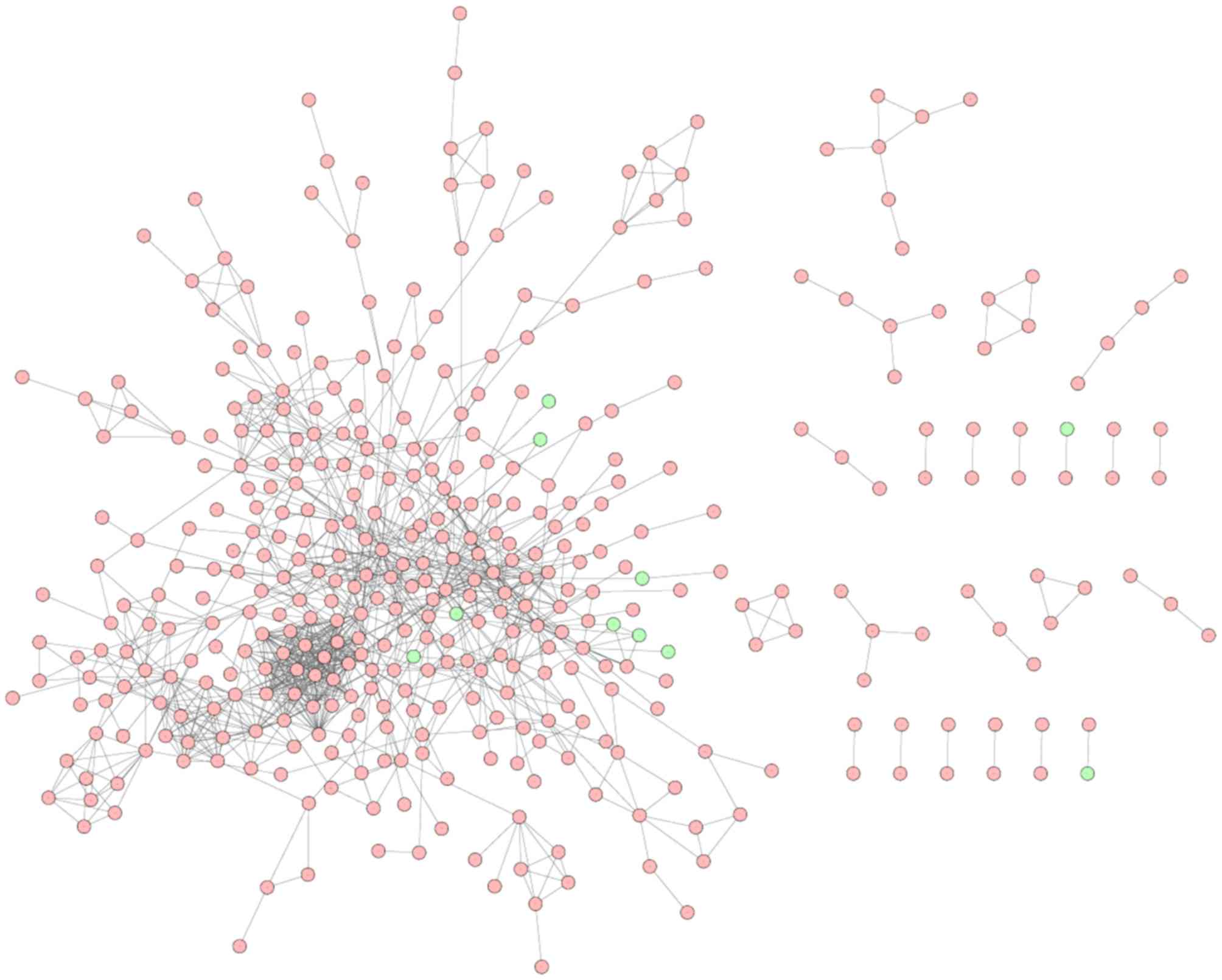
PPI network analysis
Based on the STRING database, a total of 413 nodes
and 1,114 protein pairs were obtained with a combined score >0.9
(Fig. 2). As shown in Fig. 2, the majority of the nodes in the
network were upregulated DEGs in peri-implantitis samples. The top
20 most up- and dowregulated nodes are presented in Table III. A number of protein enzyme
families, such as the proteasome subunit, α type 1 (PSMA1),
proteasome subunit, beta type 1 (PSMB1) and proteasome subunit, α
type 4 (PSMA4) were hub nodes based on the subgraph and degree
centralities. Heat shock protein HSP90AA1 (90 kDa α, member
1), Ras-related C3 botulinum toxin substrate 1, NFKB1, Jun
proto-oncogene were hub nodes with higher betweenness values.
| Table III.Nodes with higher values in subgraph
centrality, betweenness centrality and degree centrality. |
Table III.
Nodes with higher values in subgraph
centrality, betweenness centrality and degree centrality.
| Nodes | Subgragh | Nodes | Betweenness | Nodes | Degree |
|---|
| PSMA1 |
2.35×107 | HSP90AA1 |
2.78×104 | HSP90AA1 | 34 |
| PSMB1 |
2.32×107 | RAC1 |
1.77×104 | JUN | 27 |
| PSMA4 |
2.31×107 | NFKB1 |
1.38×104 | RAC1 | 27 |
| PSMB4 |
2.30×107 | JUN |
1.21×104 | PSMA1 | 25 |
| PSMA3 |
2.30×107 | HIF1A |
1.07×104 | PSMA4 | 25 |
| PSMB3 |
2.30×107 | CDH1 |
9.21×103 | PSMB1 | 25 |
| PSMB6 |
2.30×107 | SOS1 |
8.93×103 | PSMC2 | 25 |
| PSMC2 |
2.16×107 | CANX |
8.55×103 | PSMB4 | 24 |
| PSMD8 |
2.12×107 | HDAC1 |
8.35×103 | PSMA3 | 24 |
| PSMD5 |
2.08×107 | ATP5B |
8.24×103 | PSMB3 | 24 |
| PSME4 |
2.08×107 | GTF2B |
8.17×103 | PSMB6 | 24 |
| PSMD10 |
2.04×107 | HSPA5 |
7.84×103 | PSMD10 | 24 |
| PSME3 |
1.96×107 | STAT3 |
7.46×103 | PSMD8 | 23 |
| UBE2D1 |
1.54×107 | VCP |
7.32×103 | SEC61A1 | 23 |
| CDC16 |
1.36×107 | RPS5 |
7.14×103 | RPN2 | 23 |
| ANAPC5 |
1.36×107 | NME1 |
6.31×103 | NFKB1 | 23 |
| BUB3 |
1.35×107 | POLR2B |
5.90×103 | PSMD5 | 22 |
| SEC61A1 |
1.23×107 | P4HB |
5.90×103 | PSME4 | 22 |
| SKP1 |
1.16×107 | TXN |
5.67×103 | UBE2D1 | 22 |
| HSPB1 |
1.05×107 | RPN2 |
5.67×103 | RPN1 | 22 |
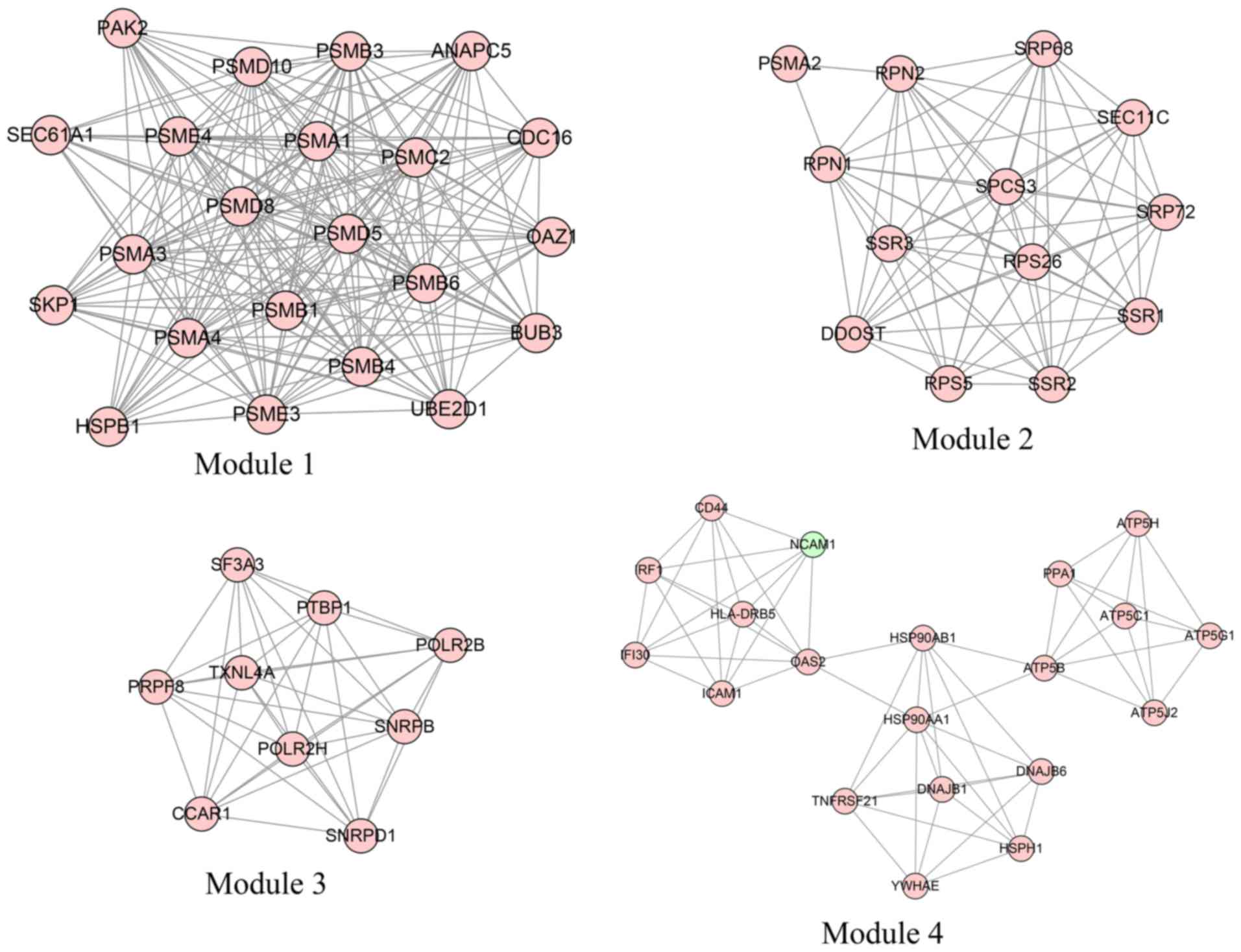
Module analysis
In total, four modules (modules 1, 2, 3 and 4) with
score >6 were detected by MCODE (Fig.
3). As shown in Table IV,
PSMA1, PSMA4 and PSMB1 were hub nodes with higher
node degrees in module 1, and ribophorin 1 (RPN1),
ribophorin 2 (RPN2) and
dolichyl-diphosphooligosaccharide-protein glycosyltransferase
subunit (DDOST) were hub nodes in module 2. The hub nodes
with higher node degrees in module 3 were pre-mRNA processing
factor 8 (PRPF8), small nuclear ribonucleoprotein D1
polypeptide (SNRPD1; 16 kDa) and small nuclear
ribonucleoprotein polypeptides B and B1 (SNRPB), and the hub
nodes in module 4 were HSP90AA1, ATP synthase, H+
transporting, mitochondrial Fo complex, Subunit F2 (ATP5J2)
and ATP synthase, H+ transporting, mitochondrial F1
complex and beta polypeptide (ATP5B). Furthermore, the
proteasome pathway was identified as the most significant pathway
in module 1.
| Table IV.KEGG pathway enriched by
differentially expressed genes in different modules
(P<0.05). |
Table IV.
KEGG pathway enriched by
differentially expressed genes in different modules
(P<0.05).
| Term | Description | P-value | Genes |
|---|
| Module1 |
|
|
|
|
hsa03050 | Proteasome |
6.77×10−17 | PSMB4, PSMA1,
PSMB6, PSMB1, PSMC2, PSMA4, PSMB3, PSMA3, PSME3, PSME4,
PSMD8 |
|
hsa04110 | Cell cycle |
9.03×10−3 | ANAPC5, SKP1,
CDC16, BUB3 |
|
hsa04120 | Ubiquitin mediated
proteolysis |
1.16×10−2 | ANAPC5, SKP1,
CDC16, UBE2D1 |
| Module2 |
|
|
|
|
hsa00510 | N-Glycan
biosynthesis |
1.63×10−3 | RPN1, RPN2,
DDOST |
|
hsa03060 | Protein export |
1.10×10−2 | SRP68,
SRP72 |
| Module3 |
|
|
|
|
hsa03040 | Spliceosome |
5.19×10−6 | PRPF8, SNRPD1,
SNRPB, TXNL4A, SF3A3 |
|
hsa03020 | RNA polymerase |
3.26×10−2 | POLR2H,
POLR2B |
| Module4 |
|
|
|
|
hsa00190 | Oxidative
phosphorylation |
1.68×10−5 | ATP5J2, ATP5B,
ATP5C1, ATP5G1, ATP5H, PPA1 |
|
hsa04612 | Antigen processing
and presentation |
1.34×10−3 | HSP90AB1,
HSP90AA1, IFI30, HLA-DRB5 |
|
hsa05012 | Parkinson's
disease |
4.63×10−3 | ATP5B, ATP5C1,
ATP5G1, ATP5H |
|
hsa05010 | Alzheimer's
disease |
9.07×10−3 | ATP5B, ATP5C1,
ATP5G1, ATP5H |
|
hsa05016 | Huntington's
disease |
1.19×10−2 | ATP5B, ATP5C1,
ATP5G1, ATP5H |
|
hsa04514 | Cell adhesion
molecules |
5.00×10−2 | NCAM1, ICAM1,
HLA-DRB5 |
Discussion
In the present study, the gene expression patterns
obtained from the GEO database revealed a total of 819 genes,
including 759 upregulated and 60 downregulated genes, that were
differently expressed in peri-implantitis samples compared with
controls. The results of the present study demonstrated that
HSP90AA1, which had the highest degrees in the PPI network,
was significantly identified in module 4. In addition, NFKB1
were also hub nodes with higher betweenness values in the PPI
network. Moreover, the proteasome pathway was the most significant
pathway in module 1, and may be key mechanisms associated with
peri-implantitis progression.
HSP90, a member of the heat shock family of
proteins, is essential in determining cell cycle control and
survival, hormone and a number of signaling pathways (24). In addition, released HSP90, which
functions as a danger signal, can elicit secretion of inflammatory
cytokines (25). A cell-impermeable
HSP90 inhibitor can prevent inflammatory responses, indicating that
extracellular HSP90 is involved in mediating and initiating sterile
inflammatory responses (26).
Moreover, previous data indicate that HSP90-targeted agents may be
helpful for the treatment of inflammatory disease, and that an
HSP90 inhibitor can affect multiple signaling processes associated
with inflammation (27).
Furthermore, inhibition of HSP90 is able to reduce innate immunity
responses and diminishes proinflammatory mediator production in
immune-stimulated macrophages (28,29). In
addition, analysis of inflammatory mediators in crevicular fluid
can be used to distinguish peri-implantitis from normal tissue
(30).
Systemic markers of inflammation are increased in
patients with peri-implantitis (30)
and high levels of inflammatory cytokines, such as IL-1β, are
associated with signs of early peri-implantitis development
(31–33). Furthermore, it has been suggested
that fibroblasts express HSP90 that also participates in the
pathogenesis of peri-implantitis (7,34).
Moreover, HSPs together with vascular and inflammatory biomarkers
may be useful as biomarkers of peri-implantitis development
(35,36). In the present study, HSP90AA1,
a hub node with higher node degrees in module 4, was enriched in
antigen processing and the presentation pathway, as well as the
NOD-like receptor-signaling pathway. Therefore, the results of the
present study are in line with results from former previous studies
and indicate that HSP90AA1 may be directly or indirectly
important in peri-implantitis development.
In the present study, NFKB1 was shown to also
have hub nodes with higher betweenness values in the PPI network.
The NFKB1 gene is known to encode the NF-KB p105/p50
isoforms (37). The central
pathological pattern of peri-implantitis is inflammatory
osteoclastogenesis, which is mediated by proinflammatory mediators
and performed by the regulators of osteoclastogenesis, including
NF-B, sRANKL and OPG (38,39). A prior study indicated that the NF-B
concentration increased 1.5–4-fold in patients with
peri-implantitis compared to those with mucositis (6). Moreover, recent studies demonstrated
that high levels of NF-B were associated with peri-implantitis
(40,41). In addition, NF-B is upregulated in
inflammatory bowel disease and is a transcription regulator of the
immune response (42). Zou et
al (43) reported that NFKB1
could regulate the transcription of genes in the immune response,
and was also a key part in coordinating the immune system. Numerous
studies have indicated that the NFKB1-94ins/del ATTG promoter
polymorphism is associated with inflammatory disease (37,42,44,45).
Furthermore, as mentioned in the aforementioned paragraph,
inflammation is associated with the pathogenesis of
peri-implantitis. Therefore, the results of the present study are
in accordance with those from previous studies and suggest that
NFKB1 may be a key gene associated with peri-implantitis
development. Notably in the current study, HSP90AA1 and
NFKB1 were enriched in the NOD-like receptor-signaling pathway.
Although the important roles of the NOD-like receptor-signaling
pathway have not been fully discussed, it is speculated that
HSP90AA1 and NFKB1 may be involved in peri-implantitis
progression via the NOD-like receptor-signaling pathway.
In addition to the two aforementioned genes, the
proteasome pathway containing the proteasome subunits (PSM)
A1, PSMA3, PSMA4, PSMB1, PSMB3,
PSMB4, PSMB6, PSMC2, PSMD8,
PSME3 and PSME4, were found to be enriched in the
most significant pathway in module 1 and to have the highest
P-value. Elliott et al (46)
indicated that key proteins modulated by the proteasome were
involved in controlling inflammatory processes. A prior study
demonstrated that the activation of the ubiquitin-proteasome by
macrophages is involved in an NF-B-dependent increase in
inflammation (47). Moreover,
previous studies have indicated that the proteasomal pathway as a
targeting goal may be a potential way to treat autoimmune and
inflammatory diseases (48–50). In addition, Wang and Maldonado
(51) demonstrated that proteasome
inhibitors could serve as novel drugs by regulating proinflammatory
protein catabolism. Furthermore, as stated above, inflammation has
been associated with the pathogenesis of peri-implantitis. Finally,
the PSMA1, PSMA3, PSMA4, PSMB1,
PSMB3, PSMB4, PSMB6, PSMC2,
PSMD8, PSME3 and PSME4 proteins are essential
subunits that contribute to the assembly of the proteasome complex
(52–55). Therefore, the proteasome and its
subunit genes may be important in peri-implantitis progression.
In conclusion, HSP90AA1, NFKB1 and the
NOD-like receptor signaling pathway, as well as the proteasome
pathway and its subunits genes may be important in the development
of peri-implantitis. However, there were certain limitations in the
present study, such as no experimental verification and the
relatively small sample size used. Therefore, further research
investigating the potential mechanisms involved in peri-implantitis
are required.
References
|
1
|
Schminke B, Vom Orde F, Gruber R,
Schliephake H, Bürgers R and Miosge N: The pathology of bone tissue
during peri-implantitis. J Dent Res. 94:354–361. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mombelli A: Microbiology and antimicrobial
therapy of peri-implantitis. Periodontol 2000. 28:177–189. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Berglundh T, Persson L and Klinge B: A
systematic review of the incidence of biological and technical
complications in implant dentistry reported in prospective
longitudinal studies of at least 5 years. J Clin Periodontol.
29:(Suppl 3). 197–212, 232–233. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Becker ST, Beck-Broichsitter BE, Graetz C,
Dörfer CE, Wiltfang J and Häsler R: Peri-implantitis versus
periodontitis: Functional differences indicated by transcriptome
profiling. Clin Implant Dent Relat Res. 16:401–411. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Theocharis AD, Seidel C, Borset M, Dobra
K, Baykov V, Labropoulou V, Kanakis I, Dalas E, Karamanos NK,
Sundan A and Hjerpe A: Serglycin constitutively secreted by myeloma
plasma cells is a potent inhibitor of bone mineralization in vitro.
J Biol Chem. 281:35116–35128. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rakic M, Struillou X, Petkovic-Curcin A,
Matic S, Canullo L, Sanz M and Vojvodic D: Estimation of bone loss
biomarkers as a diagnostic tool for peri-implantitis. J
Periodontol. 85:1566–1574. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bordin S, Flemmig TF and Verardi S: Role
of fibroblast populations in peri-implantitis. Int J Oral
Maxillofac Implants. 24:197–204. 2009.PubMed/NCBI
|
|
8
|
Irshad M, Scheres N, Anssari Moin D,
Crielaard W, Loos BG, Wismeijer D and Laine ML: Cytokine and matrix
metalloproteinase expression in fibroblasts from peri-implantitis
lesions in response to viable Porphyromonas gingivalis. J
Periodontal Res. 48:647–656. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Casado PL, Canullo L, de Almeida Filardy
A, Granjeiro JM, Barboza EP and Leite Duarte ME: Interleukins 1β
and 10 expressions in the periimplant crevicular fluid from
patients with untreated periimplant disease. Implant Dent.
22:143–150. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park SY, Kim KH, Gwak EH, Rhee SH, Lee JC,
Shin SY, Koo KT, Lee YM and Seol YJ: Ex vivo bone morphogenetic
protein 2 gene delivery using periodontal ligament stem cells for
enhanced re-osseointegration in the regenerative treatment of
peri-implantitis. J Biomed Mater Res A. 103:38–47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: affy-analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carvalho BS and Irizarry RA: A framework
for oligonucleotide microarray preprocessing. Bioinformatics.
26:2363–2367. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene Ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Altermann E and Klaenhammer TR:
PathwayVoyager: pathway mapping using the Kyoto Encyclopedia of
Genes and Genomes (KEGG) database. BMC Genomics. 6:602005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
da Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2008. View Article : Google Scholar
|
|
17
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43:D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jeong H, Mason SP, Barabási AL and Oltvai
ZN: Lethality and centrality in protein networks. Nature.
411:41–42. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Estrada E and Rodríguez-Velázquez JA:
Subgraph centrality in complex networks. Phys Rev E Stat Nonlin
Soft Matter Phys. 71:0561032005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goh KI, Oh E, Kahng B and Kim D:
Betweenness centrality correlation in social networks. Phys Rev E
Stat Nonlin Soft Matter Phys. 67:0171012003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Csardi G and Nepusz T: The igraph software
package for complex network research. Inter Journal Complex
Systems. 1695:1–9. 2006.
|
|
23
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bucci M, Roviezzo F, Cicala C, Sessa WC
and Cirino G: Geldanamycin, an inhibitor of heat shock protein 90
(Hsp90) mediated signal transduction has anti-inflammatory effects
and interacts with glucocorticoid receptor in vivo. Br J Pharmacol.
131:13–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wallin RP, Lundqvist A, Moré SH, von Bonin
A, Kiessling R and Ljunggren HG: Heat-shock proteins as activators
of the innate immune system. Trends Immunol. 23:130–135. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qin S, Ni M, Wang X, Maurier-Mahé F,
Shurland DL and Rodrigues GA: Inhibition of RPE cell sterile
inflammatory responses and endotoxin-induced uveitis by a
cell-impermeable HSP90 inhibitor. Exp Eye Res. 93:889–897. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rice JW, Veal JM, Fadden RP, Barabasz AF,
Partridge JM, Barta TE, Dubois LG, Huang KH, Mabbett SR, Silinski
MA, et al: Small molecule inhibitors of Hsp90 potently affect
inflammatory disease pathways and exhibit activity in models of
rheumatoid arthritis. Arthritis Rheum. 58:3765–3775. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
De Nardo D, Masendycz P, Ho S, Cross M,
Fleetwood AJ, Reynolds EC, Hamilton JA and Scholz GM: A central
role for the Hsp90· Cdc37 molecular chaperone module in
interleukin-1 receptor-associated-kinase-dependent signaling by
Toll-like receptors. J Biol Chem. 280:9813–9822. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shimp SK III, Parson CD, Regna NL, Thomas
AN, Chafin CB, Reilly CM and Rylander Nichole M: HSP90 inhibition
by 17-DMAG reduces inflammation in J774 macrophages through
suppression of Akt and nuclear factor-κB pathways. Inflamm Res.
61:521–533. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hultin M, Gustafsson A, Hallström H,
Johansson LÅ, Ekfeldt A and Klinge B: Microbiological findings and
host response in patients with peri-implantitis. Clin Oral Implants
Res. 13:349–358. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tokoro Y, Yamamoto T and Hara K: IL-1 beta
mRNA as the predominant inflammatory cytokine transcript:
Correlation with inflammatory cell infiltration into human gingiva.
J Oral Pathol Med. 25:225–231. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Panagakos FS, Aboyoussef H, Dondero R and
Jandinski JJ: Detection and measurement of inflammatory cytokines
in implant crevicular fluid: A pilot study. Int J Oral Maxillofac
Implants. 11:794–799. 1996.PubMed/NCBI
|
|
33
|
Aboyoussef H, Carter C, Jandinski JJ and
Panagakos FS: Detection of prostaglandin E2 and matrix
metalloproteinases in implant crevicular fluid. Int J Oral
Maxillofac Implants. 13:689–696. 1998.PubMed/NCBI
|
|
34
|
Lee SS, Jang JH, Kim KS, Yoo YJ, Kim YS
and Lee SK: Failure of bone regeneration after demineralized bone
matrix allograft in human maxillary sinus floor elevation. Basic
Appl Pathol. 2:125–130. 2009. View Article : Google Scholar
|
|
35
|
Bullon P, Fioroni M, Goteri G, Rubini C
and Battino M: Immunohistochemical analysis of soft tissues in
implants with healthy and peri-implantitis condition, and
aggressive periodontitis. Clin Oral Implants Res. 15:553–559. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Borsani E, Salgarello S, Stacchiotti A,
Mensi M, Boninsegna R, Ricci F, Zanotti L, Rezzani R, Sapelli P,
Bianchi R and Rodella LF: Altered immunolocalization of heat-shock
proteins in human peri-implant gingiva. Acta Histochem.
109:221–227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Borm ME, van Bodegraven AA, Mulder CJ,
Kraal G and Bouma G: A NFKB1 promoter polymorphism is involved in
susceptibility to ulcerative colitis. Int J Immunogenet.
32:401–405. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cochran DL: Inflammation and bone loss in
periodontal disease. J Periodontol. 79:(8 Suppl). 1569–1576. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Theoleyre S, Wittrant Y, Tat SK, Fortun Y,
Redini F and Heymann D: The molecular triad OPG/RANK/RANKL:
Involvement in the orchestration of pathophysiological bone
remodeling. Cytokine Growth Factor Rev. 15:457–475. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rakić M, Nikolić-Jakoba N, Struillout X,
Petković-Curcin A, Stamatović N, Matić S, Janković S, Aleksić Z,
Vasilić D, Leković V and Vojvodić D: Receptor activator of nuclear
factor kappa B (RANK) as a determinant of peri-implantitis.
Vojnosanit Pregl. 70:346–351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rakic M, Lekovic V, Nikolic-Jakoba N,
Vojvodic D, Petkovic-Curcin A and Sanz M: Bone loss biomarkers
associated with peri-implantitis. A cross-sectional study. Clin
Oral Implants Res. 24:1110–1116. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Karban AS, Okazaki T, Panhuysen CI,
Gallegos T, Potter JJ, Bailey-Wilson JE, Silverberg MS, Duerr RH,
Cho JH, Gregersen PK, et al: Functional annotation of a novel NFKB1
promoter polymorphism that increases risk for ulcerative colitis.
Hum Mol Genet. 13:35–45. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zou YF, Wang F, Feng XL, Tao JH, Zhu JM,
Pan FM and Su H: Association of NFKB1-94ins/delATTG promoter
polymorphism with susceptibility to autoimmune and inflammatory
diseases: A meta-analysis. Tissue antigens. 77:9–17. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li H, Gao L, Shen Z, Li CY, Li K, Li M, Lv
YJ, Li CX, Gao TW and Liu YF: Association study of NFKB1 and SUMO4
polymorphisms in Chinese patients with psoriasis vulgaris. Arch
Dermatol Res. 300:425–433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kosoy R and Concannon P: Functional
variants in SUMO4, TAB2, and NFkappaB and the risk of type 1
diabetes. Genes Immun. 6:231–235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Elliott PJ, Zollner TM and Boehncke WH:
Proteasome inhibition: A new anti-inflammatory strategy. J Mol Med
(Berl). 81:235–245. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Marfella R, D'Amico M, Di Filippo C, Baldi
A, Siniscalchi M, Sasso FC, Portoghese M, Carbonara O, Crescenzi B,
Sangiuolo P, et al: Increased activity of the ubiquitin-proteasome
system in patients with symptomatic carotid disease is associated
with enhanced inflammation and may destabilize the atherosclerotic
plaque: Effects of rosiglitazone treatment. J Am Coll Cardiol.
47:2444–2455. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Goldberg AL and Rock K: Not just research
tools-proteasome inhibitors offer therapeutic promise. Nat Med.
8:338–340. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lecker SH, Goldberg AL and Mitch WE:
Protein degradation by the ubiquitin-proteasome pathway in normal
and disease states. J Am Soc Nephrol. 17:1807–1819. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ciechanover A: The ubiquitin proteolytic
system and pathogenesis of human diseases: A novel platform for
mechanism-based drug targeting. Biochem Soc Trans. 31:474–481.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang J and Maldonado MA: The
ubiquitin-proteasome system and its role in inflammatory and
autoimmune diseases. Cell Mol Immunol. 3:255–261. 2006.PubMed/NCBI
|
|
52
|
Nothwang HG, Tamura T, Tanaka K and
Ichihara A: Sequence analyses and inter-species comparisons of
three novel human proteasomal subunits, HsN3, HsC7-I and HsC10-II,
confine potential proteolytic active-site residues. Biochim Biophys
Acta. 1219:361–368. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gallastegui N and Groll M: The 26S
proteasome: Assembly and function of a destructive machine. Trends
Biochem Sci. 35:634–642. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Le Tallec B, Barrault MB, Courbeyrette R,
Guérois R, Marsolier-Kergoat MC and Peyroche A: 20S proteasome
assembly is orchestrated by two distinct pairs of chaperones in
yeast and in mammals. Mol Cell. 27:660–674. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Murata S, Yashiroda H and Tanaka K:
Molecular mechanisms of proteasome assembly. Nat Rev Mol Cell Biol.
10:104–115. 2009. View Article : Google Scholar : PubMed/NCBI
|