Introduction
Esophageal cancer is an important public health
problem worldwide, as the eighth most common cancer and sixth most
common cause of cancer-associated mortality (1). Esophageal squamous cell carcinoma
(ESCC) accounts for 90% of all histological types of esophageal
carcinoma in the highest risk area, North-central China, which
ranks first in ESCC morbidity and mortality rates (2). Genetic heterogeneity and complex
regulatory networks make it difficult to treat ESCC (3). Thus, identifying the regulatory
pathways for the stratification of ESCC is of great clinical
significance.
MicroRNAs (miRNAs/miRs) are single-stranded RNAs of
18–24 nucleotides in length that post-transcriptionally regulate
gene expression by directly binding to the 3′-untranslated region
(UTR) of target mRNAs (4). Emerging
evidence has revealed that numerous miRs are involved in
tumorigenesis and/or tumor progression (5). The involvement of miR regulatory
mechanisms in cancer progression may be essential to understand the
complete molecular landscape of carcinoma progression. Although the
first miRNA, lin-4, was discovered in 1,993 (6) and the second, let-7, in 2,000 (7) in Caenorhabditis elegans, only
recently has the study of this class of small regulatory RNAs in
humans become more widespread. Experimental evidence that has
accumulated has led researchers to notice that unknown molecular
factors, particularly non-coding RNAs, serve important roles in
carcinogenesis (8). Approximately
70% of the genome has been identified to be transcribed in mammals
(9). Over the past decade, miRNAs
have emerged as important players in RNA interference-mediated
post-transcriptional gene regulation.
miRNAs can be transcribed from individual genes or
as clusters (10), with ~30% of
miRNAs transcribed as polycistronic clusters (11). miRNA clusters are comprised of
several miRNA genes, which are located adjacent to one another on
the chromosome and are typically highly related structural genes
with similar functions (12). The
miRNA genes in these clusters are transcribed as one long pri-miRNA
transcript, which is subsequently processed into the individual
miRNAs (12). The role of miRNA
clusters in tumorigenesis has been extensively studied; Liu et
al (13) identified that the
cluster of miR-143 and miR-145 served a role in the modulation of
ESCC metastases through targeting oncogenic fascin actin-bundling
protein 1, thus affecting the risk of ESCC progression. The
miR-106b-25 cluster is located in the thirteenth intron of the DNA
replication gene minichromosome maintenance complex component 7
(MCM7), which resides on chromosome 7 in humans and encodes three
miRNAs: miR-106b, miR-93 and miR-25. A previous study revealed that
members of the miR-106b-25 cluster are overexpressed in several
types of cancer (14).
miR-25, a member of the miR-106b-25 cluster, has
been identified to be overexpressed in a variety of types of cancer
and serve functional roles in numerous malignancy-associated
processes, including tumorigenesis, and cancer cell proliferation,
migration and metastasis (15).
Furthermore, diverse molecular signaling pathways appear to be the
targets of miR-25 in various types of cancer. For example, in
breast cancer miR-25 activates the transforming growth factor
(TGF)-β signaling pathway to induce tumorigenesis (15). However, the molecular mechanisms
underlying ESCC remain unclear; thus, further studies are required
to ascertain a more thorough understanding of the molecular basis
of ESCC.
The present study aimed to investigate the effects
of miR-25 on the metastasis and invasion of KYSE-150 and EC109 ESCC
cells. Furthermore, members of the miR-106b-25 cluster were
bioinformatically analyzed using a variety of methods, including
Gene Ontology (GO) term and Kyoto Encyclopedia of Genes and Genomes
(KEGG) pathway enrichment analyses. Downstream proteins were
analyzed and potential target genes of miR-106b, miR-93 and miR-25
were identified, laying a foundation for further research.
Materials and methods
Cell culture
The human ESCC cell lines KYSE150 and EC109 were
purchased from the Type Culture Collection of the Chinese Academy
of Sciences (Shanghai, China). KYSE150 is highly differentiated,
while EC109 is less differentiated. All cells were cultured in
RPMI-1640 (Gibco) supplemented with 10% fetal bovine serum (FBS;
Invitrogen) (both Thermo Fisher Scientific, Inc., Waltham, MA, USA)
at 37°C with 5% CO2.
Transfection
KYSE150 and EC109 were seeded into 6-well plates
(2×105 cells per well) and cultured for 5–10 min prior
to transfection. HiPerFect Transfection Reagent (Qiagen GmbH,
Hilden, Germany) was used for miRNA and siRNA transfection
according to the manufacturer's protocol. miScript miR-25 mimic
(MSY0000081), miScript miR-25 inhibitor (MIN0000081) and Allstars
negative control siRNA (cat.no. 1027280; all Qiagen GmbH) were
transfected into the cells. Transfected cells were then subjected
to reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analysis, and Transwell invasion and migration
assays.
RT-qPCR analysis
Total RNA from KYSE150 and EC109 cells was isolated
48 h following transfection using the miRNeasy Mini kit, cDNA was
reverse transcribed using the miScript II Reverse Transcription kit
and qPCR was performed using the miScript SYBR Green PCR kit (all
Qiagen GmbH), according to the manufacturer's protocol. The
thermocycling conditions were presented in Table I. The threshold cycle (Cq) was
defined as the fractional cycle number at which the fluorescence
passed the fixed threshold. The PCR primers were as follows:
miR-25, forward, 5′-CGAGCTCTTTTAAGACAGG-3′ and reverse,
5′-GCCGGCTGCATGCCTGTGG-3′; and U6, forward,
5′-GTGTGCTACGGAGTTCAGAGGTT-3′ and reverse,
5′-TGGGGTTATACATTGTGAGAGGA-3′.miRNA quantitation was performed
using a TaqMan miRNA assay (Applied Biosystems; Thermo Fisher
Scientific, Inc.) in a CFX96 real-time thermal cycler (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The amount of target gene
expression was normalized using U6 as the reference gene and the
amount of target gene in the control cells was set at 1.0.
Following the initial activaton step for 15 min at 95°C, the
reaction system entered into the 3 step cycling which included the
following steps for 40 cycles: Denaturation for 15 sec at 94°C;
annealing for 30 sec at 55°C and extension for 30 sec at 70°C. The
relative expression of miR-25 was then calculated using the
2−ΔΔCq method (16). All
samples were processed in triplicate.
 | Table I.Predicted target Genes of the
miR-106b-25 cluster. |
Table I.
Predicted target Genes of the
miR-106b-25 cluster.
| miRNA | Target genes |
|---|
| miR-106b | KAT2B, DNAJB9, E2F1,
AGFG2, APC, SMAD5, S1PR1, PLEKHM1, C5orf41, ITCH, APP, CDKN1A, RB1,
TCEAL1 |
| miR-93 | KAT2B, ENPP5, TP53,
EPHA4, ZNF512B, TRIM3, FGD5, OSR1, RAB5B, GIGYF1, PGBD5, NRP2,
NKX3-2, FGD1, MAP3K3, PAFAH1B1, NHLH1, SKI, TNRC6A, INP1, CDKN1A,
E2F1, ITGB8, TUSC2, PTEN, LATS2 |
| miR-25 | PTEN, ZNF512B, ERC2,
KAT2B, MIER3, DMXL1, TP53, EXOC5, PRKCE, PCDH11Y, PDS5B, E2F3,
LHFPL2, COL1A2, ARHGEF17, FOXG1, MAP2K4, TMEM87A, PRDM13, ZNF287,
TRAK2, ATRX, |
|
| DNAJB12, KCNK10,
C17orf39, GDF11, XYLT2, LATS2, DSCAML1, TOB1, EGR2, ADM, IQGAP2,
ADCY3, FNBP4,NAA15, SOBP, RNF44, NLK, BAZ2A, BCL2L11, CDKN1C, WDR4,
CDH1, CCL26, MDM2, EZH2, SMAD7, PRMT5, KLF4, FBXW7, NOX4, WWP2,
TMEM188 |
Transwell invasion and migration
assays
A total of 48 h after transfection the KYSE-150 and
EC109 cells were used for Transwell invasion and migration assays.
For the invasion assay, the upper chamber of a 24-well Transwell
permeable support with 8 um pores (cat. no. 5422) coated with
Matrigel (1 mg/ml; Matrigel Basement Membrane Matrix) (both Corning
Incorporated, Corning, NY, USA) was loaded with 200 µl of cell
suspension (1–4×105 cells/well) and the lower chamber
was loaded with 600 µl of RPMI-1,640 containing 10% FBS. The
Transwell was then incubated at 37°C with 5% CO2 for 48
h. Subsequently, the cells remaining on the filter surface were
fixed with 4% formaldehyde for 15 min at room temperature, stained
with 0.5% crystal violet at room temperature for 30 min and
examined under a microscope (objective, ×40; magnification, ×100).
Cells in at least six random microscopic fields were counted. A
total of five wells were tested for each group and all experiments
were performed in triplicate. The migration assay was performed in
the same way, with the exclusion of the Matrigel.
miR-106b, miR-2 and miR-93 target gene
prediction, and GO term and KEGG pathway enrichment analyses
Three target gene prediction databases were utilized
to predict the target genes of the three miRs of miR-106b-25
cluster. These were TargetScan (version 6.1; targetscan.org), PicTar (pictar.mdc-berlin.de) and miRanda (microrna.org). The intersection of the predicted
targeted genes was taken and then combined with the validated genes
that miRTarBase 6.0 (http://mirtarbase.mbc.nctu.edu.tw/php/search.php)
provided in order to obtain the target genes. GO analysis is widely
used for the functional annotation of genomic data. To explore the
functions of miR-106b, miR-2 and miR-93, GO (geneontology.org) (17)
was employed to perform biological process term enrichment
analysis. In addition, KEGG pathway enrichment analysis was
performed. These enrichment analyses were performed using the
Database for Annotation, Visualization and Integrated Discovery
(DAVID; version 6.7) (17). KEGG can
provide a valid estimate of the target genes' P-value by a Monte
Carlo simulation procedure (18).
The enrichment analysis results with P<0.05 were identified as
significant results.
Protein-protein interaction (PPI)
network construction
The identified target genes of the miR-106b-25
cluster were placed into the Search Tool for the Retrieval of
Interacting Genes (STRING) database (string-db.org), a meta resource that collects
available information on protein-protein associations, scores and
weights them, and then augments them with predicted interactions
and with the results of automatic text-mining searches in order to
construct a PPI network (17).
Statistical analysis
Data are expressed as the mean ± standard error. The
differences among groups were determined by one-way analysis of
variance, which was followed by a post hoc Dunnett's T3 (3) test when multiple groups were being
compared using SPSS 20.0 (IBM Corp., Armonk, NY, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression of miR-25 in KYSE-150 and
EC109 cells after transfection
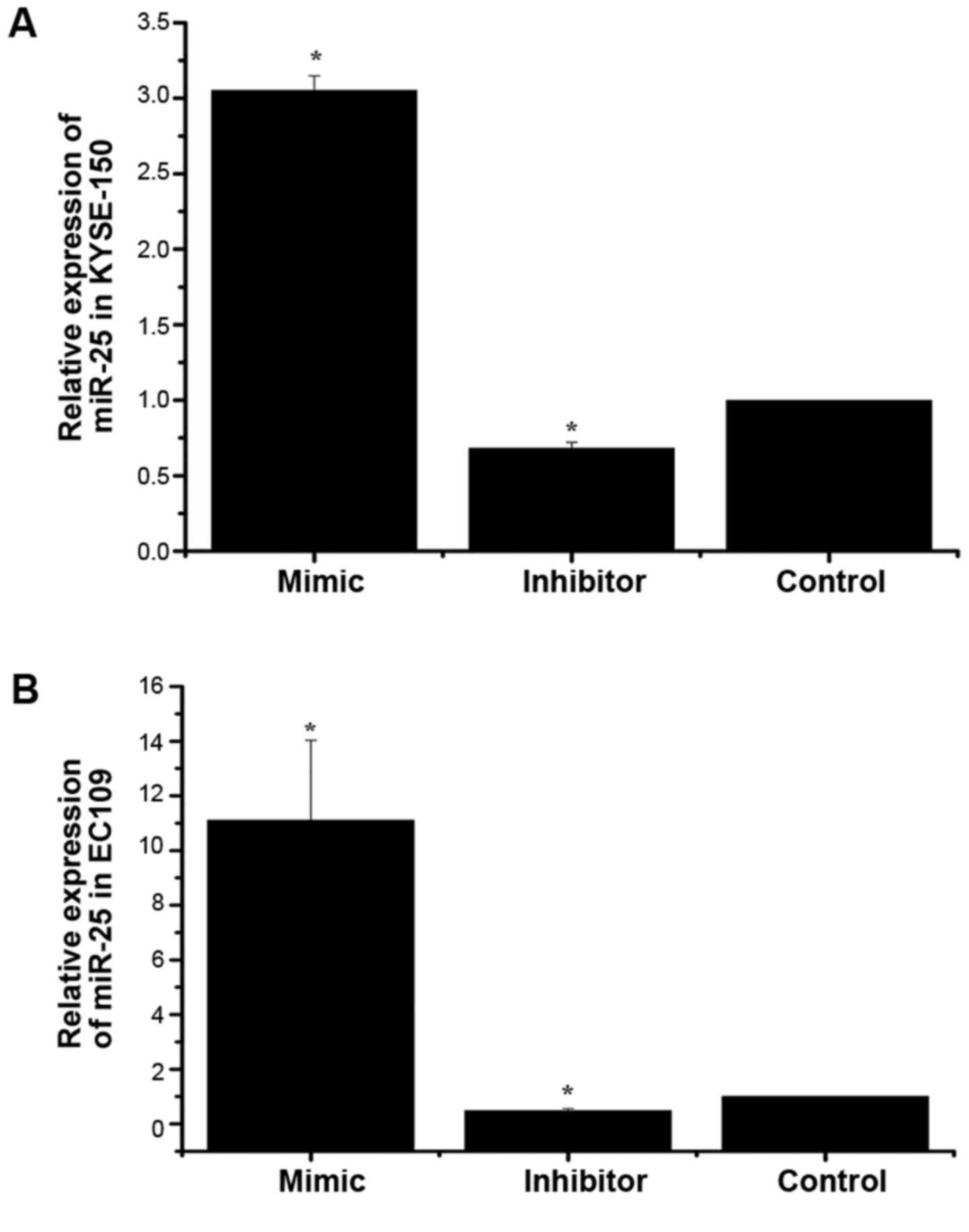
To investigate the impact of miR-25 on the growth of
KYSE-150 and EC109 cells, the expression of miR-25 was altered by
the transfection of a mimic or inhibitor. A negative control was
also transfected into a third group of cells. The expression of
miR-25 in the three groups of KYSE-150 and EC109 cells was then
detected by RT-qPCR analysis. U6 was used as an internal control.
As demonstrated in Fig. 1, the
expression of miR-25 was significantly increased in the mimic group
compared with the control group, and significantly decreased in the
inhibitor group. This indicated that cell models with differential
expression of miR-25 were successfully constructed.
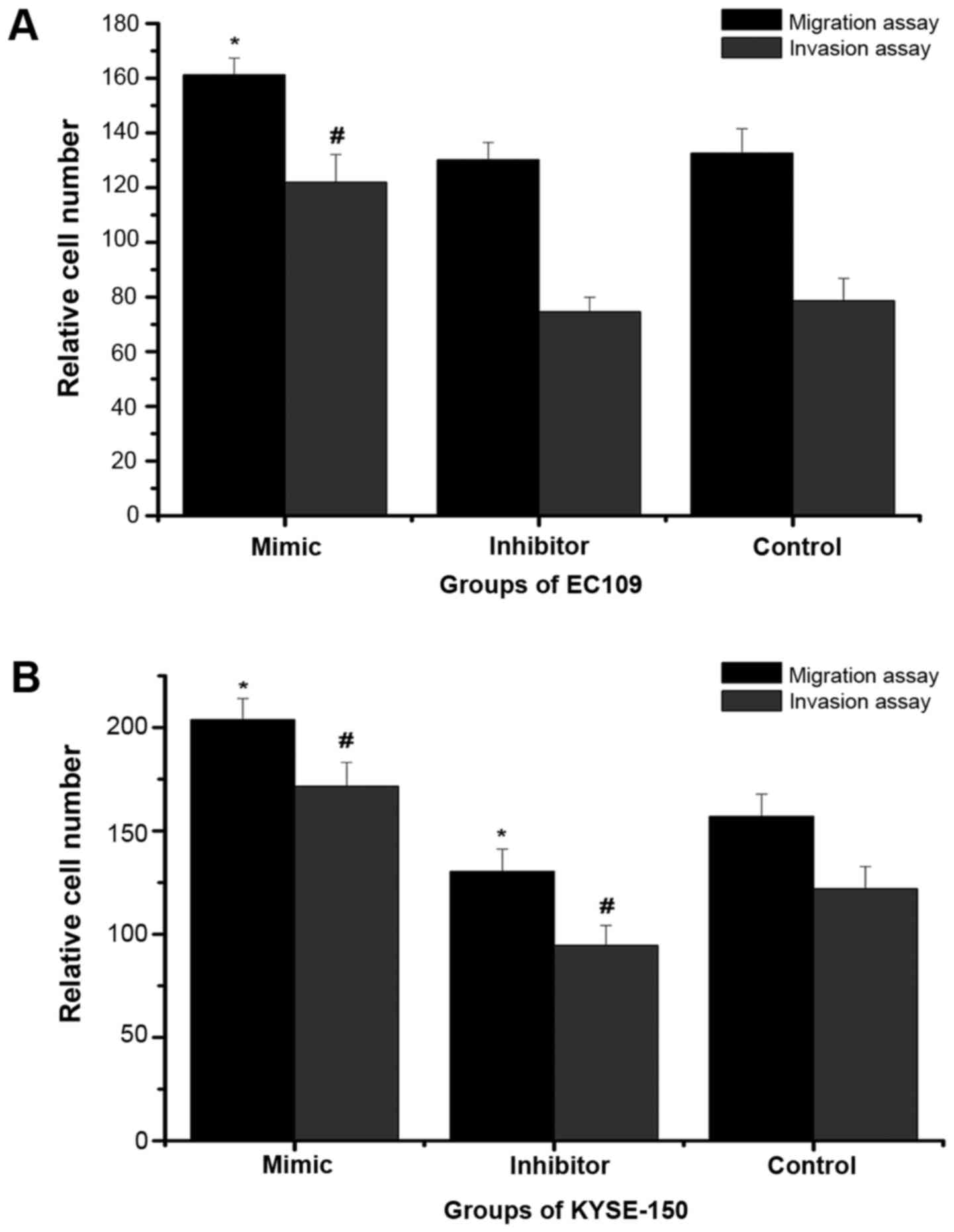
miR-25 affects the invasion and
metastasis of KYSE-150 and EC109 ESSC cells differently
In order to analyze metastasis and invasiveness,
which are important features of malignant cells, Transwell
migration and invasion assays were performed (Figs. 2 and 3). In the migration and invasion assays,
the number of KYSE-150 cells in the mimic group was significantly
higher than that in the negative control group, while the number of
cells in the inhibitor group was significantly decreased compared
with the negative control group. In the EC109 cells, similarly, the
number of cells in the mimic group was significantly higher than
that in the negative control group, however, there was no
significant difference between the numbers of cells in the
inhibitor and negative control groups. This indicated that high
expression of miR-25 may promote the invasion and metastasis of
ESCC cells, while low expression of miR-25 did not necessarily
inhibit invasion and metastasis.
Bioinformatical analysis of the
predicted target genes of miR-106b, miR-25 and miR-93
In order to investigate potential regulation
mechanisms of the miR-106b-25 cluster in the pathogenesis of ESCC,
four online bioinformatics databases (TargetScan, PicTar, miRanda
and miRTarbase) were used to identify and validate potential
targets of the three members of the cluster. A total of 14 target
genes of miR-106b, 25 target genes of miR-93 and 54 target genes of
miR-25 were identified (Table I).
Next, the biological processes that these 93 targets are involved
in were investigated using GO term and KEGG pathway enrichment
analyses (Tables II and III, respectively). The first three
pathways of the enrichment analysis indicated where the target
genes were concentrated and the results demonstrated that they
might be important genes in the regulation of macromolecule
metabolic processes and cell cycle and may serve a role in
Glioma.
| Table II.GO biological process term enrichment
analysis of the targets of the miR-106b-25 cluster. |
Table II.
GO biological process term enrichment
analysis of the targets of the miR-106b-25 cluster.
| miRNA | GO no. | Term | P-value |
|---|
| miR-106b | 0010604 | Positive regulation
of macromolecule metabolic process | <0.0001 |
|
| 0051329 | Interphase of mitotic
cell cycle |
0.000116 |
|
| 0007050 | Cell cycle
arrest |
0.000116 |
| miR-93 | 0030030 | Projection
organization |
0,000261 |
|
| 0032319 | Regulation of Rho
GTPase activity |
0.000930 |
|
| 0007049 | Cell cycle |
0.001180 |
| miR-25 | 0008285 | Negative regulation
of cell proliferation | <0.0001 |
|
| 0042127 | Regulation of cell
proliferation | <0.0001 |
|
| 0030030 | Cell projection
organization | <0.0001 |
| Table III.Kyoto Encyclopedia of Genes and
Genomes pathway enrichment analysis of the miR-106b-25 cluster. |
Table III.
Kyoto Encyclopedia of Genes and
Genomes pathway enrichment analysis of the miR-106b-25 cluster.
| miRNA | Term | P-value |
|---|
| miR-106b | Bladder cancer |
0.0018 |
|
| Glioma |
0.0040 |
|
| Melanoma |
0.0051 |
| miR-93 | Glioma |
0.0051 |
|
| Melanoma |
0.0065 |
|
| Prostate
cancer |
0.0100 |
| miR-25 | Melanoma | <0.0001 |
|
| Glioma |
0.0011 |
|
| Prostate
cancer |
0.0030 |
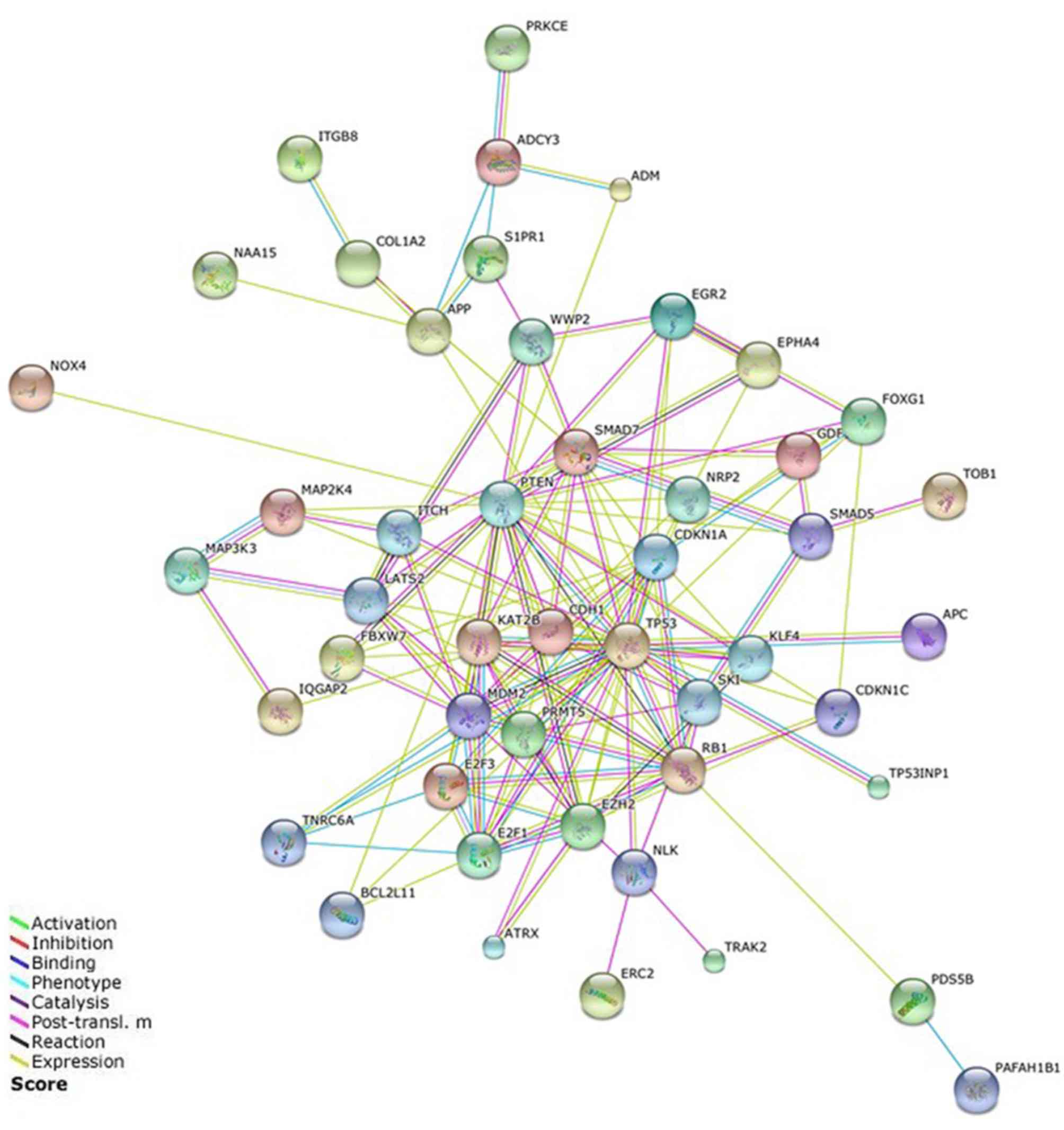
PPI network
A PPI network of the miR-106b-25 cluster target
genes was constructed using STRING (Fig.
4). A total of 45 target genes that were identified as being
linked to lax isolated nodes and not connected to any other nodes
through data analysis were removed. The PPI network created
revealed complex interactions, particularly for the target genes
PTEN, TP53, MDM2, E2F1, PRMT5, MCM2, RB1, CDKN1A, SMAD7 and EZH2.
So these genes were the focus of our study.
Discussion
Compared with normal cells, cancer cells exhibit
reduced cell adhesion, unlimited proliferation and metastasis.
Cancer cells may exhibit different degrees of differentiation. Cell
differentiation refers to the process undergone by cells of the
same origin, which produces cells with different morphological and
functional features. Selective expression of the genome in a time-
and location-dependent manner causes cell differentiation. In
vitro cultured cells grow under specific conditions, and retain
the same basic structure and function as cells in vivo. The
present study used two ESCC cells, which exhibited different
degrees of differentiation. KYSE150 is highly differentiated, while
EC109 is less differentiated. The cells were transfected with an
miR-25 mimic or inhibitor in order to investigate the role of
miR-25 in ESCC. The results indicated that overexpression of miR-25
promoted the invasion and metastasis of ESCC cells regardless of
the degree of cell differentiation. However, inhibiting miR-25 had
different effects on the invasion and metastasis abilities of ESCC
cells of different degrees of differentiation. The invasion and
metastasis of cells with a high degree of differentiation was
significantly decreased, while the abilities of poorly
differentiated cells did not change significantly. Thus, the impact
of different degrees of differentiation should be taken into
consideration in cancer studies. These results also suggest that
clinical treatment should be individualized according to the degree
of differentiation; when ESCC is well differentiated, inhibition of
miR-25 may be an effective treatment, while if it is not this
treatment would be ineffective.
Multiple steps in tumorigenesis, including
migration, cell adhesion and immunological escape, are involved in
invasion and metastasis, which serve a significant role in the
development of ESCC. Complicated processes such as these cannot be
regulated by a single molecule and require numerous molecules to
interact with one another. Research over the last decades has
identified a number of oncogenic and tumor suppressor proteins that
are associated with the tumorigenesis of ESCC (13,17,19,20),
however, molecular indicators of the origin of cellular
deregulation in ESCC have not yet been identified. In order to
improve the survival rates of patients with ESCC, it is necessary
to understand the molecular etiology of the disease, and identify
novel biomarkers for the early detection of pre-malignant lesions
and localized carcinoma (17).
miR clusters are closely related structural genes
with similar functions. Investigating how miRNA families are
expressed in clusters and control cell signaling pathways is likely
to increase understanding of cancer progression. Several studies
have identified that the polycistronic miR-106b-25 cluster is
tumorigenic (21,22). Li et al (21) reported that the miR-106b-25 cluster
may have an anti-apoptotic role and could promote cell cycling
in vitro. In the present study, bioinformatical analysis of
the miR-106b-25 cluster was performed, including target prediction,
followed by enrichment analysis these targets and PPI network
construction. A total of 93 target genes were predicted for the
three members of the cluster, including PTEN, TP53, MDM2, E2F1,
PRMT5, MCM2, RB1, CDKN1A, SHAD7 and EZH2. GO term enrichment
analysis revealed that the target genes of the cluster were
significantly enriched in cell metabolism and the cell cycle. A
previous study revealed that inhibiting the expression of the three
members of the miR-106b-25 cluster slowed the development of liver
cancer, and although the effects of the three members differed from
each other, the trend was the same, which suggests that members of
this cluster serve a role in tumor development (21). KEGG pathway enrichment analysis in
the present study revealed that the gene targets of miR-106b-25
cluster members were concentrated in signaling pathways associated
with glioma, melanoma and bladder cancer.
In the present study, a PPI network of the predicted
targets of the miR-106b-25 cluster was constructed. PTEN, MDM2 and
p53 were located centrally in this network. Research in gastric
cancer cells by Petrocca et al (23) demonstrated that the miR-106b-25
cluster was involved in E2F1 post-transcriptional regulation and
may serve a key role in the development of TGF-β resistance in
gastric cancer. E2F1 is an important transcription factor that
serves a role cancer progression (24,25)
MDM2, a negative regulator of p53, is an important regulator of the
cell cycle and apoptosis. MDM2 serves a pivotal role in p53
stabilization via phosphorylation at serine residues, and
overexpression of MDM2 has frequently been observed in solid tumors
and is associated with tumor progression (17). TP53 is at the core of the complicated
PPI network, since deregulated p53 expression is closely associated
with numerous types of cancer (17).
p53 acts as a tumor by triggering cell cycle arrest and apoptosis,
as well as maintaining genome stability (26). Xu et al (27) revealed that miR-25 promoted cell
migration and invasion in ESCC. In prostate cancer and ovarian
cancer, miR-93 can bind to and decrease PTEN expression, thereby
promoting cancer cell proliferation and invasion (28,29).
These results indicate that certain genes in the PPI network serve
important roles in tumorigenesis. However, many other genes in the
network have not been fully studied yet, including PRMT5, MDM2,
RB1, CDKN1A, SMAD7 and EZH2. These genes are located centrally in
the PPI network and have numerous links to other genes in the
network, so further experiments are required to verify their
functions.
The results of the present study and previous
research suggest that interactions between the three members of
miR-106b-25 cluster regulate multiple downstream target proteins to
promote or inhibit carcinogenesis through cell cycle regulation
mediated via several tumor-associated signaling pathways. Though
several of the target genes of the miR-106-25 cluster have been
studied, this research is still limited and knowledge about the
interaction between genes is still lacking; thus, further research
into the role of the miR-106b-25 cluster is required. This research
should include further bioinformatical analysis, which greatly
improves the efficiency of miRNA research, as it can provide
important information about genes and also provide targets for
further experiments.
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2008. View Article : Google Scholar
|
|
2
|
Ning Z, Zhu H, Li F, Liu Q, Liu G, Tan T,
Zhang B, Chen S, Li G, Huang D, et al: Tumor suppression by miR-31
in esophageal carcinoma is p21-dependent. Genes Cancer. 5:436–444.
2014.PubMed/NCBI
|
|
3
|
Guohong Z, Min S, Duenmei W, Songnian H,
Min L, Jinsong L, Hongbin L, Feng Z, Dongping T, Heling Y, et al:
Genetic heterogeneity of oesophageal cancer in high-incidence areas
of southern and northern China. PLoS One. 5:e96682010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao Z, Liu J, Wang C, Wang Y, Jiang Y and
Guo M: MicroRNA-25 regulates small cell lung cancer cell
development and cell cycle through cyclin E2. Int J Clin Exp
Pathol. 7:7726–7734. 2014.PubMed/NCBI
|
|
5
|
Li HL, Xie SP, Yang YL, Cheng YX, Zhang Y,
Wang J, Wang Y, Liu DL, Chen ZF, Zhou YN and Wu HY: Clinical
significance of upregulation of mir-196a-5p in gastric cancer and
enriched kegg pathway analysis of target genes. Asian Pac J Cancer
Prev. 16:1781–1787. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee RC, Feinbaum RL and Ambros V: The C.
Elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense
Complementarity to &II-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reinhart BJ, Slack FJ, Basson M,
Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR and Ruvkun G:
The 21-nucleotide let-7 RNA regulates developmental timing in
Caenorhabditis elegans. Nature. 403:901–906. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee NK, Lee JH, Park CH, Yu D, Lee YC,
Cheong JH, Noh SH and Lee SK: Long non-coding HOTAIR promotes
carcinogenesis and invasion of gastric adenocarcinoma. Biochem
Biophys Res Commun. 451:171–178. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Carninci P, Kasukawa T, Katayama S, Gough
J, Frith MC, Maeda N, Oyama R, Ravasi T, Lenhard B, Wells C, et al:
The transcriptional landscape of the mammalian genome. Science.
309:1559–1563. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lai EC, Tomancak P, Williams RW and Rubin
GM: Computational identification of Drosophila microRNA genes.
Genome Biology. 4:R422003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Axtell MJ, Westholm JO and Lai EC: Vive la
différence: Biogenesis and evolution of microRNAs in plants and
animals. Genome Biol. 12:2212011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Altuvia Y, Landgraf P, Lithwick G, Elefant
N, Pfeffer S, Aravin A, Brownstein MJ, Tuschl T and Margalit H:
Clustering and conservation patterns of human microRNAs. Nucleic
Acids Res. 33:2697–2706. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu R, Liao J, Yang M, Sheng J, Yang H,
Wang Y, Pan E, Guo W, Pu Y, Kim SJ and Yin L: The Cluster of
miR-143 and miR-145 affects the risk for esophageal squamous cell
carcinoma through co-regulating fascin homolog. PLoS One.
7:e339872012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Khuu C, Utheim TP and Sehic A: The Three
Paralogous MicroRNA Clusters in Development and Disease, miR-17-92,
miR-106a-363, andmiR-106b-25. Scientifica (Cairo).
2016:13796432016.PubMed/NCBI
|
|
15
|
Smith AL, Iwanaga R, Drasin DJ, Micalizzi
DS, Vartuli RL, Tan AC and Ford HL: The miR-106b-25 cluster targets
Smad7, activates TGF-β signaling and induces EMT and tumor
initiating cell characteristics downstream of Six1 in human breast
cancer. Oncogene. 31:5162–5171. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feber A, Xi L, Luketich JD, Pennathur A,
Landreneau RJ, Wu M, Swanson SJ, Godfrey TE and Litle VR: MicroRNA
Expression Profiles of Esophageal Cancer. J Thorac Cardiovasc Surg.
135:255–260. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Antonov AV, Dietmann S and Mewes HW: KEGG
spider: interpretation of genomics data in the context of the
global gene metabolic network. Genome Biol. 9:R1792008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kimura S, Naganuma S, Susuki D, Hirono Y,
Yamaguchi A, Fujieda S, Sano K and Itoh H: Expression of microRNA
in squamous cell carcinoma of human head and neck and the
esophagus: miR-205 and miR-21 are specific markers for HNSCC and
ESCC. Oncol Rep. 23:1625–1633. 2010.PubMed/NCBI
|
|
20
|
Matsumura Y, Hiraoka K, Ishikawa K, Shoji
Y, Noji T, Hontani K, Itoh T, Nakamura T, Tsuchikawa T, Shichinohe
T and Hirano S: CD40 expression in human esophageal squamous cell
carcinoma is associated with tumor progression and lymph node
metastasis. Anticancer Res. 36:4467–4475. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Tan W, Neo TW, Aung MO, Wasser S,
Lim SG and Tan TM: Role of the miR-106b-25 microRNA cluster in
hepatocellular carcinoma. Cancer Sci. 100:1234–1242. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kan T, Sato F, Ito T, Matsumura N, David
S, Cheng Y, Agarwal R, Paun BC, Jin Z, Olaru AV, et al: The
miR-106b-25 polycistron, activated by genomic amplification,
functions as an oncogene by suppressing p21 and Bim.
Gastroenterology. 136:1689–1700. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Petrocca F, Visone R, Onelli MR, Shah MH,
Nicoloso MS, de Martino I, Iliopoulos D, Pilozzi E, Liu CG, Negrini
M, et al: E2F1-regulated microRNAs impair TGFbeta-dependent
cell-cycle arrest and apoptosis in gastric cancer. Cancer Cell.
13:272–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Spender LC and Inman GJ: TGF-beta induces
growth arrest in Burkitt lymphoma cells via transcriptional
repression of E2F-1. J Biol Chem. 284:1435–1442. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen HZ, Tsai SY and Leone G: Emerging
roles of E2Fs in cancer: An exit from cell cycle control. Nat Rev
Cancer. 9:785–797. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saha MN, Qiu L and Chang H: Targeting p53
by small molecules in hematological malignancies. J Hematol Oncol.
6:232013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu X, Chen Z, Zhao X, Wang J, Ding D, Wang
Z, Tan F, Tan X, Zhou F, Sun J, et al: MicroRNA-25 promotes cell
migration and invasion in esophageal squamous cell carcinoma.
Biochem Biophys Res Commun. 421:640–645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fu X, Tian J, Zhang L, Chen Y and Hao Q:
Involvement of microRNA-93, a new regulator of PTEN/Akt signaling
pathway, in regulation of chemotherapeutic drug cisplatin
chemosensitivity in ovarian cancer cells. FEBS Lett. 586:1279–1286.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Poliseno L, Salmena L, Riccardi L, Fornari
A, Song MS, Hobbs RM, Sportoletti P, Varmeh S, Egia A, Fedele G, et
al: Identification of the miR-106b~25 microRNA cluster as a
proto-oncogenic PTEN-targeting intron that cooperates with its host
gene MCM7 in transformation. Sci Signal. 3:ra292010. View Article : Google Scholar : PubMed/NCBI
|