Introduction
Cyclophosphamide (CP) exhibits positive therapeutic
effects in clinical practice; however, it also causes undesirable
harm to normal organs and tissues, such as the testes (1). Previous studies (2–4) have
indicated that a large number of free radicals and oxidation
products are produced in mice after intraperitoneal injection of
CP, which may cause testicular oxidative stress resulting in DNA
double-strand breaks and germ cell apoptosis. Intracellular
accumulation of excess reactive oxygen species (ROS) can induce
apoptosis, resulting in programmed cell death (5). Apoptosis, a mode of programmed cell
death, has important roles in cell growth, differentiation and
development, and acts as an effective way to induce tumor cell
death. Typically, apoptosis is targeted in cancer treatment as an
anti-tumor mechanism (6,7). The specific mechanisms involved in
apoptosis are yet to be fully understood. The majority of studies
suggest that multiple mediators of cell damage, which are triggered
by adverse exogenous factors, are closely related to apoptotic
cells, including oxygen free radicals, radiation and chemotherapy
(8–10).
Squid ink polysaccharide (SIP), a natural marine
product, is considered to be a potentially effective, non-toxic,
broad-spectrumcy to protective agent due to its anti-oxidant
(1), anti-tumor (11) and anti-chemotherapy (1,12–16)
functions. SIP has been identified as a type of glycosaminoglycan,
with a unique structure: [3GlcAβ1-4(GalNAc α1-3)-Fucα1]n
(17). In previous studies, SIP
alleviated toxicity of the chemotherapeutic drug, CP, on the heart,
liver, spleen, lung, kidney and intestines (12–16,18).
There is also evidence of the protective effect of SIP on the male
reproductive system. In a previous study, SIP effectively reduced
testicular pathological damage caused by CP in mice, thus
increasing testicular weight and sperm quantity, reducing sperm
deformity rates, improving testis iconic enzyme activity and redox
homeostasis, and maintaining natural hormone levels in serum and
testes (1). Although the
preventative effects of SIP towards CP-induced damage of testes and
apoptosis in murine germ cells have been identified (10), its regulative processes remain
unclear. To elucidate the preventive mechanisms of SIP on the
testes of mice exposed to CP, apoptosis of testicular germ cells in
mice treated with CP and SIP was investigated in the present
study.
Materials and methods
Preparation of SIP
Fresh squid (Sepia esculenta) were caught
from the East coast of Beibu Gulf. Their ink sacs were removed by
the fishermen and stored at −70°C in an ultra-low temperature
freezer. SIP was prepared with a slightly modified method as
described by Chen et al (17). Briefly, the frozen squid ink was
thawed at 4°C, diluted with an equal volume of PBS (0.01 mol/l; pH
7.4) and treated by sonication in an ice bath. Following storage at
4°C for >8 h, the mixture was centrifuged (7,155 × g) at 4°C for
50 min. The supernatant was collected and hydrolyzed with papain
(1.5%) at 50°C for 90 min and heated in boiling water to denature
the protease. Proteins in the treated supernatant were removed by
the Sevag method (19). The aqueous
phase was mixed with four volumes of ethanol to precipitate the
polysaccharides. Crude polysaccharides were obtained from the
precipitate and subsequently separated into three fractions by
DEAE-52 cellulose column chromatography (GE Healthcare Life
Sciences, Chalfont, UK). The first fraction, in which the peak area
was far larger than the others, was collected, dialyzed,
concentrated, and further purified in a Sephacryl S-300HR column
(GE Healthcare Life Sciences). One elution peak was obtained from a
S-300HR column and the fraction from that peak was named SIP. The
collected SIP was dialyzed, concentrated, freeze-dried and stored
at −20°C.
Animals and experimental design
A total of 40 sexually mature male Kunming mice
(weighing 25±2 g), aged 6 weeks, were purchased from the
Experimental Animal Centre of Guangxi Medical University (Nanning,
China; SCXK(Gui) 2009-002). Mice were adaptively domesticated for
one week under the following constant experimental conditions:
Relative humidity, 55±5%; temperature, 22±2°C; quasi-diurnal cycle
of 12 h light and 12 h darkness; and ad libitum access to
food and water.
A total of 40 mice were randomly divided into four
equal experimental groups: Control group (CON; orally administered
and abdominally injected with normal saline); CP-treated group (CP;
orally administered normal saline and abdominally injected with CP
in normal saline); SIP-treated group (SIP; orally administered SIP
and abdominally injected with normal saline); and a co-treated
group (SIP+CP; orally administered SIP and abdominally injected
with CP in normal saline). SIP was administered once a day for two
weeks with 80 mg/kg body weight chosen as an optimal concentration
of SIP from our previous studies (1,20). CP
was intraperitoneally injected once on the 7th day of SIP or saline
administration; 120 mg/kg body weight was chosen as the dose to
obtain optimal effects in this model, since a large dose may result
in testicular damage due to increased cellular sensitivity when
exposed to high doses of CP (21,22).
Following treatment, all mice were sacrificed by dislocation of
cervical vertebra at 24 h after the last administration. Bilateral
testes were collected, rapidly cleared of surrounding fat and
connective tissue and stored at −70°C.
Detection of testicular oxidative
stress level
Samples were homogenised quickly in ice-cold PBS at
4°C. The homogenate was centrifuged 2,000 × g for 10 min at 4°C.
The supernatant was determined by measuring the activity of
superoxide dismutase (SOD), contents of malondialdehyde (MDA) and
reactive oxygen species (ROS) with detection kits according to
manufacturers' protocols. SOD assay kit (Hydroxylamine method, cat.
no. A001-1), ROS assay kit (Dichlorofluorescein method, cat. no.
E004) and MDA assay kit (TBA method, cat. no. A003-1) were
purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China).
Transmission electron microscopy
detection
Testes were fixed with 5% glutaraldehyde in PBS
followed by post-fixation with 1% osmic acid formulated with PBS.
Following washing three times with PBS, the tissue was dehydrated
with ethanol and acetone, embedded in Epon812 and sectioned into
60–70 nm with an ultra-microtome. Sections were stained with 2%
uranyl acetate and lead citrate and the ultra-structure of
testicular germ cells was observed by transmission electron
microscopy.
Terminal-deoxynucleoitidyl
transferase-mediated nick-end labelling (TUNEL) assay
TUNEL assay was performed according to the
manufacturer's instructions, Situ TUNEL apoptosis detection kit was
purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China). Procedure was as follows: De-waxed and hydrated testes
paraffin sections were washed three times with PBS and incubated in
DNase-free Proteinase K for 30 min at 37°C. Following washing,
sections were immersed in methanol solution at room temperature for
5 min, the excess liquid was removed with filter paper and the
sample was incubated in TdT enzyme reaction solution for 60 min at
37°C prior to washing with PBS. Excess liquid was removed with
filter paper and the sections were incubated in Streptavidin-HRP
solution at 37°C for 30 min in the dark. Following washing, each
section was stained with 3,3′-diaminobenzidinesolution at room
temperature for 10 min and washed with PBS. The sections were
observed under an optical microscope (magnification, ×200), and
brown particles were deemed to be indicative of apoptosis-positive
cells. The number of positive cells per 100 germ cells was
calculated as the positive rate of apoptosis, which indicated the
percentage of apoptosis-positive cells in the total cell count.
Western blot analysis
Western blot was used to assess the levels of B-cell
lymphoma 2 (Bcl2), Bcl2-associated X (Bax), Caspase 3, LC3B,
Beclin-1, phospho-p38, phospho-Akt and β-actin. Samples were
homogenized in a 1 ml/0.1 g tissue RIPA lysis buffer with 1 mM
phenylmethyylsulfonyl fluoride (Beyotime Institute of
Biotechnology, Shanghai, China) in a homogenizer. Following lysis
for 20 min on ice and centrifugation at 10,000 × g for 3 min at
4°C, the protein content was determined using the Enhanced BCA
Protein Assay kit (Beyotime Institute of Biotechnology). All the
resulting extracts were adjusted to the same concentration and
electrophoresed on a 10% SDS-PAGE. The loading quantity was 40 µg.
The separated proteins were then transferred onto polyvinylidene
difluoride membranes. The membranes were blocked by incubation with
a phosphate-buffered saline and Tween-20 (PBST) buffer (0.05%
Tween-20, 100 mM NaCl, 10 mM Tris-HCl pH 7.4), containing 10%
nonfat dry milk for 2 h, then incubated with one of the following
antibodies: Caspase 3 rabbit polyclonal antibody (cat. no.
19677-1-AP, Proteintech Group, Inc., Chicago, IL, USA), Bax rabbit
polyclonal antibody (cat. no. 50599-2-Ig; Proteintech Group, Inc.),
Bcl-2 rabbit polyclonal antibody (cat. no. 12789-1-AP; Proteintech
Group, Inc.), Phospho-p38 MAPK rabbit monoclonal antibody (cat. no.
4092S; Cell Signaling Technology, Inc., Danvers, MA, USA),
Akt-phospho-S473 mouse monoclonal antibody (cat. no. 66444-1-Ig;
Proteintech Group, Inc.), beta actin rabbit polyclonal antibody
(cat. no. 20536-1-AP; Proteintech Group, Inc.), beta actin mouse
monoclonal antibody (cat. no. 60008-1-Ig; Proteintech Group, Inc.),
at concentrations of 1:2,000. They were then washed three times
with 0.05% Tween-20 in PBST for 5 min by slowly shaking and finally
incubated for 2 h with anti-rabbit IgG (H+L) secondary antibody
(KPL, Inc., Gathersburg, MD, USA) at concentrations of 1:2,000. The
membranes were washed with PBST and the proteins were detected with
a substrate super ECL Plus solution (Applygen Technologies Inc.,
Beijing, China). All these steps were carried out at room
temperature. Densitometric analysis of digitalized images was done
with the Image J software (National Institutes of Health, Bethesda,
MD, USA).
Statistical analysis
All experimental data were presented as mean ±
standard deviation. Data were analysed using JMP 7.0.2 (SAS
Institute Inc., Cary, NC, USA). One-way analysis of variance and
the post-hoc Tukey's honest significant difference test were used
to evaluate differences between groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
SIP inhibits CP-induced oxidative
stress in mice testes
ROS levels, SOD activity and MDA content in testes
are shown in Table I. Significant
increases in ROS level and MDA content, and a reduction in SOD
activity were observed in mice exposed to CP (P<0.05). After SIP
treatment, SOD activity of CP-treated mice was obviously higher
(P<0.05), ROS levels and MDA content were also greatly reduced
(P<0.05). To sum up, its preventive effects on the testes of
CP-treated mice were found by evaluating the three indicators,
which implied marked inhibition of oxidative stress in CP-exposed
testes following treatment with SIP.
 | Table I.ROS level, MDA content and SOD
activity in testes. |
Table I.
ROS level, MDA content and SOD
activity in testes.
| Group | ROS (a.u./mg) | SOD (U/mg) | MDA (nmol/m) |
|---|
| CON |
32.39±5.76 |
101.17±5.34a |
11.99±3.80 |
| CP |
46.74±5.37a,b |
76.48±5.70 |
21.90±2.36a,b |
| SIP+CP |
31.42±5.35 |
99.62±4.51b |
12.10±3.89 |
| SIP |
29.87±4.84 |
96.95±6.49 |
12.21±4.41 |
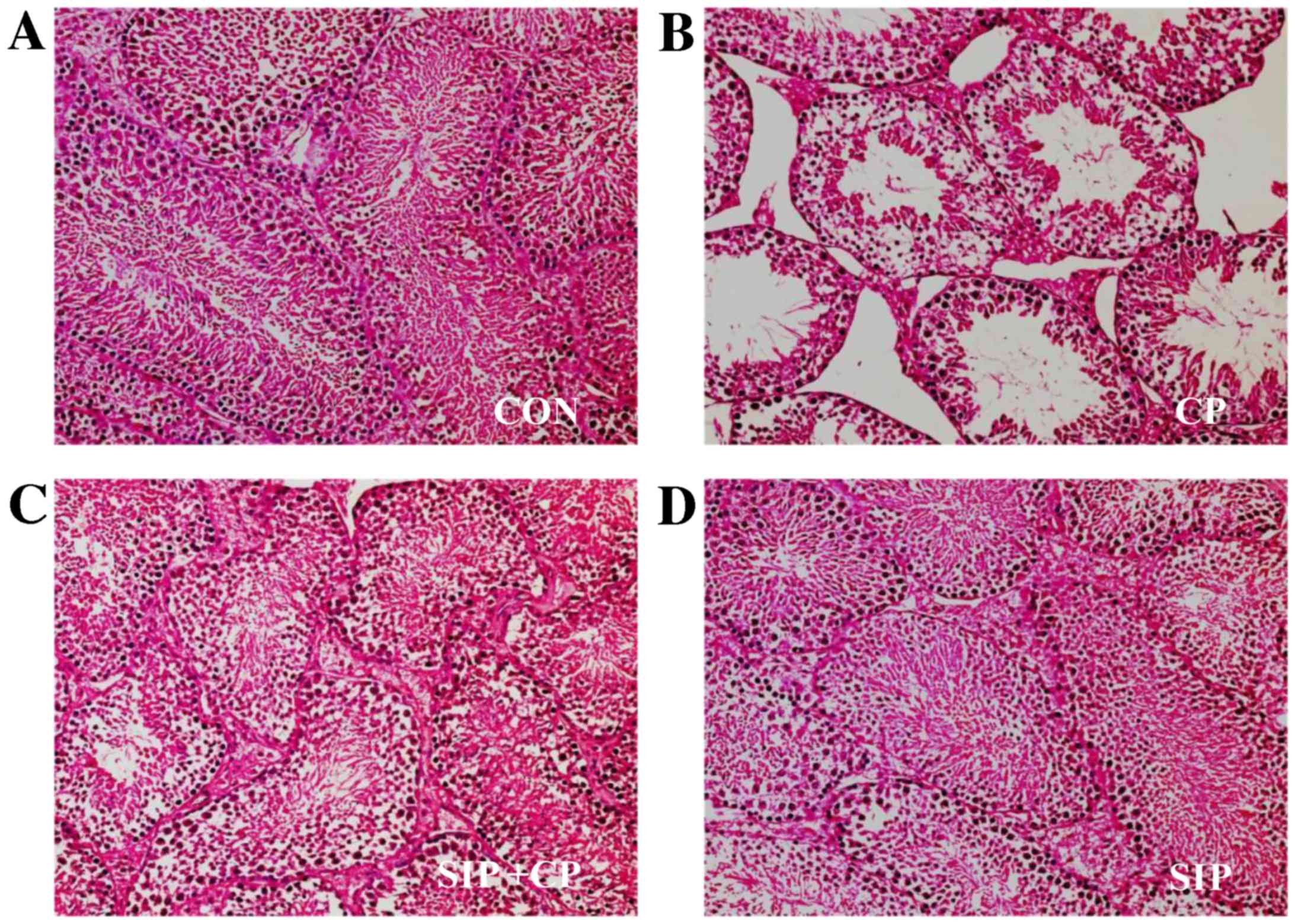
SIP prevents CP-induced damage of
histological structures in mice testes
As demonstrated in Fig.
1, testicular tissues of vehicle- and SIP-treated mice
exhibited normal histological structures, including the correct
epithelial thickness of the seminiferous tubule, plentiful layers
of cells, compactness and regularity of spermatogenic cells, as
well as rich capillaries inthe periphery of the seminiferous
tubule. However, the histological structure of the CP-treated group
showed reduced epithelial thickness of the seminiferous tubule and
fewer layers of cells. Furthermore, spermatogenic cell counts
decreased and their structures were disarranged; in addition, few
capillaries were observed in the enlarged space between
seminiferous tubules. In the group co-treated with SIP and CP,
histological structure was visibly better than that of CP-treated
mice, as indicated by comprehensive assessment of the
aforementioned parameters.
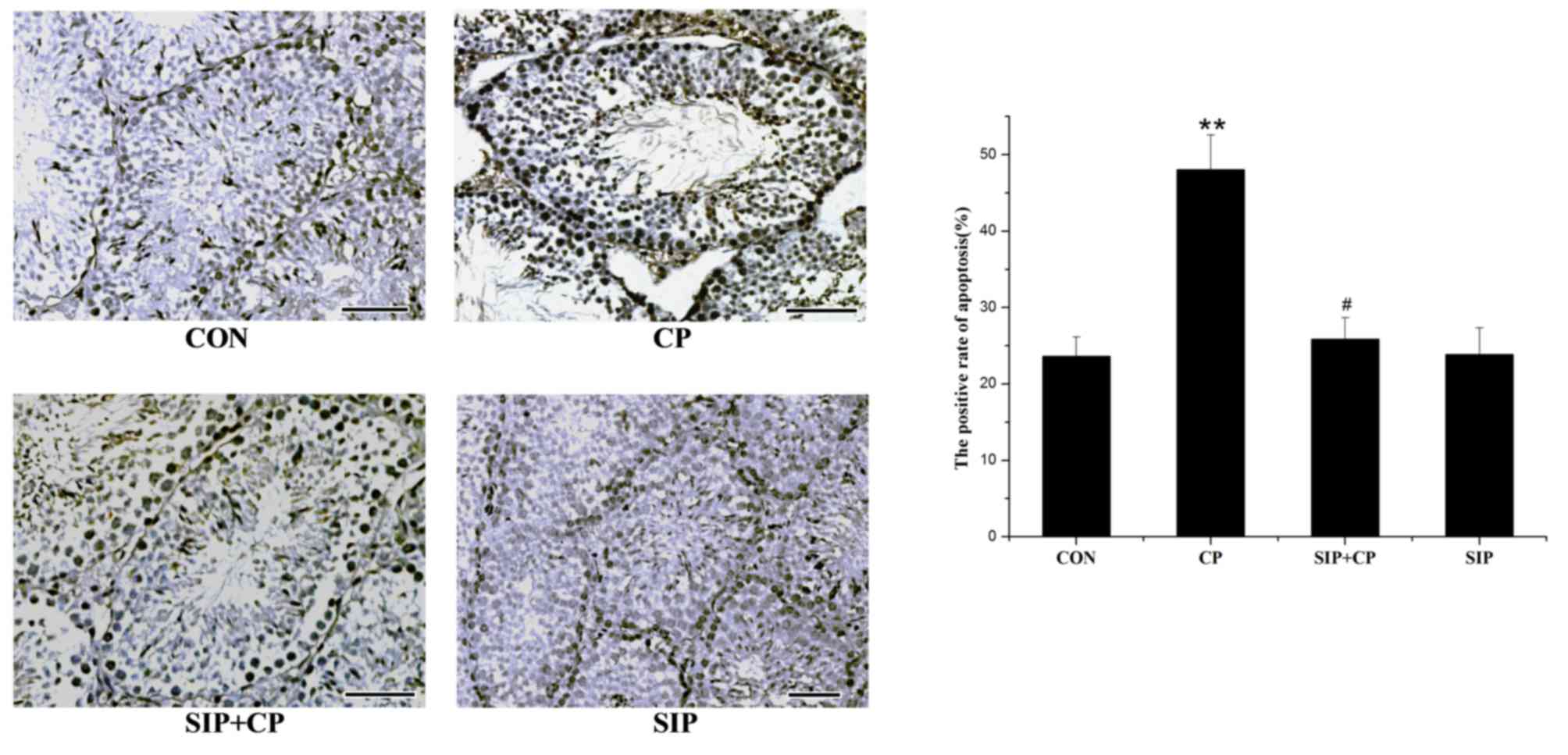
SIP prevents CP-induced apoptosis of
testicular germ cells
TUNEL was used to detect DNA strand breaks generated
during early-stage apoptosis and to observe the direct location of
apoptosis (Fig. 2). In the control
group, few apoptotic cells were present in the cell layers of
spermatogonia. CP-treated mice exhibited a large number of
apoptotic cells distributed predominantly in the spermatogonial
layer, and the positive rate of apoptosis was significantly
increased compared with the other groups (P<0.05). Similar to
the control mice, minimal evidence of apoptosis was observed in the
cell layers of the SIP-treated mice. Compared with the CP-treated
mice, mice co-treated with SIP and CP exhibited lower levels of
apoptosis in each cell layer. These results demonstrated that the
positive rate of apoptosis was significantly decreased by SIP
(P<0.05).
Effect of SIP on the CP-mediated
expression of caspase-3, Bcl-2 and Bax genes, and phosphorylation
of p38 and Akt proteins
As shown in Fig. 3,
CP upregulated Caspase-3 protein expression (P<0.01) and
improved photophosphorylation of p38 (P<0.05) and Akt proteins
(P<0.01) in mice testes. Bcl-2 protein expression was
significantly decreased by CP (P<0.01) and the Bax/Bcl-2 ratio
was significantly increased (P<0.01). Following administration
of SIP, testes of CP-exposed mice showed a significant reduction of
photophosphorylation of p38 protein (P<0.01) and a significant
decrease in the protein expression of Caspase-3 (P<0.05), while
the expression level of the Bcl-2 gene was significantly increased
(P<0.01). A significant decrease in Bax/Bcl-2 ratio was also
observed in the SIP + CP group compared with the CP group
(P<0.05). Compared with the CP-treated group, the protein
expression of p-Akt in mice co-treated with SIP and CP decreased,
but no significant difference was observed (P>0.05).
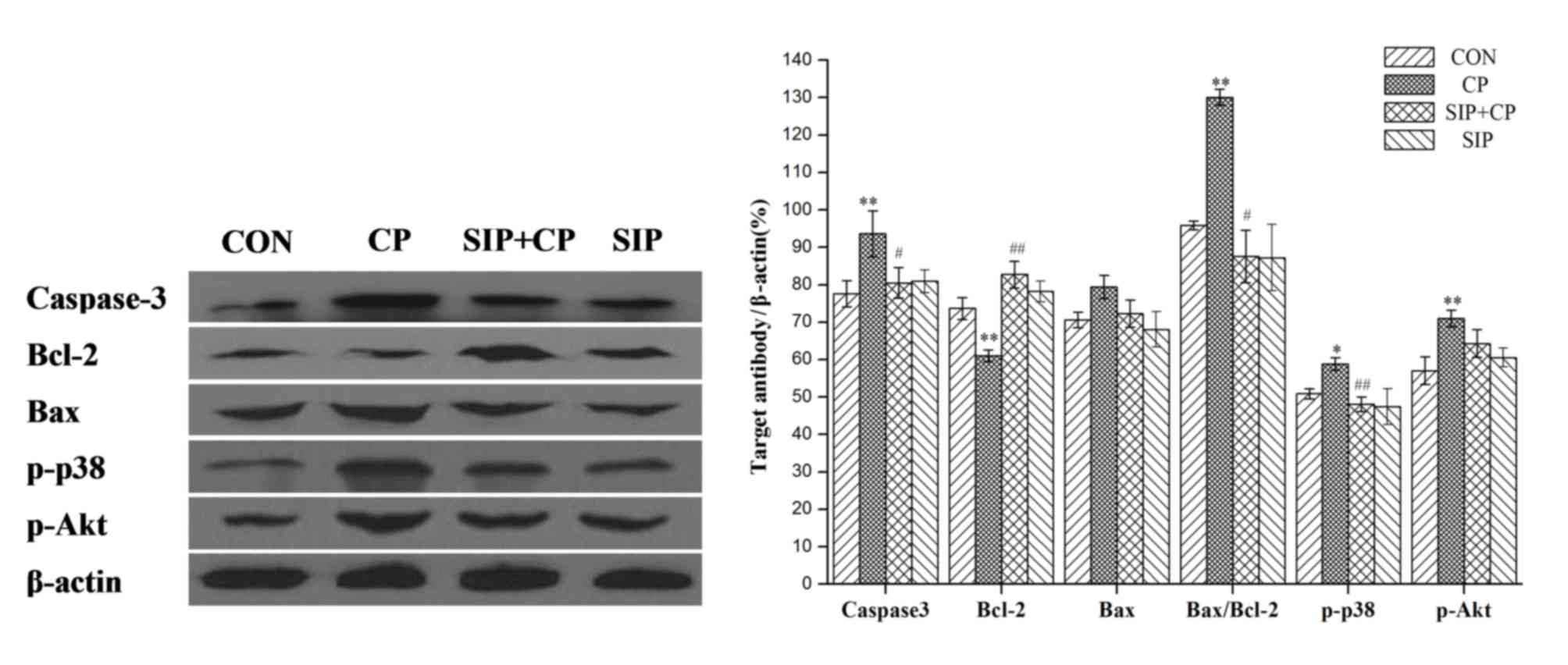 | Figure 3.Effects of SIP on CP-mediated
expression of Caspase-3, Bcl-2 and Bax genes, as well as
phosphorylation of p38 and Akt proteins. *P<0.05, **P<0.01
vs. CON. #P<0.05, ##P<0.01 vs. CP. CON,
control mice; CP, CP-treated mice; SIP+CP, mice treated with squid
ink polysaccharide and CP; SIP, squid ink polysaccharide-treated
mice; p-, phosphorylated; Akt, AKT serine/threonine kinase; Bcl-2,
B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
SIP attenuates the destruction of the
ultra-structure of testicular germ cells caused by CP
As shown in Fig. 4,
spermatogonia and Sertoli cells in control mice were healthy, as
indicated by their large nuclei, homogeneous electron density in
the nucleoplasm and prominent nucleoli. Additionally, the cytoplasm
of spermatogonia and Sertoli cells contained rod-shaped
mitochondria with substantial cristae. However, following treatment
with CP alone, spermatogonia nuclei exhibited chromatorrhexis and
chromatin pyknosis, and Sertoli cells appeared to undergo chromatin
margination. Moreover, mitochondria in the two types of cells were
swollen and displayed a lack of cristae. In some cases, spherical
cellular monolayers were evident. Under administration of SIP and
CP, spermatogonia and Sertoli cells demonstrated improved
ultrastructure, indicated by clear and evident nucleoli, the
homogenous electron density of the nucleoplasm and clear cristae
present in numerous mitochondria. In SIP-treated mice, nucleoli in
spermatogonia and Sertoli cells were clear and obvious, the
karyoplasm was light in colour and of uniform density, and the
cytoplasm had many mitochondria with rich cristae.
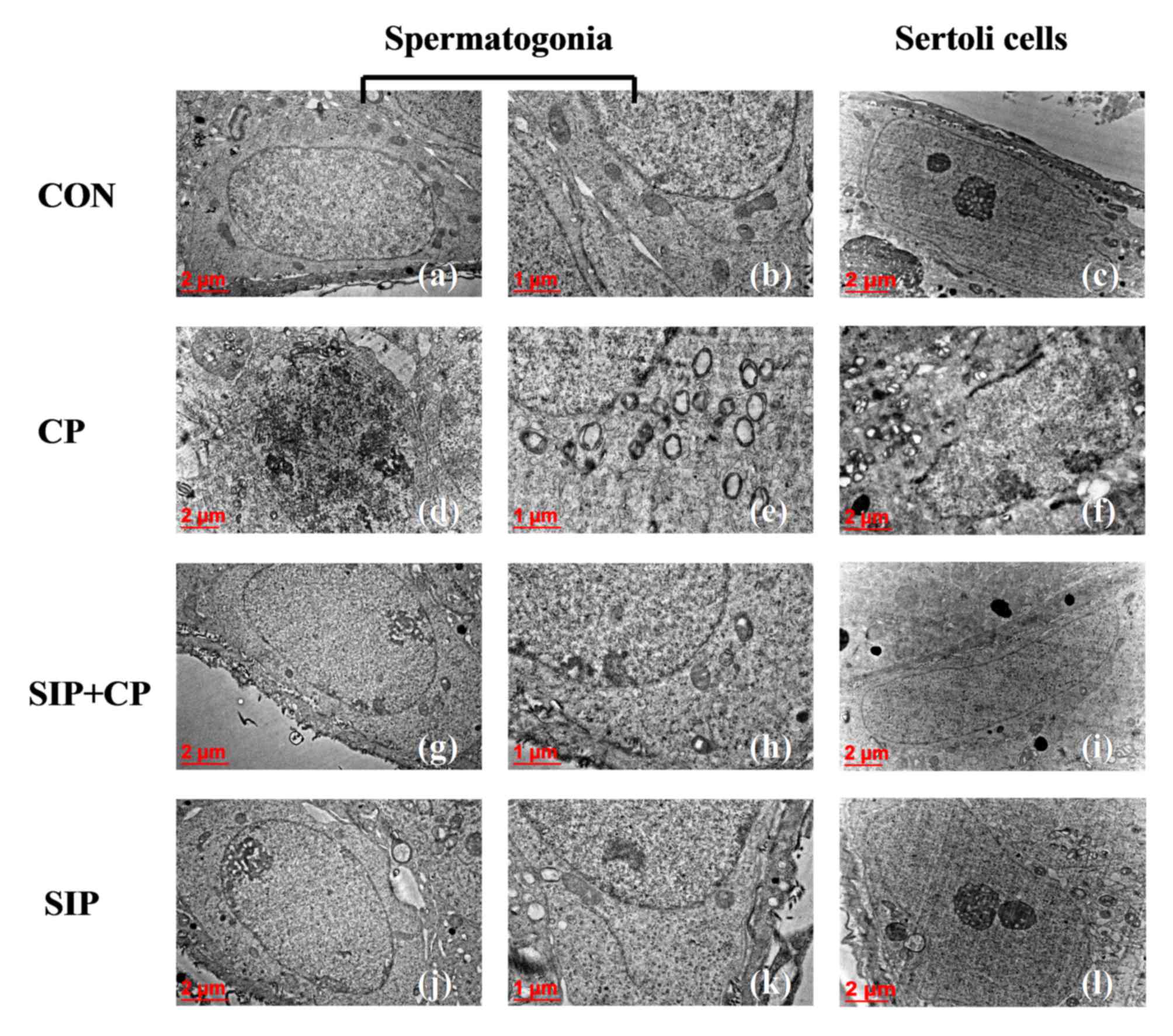 | Figure 4.Ultrastructure of testicular
spermatogonia and Sertoli cells in four groups of mice as assessed
by transmission electron microscopy. Images were obtained at
magnifications of ×12,000 (c, f, i, l), ×15,000 (a, d, g, j),
×30,000 (b, e, h, k). CON, control mice; CP, CP-treated mice;
SIP+CP, mice treated with squid ink polysaccharide and CP; SIP,
squid ink polysaccharide-treated mice. |
Discussion
As a clinical chemotherapeutic agent that is widely
used, CP inevitably has some negative effects on the normal tissues
and organs of patients with cancer, such as infertility or genetic
toxicity (23,24). Two main physiological functions of
testes is the generation of sperm and endocrine regulation, the
latter restricts the former through they interact with each other
(25,26). Apoptosis within seminiferous tubules
is necessary to maintain a dynamic equilibrium in the testes and
the generation of healthy sperm. However, chemotherapeutic drugs,
such as CP, may generate excess free radicals and oxidation
products and weaken testicular antioxidant functions, which may
destroy the oxidation-antioxidation balance system in testes and
result in testicular oxidative stress (27,28).
Oxidative stress-induced accumulation of ROS may result in serious
damage to mitochondria, including DNA mutation, lipid peroxidation
and mitochondrial membrane channel opening, which leads to reduced
mitochondrial function and male reproductive disorder (29). In the present study, CP induced
injury to testis in male mice, including excessive ROS and MDA,
decreased SOD activity and number of germ cells, as well as
hollowed seminiferous tubules. Moreover, nuclear fragmentation and
condensation, enlarged mitochondria and reduced cristae were
observed in spermatogonial cells and Sertoli cells, the number of
apoptosis-positive cells was elevated and these apoptotic cells
were predominantly distributed in spermatogonial cell layers. These
data support the CP-induced theory of oxidative stress (1,20).
Additionally, mitochondria are organelles that are highly sensitive
to stressful stimulations, oxidative stress alters mitochondrial
membrane permeability and potential, which leads to mitochondrial
swelling (29). In the present
study, mitochondria were swollen and displayed a lack of cristae in
spermatogonial cells and Sertoli cells. Therefore, cristae are
important structures involved in mitochondrial energy metabolism,
and their destruction inevitably causes mitochondrial energy
metabolism disorder (30). Thus,
based on the results of the current study, there was an association
between oxidative stress and the apoptosis in cells, with low
activities of antioxidative enzymes, nuclear pyknosis,
mitochondrial swelling and vesiculation.
The marine polysaccharide, SIP, has demonstrated
strong antioxidant capacity and it is able to effectively weaken
the toxicity of CP on the male reproductive system (1). The present findings, demonstrated that
SIP inhibited apoptosis of germ cells via improving antioxidant
ability by hindering oxidative damage in mitochondria and
suppressing activation of Caspase-3 protein in CP-exposed mice.
Furthermore, SIP depleted ROS and therefore reduced the expression
levels of pro-apoptotic Bax genes in spermatogonia and Sertoli
cells, allowing these cells to maintain their morphological
structural integrity. As a result, the blood-testis barrier was
protected and normal physiological functions in the testes were
conserved.
Multiple factors of signaling pathways are involved
in the inhibition of apoptosis. Anti-apoptotic protein, Bcl-2,
combines with Bax to form a homodimeror heterodimer that inhibits
apoptosis (31). The present study
indicated that SIP increased the content of Bcl-2 and decreased the
Bax/Bcl-2 ratio, which suggested that SIP inhibited CP-induced
apoptosis by regulating expression levels of Bcl-2 and Bax genes in
the testes of CP-treated mice. The p38 mitogen-activated protein
kinase (MAPK) signal transduction pathway relies on p38, which is
an important member of the MAPK familythat can be activated by
anti-tumor drugs or ROS; p38 has been used to inhibit tumors by
inducing tumor cells apoptosis (32). The present results suggested that
CP-activated p38 protein was dephosphorylated by SIP, or that SIP
suppressed the phosphorylation of p38 protein induced by CP, which
indicated that SIP may impair p38 activation by scavenging
CP-mediated ROS to depress apoptosis. Akt is an important target
kinase in the downstream actions of phosphatidylinositol-2-kinase,
its phosphorylation can activate or inhibit downstream target
proteins such as Bad, Caspase-9, nuclear factor kappa B and
prostate apoptosis response-4 to block apoptosis (33–36). Akt
signaling is considered to be an important mechanism that
contributes toward sapoptosis inhibition (37,38). In
the present study, it was identified that CP upregulated the
content of phospho-Akt and the level of Akt phosphorylation was not
significantly reduced after inhibition of apoptosisby SIP. We
speculate that SIP may improve the cellular environmental impact of
CP and contribute to Akt-mediated inhibition of apoptosis.
In conclusion, SIP weakened CP-induced oxidative
stress to improve the antioxidant ability, protected functions of
mitochondria, increased expression levels of Bcl-2 and reduced
expression levels of Caspase-3 and Bax. Furthermore, SIP inhibited
the p38 MAPK signal pathway, thus suppressing CP-induced apoptosis
in testicular germ cells.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 31171667), the Special Fund
for Distinguished Experts in Guangxi and the Guangxi Talent
Highland of Preservation and Deep Processing Research in Fruit and
Vegetables.
References
|
1
|
Le XY, Luo P, Gu YP, Tao YX and Liu HZ:
Interventional effects of squid ink polysaccharides on
cyclophosphamide-associated testicular damage in mice. Bratisl Lek
Listy. 116:334–339. 2015.PubMed/NCBI
|
|
2
|
Mythili Y, Sudharsan PT, Selvakumar E and
Varalakshmi P: Protective effect of DL-alpha-lipoic acid on
cyclophosphamide induced oxidative cardiac injury. Chem Biol
Interact. 151:13–19. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aguilar-Mahecha A, Hales B and Robaire B:
Effects of acute and chronic cyclophosphamide treatment on meiotic
progression and the induction of DNA double-strand breaks in rat
spermatocytes. Biol Reprod. 72:1297–1304. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang J, Ma K and Wang H: Cyclophosphamide
suppresses thioredoxin reductase in bladder tissue and its adaptive
response via inductions of thioredoxin reductase and gluththione
peroxidase. Chem Biol Interact. 162:24–30. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Faleiro L and Lazebnik Y: Caspases disrupt
the nuclear-cytoplasmic barrier. J Cell Biol. 151:951–959. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu Z, Wu L, Li L, Tashiro S, Onodera S and
Ikejima T: p53-mediated cell cycle arrest and apoptosis induced by
shikonin via a caspase-9-dependent mechanism in human malignant
melanoma A375-S2 cells. J Pharmacol Sci. 94:166–176. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maiuri MC, Tasdemir E, Criollo A, Morselli
E, Vicencio JM, Carnuccio R and Kroemer G: Control of autophagy by
oncogenes and tumor suppressor genes. Cell Death Differ. 16:87–93.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Simon HU, Haj-Yehia A and Levi-Schaffer F:
Role of reactive oxygen species (ROS) in apoptosis induction.
Apoptosis. 5:415–418. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Zhang X, Rabbani ZN, Jackson IL
and Vujaskovic Z: Oxidative stress mediates radiation lung injury
by inducing apoptosis. Int J Radiat Oncol Biol Phys. 83:740–748.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cai L, Hales BF and Robaire B: Induction
of apoptosis in the germ cells of adult male rats after exposure to
cyclophosphamide. Biol Reprod. 56:1490–1497. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zong A, Zhao T, Zhang Y, Song X, Shi Y,
Cao H, Liu C, Cheng Y, Qu X, Cao J and Wang F: Anti-metastatic and
anti-angiogenic activities of sulfated polysaccharide of Sepiella
maindroni ink. Carbohyd Polym. 91:403–409. 2013. View Article : Google Scholar
|
|
12
|
Zuo T, He X, Cao L, Xue C and Tang QJ: The
dietary polysaccharide from Ommastrephes bartrami prevents
chemotherapeutic mucositis by promoting the gene expression of
antimicrobial peptides in Paneth cells. J Funct Foods. 12:530–539.
2015. View Article : Google Scholar
|
|
13
|
Zuo T, Cao L, Xue C and Tang QJ: Dietary
squid ink polysaccharide induced goblet cells to protect small
intestine from chemotherapy induced injury. Food Funct. 6:981–986.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tang Q, Zuo T, Lu S, Wu J, Wang J, Zheng
R, Chen S and Xue C: Dietary squid ink polysaccharides ameliorated
the intestinal microflora dysfunction in mice undergoing
chemotherapy. Food Funct. 5:2529–2535. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zuo T, Cao L, Li X, Zhang Q, Xue C and
Tang QJ: The squid ink polysaccharides protect tight junctions and
adherens junctions from chemotherapeutic injury in the small
intestinal epithelium of mice. Nutr Cancer. 67:364–371. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zuo T, Cao L, Sun X, Li X, Wu J, Lu S, Xue
C and Tang Q: Dietary squid ink polysaccharide could enhance SIgA
secretion in chemotherapeutic mice. Food Funct. 5:3189–3196. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen S, Xu J, Xue C, Dong P, Sheng W, Yu G
and Chai W: Sequence determination of a non-sulfated
glycosaminoglycan-like polysaccharide from melanin-free ink of the
squid Ommastrephes bartrami by negative-ion electrospray tandem
mass spectrometry and NMR spectroscopy. Glycoconj J. 25:481–492.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu HZ, Wang G, Wu JL, Shi LS, Zhong JP
and Pan JQ: Amelioratory effects of squid ink polysaccharides on
partial internal organs injuried by cyclophosphamide. Chinese J Mod
Appl Pharm. 2:89–93. 2012.
|
|
19
|
Staub AM: Removal of protein-Sevag method.
Methods Carbohydrate Chem. 5:5–6. 1965.
|
|
20
|
Le X, Luo P, Gu Y, Tao Y and Liu H: Squid
ink polysaccharides reduces cyclophosphamide-induced testicular
damage via Nrf2/ARE activation pathway in mice. Iran J Basic Med
Sci. 18:827–831. 2015.PubMed/NCBI
|
|
21
|
Elangovan N, Chiou TJ, Tzeng WF and Chu
ST: Cyclophosphamide treatment causes impairment of sperm and its
fertilizing ability in mice. Toxicology. 222:60–70. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brodsky RA: High-dose cyclophosphamide for
autoimmunity and alloimmunity. Immunol Res. 47:179–184. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Motoyoshi Y, Kaminoda K, Saitoh O,
Hamasaki K, Nakao K, Ishii N, Nagayama Y and Eguchi K: Different
mechanisms for anti-tumor effects of low- and high-dose
cyclophosphamide. Oncol Rep. 16:141–146. 2006.PubMed/NCBI
|
|
24
|
Guittin P, Labbe V, Cavalier JC, Delongeas
JL and Hodge T: Effects of cyclophosphamide administration on
different fertility evaluation tests in male rats. Teratology.
53:30A1996.
|
|
25
|
Luboshitzky R, Shen-Orr Z and Herer P:
Seminal plasma melatonin and gonadal steroids concentrations in
normal men. Arch Androl. 48:225–232. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martin CW, Riley SC, Everington D, Groome
NP, Riemersma RA, Baird DT and Anderson RA: Dose-finding study of
oral desogestrel with testosterone pellets for suppression of the
pituitary-testicular axis in normal men. Hum Reprod. 15:1515–1524.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iuchi Y, Kaneko T, Matsuki S, Ishii T,
Ikeda Y, Uchida K and Fujii J: Carbonyl stress and detoxification
ability in the male genital tract and testis of rats. Histochem
Cell Biol. 121:123–130. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Selvakumar E, Prahalathan C, Sudharsan PT
and Varalakshmi P: Protective effect of lipoic acid on
cyclophosphamide-induced testicular toxicity. Clin Chim Acta.
367:114–119. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Motawi TM, Sadik NA and Refaat A:
Cytoprotective effects of DL-alpha-lipoic acid or squalene on
cyclophosphamide-induced oxidative injury: An experimental study on
rat myocardium, testicles and urinary bladder. Food Chem Toxicol.
48:2326–2336. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tinari A, Garofalo T, Sorice M, Esposti MD
and Malorni W: Mitoptosis: Different pathways for mitochondrial
execution. Autophagy. 3:282–284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lindsay J, Esposti MD and Gilmore AP:
Bcl-2 proteins and mitochondria-specificity in membrane targeting
for death. Biochim Biophys Acta. 1813:532–539. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie N, Li H, Wei D, LeSaga G, Chen L, Wang
S, Zhang Y, Chi L, Ferslew K, He L, et al: Glycogen synthase
kinase-3 and p38 MAPK are required for opioid-induced microglia
apoptosis. Neuropharmacology. 59:444–451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cardone MH, Roy N, Stennicke HR, Salvesen
GS, Franke TF, Stanbridge E, Frisch S and Reed JC: Regulation of
cell death protease caspase-9 by phosphorylation. Science.
282:1318–1321. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Niquet J and Wasterlain CG: Bim, Bad, and
Bax: A deadly combination in epileptic seizures. J Clin Invest.
113:960–962. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jeong SJ, Pise-Masison CA, Radonovich MF,
Park HU and Brady JN: Activated AKT regulates NF-kappaB activation,
p53 inhibition and cell survival in HTLV-1-transformed cells.
Oncogene. 24:6719–6728. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Goswami A, Burikhanov R, de Thonel A,
Fujita N, Goswami M, Zhao Y, Eriksson JE, Tsuruo T and Rangnekar
VM: Binding and phosphorylation of par-4 by akt is essential for
cancer cell survival. Mol Cell. 20:33–44. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dan HC, Sun M, Kaneko S, Feldman RI,
Nicosia SV, Wang HG, Tsang BK and Cheng JQ: Akt phosphorylation and
stabilization of X-linked inhibitor of apoptosis protein (XIAP). J
Biol Chem. 291:228462016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang F, Lee JT, Navolanic PN, Steelman
LS, Shelton JG, Blalock WL, Franklin RA and McCubrey JA:
Involvement of PI3K/Akt pathway in cell cycle progression,
apoptosis, and neoplastic transformation: A target for cancer
chemotherapy. Leukemia. 17:590–603. 2003. View Article : Google Scholar : PubMed/NCBI
|