Introduction
The prevalence of mental disorders such as
depression has increased over the past few years and such disorders
negatively affect the quality of life of many people around the
world (1). It has been demonstrated
that environmental stress is a risk factor for a number of mental
disorders. Exposure to psychological and physiological-related
chronic stress contributes to the dysfunction of
hypothalamic-pituitary-adrenal (HPA) axis and causes a sustained
elevation of glucocorticoid (GC) levels, which may result in the
development of mental disorders (2).
In vitro, a high concentration of GCs suppresses neuronal
proliferation, growth and differentiation and may even inhibit
neuronal cell death (3). It has been
demonstrated that a number of classical antidepressants protect
against the cytotoxicity induced by high concentrations of GCs in
neural cells (4).
The consumption of green tea is very popular
worldwide and it may exhibit beneficial pharmacological actions,
including anti-oxidant, anti-carcinogenic, anti-tumor and
anti-neurodegenerative effects (5–7). A
number of studies have demonstrated that the green tea induces
marked anti-depressive effects in animal models of depression
(8,9)
and that catechins, the major polyphenolic compounds found in green
tea, decrease depression- and anxiety-like behaviors in a rat model
induced following the injection of chronic corticosterone (CORT)
(10). The catechin
(−)-epigallocatechin-3-gallate (EGCG) is the primary constituent of
the green tea polyphenols and exhibits the strongest biological
activity, including antioxidant, anti-inflammatory and
neuroprotective effects (11,12). It
has been demonstrated that EGCG significantly improves chronic
unpredictable mild stress-induced behavior alterations in rats
(13) and exerts neuroprotective
effects against different types of stress-related damage, including
trauma and neuronal damage induced by L-3,4-dihydroxyphenylalanine
(L-DOPA) (14,15). However, few studies have been
completed to assess the effect of EGCG on neural cells damaged
following exposure to high concentrations of CORT.
The adrenal phaeochromocytoma PC12 cell line is one
of the most widely used neuronal cell lines and is commonly used to
study the neuronal damage induced by GCs in vitro (16) and also used as an in vitro
experimental model of depression (17,18).
Although previous studies have suggested that EGCG has
neuroprotective effects, the effect of EGCG on neuronal cells
exposed to high concentrations of CORT remains to be elucidated.
The present study examined the neuroprotective activity and
associated potential mechanisms of EGCG in CORT-injured PC12
cells.
Materials and methods
Materials
The PC12 cell line was supplied by the Central
Laboratory of the Central Hospital of Wuhan (Wuhan, China). The
RPMI 1640 medium was purchased from Hyclone; GE Healthcare Life
Sciences (Logan, UT, USA). Fetal bovine serum (FBS) was purchased
from Zhejiang Tianhang Biotechnology Co., Ltd. (Hangzou, China).
EGCG (>97.0%) was purchased from Selleck Chemicals (Houston, TX,
USA). CORT (>97.0%) and the hedgehog-smoothened inhibitor
cyclopamine (>99.0%) were purchased from Shanghai Aladdin
Biochemical Technology Co., Ltd., (Shanghai, China). Desipramine
(DIM; >98.0%), a well-known antidepressant (19) which was used as the positive control
(18) in the present study, was
purchased from Sigma Aldrich; Merck KGaA (Darmstadt, Germany). All
pharmacological agents were prepared as a stock solution and stored
at −20°C.
Cell culture and treatment
PC12 cells were maintained in 1640 supplemented with
10% fetal bovine serum, 100 U/ml penicillin and 100 µg/ml
streptomycin in a humidified atmosphere containing 5%
CO2 atmosphere at 37°C. Cells were seeded at a density
of 8×103 cells/well in 96-well plates and incubated for
2–3 days for MTT assay; cells were seeded at a density of
4×103 cells/well in 96-well plates and incubated for 1
days for the CCK8 assay. The cells were divided into five groups:
Control group, where PC12 cells were not treated; CORT group, where
PC12 cells were treated with CORT; CORT+EGCG group, where PC12
cells were treated with CORT and EGCG, and EGCG was added 1 h
before CORT; CORT+DIM group, where PC12 cells were treated with
CORT and DIM, and DIM was added 1 h before CORT; and
Cyclopamine+EGCG+CORT group, where PC12 cells were treated CORT,
EGCG and cyclopamine, and cyclopamine was added 30 min before EGCG,
and then EGCG was added 1 h before CORT.
MTT assay
A
3-(4,5-Desethyithiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT,
Biosharp, Inc., Pivotal Scientific, Wuhan, China) assay was used to
measure cell viability. Cells were seeded at a density of
8×103 cells/well in 96-well plates and cultured, and
following incubation of the cells with the different drugs for 24
h, they were treated with the MTT solution and incubated at 37°C
for 4 h. The dark blue formazan crystals that formed in the wells
were solubilized with dimethyl sulfoxide for 10 min at 37°C.
Absorbance at 570 nm was measured using a microplate reader
(PerkinElmer, Inc, Waltham, MA, USA).
Morphological changes
Following treatment of PC12 cells with different
drugs, grown medium was removed by PBS. Cellular morphology was
observed using a fluorescence microscope (BX-50-FLA, Olympus
Corporation, Tokyo, Japan).
Hoechst 33342 staining
Cells were treated with different drugs and
incubated for 24 h. The medium was subsequently replaced by Hoechst
33342 (Beyotime Biotechnology Institute of Biotechnology, Nanjing,
China) solution and incubated at 37°C for 15 min. Cells were then
observed under an inverted fluorescence microscope (BX-50-FLA,
Olympus Corporation). Apoptotic cells exhibited strong blue
fluorescence and shrunken nuclei, whereas non-apoptotic cells
exhibited weak blue fluorescence and normal nuclei.
Cell counting kit-8 (CCK-8) assay
A CCK-8 assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) was used to measure cell proliferation. Cells were
seeded and cultured in 96-well plates and after the cells were
treated with the different drugs and incubated for 0, 12, 24, 36,
48 and 60 h, the medium was replaced with CCK-8 reagent (Dojindo
Molecular Technologies, Inc.) and cells were incubated for a
further 2 h at 37°C. Finally, the optical density of each well was
measured using a microplate reader at a wavelength of 450 nm. The
percentage of the control samples of each cell line was calculated
thereafter.
Preparation of total RNA and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Cells were treated with the aforementioned drugs and
cultured for 24 h. Subsequently, cells were harvested and total RNA
was extracted using TRIzol reagent (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The purity of total RNA was evaluated using the
optical density 260/280 ratio and a UV spectrophotometer (Beckman
Coulter, Inc., Brea, CA, USA). RNA purity was determined to be
between 1.7 and 2.0.
Reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) was performed to determine mRNA expression of
proteins in the hedgehog signaling pathway. Reverse transcription
was performed using cDNA Synthesis kit (Toyobo, Co., Ltd., Osaka,
Japan) following the manufacturer's protocol. cDNA was subjected to
qPCR using the Power SYBR® Green PCR Master mix
(CWBIOTECH, Beijing, China). The specific sequences of the primers
used were as follows: GAPDH, forward 5′-GATGGTGAAGGTCGGTGTGA-3′ and
reverse 5′-GTCAATGAAGGGGTCGTTGA-3′; Shh, forward
5′-TATGAGGGTCGAGCAGTGGA-3′ and reverse 5′-AGTGGATGCGAGCTTTGGAT-3′;
Gli1, forward 5′-GTCACTACCTGGCCTCACAC-3′ and reverse
5′-CCCCTGCATTGGGGTTGTAT-3′; and N-myc, forward
5′-GATGACTTCTACTTCGGCGGT-3′ and reverse 5′-CCAAACGCATCCTCCTCGG-3′.
qPCR was performed under the following conditions: 95°C for 30 sec,
followed by 45 cycles at 95°C for 5 sec, 60°C for 30 sec in the ABI
7900 Real-Time PCR System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The relative quantification of gene expression
was calculated using the 2−ΔΔCq method (20). Each gene analysis was performed in
triplicate.
Western blot analysis
Cells were lysed with ice-cold
radioimmunoprecipitation assay lysis buffer with 10% phenylmethane
sulfonyl fluoride (Beyotime Institute of Biotechnology, Haimen,
China) and the protein was collected after being centrifuged at
12,000 × g for 30 min at 4°C. Protein concentration was measured
using a BCA protein assay kit (Beyotime Institute of Biotechnology)
following the manufacturer's protocol. Equal amounts of protein (20
µg per lane) were electrophoresed on 10% density SDS acrylamide
gels and then proteins were transferred to nitrocellulose
membranes. Non-specific binding was blocked with 5% skim milk in
TBST buffer at 4°C for 2–3 h, prior to incubation with rabbit
anti-N-myc antibody (1:200, sc-791; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) and rabbit anti-GAPDH antibody (1:1,000, sc-25778;
Santa Cruz Biotechnology, Inc.,) overnight at 4°C. Membranes were
then incubated with FITC-conjugated goat anti-rabbit IgG secondary
antibody (1:10,000, BL033A; Biosharp Inc., Pivotal Scientific) for
1 h at room temperature. Finally, detection was performed using
enhanced chemiluminscence (GE Healthcare Life Sciences) and
quantified using Quantity One 4.6.2 imaging software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Data were expressed as the mean ± standard error of
the mean. Multiple group comparisons were performed using one-way
analysis of variance (ANOVA) followed by Dunnett's test using
GraphPad Prism 5.0 software (GraphPad software, Inc., La Jolla, CA,
USA) to detect any inter-group differences. P<0.05 was
considered to indicate a statistically significant difference.
Results
EGCG protects against the cytotoxicity
induced by CORT
Differentiated PC12 cells were treated with 50, 100,
150, 250 and 400 µM CORT for 24 h. Subsequently, the effect of CORT
on PC12 cell viability was determined using a MTT assay. Different
concentrations of CORT decreased cell viability in a dose-dependent
manner (Fig. 1A). The concentration
of 400 µM CORT, which significantly decreased cell viability to
35.6% (P<0.01; Fig. 1A) was
selected for subsequent experiments. The protective effects of
different concentrations of EGCG was subsequently investigated.
EGCG induced no cytotoxic effects at concentrations between 1 and
20 µM (Fig. 1B), although
concentrations of EGCG >30 µM significantly decreased cell
viability compared with the control (P<0.05). It was also
demonstrated that EGCG attenuated the decrease in cell viability
induced by CORT in a dose-dependent manner and that the activity of
EGCG was optimal at a concentration of 20 µM (Fig. 1C). The viability of PC12 cells
treated with 20 µM EGCG and 400 µM CORT increased significantly by
compared with cells that underwent treatment with 400 µM CORT alone
(P<0.01; Fig. 1C). DIM
significantly increased the viability of PC12 cells (P<0.05) at
a concentration of 1 µM compared with the group treated with 400 µM
CORT alone (Fig. 1D). To determine
the protective effect of EGCG on CORT-injured PC12 cells, the
morphological changes of PC12 cells treated with different drugs
were observed using an inverted microscope (Fig. 1E). The morphology of cells treated
with 400 µM CORT changed and the axons shrunk markedly. However,
these alterations were attenuated when cells in the 400 µM CORT
group were pretreated with 20 µM EGCG or 1 µM DIM. Thus, EGCG may
protect PC12 cells against the cytotoxicity induced by CORT.
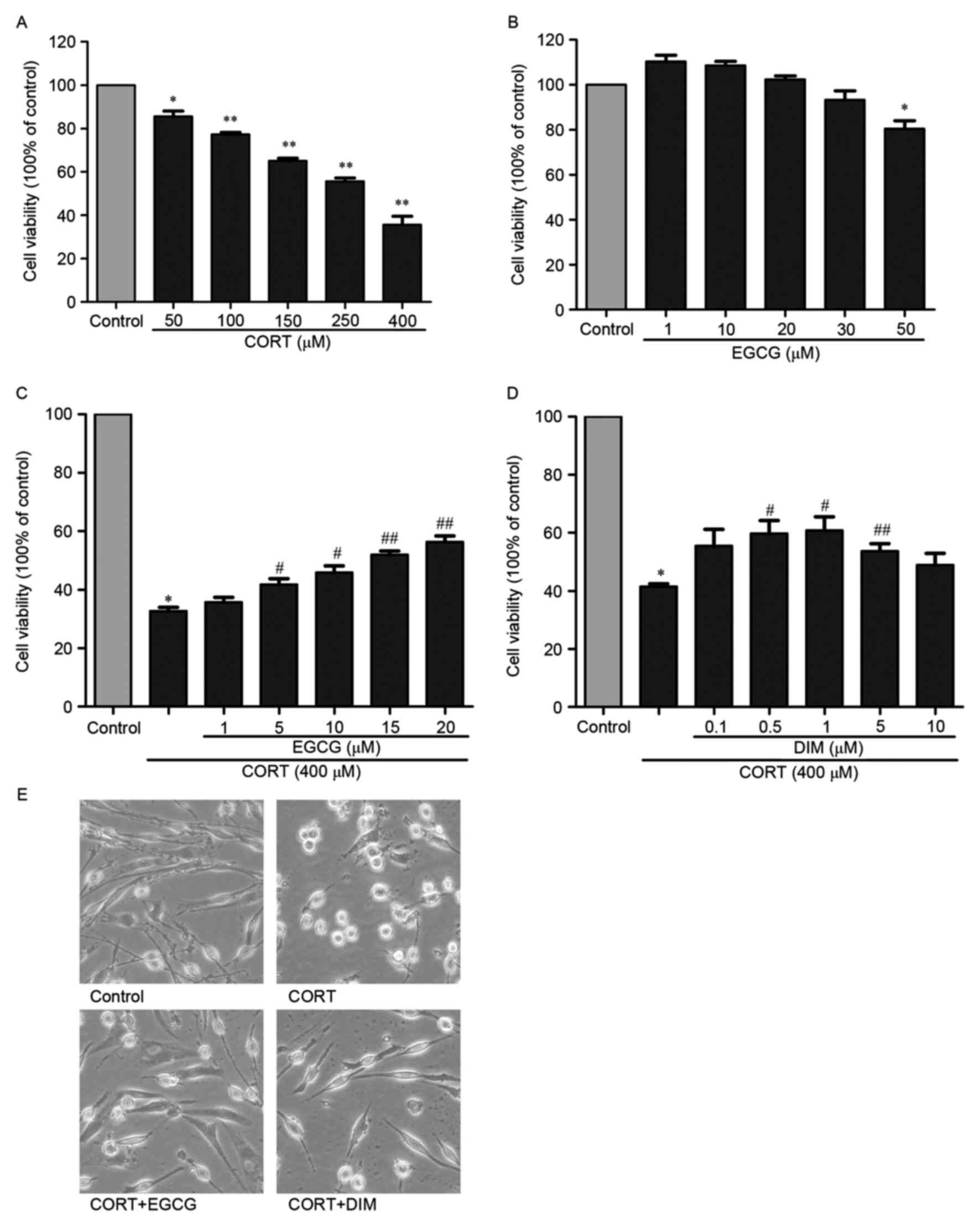 | Figure 1.Protective effect of EGCG against the
cytotoxicity induced by CORT in PC12 cells. (A) The viability of
PC12 cells treated with 50, 100, 150, 250, 400 µM CORT was measured
using an MTT assay 24 h after treatment of cells with CORT. (B) The
viability of PC12 cells treated with 1, 10, 20, 30, 50 µM EGCG was
measured using an MTT assay 24 h after cells were treated with
EGCG. (C) The viability of PC12 cells treated with 1, 5, 10, 15, 20
µM EGCG and 400 µM CORT was measured using an MTT assay. PC12 cells
were pretreated with different concentrations of EGCG 1 h before
treatment with 400 µM CORT and incubated for 24 h prior to the MTT
assay. (D) The viability of PC12 cells treated with 0.1, 0.5, 1, 5,
10 µM DIM (positive control) and 400 µM CORT was measured using an
MTT assay. PC12 cells were pretreated with different concentrations
of DIM 1 h before incubation with 400 µM CORT for 24 h prior to an
MTT assay. (E) Images of cell morphology were taken 24 h after the
cells were treated with 400 µM CORT alone, 400 µM CORT and 20 µM
EGCG or 400 µM CORT and 1 µM DIM (magnification, ×200). All results
are presented as the mean ± standard error of the mean of three
independent experiments performed in triplicate. *P<0.05 and
**P<0.01 vs. control; #P<0.05 and
##P<0.01 vs. CORT group. EGCG,
(−)-Epigallocatechin-3-gallate; CORT, corticosterone; DIM,
desipramine. |
EGCG attenuates CORT-induced cell
apoptosis and inhibition of cell proliferation
Cellular apoptosis induced by different drugs was
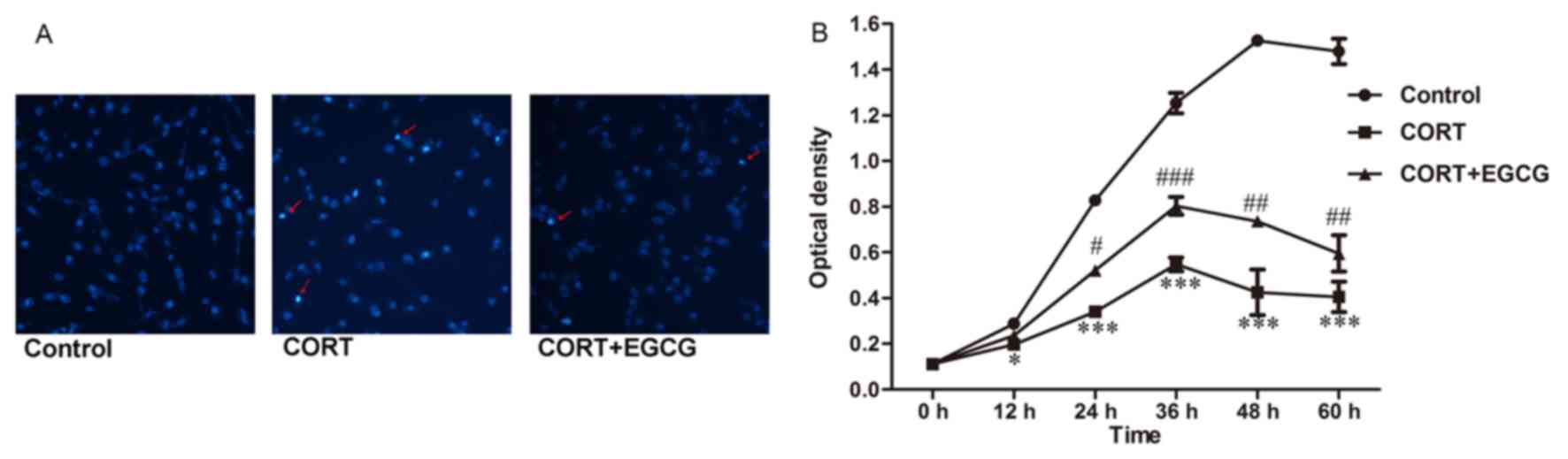
detected by staining with Hoechst 33342 (Fig. 2A). The results demonstrated that
treatment with 400 µM CORT markedly increased the amount of
condensed chromatin in PC12 cells. The addition of 20 µM EGCG
decreased the amount of condensed chromatin compared with the CORT
group. The optical density (OD) of PC12 cells treated with 400 µM
CORT for 0, 12, 24, 36, 48 and 60 h was determined using a CCK8
assay. Compared with the control group, the OD of the cells were
significantly decreased when treated with 400 µM CORT for 12
(P<0.05, Fig. 2B), 24
(P<0.001, Fig. 2B), 36
(P<0.001, Fig. 2B), 48
(P<0.001, Fig. 2B) and 60 h
(P<0.001, Fig. 2B). The OD of the
cells pretreated with 20 µM EGCG increased significantly when
compared with the OD of cells treated 400 µM CORT for 24
(P<0.05, Fig. 2B), 36
(P<0.001, Fig. 2B), 48
(P<0.01, Fig. 2B) and 60 h
(P<0.01, Fig. 2B). These findings
suggested that cellular proliferation was reduced following
exposure to 400 µM CORT; however, this effect was reversed
following treatment with 20 µM EGCG. These results indicate that
EGCG may attenuate cellular apoptosis and the inhibition of cell
proliferation induced by CORT.
CORT downregulates the Shh signaling
pathway
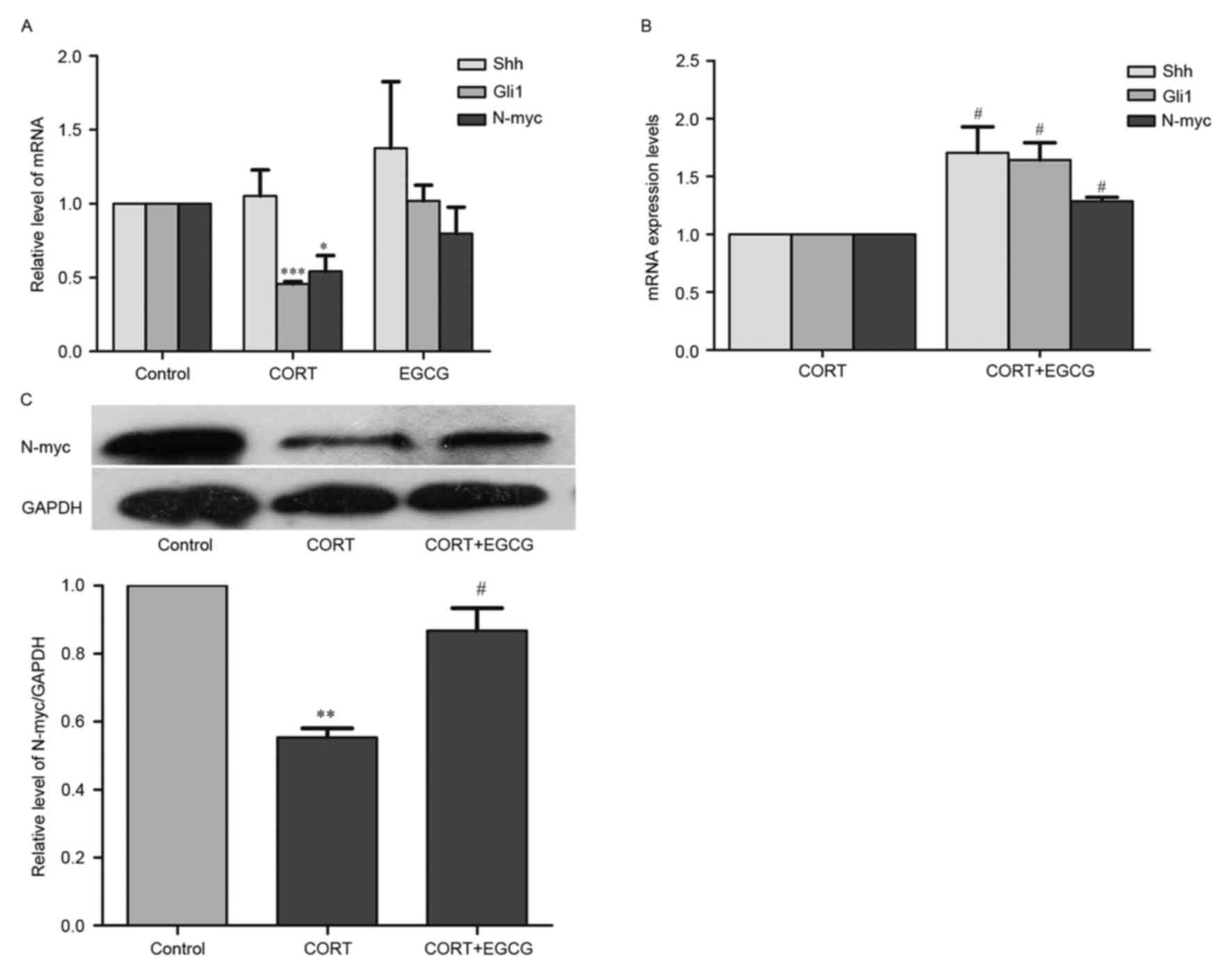
To determine whether the Shh signaling pathway
regulates CORT-induced cytotoxicity, the levels of Shh, N-myc and
Gli1 mRNA were measured using RT-qPCR. The expression of N-myc
protein was evaluated using western blot analysis to determine the
downstream effects of the Shh signaling pathway (Fig. 3). Cells were then treated with 400 µM
CORT 24 h prior to extraction of total RNA and the results of
RT-qPCR indicated that the expression of Shh mRNA did not differ
significantly between control cells and cells treated with 400 µM
CORT (Fig. 3A). However, in PC12
cells treated with 400 µM CORT, levels of Gli1 and N-myc mRNA
(Fig. 3A) as well as levels of N-myc
protein (Fig. 3C) were significantly
lower than controls (P<0.05), demonstrating that CORT inhibits
downstream Shh signaling.
EGCG reverses the downregulation of
Shh signaling induced by CORT
PC12 cells were co-treated with 20 µM EGCG and 400
µM CORT for 24 h to determine the effects of EGCG on CORT-induced
signaling inhibition. It was determined that 20 µM EGCG did not
significantly alter the levels of Shh, Gli1 and N-myc mRNA in
control cells (Fig. 3A), however it
reversed the decrease in the levels of Shh, Gli1 and N-myc mRNA
(P<0.05; Fig. 3B) and expression
of N-myc protein (P<0.05; Fig.
3C) induced by 400 µM CORT. This suggests that EGCG contributes
to the activation of Shh pathway in CORT-treated PC12 cells and
this signaling may be involved in the protection of EGCG in
CORT-injured PC12 cells.
The Shh signaling pathway is involved
in the protective effect of EGCG
To identify the role of Shh signaling pathway in the
protective effect of EGCG, prior to treatment with 20 µM EGCG and
400 µM CORT, cells were pretreated with 20 µM cyclopamine, which
targets the Sonic hedgehog signaling pathway by blocking the
activity of the smoothened frizzled class transmembrane receptor
(SMO). The results demonstrated that, compared with the EGCG + CORT
group, pretreatment with 20 µM cyclopamine significantly decreased
the cell viability (P<0.01; Fig.
4A), although it could not decrease the cell viability to a
similar level in the CORT group (Fig.
4A), which showed that EGCG's effect of promoting cell survival
was partly inhibited by 20 µM cyclopamine. Furthermore, compared
with the EGCG and CORT group, the axons of the cells were markedly
reduced and cell apoptosis was markedly decreased after the cells
were treated with 20 µM cyclopamine (Fig. 4B and C, respectively). Compared with
the EGCG and CORT group, the OD of the cells pretreated with 20 µM
cyclopamine were significantly decreased after treated with 400 µM
CORT for 24 (P<0.05; Fig. 4D), 36
(P<0.001) and 48 h (P<0.01; Fig.
4D). Furthermore, cyclopamine also significantly reversed the
increase in levels of Gli1 and N-myc mRNA (P<0.05; Fig. 4E) and levels of N-myc protein
(P<0.01; Fig. 4F) by 20 µM EGCG
in PC12 cells treated with 400 µM CORT. These findings imply that
the Shh signaling pathway is involved in the neuroprotective effect
of EGCG.
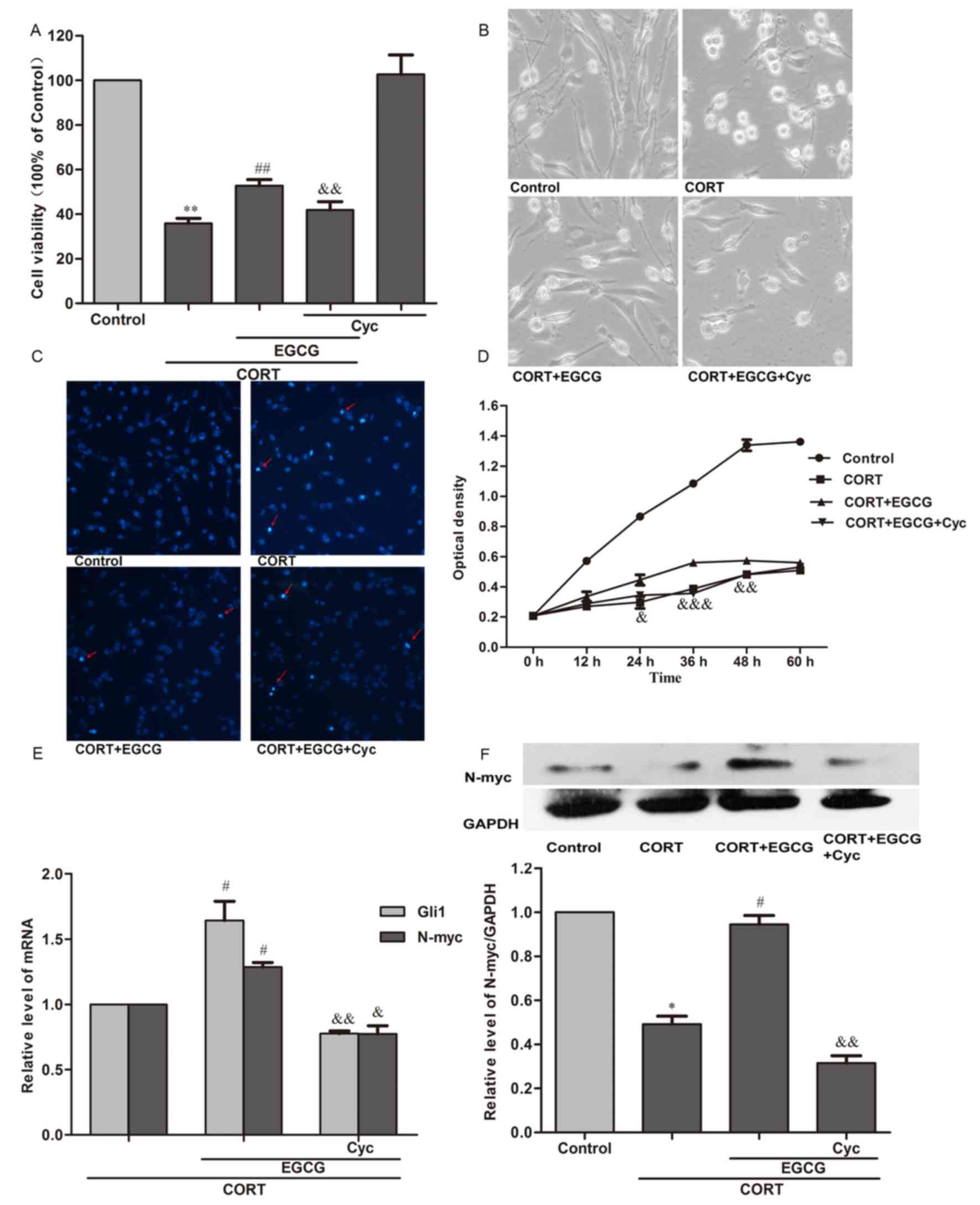 | Figure 4.Cyclopamine inhibits the
neuroprotective effect of EGCG. PC12 cells were treated with 20 µM
cyclopamine 30 min prior to treatment with 20 µM EGCG, which
occurred 1 h before treatment with 400 µM CORT. (A) Cell viability
was measured using an MTT assay, conducted 24 h following the
treatment of cells with 400 µM CORT. (B) Morphological images of
cells were taken using a fluorescence microscope (magnification,
×200) 24 h after treatment of cells with 400 µM CORT. (C) Cellular
apoptosis was measured using Hoechst 33342 staining (magnification,
×200). Red arrows indicated apoptotic nuclei. Observations were
made 24 h after the treatment of cells with 400 µM CORT. (D)
Cellular proflieration was measured using a CCK-8 assay 0, 12, 24,
36, 48 and 60 h after the treatment of cells with 400 µM CORT. (E)
The relative mRNA expression of Gli1 and N-myc was measured using
reverse transcription-quantitative polymerase chain reaction. mRNA
levels were measured 24 h after the treatment of cells with 400 µM
CORT. (F) Relative expression of N-myc protein as determined by
western blot analysis. Protein expression was measured 24 h after
the treatment of cells with 400 µM CORT. All results are expressed
as the mean ± standard error of the mean of three independent
experiments performed in triplicate. *P<0.05 and **P<0.01 vs.
control; #P<0.05 and ##P<0.01 vs. CORT
group; &P<0.05, &&P<0.01 and
&&&P<0.01 vs. CORT+EGCG group. CCK-8, cell counting
kit-8; EGCG, (−)-Epigallocatechin-3-gallate; CORT,
corticosterone. |
Discussion
Supraphysiological GC levels may have detrimental
effects on neurodevelopment (21)
and a significant chronic rise in GC levels induced by stress is
associated with pathological changes in the hippocampus, which
contributes to the pathogenesis of depression. Previous studies
have demonstrated that exposure to high levels of GCs may suppress
neuronal proliferation, growth and differentiation and lead to the
death of neuronal cells, including PC12 cells, in vitro
(3,22). Consistent with these observations,
the present study demonstrated that treatment with 400 µM CORT
induced a significant decrease in cell survival and growth as well
as the shrinking of cells, confirming that CORT induces
neurotoxicity. EGCG alleviated cytotoxicity induced by CORT but had
no effect on normal cells, indicating that EGCG reverses
CORT-induced neuronal damage.
The present study demonstrated that EGCG increases
the viability and proliferation of PC12 cells in vitro
following CORT injury and that treatment with 20 µM induces the
greatest effects. Higher doses of EGCG, such as 100 µM EGCG, are
harmful to normal PC12 cells, thus confirming the hypothesis that
the biological activity of EGCG follows a characteristic biphasic
pattern: Higher doses of EGCG exhibit pro-oxidant and pro-apoptotic
effects, whereas lower doses of EGCG have neuroprotective effects
(23,24). Previous studies have suggested that
EGCG has a neuroprotective function. It has been demonstrated that
EGCG protects the brain against traumatic injury (14), inhibits the oxidative stress induced
by 1-methyl-4-phenylpyridinium in neuronal cells (25), attenuates the neurotoxicity induced
by L-DOPA or oxygen-glucose deprivation (15,26) and
promotes the neurogenesis of adult hippocampus in vivo and
in vitro (27). The current
study confirmed that EGCG protects neuronal cells against
CORT-induced damage, indicating that EGCG is neuroprotective.
However, the precise cellular and molecular
mechanisms underlying CORT-induced neurotoxicity are not fully
understood. It has been demonstrated that in prenatally stressed
rats and hippocampus neural cells treated with high concentrations
of cortisol, the inhibition of Hedgehog (Hh) signaling by high
concentrations of GCs contributes to neurotoxicity (28) and that activation of Shh signaling is
protective against GC-induced neurotoxicity (29,30). Hh
signaling serves an important role in neuronal development and
growth, as well as the repair of damage (31) and is activated by the binding of
secreted Hh proteins, including Shh, Desert hedgehog and Indian
hedgehog, to the transmembrane receptor Patched (Ptch). Upon
binding to Ptch, the Hh protein reverses the repression of the
transmebrane receptor SMO, thus activating the major transcription
factor Gli that regulates the expression of genes, such as those
encoding N-myc, to influence the survival, growth and
differentiation of cells (32). The
present study demonstrated that Gli1 and N-myc were inhibited in
PC12 cells by high concentrations of CORT, which is similar to the
results of studies that indicated chronic GC treatment reduced the
expression of Gli1 and N-myc in neonatal brain or medulloblastoma
(30,33). Furthermore, consistent with the
results of a study by Heine et al (30), the current study did not detect any
changes in Shh expression, demonstrating that CORT acted downstream
of Shh in this pathway and that Gli1 may be a target of CORT.
Activation of Shh signaling is involved in
protection against neuronal damage (34), however little is known regarding its
role in EGCG-enhanced protection following CORT-induced
neurotoxicity. EGCG stimulates intracellular signaling pathways
associated with cell survival and growth and Shh signaling is
involved in EGCG-mediated neuroprotection (35). In the current study, it was
determined that pretreatment with EGCG attenuated the CORT-induced
downregulation of Gli1 and N-myc, implying that EGCG may contribute
to the activation of the Shh signaling pathway. By contrast, when
cells were treated with the Shh inhibitor cyclopamine prior to
EGCG, the increase in cell viability and proliferation was
reversed. Furthermore, the expression of Gli1 and N-myc were
suppressed by cyclopamine, indicating that Shh signaling
contributes to the protective effect of EGCG. Although cyclopamine
reversed the effect of EGCG on promoting cell survival, it could
not decrease the cell viability to a similar level with the CORT
group. This suggested that the increase in cell viability was not
completely blocked by cyclopamine, indicating that Shh signaling
may be not the only mechanism by which EGCG acts (13). Therefore, further studies are
required to elucidate these other potential mechanisms of
action.
In conclusion, the current study demonstrated that
EGCG exhibits protective activities and contributes to the
activation of the Shh signaling pathway in CORT-injured PC12 cells.
Furthermore, Shh signaling may be involved in the neuroprotective
effects of EGCG against CORT-induced neurotoxicity. The results of
the current study may be useful in improving understanding
regarding the neuroprotective effects of the various compounds
contained in green tea. However, further experimental studies in
vivo are required to examine the effects of EGCG and other
compounds in green tea on stress-related neurotoxicity and
neurological disorders.
Acknowledgements
The current study was supported by the National
Natural Science Foundation of China (grant no. 81603013), the Basic
Research of Wuhan Science and Technology Bureau (grant no.
2017060201010204) and the Chenguang Plan of Wuhan Manicipal Science
and Technology Bereau (grant no. 201407040401010226).
References
|
1
|
Spielmans GI, Berman MI, Linardatos E,
Rosenlicht NZ, Perry A and Tsai AC: Adjunctive atypical
antipsychotic treatment for major depressive disorder: A
meta-analysis of depression, quality of life, and safety outcomes.
PLoS Med. 10:e10014032013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Watson S, Gallagher P, Del-Estal D, Hearn
A, Ferrier IN and Young AH: Hypothalamic-pituitary-adrenal axis
function in patients with chronic depression. Psychol Med.
32:1021–1028. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao J, Peng L, Zheng W, Wang R, Zhang L,
Yang J and Chen H: Chemically bonding of amantadine with
gardenamide a enhances the neuroprotective effects against
corticosterone-induced insults in PC12 cells. Int J Mol Sci.
16:22795–22810. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li YF, Liu YQ, Huang WC and Luo ZP:
Cytoprotective effect is one of common action pathways for
antidepressants. Acta Pharmacol Sin. 24:996–1000. 2003.PubMed/NCBI
|
|
5
|
Bastianetto S, Zheng WH and Quirion R: The
Ginkgo biloba extract (EGb 761) protects and rescues hippocampal
cells against nitric oxide-induced toxicity: Involvement of its
flavonoid constituents and protein kinase C. J Neurochem.
74:2268–2277. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dong Z, Ma W, Huang C and Yang CS:
Inhibition of tumor promoter-induced activator protein 1 activation
and cell transformation by tea polyphenols, (-)-epigallocatechin
gallate, and theaflavins. Cancer Res. 57:4414–4419. 1997.PubMed/NCBI
|
|
7
|
Song DU, Jung YD, Chay KO, Chung MA, Lee
KH, Yang SY, Shin BA and Ahn BW: Effect of drinking green tea on
age-associated accumulation of Maillard-type fluorescence and
carbonyl groups in rat aortic and skin collagen. Arch Biochem
Biophys. 397:424–429. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Y, Jia G, Gou L, Sun L, Fu X, Lan N,
Li S and Yin X: Antidepressant-like effects of tea polyphenols on
mouse model of chronic unpredictable mild stress. Pharmacol Biochem
Behav. 104:27–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhu WL, Shi HS, Wei YM, Wang SJ, Sun CY,
Ding ZB and Lu L: Green tea polyphenols produce antidepressant-like
effects in adult mice. Pharmacol Res. 65:74–80. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee B, Sur B, Kwon S, Yeom M, Shim I, Lee
H and Hahm DH: Chronic administration of catechin decreases
depression and anxiety-like behaviors in a rat model using chronic
corticosterone injections. Biomol Ther (Seoul). 21:313–322. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cai J, Jing D, Shi M, Liu Y, Lin T, Xie Z,
Zhu Y, Zhao H, Shi X, Du F and Zhao G: Epigallocatechin gallate
(EGCG) attenuates infrasound-induced neuronal impairment by
inhibiting microglia-mediated inflammation. J Nutr Biochem.
25:716–725. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xie J, Jiang L, Zhang T, Jin Y, Yang D and
Chen F: Neuroprotective effects of Epigallocatechin-3-gallate
(EGCG) in optic nerve crush model in rats. Neurosci Lett.
479:26–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gu HF, Nie YX, Tong QZ, Tang YL, Zeng Y,
Jing KQ, Zheng XL and Liao DF: Epigallocatechin-3-gallate
attenuates impairment of learning and memory in chronic
unpredictable mild stress-treated rats by restoring hippocampal
autophagic flux. PLoS One. 9:e1126832014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Itoh T, Imano M, Nishida S, Tsubaki M,
Mizuguchi N, Hashimoto S, Ito A and Satou T:
(-)-Epigallocatechin-3-gallate increases the number of neural stem
cells around the damaged area after rat traumatic brain injury. J
Neural Transm (Vienna). 119:877–890. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee MY, Choi EJ, Lee MK and Lee JJ:
Epigallocatechin gallate attenuates L-DOPA-induced apoptosis in rat
PC12 cells. Nutr Res Pract. 7:249–255. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li ZY, Jiang YM, Liu YM, Guo Z, Shen SN,
Liu XM and Pan RL: Saikosaponin D acts against
corticosterone-induced apoptosis via regulation of mitochondrial GR
translocation and a GR-dependent pathway. Prog Neuropsychopharmacol
Biol Psychiatry. 53:80–89. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang Z, Mao QQ, Zhong XM, Feng CR, Pan AJ
and Li ZY: Herbal formula SYJN protect PC12 cells from
neurotoxicity induced by corticosterone. J Ethnopharmacol.
125:456–460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou H, Li X and Gao M: Curcumin protects
PC12 cells from corticosterone-induced cytotoxicity: Possible
involvement of the ERK1/2 pathway. Basic Clin Pharmacol Toxicol.
104:236–240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cappiello A, McDougle CJ, Delgado PL,
Malison RT, Jatlow P, Charney DS, Heninger GR and Price LH: Lithium
and desipramine versus desipramine alone in the treatment of severe
major depression: A preliminary study. Int Clin Psychopharmacol.
13:191–198. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Crowther CA, Doyle LW, Haslam RR, Hiller
JE, Harding JE and Robinson JS: ACTORDS Study Group: Outcomes at 2
years of age after repeat doses of antenatal corticosteroids. N
Engl J Med. 357:1179–1189. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang H, Zhou X, Huang J, Mu N, Guo Z, Wen
Q, Wang R, Chen S, Feng ZP and Zheng W: The role of Akt/FoxO3a in
the protective effect of venlafaxine against corticosterone-induced
cell death in PC12 cells. Psychopharmacology (Berl). 228:129–141.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mandel S, Weinreb O, Amit T and Youdim MB:
Cell signaling pathways in the neuroprotective actions of the green
tea polyphenol (-)-epigallocatechin-3-gallate: Implications for
neurodegenerative diseases. J Neurochem. 88:1555–1569. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weinreb O, Amit T, Mandel S and Youdim MB:
Neuroprotective molecular mechanisms of
(-)-epigallocatechin-3-gallate: A reflective outcome of its
antioxidant, iron chelating and neuritogenic properties. Genes
Nutr. 4:283–296. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ye Q, Ye L, Xu X, Huang B, Zhang X, Zhu Y
and Chen X: Epigallocatechin-3-gallate suppresses
1-methyl-4-phenyl-pyridine-induced oxidative stress in PC12 cells
via the SIRT1/PGC-1α signaling pathway. BMC Complement Altern Med.
12:822012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gundimeda U, McNeill TH, Elhiani AA,
Schiffman JE, Hinton DR and Gopalakrishna R: Green tea polyphenols
precondition against cell death induced by oxygen-glucose
deprivation via stimulation of laminin receptor, generation of
reactive oxygen species, and activation of protein kinase Cε. J
Biol Chem. 287:34694–34708. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ortiz-López L, Márquez-Valadez B,
Gómez-Sánchez A, Silva-Lucero MD, Torres-Pérez M,
Téllez-Ballesteros RI, Ichwan M, Meraz-Ríos MA, Kempermann G and
Ramírez-Rodríguez GB: Green tea compound
epigallo-catechin-3-gallate (EGCG) increases neuronal survival in
adult hippocampal neurogenesis in vivo and in vitro. Neuroscience.
322:208–220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Anacker C, Cattaneo A, Luoni A, Musaelyan
K, Zunszain PA, Milanesi E, Rybka J, Berry A, Cirulli F, Thuret S,
et al: Glucocorticoid-related molecular signaling pathways
regulating hippocampal neurogenesis. Neuropsychopharmacology.
38:872–883. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Heine VM, Griveau A, Chapin C, Ballard PL,
Chen JK and Rowitch DH: A small-molecule smoothened agonist
prevents glucocorticoid-induced neonatal cerebellar injury. Sci
Transl Med. 3:105ra1042011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Heine VM and Rowitch DH: Hedgehog
signaling has a protective effect in glucocorticoid-induced mouse
neonatal brain injury through an 11betaHSD2-dependent mechanism. J
Clin Invest. 119:267–277. 2009.PubMed/NCBI
|
|
31
|
Gulino A, Di Marcotullio L, Ferretti E, De
Smaele E and Screpanti I: Hedgehog signaling pathway in neural
development and disease. Psychoneuroendocrinology. 32 Suppl
1:S52–S56. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Varjosalo M and Taipale J: Hedgehog:
Functions and mechanisms. Genes Dev. 22:2454–2472. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Heine VM, Priller M, Ling J, Rowitch DH
and Schüller U: Dexamethasone destabilizes Nmyc to inhibit the
growth of hedgehog-associated medulloblastoma. Cancer Res.
70:5220–5225. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bambakidis NC, Petrullis M, Kui X,
Rothstein B, Karampelas I, Kuang Y, Selman WR, LaManna JC and
Miller RH: Improvement of neurological recovery and stimulation of
neural progenitor cell proliferation by intrathecal administration
of Sonic hedgehog. J Neurosurg. 116:1114–1120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Y, Li M, Xu X, Song M, Tao H and Bai
Y: Green tea epigallocatechin-3-gallate (EGCG) promotes neural
progenitor cell proliferation and sonic hedgehog pathway activation
during adult hippocampal neurogenesis. Mol Nutr Food Res.
56:1292–1303. 2012. View Article : Google Scholar : PubMed/NCBI
|