Introduction
In recent years, the use of blood components for
purposes other than classic transfusion has become more popular. It
has emerged that blood cells and/or plasma-derived factors may have
therapeutic effects in various diseases. There is increasing
interest in the use of platelet derivatives, particularly
platelet-rich plasma (PRP), which is classically described as ‘a
volume of plasma that has a platelet count above baseline (of whole
blood)’ (1). Platelets are not only
involved in a well-documented haemostatic function, but also play a
fundamental role in tissue repair and regeneration. They have the
ability to store numerous growth factors (GFs), enzymes and other
bioactive molecules, which are rapidly released following platelet
activation (2–4). Platelets are also able to stimulate
certain key events in the reparative processes, such as replication
of cells of mesenchymal origin (including fibroblasts, osteoblasts
and endothelial cells), or chemotactic effects (2–5).
Various methods have been developed for the topical
use of platelet concentrates (PCs) in order to stimulate tissue
growth and regeneration. Their clinical use in wound regeneration
has gradually expanded over recent years, being applied in various
fields such as ophthalmology, pain management, oral and
maxillofacial surgery, orthopedics, plastic, periodontal and
cardiac surgery and sports medicine (5–11).
PRP contributes to tissue healing since it is a
carrier of all the individual elements involved in this process
(including platelets, leukocytes and GFs). Establishing the role
and importance of each of these factors is essential in order to
identify the most effective product to induce tissue healing.
The purpose of the present study was to evaluate to
what extent the presence of leukocytes in PRP influences its
ability to induce, in vitro, the biological activities that
are necessary for wound repair in cell types that are involved in
the tissue regeneration process. These cell types included
endothelial cells (required for angiogenesis) and fibroblasts
(required for matrix remodeling).
Materials and methods
Platelet gel (PG)-released supernatant
preparation
Whole blood samples (each 450 ml) were collected by
standard homologous blood donation using triple bags (Teruflex with
CPD; S.A.G.M., Terumo, Rome, Italy). Each donor provided consent
according to current laws (Decree Law 3, March 2005, and Law 21,
October 2005, n. 219).
PRP was produced from the blood donation of each
patient. Fractionation was carried out by initial centrifugation of
the bag for 10 min at 22°C at 462 × g using a Heraeus Cryofuge
6000i centrifuge [AHSI SpA, Massa Martana (PG), Italy] to obtain
PRP and red cell concentrates. Subsequently, the obtained PRP was
subjected to a second centrifugation for 6 min at 22°C at 3,932 × g
to produce the PC and platelet-poor plasma. Finally, the platelets
were hyperconcentrated in 10–15 ml of plasma.
This process produced leukocyte-rich PC (LR-PC),
which was later divided into two aliquots. One aliquot was deprived
of leukocytes through filtration on TERUFLEX BP-KIT®
filters (Terumo, Rome, Italy) to obtain leukocyte-poor PC
(LP-PC).
In order to measure platelet content and, when
necessary, the depletion of leukocytes, both whole blood and
obtained PCs were analyzed using a Coulter®
Ac·T 5diff AL hemocytometer (Beckman Coulter, Inc.,
Brea, CA, USA).
LR-PG or LP-PG was produced by placing LR-PC or
LP-PC, respectively, in Vacutainer Plus tubes (BD Biosciences,
Plymouth, UK) containing 5 NIH units thrombin. Calcium gluconate
[Bioindustria Laboratorio Italiano Medicinali SpA, Novi Ligure
(AL), Italy] was added at a 1:20 dilution. Subsequently, the
solutions were allowed to clot for 5–10 min at 37°C. After a
further 20–25 min, the clots were fully retracted and were
centrifuged for 10 min at 153 × g to obtain a supernatant that was
rich in GFs released from the activated platelets and, where
present, leukocytes. The supernatants were subjected to a
succession of centrifugations to remove red cells, debris and
cellular stroma, and were immediately used in experiments. For
convenience, LR-PG- and LP-PG-derived supernatants will be
identified hereafter as ‘LR-PGs’ and ‘LP-PGs’.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
isolated from umbilical cord veins. The cells were grown at 37°C
and 5% CO2 on 1% gelatin-coated flasks in DMEM
supplemented with 10% fetal bovine serum (FBS), 10% newborn calf
serum (NCS), 20 mM N-(2-hydroxyethyl)
piperazine-N′-(2-ethanesulfonic acid) (HEPES), 6 U/ml heparin, 2 mM
glutamine, 50 µg/ml endothelial cell growth factor (ECGF),
penicillin and streptomycin. The cells were used between the third
and fifth passage.
Normal human dermal fibroblasts (NHDFs) were
purchased from Lonza (Walkersville, MD, USA) and grown at 37°C and
5% CO2 in DMEM supplemented with 10% FBS, 2 mM
glutamine, penicillin and streptomycin. The cells were used before
reaching their 15th doubling, as recommended by the supplier.
HEPES and ECGF were purchased from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). FBS, DMEM, glutamine, penicillin
and streptomycin were purchased from Euroclone SpA (Milan, Italy).
NCS was purchased from Gibco (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA).
Cell proliferation
Cell proliferation was determined using an XTT assay
(Sigma-Aldrich; Merck KGaA). Living cells metabolically reduce XTT
mixed with phenazine methosulfate (Sigma-Aldrich; Merck KGaA) to
produce a colored, non-toxic, water-soluble formazan; its optical
density is directly proportional to the number of viable cells.
Briefly, 1,250 HUVECs/well were seeded into a 1%
gelatin-coated 96-well plate, then treated with LR-PGs and LP-PGs
(0–2.5×106 platelets/µl), diluting the original
preparation with DMEM supplemented with HEPES, heparin, ECGF and 2%
FBS; the same medium was used for the control (untreated cells).
NHDFs were seeded at 1,000 cells/well into a 96-well plate and then
treated with LR-PGs or LP-PGs (0–3×106 platelets/µl),
diluting the original preparation with DMEM supplemented with 1%
FBS; the same medium was used for the control (untreated
cells).
After seeding, HUVECs and NHDFs were incubated for
24 or 48 h, respectively, in complete medium at 37°C and 5%
CO2 to enable cell adhesion and spreading prior to PG
treatments. Once treated, cells were incubated at 37°C with 5%
CO2 for 72 h. At the end of this period, an XTT assay
was performed and the optical density was evaluated at 450 nm. XTT
tests were performed before the cells reached confluence to prevent
possible artifact decreases in the results due to contact
inhibition.
Each experiment was performed in triplicate and
repeated at least twice. Data are expressed as the mean ± standard
deviation.
Tube formation assay
A tube formation assay was performed to measure the
ability of endothelial cells to invade, migrate, organize, and
differentiate into capillary-like tubular structures. This was
assessed within a three-dimensional matrix constituted by 10 mg/ml
Matrigel Growth Factor Reduced (BD Biosciences, Franklin Lakes, NJ,
USA). HUVECs (20,000 cells/well) were seeded onto Matrigel-coated
96-well plates in DMEM containing 1% FBS or in LR-PGs or LP-PGs
diluted in the same medium to reach the desired concentration.
After 6 h, the formation of tubes was photographed using an
inverted optical microscope (Zeiss, Oberkochen, Germany) supported
by a Nikon camera (Nikon, Tokyo, Japan). The images were
independently scored by two blinded observers. Several images were
acquired per well and processed using the Angiogenesis Analyzer
plugin on ImageJ software.
In vitro wound-healing assay
The wound-healing assay is one of the earliest
developed tests to study directional cell migration in
vitro. It is based on the observation of cell migration into a
scratch ‘wound’ created on a cell monolayer.
HUVECs and NHDFs were cultured in 24-well
microplates under normal culture conditions and allowed to reach
maximum confluence. A previously sterilized round-tipped steel
needle was used to create several scratch wounds of approximately
0.2 mm in the cellular stratum. The microplates were washed three
times with DMEM and the cells were cultured in a low 1% FBS medium
(untreated cells) or with LR-PGs and LP-PGs diluted to the desired
concentrations (0.3×106, 1.5×106 and
2.5×106 platelets/ml) with the same medium. The status
of the scratch wounds was controlled using contrast-phase
microscopy at the beginning of the assay and at regular intervals.
Representative images were obtained (after 6 h for HUVECs and 24 h
for NHDFs) using a Nikon camera (Nikon, Tokyo, Japan). Where
possible, the percentage of closed area relative to the original
wound area was measured using ImageJ software.
Zymography
Gelatin zymography was performed using sodium
dodecyl sulfate-polyacrylamide gels (SDS-PAGE; 7.5%) co-polymerized
with 1 mg/ml gelatin type B (Sigma-Aldrich; Merck KGaA).
Casein-plasminogen zymography was performed using 10% SDS-PAGE
co-polymerized with 0.2% casein and 10 mg/ml plasminogen (both from
Sigma-Aldrich; Merck KGaA). Zymographies were performed on
supernatants diluted in SDS-PAGE sample buffer in non-reducing
conditions without heating; volumes corresponding to
5×106 and 20×106 platelets (for gelatin and
casein-plasminogen zymography, respectively) were loaded for both
LR-PGs and LP-PGs. After electrophoresis, the gels were washed
twice for 30 min in 50 mM Tris (pH 7.4) with 2.5% Triton X-100 at
room temperature and incubated overnight at 37°C in activating
buffer [50 mM Tris-HCl (pH 7.4), containing 5 mM CaCl2
and 150 mM NaCl for gelatinases; 50 mM Tris (pH 7.4), for
plasminogen activators (PAs)]. The gels were stained with Coomassie
Blue R 250 (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
dissolved in a mixture of methanol-acetic acid-water (4:1:5) for 1
h. The gels were then washed in the same solution without dye.
Enzyme activity was visualized as distinct bands, indicating
proteolysis of the substrate. Images were recorded and band
intensities were quantified using the Alliance LD2 system (UVItec,
Cambridge, UK).
Quantification of GFs
LR-PGs and LP-PGs samples were assayed for vascular
endothelial growth factor (VEGF), thrombospondin-1 (TSP-1),
interferon-γ (IFN-γ), platelet derived growth factor (PDGF)-BB, -AA
and -B, tumor growth factor-β1 (TGF-β1), tumor necrosis factor-α
(TNF-α) and fibroblast growth factor-7 (FGF-7, also known as
keratinocyte growth factor). Commercially available human ELISA
kits were used. VEGF-A ELISA kit was purchased from Diaclone
(Besançon, France; cat no. 650.080.096). TSP-1 ELISA kit was
purchased from Cusabio Biotech Co., Ltd. (College Park, MD, USA;
cat no. CSB-E08763 h). IFN-γ, PDGF-BB, -AA and -B, TGF-β1, TNF-α
and FGF-7 ELISA kits were purchased from Elabscience (Bethesda, MD,
USA; cat nos. E-EL-H0108, E-EL-H1577, E-EL-H1575, E-EL-H1670,
E-EL-H10110, E-EL-H0109 and E-EL-H0092). Samples and standards were
analyzed in duplicate according to the manufacturer's protocol. The
concentration of each GF in the original standard or sample was
proportional to the amount of signal produced and was extrapolated
from standard curves.
GFs were also measured to test their stability over
time and temperature. Freshly obtained supernatants from 2 LR-PG
were prepared as described above and divided into aliquots. One
aliquot was immediately stored at −80°C to prevent GF degradation,
while the others were stored at 4, 22 or 37°C for 48 or 168 h. As
soon as the incubation time expired, the supernatants were stored
at −80°C. At 37°C, incubation periods of 24 and 72 h were also
evaluated. Samples were successively analyzed using ELISA kits.
Platelet and white blood cell counts of LR-PGs and
LP-PGs samples used for GF quantification are presented in Table I.
 | Table I.Platelets and white blood cell count
of samples used for growth factor quantification. |
Table I.
Platelets and white blood cell count
of samples used for growth factor quantification.
|
| Leukocyte-rich
platelet concentrate | Leukocyte-poor
platelet concentrate |
|---|
|
|
|
|
|---|
| Sample | Platelets | White blood
cells | Platelets | White blood
cells |
|---|
| 1 |
2.35×106/µl |
8.6×103/µl |
1.99×106/µl | 0 |
| 2 |
3.42×106/µl |
13.2×103/µl |
2.39×106/µl | 0 |
| 3 |
2.09×106/µl |
11.8×103/µl |
1.69×106/µl | 0 |
| 4 |
2.56×106/µl |
10.6×103/µl |
2.74×106/µl | 0 |
| 5 |
1.84×106/µl |
9.8×103/µl |
1.63×106/µl | 0 |
| 6 |
2.41×106/µl |
11.6×103/µl |
|
|
| 7 |
1.85×106/µl |
8.1×103/µl |
|
|
Statistical analysis
All data shown are from at least three independent
experiments and are expressed as the mean ± standard deviation.
Statistically significant differences between groups were
determined using the Student's t-test. Calculations were performed
using GraphPad Prism 4 software (GraphPad Software, Inc., La Jolla,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
LR-PGs and LP-PGs affect
angiogenesis-related processes
The current study aimed to evaluate whether
leukocyte depletion could alter the effectiveness of PG at
stimulating angiogenesis-related activities in endothelial cells
(12,13). Thus, in vitro experiments were
conducted on HUVECs treated with different concentrations of LR-PGs
and LP-PGs.
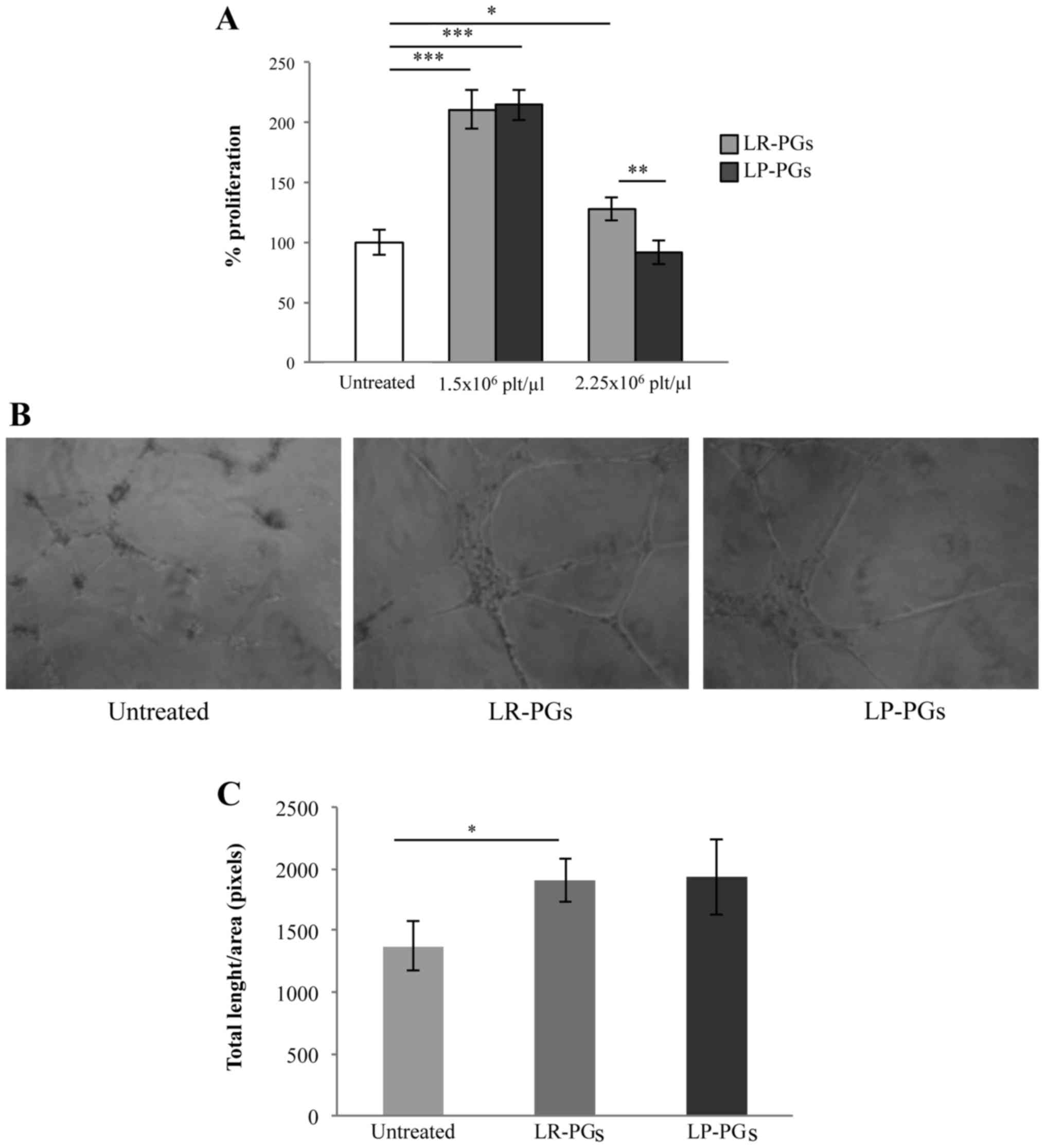
Cells were treated with 1.5×106 and
2.5×106 platelets/µl for 72 h. At the lower
concentration, both LR-PGs and LP-PGs stimulated proliferation to a
greater extent (approximately 2-fold) compared with untreated
cells, and no significant differences between LR-PGs and
LP-PGs-treated cells were observed. The higher platelet
concentration exhibited a weak stimulation in LR-PGs, while LP-PGs
appeared to have a slight negative effect compared with untreated
cells (Fig. 1A).
To investigate any differences in the ability of
LR-PGs and LP-PGs to induce formation of a capillary-like network,
a tube formation assay was performed. LR-PGs and LP-PGs
(1.5×106 platelets/µl) both stimulated the formation of
more structured tubes compared with untreated cells (Fig. 1B). No significant differences were
observed in the effects of LR-PGs and LP-PGs (Fig. 1C).
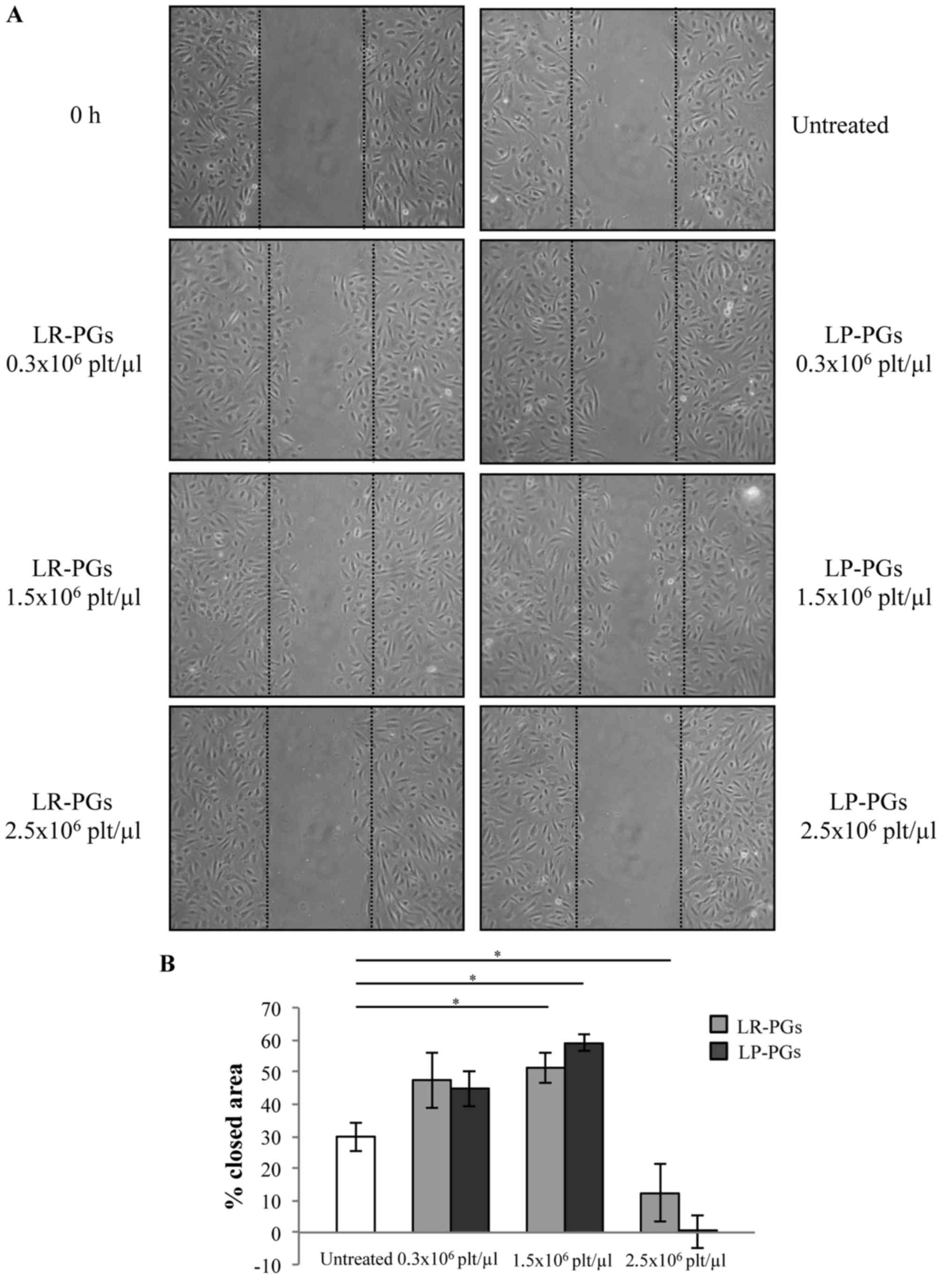
The effect of LR-PGs and LP-PGs on HUVEC motility
was evaluated using a wound-healing assay. At lower concentrations
(0.3×106 and 1.5×106 platelets/µl), cells
were able to heal the wound to a greater extent compared with the
higher concentration (2.5×106 platelets/µl; Fig. 2). With the exception of the highest
concentration (at which LP-PGs exhibited a greater negative effect
compared with LR-PGs), no significant differences were observed
between LR-PGs- and LP-PGs-treated cells.
LR-PGs and LP-PGs affect human
fibroblast behavior
The role of PG-derived GFs on human fibroblasts
cells has been described previously (14). The present study investigated whether
leukocyte depletion could affect some of the cell properties
associated with tissue repair, such as proliferative and motile
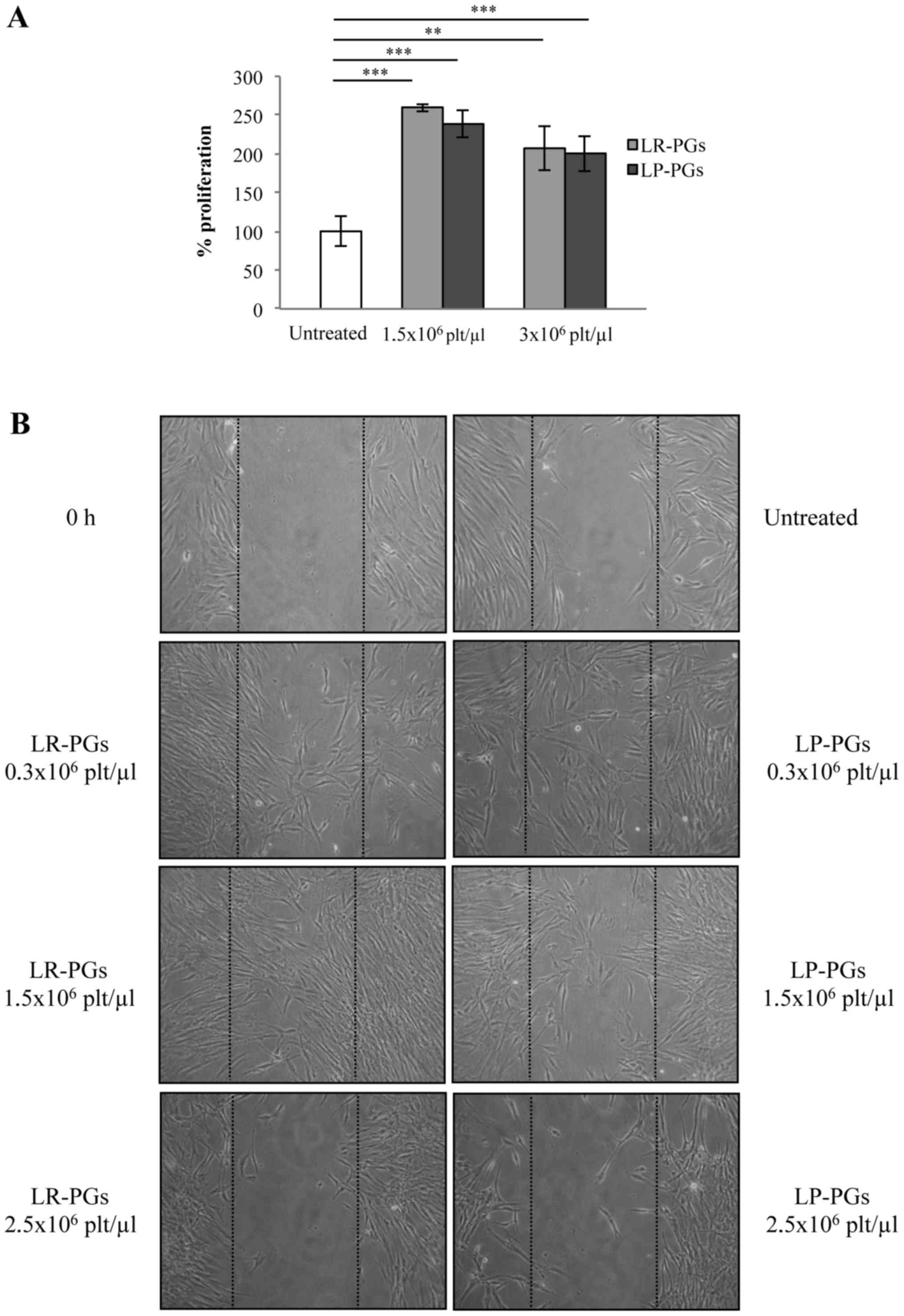
capacities. Fibroblasts were treated with LR-PGs or LP-PGs at
concentrations of 1.5×106 and 3.0×106
platelets/µl and proliferation was evaluated. Both supernatants
stimulated proliferation to a greater extent (approximately
2.5-fold) compared with untreated cells. No significant differences
in the proliferative response were observed between LR-PGs- and
LP-PGs-treated cells (Fig. 3A).
In a wound-healing assay, cells stimulated with
lower concentrations (0.3×106 and 1.5×106
platelets/µl) were able to migrate to a greater extent compared
with untreated cells. At a higher concentration (2.5×106
platelets/µl) fibroblast motility was strongly inhibited (Fig. 3B). No marked differences were evident
between LR-PGs- and LP-PGs-treated cells, with the exception of the
1.5×106 platelets/µl concentration, where LR-PGs
appeared to exert a greater effect on cell motility compared with
LP-PGs.
Molecular characterization of LR-PGs
and LP-PGs
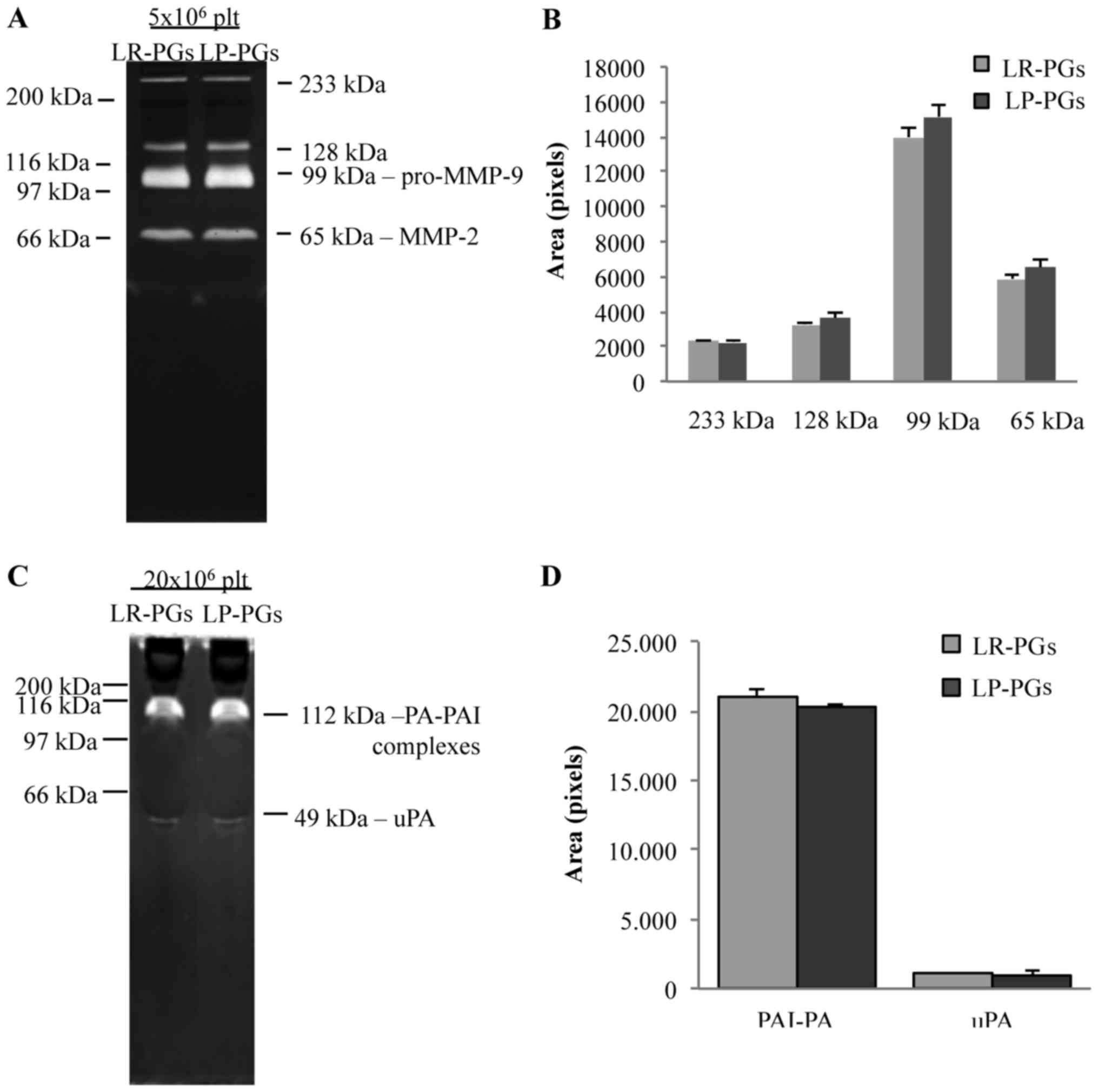
The activity of gelatinases (MMP-2 and MMP-9) and
PAs was assessed using zymography techniques (Fig. 4). Zymography on gelatin substrate
identified in both samples (LR-PGs and LP-PGs) the active form of
MMP-2 (65 kDa), the pro-enzyme form of MMP-9 (99 kDa) and two
high-molecular-weight bands (128 and 233 kDa), likely corresponding
to gelatinase complexes with their inhibitors (tissue inhibitors of
metalloproteinases) (Fig. 4A). No
statistically significant differences were evident between the
samples (Fig. 4B).
Similarly, zymography of casein-plasminogen
substrate showed no significant differences between the two
analyzed samples (Fig. 4D). In both
samples, the urokinase-type PA (49 kDa) and the complex of PA with
its own inhibitor (112 kDa) were present (Fig. 4C).
GF content was analyzed in 5 PGs (PGs 1–5 from
Table I) using ELISA kits. The
results are presented in Table II.
For the majority of GFs, there were no significant differences in
GFs content of LR-PGs and LP-PGs. The only exceptions were GP 4 (in
which there was a significant decrease in VEGF in LP-PGs compared
with LR-PGs), GPs 2 and 5 (in which there was a significant
increase in IFN-γ in LP-PGs compared with LR-PGs) and GP3 (in which
there was a significant increase in PDGF-B in LP-PGs compared with
LR-PGs). TNF-α and FGF-7 levels were also measured, but were not
detectable.
| Table II.GFs content in LR-PG and LP-PG
derived supernatants. |
Table II.
GFs content in LR-PG and LP-PG
derived supernatants.
| Variables | VEGF pg/ml | TSP-1 µg/ml | INFγ pg/ml | PDGF-BB pg/ml | PDGF-AA pg/ml | PDGF-B pg/ml | TGF-β1 µg/ml |
|---|
| LR-PG 1 | 1,774.4±87.8 | 567.5±51.3 | 402.4±11.4 | 38.7±4.5 | 16.3±1.4 | 20.9±0.8 | 44.3±5.7 |
| LP-PG 1 | 1,954.1±24.7 | 576.5±23.6 | 510.9±57.1 | 29.5±6.7 | 15.1±3.1 | 289±4.7 | 54.2±4.8 |
| LR-PG 2 | 686.7±15.9 | 342.1±16.7 | 241.6±15.9 | 40.4±3.8 | 33.9±7.4 | 59.1±10.2 | 36.8±7.9 |
| LP-PG 2 | 712.3±20.5 | 483.2±23.8 | 438.8±2.2 | 63.9±4.7 | 58.3±12.1 | 72.1±9.1 | 59.4±6.9 |
| LR-PG 3 | 1,166.0±7.8 | 559.1±19.7 | 547.5±4.4 | 125.7±11.3 | 57.7±4.2 | 80.4±0.8 | 64.6±4.2 |
| LP-PG 3 | 1,080.1±51.3 | 501.3±18.7 | 653.6±35.0 | 99.4±8.7 | 60.6±0.8 | 93.6±3.6 | 88.8±9.8 |
| LR-PG 4 | 1,423.9±10.6 | 487.3±39.6 | 362.1±11.3 | 117.7±9.7 | 54.9±1.1 | 101.6±9.4 | 58.9±4.7 |
| LP-PG 4 | 1,261.5± 31.8 | 474.9±41.3 | 353.6±13.5 | 144.3±12.1 | 63.3±7.9 | 80.7±2.3 | 48.7±6.9 |
| LR-PG 5 | 2,663.3±60.1 | 421.9±54.8 | 282.9±10.7 | 162.9±8.2 | 72±5.9 | 86.4±5.3 | 43.8±11.3 |
| LP-PG 5 | 2,464.8±73.6 | 546.7±51.3 | 471.5±6.0 | 141.6±9.2 | 76.2±5.2 | 97.8±32.9 | 73.0±8.7 |
Effects of time and temperature on GF
stability
Two samples (PGs 6 and 7 from Table I) were assayed to verify whether
short-term storage at different temperatures could cause
degradation of GFs. VEGF, TSP-1, IFN-γ, PDGF-AA and PDGF-B levels
were measured using ELISA kits. The results are presented in
Table III. For each sample, no
significant differences were observed in aliquots maintained for
24–168 h at 4, 22 or 37°C compared with the aliquot at point
zero.
| Table III.Time and temperature effects on GFs
contained in LR-PG derived supernatants. |
Table III.
Time and temperature effects on GFs
contained in LR-PG derived supernatants.
| Variables | VEGF pg/ml | TSP-1 µg/ml | INFγ pg/ml | PDGF-AA pg/ml | PDGF-B pg/ml |
|---|
| PG 6 point
zero | 2,217.0±94.9 | 196.7±17.4 | 918.0±41.8 | 49.1±11.1 | 30.5±2.1 |
| PG 6 48 h 4° | 2,122.7±24.9 | 152.3±9.6 | 913.3±20.6 | 44.1±11.4 | 35.7±2.7 |
| PG 6 1 week 4° | 1,927.9±11.3 | 148.2±9.6 | 936.9±5.6 | 52.5±3.1 | 31.2±3 |
| PG 6 48 h 22° | 2,130.7±31.2 | 169.8±11.8 | 834.6±108.9 | 43.5±8.9 | 36.5±2.6 |
| PG 6 1 week
22° | 2,022.1±58.7 | 244.8±17.5 | 932.5±31.9 | 57.3±7.5 | 28.7±4.2 |
| PG 6 24 h 37° | 2,119.5±15.8 | 161.1±10.3 | 902.0±28 | 43.4±9.2 | 27.9±3 |
| PG 6 48 h 37° | 2,202.6±169.4 | 160.4±9.5 | 949.1±8.4 | 39.7±4.9 | 24.9±3.7 |
| PG 6 72 h 37° | 2,453.4±207.8 | 157.2±13.6 | 900.2±77.5 | 41.6±9.2 | 35.1±2.1 |
| PG 6 1 week
37° | 2,547.6±24.8 | 146.2±13.9 | 794.6±21.2 | 41.9±9.5 | 37.8±5.2 |
| PG 7 point
zero | 656.3±44.2 | 206.4±12.9 | 1,195.7±13.6 | 27.2±4.9 | 22.1±5.4 |
| PG 7 48 h 4° | 539.6±56 | 198.2±18.8 | 1,189.1±51.9 | 26.2±3.2 | 21.4±4.7 |
| PG 7 1 week 4° | 541.7±23.6 | 214.2±11.8 | 1,243.7±2.9 | 25.1±2.4 | 40.1±8.1 |
| PG 7 48 h 22° | 591.7±5.9 | 213.2±11.4 | 1,206.6±28.3 | 26.5±2.2 | 24±4.1 |
| PG 7 1 week
22° | 606.3±14.7 | 210.7±5.7 | 1,245.7±25.9 | 24.3±0.9 | 22.2±3.2 |
| PG 7 24 h 37° | 658.4±41.3 | 164.7±14.3 | 1,245.7±23.6 | 26.9±0.4 | 22±2.1 |
| PG 7 48 h 37° | 623.0±26.5 | 177.4±12.1 | 1,220.3±24.7 | 28.3±3.5 | 37.1±9.6 |
| PG 7 72 h 37° | 785.5±55.9 | 170.2±15.9 | 1,245.73±25.6 | 45.0±24.1 | 21.8±1.4 |
| PG 7 1 week
37° | 887.6±70.7 | 224.0±8.6 | 1,221.4±34.5 | 24.6±1.6 | 23±1.2 |
Discussion
It is well established that platelet-enriched blood
derivatives, such as PRP, contribute to tissue healing, since they
carry all the elements involved in this process (platelets,
leukocytes and GFs). Establishing the role and importance of each
of these factors is essential in order to identify the most
effective product to induce tissue healing. As the breadth of
knowledge has increased in this field, and the clinical use of PRP
has become more common, novel methods of preparation have been
developed, offering specific platelet/leukocyte concentrations
(15). Different classifications of
PCs have been proposed based on their content of leukocytes; LR-PRP
and LP-PRP both contain hyperconcentrated platelets but with and
without leukocytes, respectively (16–18).
It is still a subject of debate whether leukocytes
have a positive or negative effect on healing processes (15). They may have beneficial effects
because they stimulate the immune response against infections
(19), but it has been recently
demonstrated that L-PRP and P-PRP generally inhibit bacterial
growth in a comparable way (20);
leukocytes also increase GF release, contributing to angiogenesis,
matrix production and hypercellularity (21). On the other hand, leukocytes may
release inflammatory cytokines (such as TNF-α and reactive oxygen
species) that may increase inflammation, having a detrimental
effect by delaying tissue healing (17,22). In
addition, leukocytes could increase levels of MMPs, which could
play an important role in causing matrix degradation and, thus,
inferior repair of wounded tissues and scar formation (15); some MMPs, including MMP-9, are stored
in circulating platelets and also in neutrophils and are released
after their activation (23).
Certain studies, mainly conducted in the orthopedic field, have
suggested that LP-PRP could induce more effective tissue healing
when compared to LR-PRP (15,24,25),
while other studies highlight no significant differences between
the two (26,27).
It is likely that both LR- and LP-PRP could have
positive or negative effects depending on the particular clinical
conditions (2,17). In our opinion, there are some aspects
still to be examined in relation to the effects of leukocytes
contained in PRP on the healing process. In particular, studies
outside the areas of tendinopathy and orthopedics are required, in
order to investigate effects in a generic wound, where several cell
systems are involved. These include endothelial cells, which are
responsible for angiogenesis, the formation of new blood vessels
that support healing by providing nutrients, promoting granulation
tissue formation and facilitating the clearance of debris (28). Fibroblasts are also involved in wound
healing, since they play a major role in producing collagen and
other matrix components. The current paper, therefore, aimed to
provide in vitro data about the positive or negative effects
of leukocytes contained in PRP in the healing process, in the
context of PG preparations/formulations.
For this purpose, we obtained PCs from whole blood
that were divided into two aliquots, one of which was maintained
unaltered (LR-PC), while the other one was leukocyte-depleted by
means of filtration (LP-PC). Platelet clotting to obtain LR-PG and
LP-PG was induced by calcium and thrombin addition, in order to
mimic a ‘physiological’ activation. During the 30-min period from
first clot formation to supernatant recovery, both platelets and
leukocytes should be activated; platelet activation and
degranulation are simultaneous to clotting, and leukocytes are
activated by PDGF released from platelets in a short time period
(29,30). The obtained supernatant contains GFs
and other biological active molecules released from platelets and
leukocytes during activation, which support the tissue
repair/regeneration process (2–4,31). This was previously demonstrated to be
equivalent, in terms of its in vitro effect, to PG itself
(13). Thus, in the present study,
for greater ease of use, experiments were performed with
supernatants. Since preliminary studies already highlighted that
different concentrations (expressed as platelets/µl) exert
different biological effects (12–14,32),
specific concentrations were examined. Finally, since the
concentration of biologically relevant molecules released from
platelets is proportional to the initial concentration of
platelets/µl, we assumed that the supernatant would maintain the
same platelets/µl concentration of PG from which it was obtained,
even if the platelets were no longer physically present.
HUVECs were tested for the characteristics required
for the angiogenesis process, such as proliferation and motility.
As it was already known that some concentrations are more effective
than others (13), cells were
treated with 1.5×106 platelets/µl and 2.5×106
platelets/µl (optimal and counterproductive concentrations for
endothelial cells activity, respectively). LR-PGs and LP-PGs both
stimulated proliferation and motility, but no significant
differences between LR-PGs- and LP-PGs-treated cells were observed.
The tube formation assay, which was only conducted at the optimal
concentration for endothelial cells, indicated the same trend.
Human fibroblasts were also evaluated for
proliferation and motility in response to LR-PGs and LP-PGs. The
results were consistent with those obtained for endothelial cells.
In the wound-healing assay, unlike HUVEC cells (that move into the
wound as a compact layer) fibroblasts migrate into the wounded
space individually. This made it impossible to perform an accurate
quantification of the closed area; however, upon simple observation
of the images, it is evident that at lower concentrations, cells
are able to migrate into the wound to a greater extent compared
with untreated cells, whereas motility was strongly inhibited at
the highest concentration. No notable differences, however, were
observed between LR-PGs- and LP-PGs-treated cells.
In conclusion, it seems that LR-PGs and LP-PGs are
equally effective in inducing biological activities relevant for
tissue healing in endothelial cells and fibroblasts if used at
optimal concentrations. At higher, detrimental concentrations, the
LP-PGs exerts a slight negative effect compared with LR-PGs.
Enzyme levels were measured in order to assess
whether LR-PGs and LP-PGs contribute differently in terms of
proteolytic enzyme activity during tissue repair. White blood cells
release MMPs and PAs, which are able to remodel the extracellular
matrix (33–35); however, if excessive amounts are
released, this could counteract tissue healing (17). Zymographies showed similar levels of
expression of both gelatinases and PAs, suggesting that leukocyte
depletion does not affect these enzyme activities.
In order to understand whether leukocyte removal
impoverishes the PG in terms of GFs that promote healing processes,
it was critical to assess GFs in LR-PGs and LP-PGs. Some studies on
GFs in PRP have already been conducted, but these are not relevant
if quantification was performed on non-activated platelet
suspension; in order to properly measure GF content, the PRP needs
to be activated in PG (2,21). In the present study, several GFs were
assayed to measure their levels in LR-PGs and LP-PGs. The GFs we
selected are involved in various aspects of wound repair: VEGF has
a leading role in angiogenesis (36); TSP-1 has a well-defined role in
latent TGF-β1 activation and in inflammation (37,38);
IFN-γ can delay the wound healing process (39); PDGFs are involved in tissue
remodeling (40); TGF-β affects
inflammation, angiogenesis and fibroblast activities (41); TNF-α stimulates fibroblast
proliferation and angiogenesis (42); and FGF-7 stimulates proliferation,
migration and angiogenesis (42).
When LR-PGs and LP-PGs were compared for each GF, there were few
differences, generally supporting the proposal that no differences
are induced by the leukocyte depletion process. A small number of
GFs showed significant differences: In GP 4, the VEGF level
decreased in the LP-PGs compared with LR-PGs, while in GPs 2 and 5,
the IFN-γ level increased in LP-PGs compared with LR-PGs. We
hypothesized that the eventual lower content of VEGF (which
positively drives angiogenesis) and higher content of IFN-γ (which
negatively affects wound healing) in LP-PGs could partially explain
the more negative effect induced by LP-PGs in the proliferation and
motility tests.
The present data confirmed the ability of PG to
stimulate the biological mechanisms involved in the
repair/regeneration of wounds. The data also revealed a substantial
overlap of performance between LR-PG and LP-PG, which could
probably be explained by a substantial equivalence in GFs and
enzyme content. GFs and enzymes are not negatively affected by
leukocyte depletion and are still present in sufficient amounts to
sustain in vitro repair of endothelial cell and fibroblast
cultures.
Finally, in a preliminary way, some GFs were also
analyzed in two samples to verify how short-term storage at various
temperatures could affect GF stability. The results did not support
the hypothesis that preservation for short periods (24 h to 1 week)
at easily obtainable temperatures [such as refrigerator temperature
(~4°C) or room temperature (~22°C)] can influence the content in
GFs. Even maintaining the samples at body temperature (37°C) for 24
h to 1 week did not significantly affect the stability of the GFs,
suggesting that they remained fully active during the entire
duration of clinical treatments. However, assays with a larger
number of samples are required to confirm these findings.
Although the present study was limited by the
absence of in vivo experiments, we hypothesize that, in
in vivo applications, a priori removal of leukocytes
is not a fundamental requirement to obtain greater efficacy of PG.
Furthermore, each clinical situation will require careful
consideration. Data from the analyses on GF stability also suggest
it may be possible to design PG formulations that can be safely
stored by patients in a domestic setting and autonomously applied
(for example, eye drops for ophthalmologic treatments) (9). This would avoid the requirement for
daily medical attention.
Acknowledgements
This study was partially supported by Fondazione
Cassa di Risparmio della Provincia dell'Aquila.
References
|
1
|
Marx RE: Platelet-rich plasma (PRP): What
is PRP and what is not PRP? Implants Dent. 10:225–228. 2001.
View Article : Google Scholar
|
|
2
|
Arnoczky SP, Delos D and Rodeo SA: What is
platelet-rich plasma? Oper Tech Sports Med. 19:142–148. 2011.
View Article : Google Scholar
|
|
3
|
Blair P and Flaumenhaft R: Platelet
alpha-granules: Basic biology and clinical correlates. Blood Rev.
23:177–189. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leslie M: Cell biology. Beyond clotting:
The powers of platelets. Science. 328:562–564. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Knezevic NN, Candido KD, Desai R and Kaye
AD: Is platelet-rich plasma a future therapy in pain management?
Med Clin North Am. 100:199–217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lacci KM and Dardik A: Platelet-rich
plasma: Support for its use in wound healing. Yale J Biol Med.
83:1–9. 2010.PubMed/NCBI
|
|
7
|
Mihaylova Z, Mitev V, Stanimirov P, Isaeva
A, Gateva N and Ishkitiev N: Use of platelet concentrates in oral
and maxillofacial surgery: An overview. Acta Odontol Scand.
75:1–11. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fréchette JP, Martineau I and Gagnon G:
Platelet-rich plasmas: Growth factor content and roles in wound
healing. J Dent Res. 84:434–439. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van der Meer PF, Seghatchian J and Marks
DC: Quality standards, safety and efficacy of blood-derived serum
eye drops: A review. Transfus Apher Sci. 54:164–167. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hsu WK, Mishra A, Rodeo SR, Fu F, Terry
MA, Randelli P, Canale ST and Kelly FB: Platelet-rich plasma in
orthopaedic applications: Evidence-based recommendations for
treatment. J Am Acad Orthop Surg. 21:739–748. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee KS: Platelet-rich plasma injection.
Semin Musculoskelet Radiol. 17:91–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rughetti A, Giusti I, D'Ascenzo S, Leocata
P, Carta G, Pavan A, Dell'Orso L and Dolo V: Platelet gel-released
supernatant modulates the angiogenic capability of human
endothelial cells. Blood Transfus. 6:12–17. 2008.PubMed/NCBI
|
|
13
|
Giusti I, Rughetti A, D'Ascenzo S,
Millimaggi D, Pavan A, Dell'Orso L and Dolo V: Identification of an
optimal concentration of platelet gel for promoting angiogenesis in
human endothelial cells. Transfusion. 49:771–778. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giusti I, Rughetti A, D'Ascenzo S, Di
Stefano G, Nanni MR, Millimaggi D, Dell'orso L and Dolo V: The
effects of platelet gel-released supernatant on human fibroblasts.
Wound Repair Regen. 21:300–308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cross JA, Cole BJ, Spatny KP, Sundman E,
Romeo AA, Nicholson GP, Wagner B and Fortier LA: Leukocyte-reduced
platelet-rich plasma normalizes matrix metabolism in torn human
rotator cuff tendons. Am J Sports Med. 43:2898–2906. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dohan Ehrenfest DM, Bielecki T, Mishra A,
Borzini P, Inchingolo F, Sammartino G, Rasmusson L and Evert PA: In
search of a consensus terminology in the field of platelet
concentrates for surgical use: Platelet-rich plasma (PRP),
platelet-rich fibrin (PRF), fibrin gel polymerization and
leukocytes. Curr Pharm Biotechnol. 13:1131–1137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou Y, Zhang J, Wu H, Hogan MV and Wang
JH: The differential effects of leukocyte-containing and pure
platelet-rich plasma (PRP) on tendon stem/progenitor
cells-implications of PRP application for the clinical treatment of
tendon injuries. Stem Cell Res Ther. 6:1732015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duif C, Vogel T, Topcuoglu F, Spyrou G,
von Schulze Pellengahr C and Lahner M: Does intraoperative
application of leukocyte-poor platelet-rich plasma during
arthroscopy for knee degeneration affect postoperative pain,
function and quality of life? A 12-month randomized controlled
double-blind trial. Arch Orthop Trauma Surg. 135:971–977. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moojen DJ, Everts PA, Schure RM,
Overdevest EP, van Zundert A, Knape JT, Castelein RM, Creemers LB
and Dhert WJ: Antimicrobial activity of platelet-leukocyte gel
against Staphylococcus aureus. J Orthop Res. 26:404–410. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mariani E, Canella V, Berlingeri A, Bielli
A, Cattini L, Landini MP, Kon E, Marcacci M, Di Matteo B and
Filardo G: Leukocyte presence does not increase microbicidal
activity of Platelet-rich Plasma in vitro. BMC Microbiol.
15:1492015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dohan Ehrenfest DM, Bielecki T, Jimbo R,
Barbé G, Del Corso M, Inchingolo F and Sammartino G: Do the fibrin
architecture and leukocyte content influence the growth factor
release of platelet concentrates? An evidence-based answer
comparing a pure platelet-rich plasma (P-PRP) gel and a leukocyte-
and platelet-rich fibrin (L-PRF). Curr Pharm Biotechnol.
13:1145–1152. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dragoo JL, Braun HJ, Durham JL, Ridley BA,
Odegaard JI, Luong R and Arnoczky SP: Comparison of the acute
inflammatory response of two commercial platelet-rich plasma
systems in healthy rabbit tendons. Am J Sports Med. 40:1274–1281.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hasty KA, Pourmotabbed TF, Goldberg GI,
Thompson JP, Spinella DG, Stevens RM and Mainardi CL: Human
neutrophil collagenase: A distinct gene product with homology to
other matrix metalloproteinases. J Biol Chem. 265:11421–11424.
1990.PubMed/NCBI
|
|
24
|
McCarrel TM, Minas T and Fortier LA:
Optimization of leukocyte concentration in platelet rich plasma for
the treatment of tendinopathy. J Bone Joint Surg Am.
94:e143(18)2012. View Article : Google Scholar
|
|
25
|
Yin WJ, Xu HT, Sheng JG, An ZQ, Guo SC,
Xie XT and Zhang CQ: Advantages of pure platelet-rich plasma
compared with leukocyte- and platelet-rich plasma in treating
rabbit knee osteoarthritis. Med Sci Monit. 22:1280–1290. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Filardo G, Kon E, Pereira Ruiz MT, Vaccaro
F, Guitaldi R, Di Martino A, Cenacchi A, Fornasari PM and Marcacci
M: Platelet-rich plasma intra-articular injections for cartilage
degeneration and osteoarthritis: Single-vs. double-spinning
approach. Knee Surg Sports Traumatol Arthrosc. 20:2082–2091. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Riboh JC, Saltzman BM, Yanke AB, Fortier L
and Cole BJ: Effect of leukocyte concentration on the efficacy of
platelet rich plasma in the treatment of knee osteoarthritis. Am J
Sports Med. 44:792–800. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Diegelmann RF and Evans MC: Wound healing:
An overview of acute, fibrotic and delayed healing. Front Biosci.
9:283–289. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tzeng DY, Deuel TF, Huang JS and Baehner
RL: Platelet-derived growth factor promotes human peripheral
monocyte activation. Blood. 66:179–183. 1985.PubMed/NCBI
|
|
30
|
Elstad MR, McIntyre TM, Prescott SM and
Zimmerman GA: The interaction of leukocytes with platelets in blood
coagulation. Curr Opin Hematol. 2:47–54. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Grazul-Bilska AT, Johnson ML, Bilski JJ,
Redmer DA, Reynolds LP, Abdullah A and Abdullah KM: Wound healing:
The role of growth factors. Drugs Today (Barc). 39:787–800. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Giusti I, D'Ascenzo S, Mancò A, Di Stefano
G, Di Francesco M, Rughetti A, Dal Mas A, Properzi G, Calvisi V and
Dolo V: Platelet concentration in platelet-rich plasma affects
tenocyte behavior in vitro. Biomed Res Int. 2014:6308702014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Granelli-Piperno A, Vassalli JD and Reich
E: Secretion of plasminogen activator by human polymorphonuclear
leukocytes. Modulation by glucocorticoids and other effectors. J
Exp Med. 146:1693–1706. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Salamonsen LA and Lathbury LJ: Endometrial
leukocytes and menstruation. Hum Reprod Update. 6:16–27. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xue M, Thompson PJ, Clifton-Bligh R,
Fulcher G, Gallery ED and Jackson C: Leukocyte matrix
metalloproteinase-9 is elevated and contributes to lymphocyte
activation in type I diabetes. Int J Biochem Cell Biol.
37:2406–2416. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hoeben A, Landuyt B, Highley MS, Wildiers
H, Van Oosterom AT and DeBruijn EA: Vascular endothelial growth
factor and angiogenesis. Pharmacol Rev. 56:549–580. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Soto-Pantoja DR, Shih HB, Maxhimer JB,
Cook KL, Ghosh A, Isenberg JS and Roberts DD: Thrombospondin-1 and
CD47 signaling regulate healing of thermal injury in mice. Matrix
Biol. 37:25–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Blanco-Mezquita JT, Hutcheon AE and Zieske
JD: Role of thrombospondin-1 in repair of penetrating corneal
wounds. Invest Ophthalmol Vis Sci. 54:6262–6268. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Laato M, Heino J, Gerdin B, Kähäri VM and
Niinikoski J: Interferon-gamma-induced inhibition of wound healing
in vivo and in vitro. Ann Chir Gynaecol. 90 Suppl 205:S19–S23.
2001.
|
|
40
|
Martínez CE, Smith PC and Palma Alvarado
VA: The influence of platelet-derived products on angiogenesis and
tissue repair: A concise update. Front Physiol. 6:2902015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Penn JW, Grobbelaar AO and Rolfe KJ: The
role of the TGF-β family in wound healing, burns and scarring: A
review. Int J Burns Trauma. 2:18–28. 2012.PubMed/NCBI
|
|
42
|
Rozman P and Bolta Z: Use of platelet
growth factors in treating wounds and soft-tissue injuries. Acta
Dermatovenerol Alp Pannonica Adriat. 16:156–165. 2007.PubMed/NCBI
|