Introduction
Bronchopulmonary dysplasia (BPD) is a common chronic
lung disease in neonates, and an important cause of neonatal death.
BPD is closely related to infection, premature delivery and
long-term inhalation of high-concentration oxygen, characterized by
alveolar developmental block and impaired pulmonary microvascular
development (1). This disease has a
high mortality rate, and survivors are often suffering from lung
dysfunction and lung developmental disorder, even delayed
neurodevelopment and cerebral palsy (2). In recent years, with the continuous
development of medical technology, the incidence rate of typical
BPD has been gradually reduced, but that of new BPD remained
unchanged (3). Previous studies have
confirmed that pulmonary angiogenesis is the prerequisite for
normal alveolar development. This process involves mutual
regulation among multiple factors, among which the transcription
factor CASZI is an important factor in pulmonary angiogenesis
(4,5). Only few studies have been conducted on
the role of CASZI in bronchopulmonary development, and in this
study we analyzed the relationship between CASZI and pulmonary
angiogenesis using neonatal rats exposed to hyperoxia.
Materials and methods
Experimental animals and groups
Forty Sprague-Dawley neonatal rats aged 1–3 days of
either sex were purchased from Shanghai Experimental Animal Center.
Rats were randomly divided into the control group (n=20) and the
experimental group (n=20) using a random number table.
Materials
We used total RNA extraction kit; RT-PCR kit (both
from Active Motif, Carlsbad, CA, USA); rabbit-anti mouse β-actin
monoclonal antibody (dilution, 1:1,000; cat. no. CW0096; Beijing
ComWin Biotech Co., Ltd., Beijing, China); whole protein extraction
kit (Beyotime Biotechnology Institute); rabbit-anti mouse CASZ1,
Rho kinase, RhoA and ET-A receptor monoclonal antibodies (dilution,
1:1,000; cat. nos. sc-398303, sc-374388, sc-365241 and sc-376815;
Santa Cruz Biotechnology, Inc., CA, USA); infrared-labeled
fluorescent goat anti-rabbit secondary polyclonal antibody
(dilution, 1:2,000; cat. no. sc-2007; Santa Cruz Biotechnology,
Inc.); western blotting kit (Shanghai Kangcheng Biotech Co., Ltd.,
Shanghai, China). All reagents were analytically pure and made in
China.
The main instruments
Super clean bench (Suzhou Changqiao Purification
Equipment Factory, Suzhou, China); Synergy 2 multifunctional
microplate reader (BioTek Instruments, Winooski, VT, USA);
ultraviolet spectrophotometer (Third Shanghai Analytical Instrument
Factory, Shanghai, China); electronic scales (Sanyo, Tokyo, Japan);
electrophoresis apparatus (Liuyi, Beijing, China); RT-PCR
instrument (Shanghai Saimo Biological Science and Technology
Development Co., Ltd., Shanghai, China).
Establishment of animal model
Three different hole areas, inlet hole,
oxygen-measuring hole and outlet hole, were set up in a self-made
oxygen box. The neonatal rats in the experimental group were fed in
the self-made oxygen box (feeding conditions: oxygen concentration
≥95%, humidity, 50–70% and temperature, 22–25°C). Water and fodder
were added regularly every day and the padding was replaced. Rats
in the control group were placed in the standard mouse cage and fed
by indoor air. The breathing, growth, development and activity of
neonatal rats in both groups were monitored daily.
Detection indexes and methods
Specimen collection
At 14 days, rats were sacrificed using
intraperitoneal injection of 10% chloral hydrate and the right lung
was removed. Pieces of lung tissues were cryopreserved using liquid
nitrogen and preserved at −80°C. The remaining lung tissues were
fixed with 4% paraformaldehyde overnight for immunohistochemical
and hematoxylin and eosin (H&E) staining. The study was
approved by the Ethics Committee of Children's Hospital of
Zhengzhou and informed consents were signed by the patients and/or
guardians.
Pulmonary pathological examination
Lung tissues were fixed with 4% paraformaldehyde
overnight and were embedded in paraffin and cut into slices. After
H&E staining, radial alveolar count (RAC) and morphological
changes in lung tissues were observed under microscope. The
thickness of respiratory membrane was also detected using IPP6
image.
CASZI, Rho kinase, RhoA and ET-A receptor mRNAs
expression levels in lung tissues
Rat kidneys were removed and the total RNA was
extracted using TRIzol method. Target gene primer sequences used
were: GADPH (305 bp) forward, 5′-TGGTGAAGGTCGGTGTGAAC-3′ and
reverse primer, 5′-GGTGGTGAAGACGCCAGTAG-3′; CASZI (245 bp) forward,
5′-GGCACATTCCGGGACAGAGAGAA-3′ and reverse primer,
5′-CGGTTTACCCATGTGGTGCCTC-3′; RhoA forward,
5′-ACCAGTTCCCAGAGGTGTATG-3′ and reverse primer,
5′-TTGGGACAGAAGTGCTTGACTTC-3′; Rho kinase forward,
5′-CTGCGGGTACGAAGGTATCG-3′ and reverse primer,
5′-AGCATCCAATCCATCCAGCA-3′; ET-A receptor forward,
5′-CCGAGGAGCTCTAAGGGGAA-3′ and reverse primer,
5′-CCAAAAGGACGCCAGAAAGC-3′. The mRNA transcriptional levels of
CASZI, Rho kinase, RhoA and ET-A receptor were detected via RT-PCR,
and glyceraldehyde 3-phosphate dehydrogenase (GADPH) was used as
internal reference.
CASZI, Rho kinase, RhoA and ET-A receptor
proteins expression levels in lung tissues
Rat kidneys were removed and cut up using scissors.
The total protein was extracted from the kidney tissues using
pre-cooled tissue lysis buffer. The protein content in specimens
was determined using Bradford method. The protein was separated on
a 12% gel, and proteins were transferred onto the PVDF membrane
using the membrane transfer instrument under 100 V for 1.5 h.
Membrane was sealed using skim milk powder for 2 h. Membrane was
washed, and consequently diluted monoclonal antibodies (1:1,000)
anti-CASZI, Rho kinase, RhoA and ET-A receptor were added. After
overnight incubation, membrane was washed, and the diluted
secondary antibody was added followed by incubation at room
temperature for 60 min. BeyoECL Plus was used for color
development, color-developing bands were collected using
gel-imaging and chemiluminescence analysis system. Quantity One
software was used for data analysis of protein bands.
Statistical analysis. All data were analyzed and
treated using SPSS 18.0 software (SPSS, Inc., Chicago, IL, USA).
Measurement data were presented as mean ± SD and t-test was
performed. P<0.05 is considered to indicate a statistically
significant difference.
Results
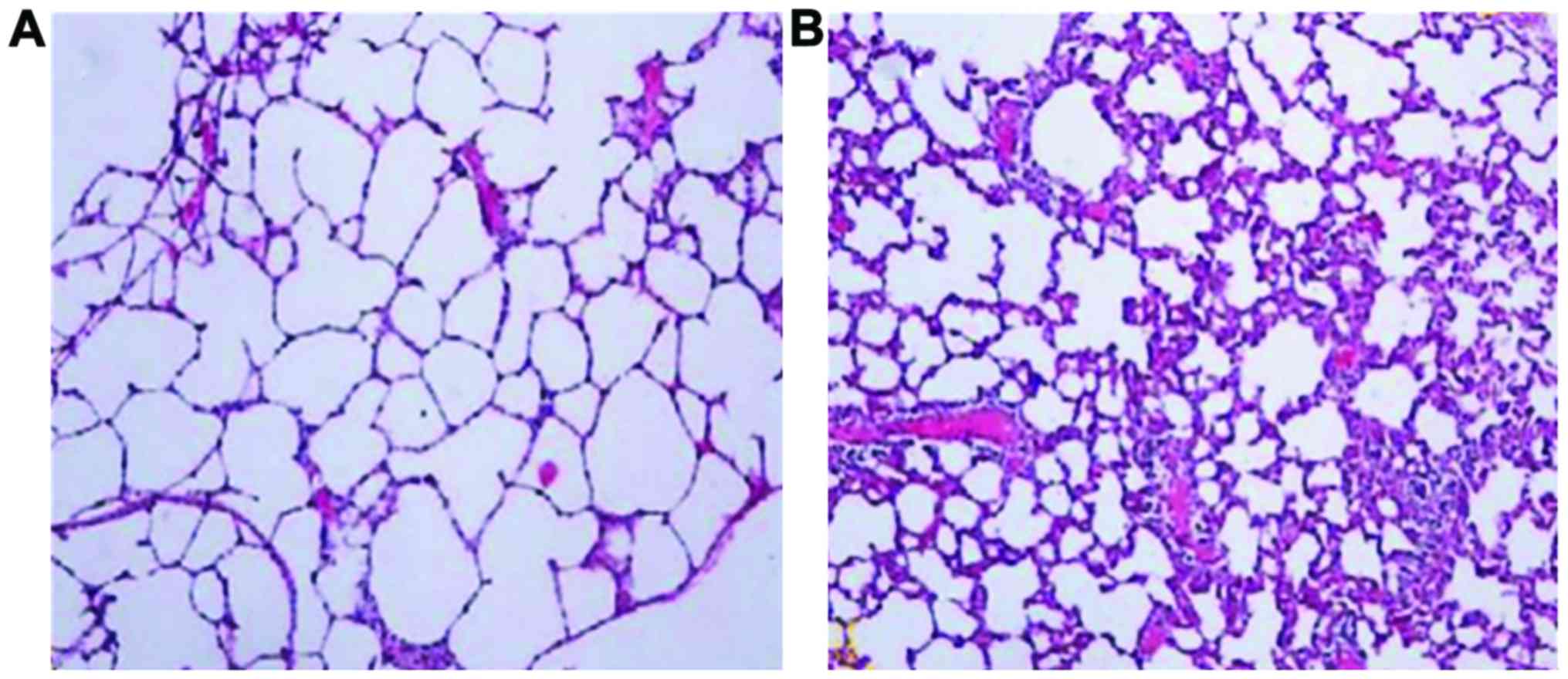
Morphological changes in lung
tissues
In neonatal rats in the control group, alveoli had a
uniform size and clear structure, the alveolar cavity was smaller
and the alveolar septum was thicker without any obvious
pathological changes. In neonatal rats in the experimental group,
alveoli had different sizes with disordered structures. The
alveolar wall was thinner, and we observed an increased alveolar
volume, reduced area and fusion, suggesting that BPD occurred in
lung tissue structures (Fig. 1).
RAC and thickness of respiratory
membrane
RAC in the experimental group was significantly
lower than that in the control group (P<0.05) and the thickness
of respiratory membrane was significantly higher in the
experimental group (P<0.05) (Table
I).
 | Table I.RAC and thickness of respiratory
membrane of neonatal rats in experimental group and control
group. |
Table I.
RAC and thickness of respiratory
membrane of neonatal rats in experimental group and control
group.
| Group | RAC (n) | Thickness of
respiratory membrane (µm) |
|---|
| Experimental
group | 5.6±0.5 | 11.9±2.3 |
| Control group | 8.2±0.9 | 6.9±1.5 |
| t-value | 2.308 | 2.275 |
| P-value | 0.015 | 0.022 |
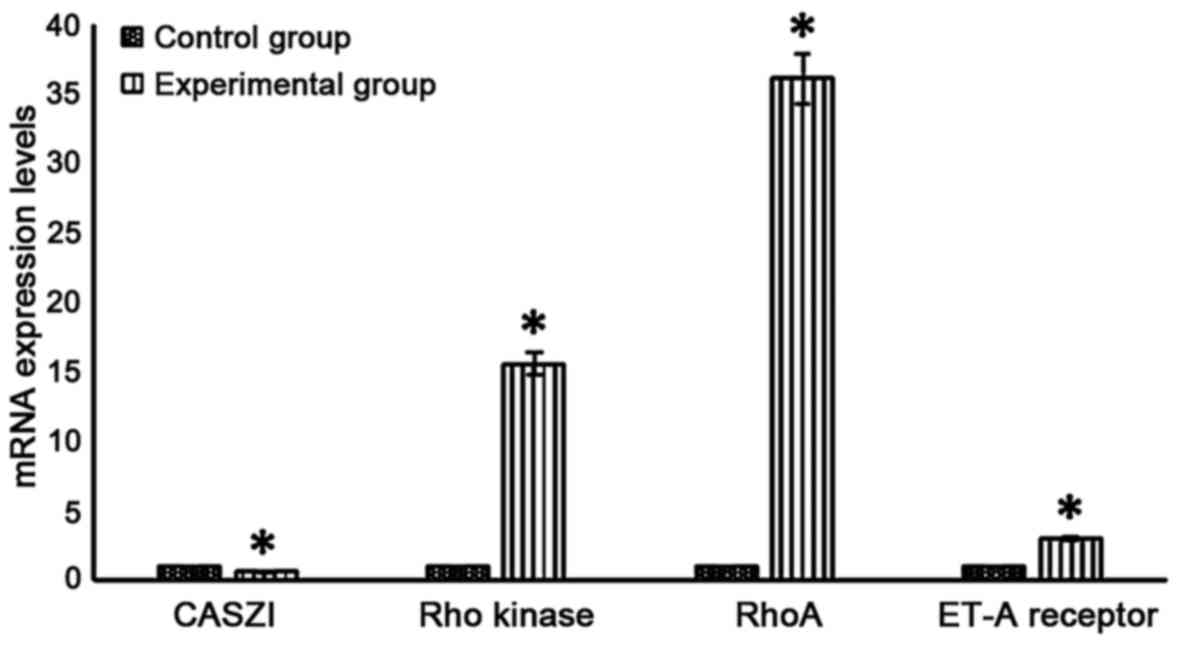
CASZI, Rho kinase, RhoA and ET-A
receptor mRNA expression levels in lung tissues
CASZI mRNA level in the experimental group was
significantly lower than that in the control group (P<0.05), and
Rho kinase, RhoA and ET-A receptor mRNA levels were significantly
higher than those in the control group (P<0.05) (Fig. 2).
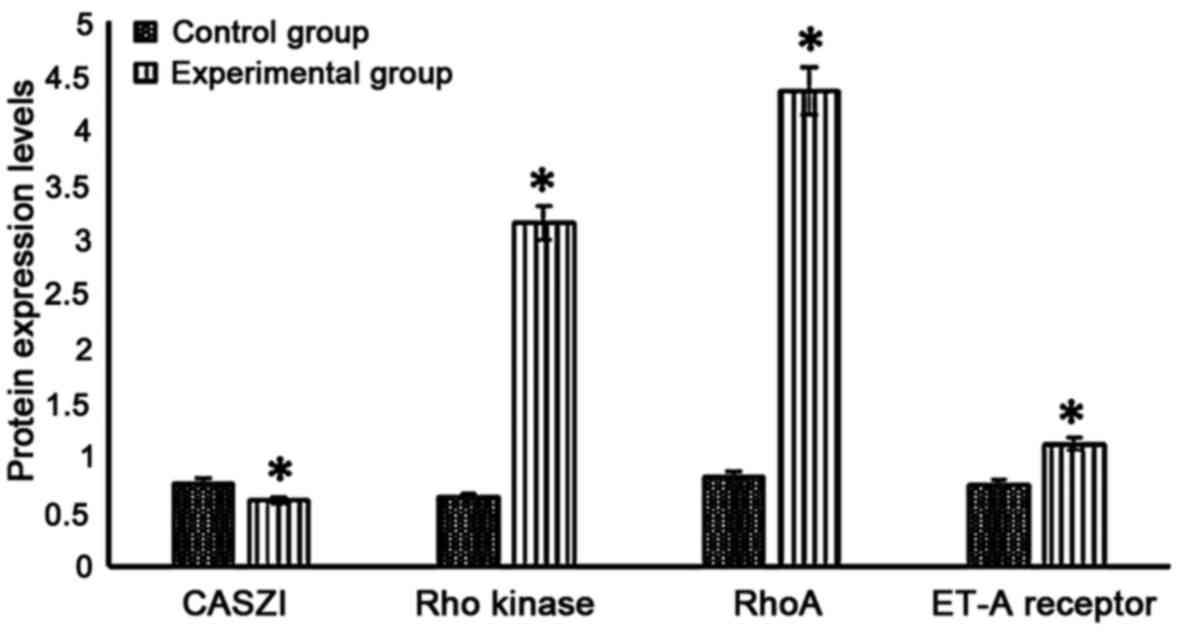
CASZI, Rho kinase, RhoA and ET-A
receptor protein expression levels in lung tissues
CASZI protein level in the experimental group was
significantly lower than that in the control group (P<0.05),
while Rho kinase, RhoA and ET-A receptor proteins levels were
significantly higher in the experimental group (P<0.05)
(Fig. 3).
Discussion
The complex pathogenesis of BPD is a multi-factor
process involving the interaction among epithelial cells, pulmonary
interstitium and pulmonary microvascular system (6). The alveolarization damage and pulmonary
microvascular abnormalities are the two important poles in the
pathogenesis of BPD (7). In recent
years, more attention has been focused on the importance of the
pulmonary microvascular abnormalities in BPD, and to emphasize this
importance, some scholars have introduced the ‘vascular hypothesis’
(8).
In this study, BPD rat model was established using
95% hyperoxia. The lung development in neonatal rats was in the
vesicular stage, equivalent to human gestational age of ~28 weeks.
At this time, the lung development was not perfect with active
shaping and remodeling. Hyperoxia method induced was similar to the
occurrence and development process of BPD in premature infants, and
with the ability of damaging and inhibiting the alveoli and can
cause acute lung injury in the short term. The experimental data
showed that in neonatal rats in the control group, alveoli had a
uniform size with clear structure. The alveolar cavity was smaller
and the alveolar septum was thicker without obviously pathological
changes. In the experimental group, alveoli had various sizes with
disordered structures. The alveolar wall was thinner, and an
increased alveolar volume was observed. In the experimental group,
we observed lower area and fusion, suggesting that BPD occurred in
lung tissue structures. RAC in the experimental group was
significantly lower than that in the control group (P<0.05) and
the thickness of respiratory membrane was significantly higher than
that in the control group (P<0.05). These observations showed
that BPD rat model was successfully established using 95%
hyperoxia.
Vasculogenesis and angiogenesis are two major
processes of vascular generation, the former is observed when the
new blood vessels are formed by endothelial cells, endothelial
progenitor cells or hemangioblastoma from nothing, and the latter
is observed when the new blood vessels are formed based on the
existing blood vessels through promoting the vascular endothelial
cell migration and proliferation (9,10).
Pulmonary angiogenesis involves a complex regulatory factors
network, and several pro-vascular growth factors have been already
discovered. These factors include angiopoietin (Ang), vascular
endothelial growth factor (VEGF), placental growth factor (PlGF),
tyrosine kinase receptor-ligand (EGF/Ephrin), platelet-derived
growth factor (PDGF), basic fibroblast growth factor (bFGF),
transforming growth factor-β (TGF-β), hepatocyte growth factor
(HGF) and monocyte chemoattractant protein-1 (11–13).
CASZI is a recently-discovered transcription factor which belongs
to the zinc finger gene family. CASZI is widely distributed in the
vascular endothelial cells in lung, stomach, pancreas, testis,
skeletal muscle, heart and small intestine. CASZI binds to DNA and
plays an important role in cell proliferation, differentiation and
apoptosis. It has been proved that CASZI can promote the formation
and branching of vascular networks, as well as promoting vascular
buds (14). The results of this
study showed that CASZI mRNA and protein expression levels in the
experimental group were significantly lower than those in the
control group (P<0.05). These results suggested that
high-concentration of oxygen could inhibit the expression of CASZI,
and CASZI may play an important role in the development of BPD.
In order to further study the regulatory mechanism
of lung development, Rho kinase, RhoA and ET-A receptor mRNA and
protein expression levels in RhoA/ROCK were analyzed. RhoA/ROCK is
a downstream signaling pathway of CASZI, and RhoA is an important
cellular signal transduction molecule, which is essentially a
GTPase (15,16), and its main role is to hydrolyze
ornithine. Activated RhoA has the capability of influencing a
variety of biological behaviors, such as cell cycle regulation,
transcription factor activation and smooth muscle contraction
(16). Rho kinase is a downstream
signaling protein of RhoA, and G-protein-coupled receptor can
activate RhoA through ET receptor, thus activating Rho kinase and
mediating the differentiation, proliferation and migration of
vascular endothelial cells (17,18).
Prior studies have confirmed that CASZI can regulate the
endothelial cell adhesion through RhoA, and promote vascular buds.
The experimental data showed that Rho kinase, RhoA and ET-A
receptor protein expression levels in the experimental group were
significantly higher than those in the control group (P<0.05).
Therefore, it is speculated that hyperoxia may inhibit CASZI
expression, thus downregulating the expression of RhoA, activating
RhoA/ROCK pathway and reducing the formation and branching of
pulmonary vascular network.
We concluded that the expression of CASZI, during
the development of BPD, was downregulated, and this influenced the
pulmonary microvascular regeneration through RhoA/ROCK signaling
pathway, thus participated in the pathogenesis of BPD.
References
|
1
|
Gien J, Kinsella J, Thrasher J, Grenolds
A, Abman SH and Baker CD: Retrospective analysis of an
interdisciplinary ventilator care program intervention on survival
of infants with ventilator-dependent bronchopulmonary dysplasia. Am
J Perinatol. 34:155–163. 2017.PubMed/NCBI
|
|
2
|
Nair V, Loganathan P and Soraisham AS:
Azithromycin andother macrolides for prevention of
bronchopulmonarydysplasia: A systematic review and meta-analysis.
Neonatology. 106:337–347. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gadhia MM, Cutter GR, Abman SH and
Kinsella JP: Effects of early inhaled nitric oxide therapy and
vitamin A supplementation on the risk for bronchopulmonary
dysplasia in premature newborns with respiratory failure. J
Pediatr. 164:744–748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu Q, Liu X and Zhou G: Effects of
ligustrazine on hypoxia-inducible factor pathway expression in
neonatal rats with BPD. J Pediatr Pharm. 20:1–5. 2014.
|
|
5
|
Golovnin AK, Kostyuchenko MV, Georgiev PG
and Melnikova LS: Mod(mdg4)-58.8, isoform of mod(mdg4) loci,
directly interacts with MTACP1A and MTACP1B proteins of
Drosophila melanogaster. Dokl Biochem Biophys. 466:5–8.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vom Hove M, Prenzel F, Uhlig HH and
Robel-Tillig E: Pulmonary outcome in former preterm, very low birth
weight children with bronchopulmonary dysplasia: A case-control
follow-up at school age. J Pediatr. 164:40–45.e4. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Balany J and Bhandari V: Understanding the
impact of infection, inflammation, and their persistence in the
pathogenesis of bronchopulmonary dysplasia. Front Med (Lausanne).
2:902015.PubMed/NCBI
|
|
8
|
Del Re DP, Miyamoto S and Brown JH:
RhoA/Rho kinase upregulate Bax to activate a mitochondrial death
pathway and induce cardiomyocyte apoptosis. J Biol Chem.
282:8069–8078. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Monteiro CB, Midão L, Rebelo S, Reguenga
C, Lima D and Monteiro FA: Zinc finger transcription factor Casz1
expression is regulated by homeodomain transcription factor Prrxl1
in embryonic spinal dorsal horn late-born excitatory interneurons.
Eur J Neurosci. 43:1449–1459. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang L, Feng Z and Lv H: Expressions of
vascular endothelial growth factor and angiopoietin-1 in neonatal
rats with hypoxia-induced BPD and effects on lung development. J
Pract Med. 20:525–528. 2014.
|
|
11
|
Tang H: ‘Vascular hypothesi’ of BPD and
progress of Angiogenesis-promoting Treatment. J Clin Pediatr.
34:5532016.
|
|
12
|
Liu Z, Lam N and Thiele CJ: Zinc finger
transcription factor CASZ1 interacts with histones, DNA repair
proteins and recruits NuRD complex to regulate gene transcription.
Oncotarget. 6:27628–27640. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Q, Ma X and Zhang X: Pulmonary
microvascular development and CD34 expression in rats with
oxygen-induced BPD. J PLA Med. 26:25–28. 2014.
|
|
14
|
Sauzeau V, Rolli-Derkinderen M, Lehoux S,
Loirand G and Pacaud P: Sildenafil prevents change in RhoA
expression induced by chronic hypoxia in rat pulmonary artery. Circ
Res. 93:630–637. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pierro M, Ionescu L, Montemurro T, Vadivel
A, Weissmann G, Oudit G, Emery D, Bodiga S, Eaton F, Péault B, et
al: Short-term, long-term and paracrine effect of human umbilical
cord-derived stem cells in lung injury prevention and repair in
experimental bronchopulmonary dysplasia. Thorax. 68:475–484. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Julian L and Olson MF: Rho-associated
coiled-coil containing kinases (ROCK): Structure, regulation, and
functions. Small GTPases. 5:e298462014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stiles JM, Kurisetty V, Mitchell DC and
Bryan BA: Rho kinase proteins regulate global miRNA expression in
endothelial cells. Cancer Genomics Proteomics. 10:251–263.
2013.PubMed/NCBI
|
|
18
|
Kaya G, Sivasli E, Oztuzcu S, Melekoglu
NA, Ozkara E, Sarikabadayi U and Demiryürek AT: Association of
Rho-kinase gene polymorphisms with respiratory distress syndrome in
preterm neonates. Pediatr Neonatol. 58:36–42. 2017. View Article : Google Scholar : PubMed/NCBI
|