Introduction
Cervical carcinoma represents the fourth most common
cancer type worldwide in women, and the third highest cause of
cancer-associated mortality among females in developing countries
(1). According to statistical data,
there were 527,600 new cervical carcinoma cases and 265,700
associated mortalities worldwide in 2012 (2). Current therapeutic strategies of
cervical carcinoma, including surgery, radiation therapy and
chemotherapy, have a high recurrence rate, limited effectiveness
and various side effects. Thus, it is urgent to explore novel
potential therapies for enhancing the prevention and treatment of
cervical carcinoma.
Recently, numerous scholars focused on exploring
natural compounds for the treatment of cervical cancer, due to
their multiple pharmacological activities and low side effects
(3–5). Ellagic acid, with the chemical formula
4,4′,5,5′,6,6′-hexahydroxydiphenic acid-2,6,2′,6′-dilactone, is
widely found in various fruits and plants, including pomegranates,
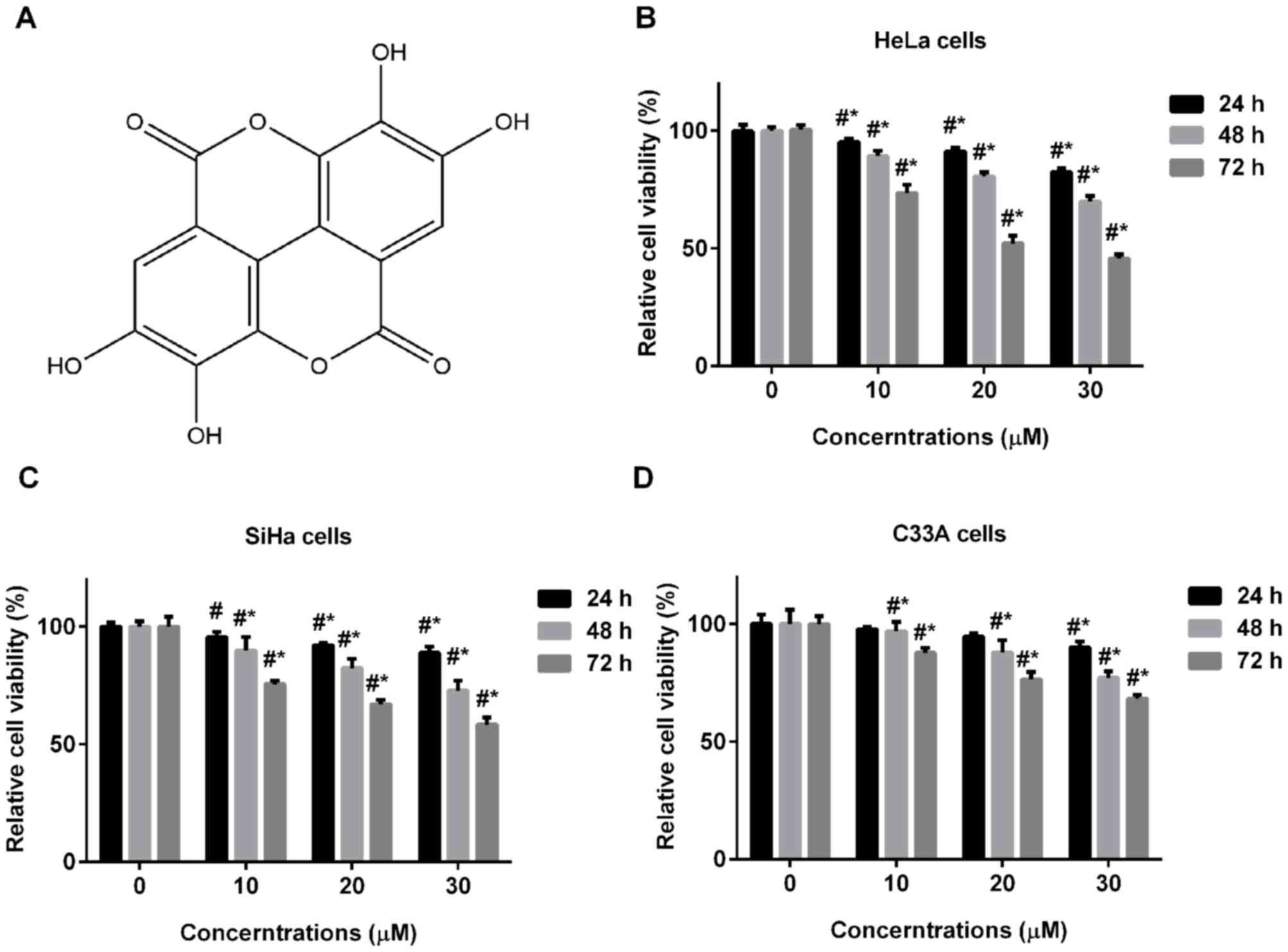
strawberries, raspberries and blackberries (6). The structure of ellagic acid is shown
in Fig. 1A. Ellagic acid has been
reported to possess various effects, including anti-inflammatory
(7), anti-oxidative (8) and antiviral (9) activities. Previous studies have
demonstrated that ellagic acid was able to inhibit tumor growth in
a human pancreatic cancer cell xenografted mice (10), and to induce cell apoptosis in
TSGH8301 human bladder cancer cells via endoplasmic reticulum
stress- and mitochondria-dependent signaling pathways (11). In addition, ellagic acid potentiated
the differentiation of human leukemia cells (12) and presented an anti-angiogenesis
activity in breast cancer (13),
suggesting the strong anticancer activity of ellagic acid.
Furthermore, a previous study indicated that ellagic acid treatment
inhibited cell growth and induced cell apoptosis in human cervical
carcinoma CaSki cells (14).
However, the molecular mechanism underlying the effect of ellagic
acid on cervical carcinoma remains unclear. A recent study
demonstrated that ellagic acid significantly decreased cell
proliferation via its inhibitory effects on signal transducer and
activator of transcription 3 (STAT3) signaling in human prostate
cancer cells (15). Additionally, it
is widely identified that STAT3, which functions as an oncogenic
transcription factor, serves a key role in regulating tumor cell
proliferation, survival, apoptosis and angiogenesis in numerous
types of cancer (16,17). Clinical and pro-clinical studies have
noted that STAT3 is aberrantly expressed in cervical carcinoma
tissues and cell lines (18,19). Increasing evidence also demonstrated
that STAT3serves a key role in the tumor microenvironment (20) and human papillomaviruses infection
(18,21) during the development of cervical
cancer. Based on the aforementioned observations, it is
hypothesized that ellagic acid may inhibit human cervical carcinoma
cells, and the mechanism may be associated with the regulation of
STAT3 signaling.
Therefore, the current study aimed to investigate
the effect and molecular mechanism of ellagic acid on cervical
carcinoma cells. Herein, it was observed that ellagic acid
significantly inhibited the growth of human cervical carcinoma cell
lines, induced cell apoptosis and arrested the cell cycle at G1
phase in human HeLa cells, possibly by regulating the activation of
STAT3 signaling.
Materials and methods
Chemicals and antibodies
Dulbecco's modified Eagle's medium (DMEM), fetal
bovine serum (FBS), penicillin and streptomycin were purchased from
Thermo Fisher Scientific, Inc. (Waltham, MA, USA). Ellagic acid,
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and propidium iodide (PI) were purchased from Sigma-Aldrich (Merck
KGaA, Darmstadt, Germany). Antibodies against phosphorylated-Janus
kinase 2 (p-JAK2; cat. no. 3771), p-STAT3 (Ser727; cat. no. 94994),
p-STAT3 (Tyr705; cat. no. 9145), CyclinD1 (cat. no. 2922), B-cell
lymphoma-extra large (Bcl-xl; cat. no. 2764) and myeloid cell
leukemia 1 (Mcl-1; cat. no. 5453) were obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA; All 1:1,000). Since Tyr705 is
required for STAT3 activation and the present study involves the
Ser727 signaling pathway, Ser727 and Tyr705 (two sites of STAT3)
were examined. Antibodies against glyceraldehyde-3-phosphate
dehydrogenase (GAPDH; cat. no. sc-365062) and horseradish
peroxidase (HRP)-conjugated anti-rabbit IgG (cat. no. sc-2005) were
purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA;
all 1:5,000). All other chemicals and reagents were of analytical
grade.
Cell culture
Human cervical carcinoma HeLa, SiHa and C33A cells
were obtained from the Shanghai Cell Bank (Shanghai, China). HeLa,
SiHa and C33A cells were cultured in DMEM supplemented with 10%
FBS, 100 U/ml penicillin and 100 µg/ml streptomycin at 37°C under a
humidified 5% CO2 atmosphere.
Cell viability assay
MTT assay was performed to investigate the cell
proliferation. Briefly, cells were seeded in 96-well plates at a
density of 2×103cells per well and cultured overnight at
37°C. The control group was treated with 0.1% (v/v) dimethyl
sulfoxide (DMSO), serving as the vehicle. The cells were then
incubated at 37°C with different concentrations (0, 10, 20 and 30
µM) of ellagic acid at different time points (24, 48 and 72 h).
Subsequently, 20 µl MTT (5 mg/ml) were added to each well and
incubated for an additional 3 h at 37°C. Following lysis with 150
µl DMSO, the absorbance was detected using a microplate reader
(Thermo Fisher Scientific, Inc.).
Cell apoptosis detection
HeLa cells were treated with different
concentrations of ellagic acid for 72 h. Next, cell apoptosis was
evaluated with an Annexin V-FITC/PI Apoptosis Detection kit (BD
Pharmingen, San Diego, CA, USA) according to the manufacturer's
instructions. The apoptosis ratio was then measured using a Beckman
Coulter CyAn ADP Flow Cytometer (Becton Dickinson, San Jose, CA,
USA).
Cell cycle progression
examination
HeLa cells were treated with different
concentrations (0, 10, 20 and 30 uM) of ellagic acid for 72 h and
cell apoptosis was evaluated with a Cell Cycle Analysis kit (BD
Pharmingen) following the instructions provided by the
manufacturer. Briefly, subsequent to treating with various
concentrations of ellagic acid for 48 h, HeLa cells were
trypsinized and centrifuged at a speed of 1,000 × g for 10 min at
room temperature. The pellet was washed three times with
phosphate-buffered saline and fixed with 70% alcohol. Subsequently,
the nuclei were stained with PI and the DNA content was measured by
flow cytometry (Becton Dickinson) in accordance with a previous
study (22).
Western blot analysis
Following incubation with various concentrations of
ellagic acid for 72 h, the total protein was extracted from HeLa
cells and the protein concentrations were determined using a is
Pierce™ BCA Protein assay kit (cat. no. 23225; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Equal amounts of protein
(20 µg) were separated by 10% sodium dodecyl sulfate-polyacrylamide
gel electrophoresis and then transferred to polyvinylidene
difluoride membranes. Blocking was then performed by overnight
incubation in Tris-buffered saline/Tween 20 (TBST) containing 5%
non-fat dried milk. The membranes were then incubated with the
primary antibodies against p-JAK2, p-STAT3 (Ser727), p-STAT3
(Tyr705), CyclinD1, Bcl-xl, Mcl-1 and GAPDH overnight at 4°C.
Subsequent to washing with TBST, the blots were incubated with the
aforementioned secondary antibodies coupled to HRP for 1 h at 37°C.
The signal was visualized with an enhanced chemiluminescence kit
(EMD Millipore, Billerica, MA, USA) and GAPDH was used as an
internal loading control. The gray value of the target protein and
GAPDH were analyzed using Image J software (version 4.0; National
Institutes of Health, Bethesda, MD, USA).
Statistical analysis
The data are expressed as the mean ± standard
deviation. All experiments were performed three times and the data
were analyzed by the GraphPad Prism 5 (GraphPad Software, Inc., La
Jolla, CA, USA). Statistical analysis between different groups was
performed using the one-way analysis of variance (ANOVA) followed
by a post-hoc Bonferroni test. A two-way ANOVA was used in the cell
proliferation (MTT) assay. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of ellagic acid on cell
proliferation of human cervical carcinoma cells
To investigate the effect of ellagic acid on cell
proliferation of human cervical carcinoma cells, HeLa, SiHa and
C33A cells were treated with various concentrations of ellagic acid
for 24, 48 or 72 h. As ellagic acid treatment time and
concentration increased, the cell viability of the three cervical
carcinoma cell lines, including HeLa (Fig. 1B), SiHa (Fig. 1C) and C33A cells (Fig. 1D) significantly decreased.
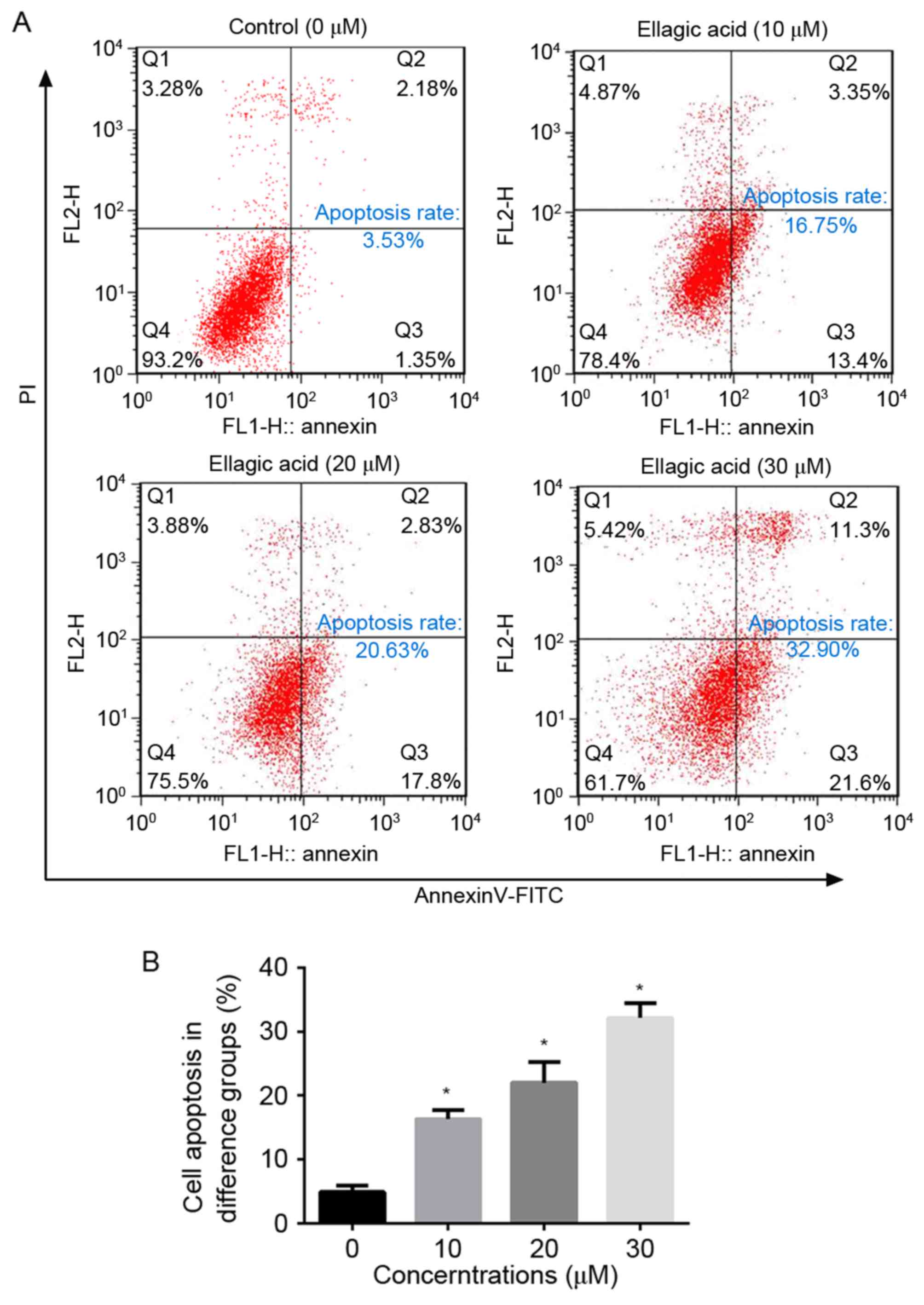
Ellagic acid stimulates cell apoptosis
in HeLa cells
To evaluate the effect of ellagic acid on cell
apoptosis of cervical carcinoma cells, HeLa cells were treated with
various concentrations of ellagic acid and examined by flow
cytometry. As illustrated in Fig. 2,
ellagic acid dosage-dependently increased the number of apoptotic
HeLa cells. In comparison with the NC group (4.90±0.98%), the
apoptosis rates upon treatment with 10 (16.3±1.38%), 20
(22.0±3.24%) and 30 (32.1±2.27%) µM ellagic acid were significantly
increased (P<0.05). These observations suggest that ellagic acid
induces cell apoptosis, which may contribute to the anticancer
effect of ellagic acid in HeLa cells.
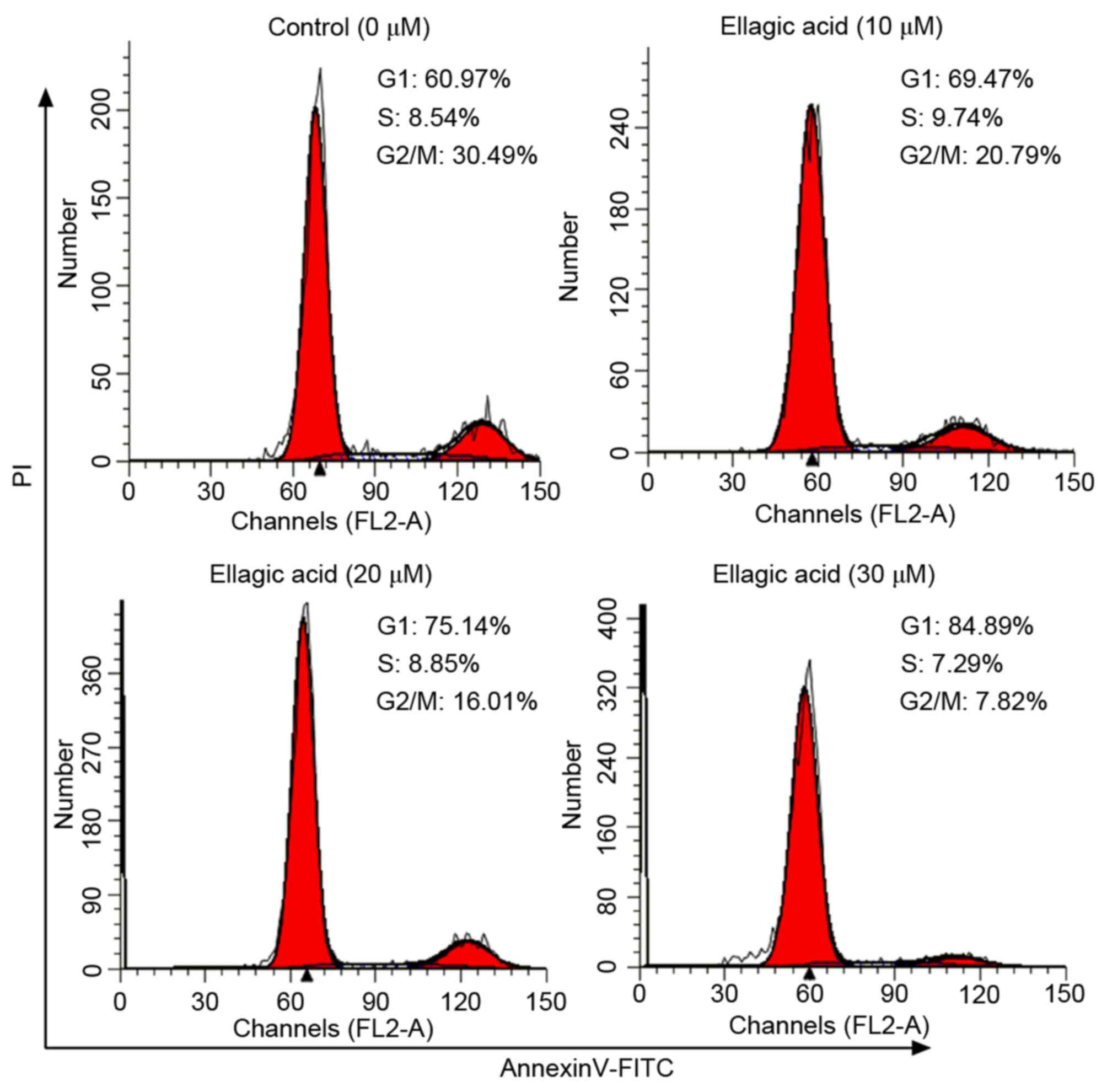
Effect of ellagic acid on cell cycle
distribution of HeLa cells
The present study subsequently investigated whether
ellagic acid was able to affect the cell cycle distribution of HeLa
cells by flow cytometry with PI staining, and the results are shown
in Fig. 3 and Table I. Ellagic acid at the three
investigated doses induced G1 cell cycle arrest in HeLa cells, as
evidenced by the significantly increased proportion of cells at the
G1 phase (69.9±3.55, 75.7±3.27 and 84.7±3.01% for 10, 20 and 30 µM
ellagic acid, respectively; all P<0.05) in HeLa cells compared
with the control group (60.3±4.21%). The results demonstrated that
the proportion of cells at the G2 phase were significantly
decreased in HeLa cells compared with the control group (20.4±2.94,
15.6±2.87 and 9.38±1.37 for 10, 20 and 30 µM ellagic acid,
respectively; all P<0.05).
 | Table I.Proportion of HeLa cells in G1, S and
G2 phases of the cell cycle following ellagic acid treatment. |
Table I.
Proportion of HeLa cells in G1, S and
G2 phases of the cell cycle following ellagic acid treatment.
|
|
| Ellagic acid dose
µM |
|---|
|
|
|
|
|---|
| Cell cycle phase | Control | 10 | 20 | 30 |
|---|
| G1 | 60.3±4.21 |
69.9±3.55a |
75.7±3.27a |
84.7±3.01a |
| S | 8.44±0.39 | 9.65±0.61 | 8.71±0.41 | 7.56±0.37 |
| G2 | 31.2±3.82 |
20.4±2.94a |
15.6±2.87a |
9.38±1.37a |
Effect of ellagic acid on STAT3
signaling in HeLa cells
Given that STAT3 signaling serves a pivotal role in
tumor cell proliferation, survival and apoptosis, the current study
assessed the effect of ellagic acid on the expression levels of
STAT3 and associated proteins by western blot analysis. As shown in
Fig. 4, compared with the control.
the cells treated with ellagic acid presented significant decreased
levels ofp-JAK2 (10 µM, 0.59±0.066; 20 µM, 0.44±0.089; 30 µM,
0.27±0.088), p-STAT3 (Ser727) (10 µM, 0.69±0.085; 20 µM,
0.49±0.097; 30 µM, 0.39±0.086) and p-STAT3 (Tyr705)(10 µM,
0.77±0.092; 20 µM, 0.58±0.104; and 30 µM, 0.33±0.116), when
compared with the control group levels (1.00±0.186, 1.00±0.129 and
1.00±0.141, respectively; all P<0.05). This indicates that
ellagic acid treatment may suppress the activation of STAT3
signaling in HeLa cells. Furthermore, the protein expression levels
of downstream genes were evidently downregulated by the
administration of ellagic acid, including the levels of CyclinD1
(10 µM, 0.81±0.107; 20 µM, 0.55±0.073; 30 µM, 0.34±0.073), Bcl-xl
(10 µM, 0.73±0.060; 20 µM, 0.58±0.049; 30 µM, 0.31±0.079) and Mcl-1
(10 µM, 0.76±0.057; 20 µM, 0.54±0.070; and 30 µM, 0.31±0.076),
compared with the control group levels (1.00±0.107, 1.00±0.155 and
1.00±0.156; all P<0.05). These results suggest that ellagic acid
induces cell cycle arrest at the G1 phase and that cell apoptosis
may be induced via the suppression of STAT3 signaling.
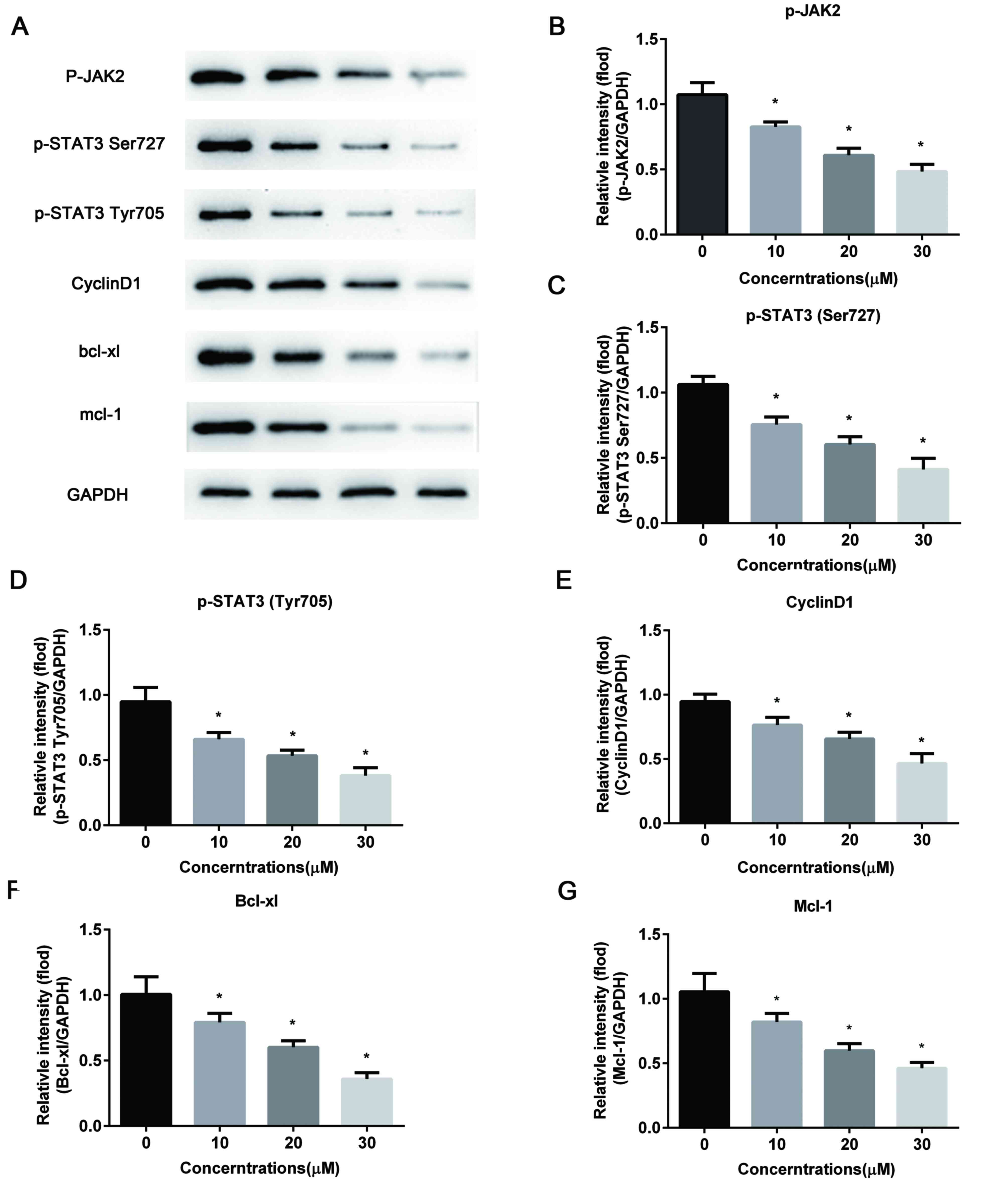 | Figure 4.Ellagic acid suppresses STAT3
signaling pathway in HeLa cells. (A) Representative western blot
bands of the proteins associated with STAT3 signaling. The protein
expression levels of (B) p-JAK2, (C) p-STAT3 (Ser727), (D) p-STAT3
(Tyr705), (E) CyclinD1, (F) Bcl-xl and (G) Mcl-1 were detected by
western blot analysis and normalized to GAPDH. All data are
expressed as the mean ± standard deviation.*P<0.05 vs. the
control group. STAT3, signal transducer and activator of
transcription 3; JAK2, Janus kinase 2; p-, phosphorylated; Bcl-xl,
B-cell lymphoma-extra large; Mcl-1, myeloid cell leukemia 1; GAPDH,
glyceraldehyde-3-phosphate dehydrogenase. |
Discussion
The current study provided insights into the effect
of ellagic acid on the cell proliferation, cell cycle progression
and apoptosis of HeLa cervical carcinoma cells. The most notable
findings of the present study were that ellagic acid arrested the
cell cycle at the G1 phase, induced cell apoptosis, suppressed the
phosphorylationof JAK2 and STAT3, and modulated the expression of
associated proteins. To the best of our knowledge, the current
study is the first to demonstrate that ellagic acid exerts an
anticancer activity in cervical carcinoma HeLa cells possibly by
inducing cell apoptosis and arresting the cell cycle via the
regulation of STAT3 signaling.
Ellagic acid, a plant polyphenolic compound, has
been demonstrated to exert an antitumor activity in several types
of cancer (23–25). The MTT assay conducted in the present
study further confirmed the anticancer property of ellagic acid in
human cervical carcinoma cell lines. It was demonstrated that
ellagic acid reduced the proliferation of the human cervical
carcinoma HeLa, SiHa and C33A cell lines in a dosage- and
time-dependent manner. Another study has previously reported that
ellagic acid dose-dependently suppressed HeLa cell growth (26), which is in agreement with the current
observations. Of note, the present study results revealed that the
inhibitory effects on HeLa cells were more significant compared
with those on SiHa and C33A cells, thus the HeLa cell line was used
for subsequent experiments to explore the molecular mechanism
underlying the effects of ellagic acid.
The mechanism of the anticancer effect of ellagic
acid is widely considered to involve the regulation of the cell
cycle and apoptosis. For instance, ellagic acid inhibited the
growth of MCF-7 breast cancer cells via inducing G0/G1 arrest
(27), as well as suppressed cell
growth by arresting the cell cycle in the G1 phase and inducing
cell apoptosis in ovarian carcinoma cells (28). To further examine the exact mechanism
of ellagic acid-induced inhibition of HeLa proliferation, the
effects of ellagic acid on HeLa cell cycle and apoptosis were
determined by flow cytometry. According to these experiments, the
present study observed that ellagic acid treatment markedly
arrested the cell cycle in the G1 phase and stimulated cell
apoptosis in HeLa cells at a dosage-dependent manner. Furthermore,
STAT3 is generally identified as an oncogenic transcription factor,
which is involved in the regulation of tumor cell proliferation,
survival, apoptosis and angiogenesis in various types of cancer
(16,17). In the current study, it was revealed
that the phosphorylation of JAK2 and STAT3 were significantly
inhibited by treatment with ellagic acid, implying the inhibitory
effect of ellagic acid on STAT3 signaling activation. Furthermore,
STAT3 has been demonstrated to regulate tumorigenic proteins,
including CyclinD1, Bcl-xl and Mcl-1 (29,30).
CyclinD1, the major regulator of cyclin-dependent kinase (CDK) 4 or
CDK6 activation, drives the progression from G1 to S phase
(31). Notably, the results of the
current study demonstrated that ellagic acid evidently
downregulated the expression of CyclinD1, which is consistent with
the results of the cell cycle assay. Bcl-xl and Mcl-1, two
anti-apoptotic proteins involved in the apoptosis and survival of
various cancer types, were also significantly decreased by the
treatment with ellagic acid in HeLa cells. Collectively, the
aforementioned observations indicate that ellagic acid induces cell
cycles arrest in the G1 phase and cell apoptosis possibly via the
suppression of STAT3 signaling. However, further experimental
studies are required to validate these findings in vivo and
to clarify the precise mechanism by which ellagic acid regulates
STAT3 signaling in cervical carcinoma cells.
In conclusion, the data of the present study
indicated that treatment with ellagic acid markedly inhibited cell
proliferation, and induced G1 arrest and apoptosis by suppressing
the activation of STAT3 signaling in cervical carcinoma HeLa cells.
Thus, these findings suggest the potential application of ellagic
acid for the management of patients with cervical carcinoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Scientific
Research Project of Hei Long-jiang Provincial Health Bureau of
China (grant no. 2016-105) and the Administration of Traditional
Chinese Medicine of Heilongjiang Province, China (grant no.
ZHY16-110).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL wrote the manuscript and analyzed the data; CN
and ST analyzed the data and revised the manuscript; JC analyzed
the data; RM and YG collected the data; and GL designed the
study.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA:
a cancer journal for clinicians. 65:87–108. 2015.PubMed/NCBI
|
|
3
|
Liu J, Sun Y, Zhang H, Ji D, Wu F, Tian H,
Liu K, Zhang Y, Wu B and Zhang G: Theanine from tea and its
semi-synthetic derivative TBrC suppress human cervical cancer
growth and migration by inhibiting EGFR/Met-Akt/NF-κB signaling.
Eur J Pharmacol. 791:297–307. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Munagala R, Aqil F, Jeyabalan J and Gupta
RC: Tanshinone IIA inhibits viral oncogene expression leading to
apoptosis and inhibition of cervical cancer. Cancer Lett.
356:536–546. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim B, Kim HS, Jung EJ, Lee JYBKT, Lim JM
and Song YS: Curcumin induces ER stress-mediated apoptosis through
selective generation of reactive oxygen species in cervical cancer
cells. Mol Carcinog. 55:918–928. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Promsong A, Chung WO, Satthakarn S and
Nittayananta W: Ellagic acid modulates the expression of oral
innate immune mediators: Potential role in mucosal protection. J
Oral Pathol Med. 44:214–221. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seo CS, Jeong SJ, Yoo SR, Lee NR and Shin
HK: Quantitative analysis and in vitro anti-inflammatory effects of
gallic acid, ellagic acid, and quercetin from radix sanguisorbae.
Pharmacogn Mag. 12:104–108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baek B, Lee SH, Kim K, Lim HW and Lim CJ:
Ellagic acid plays a protective role against UV-B-induced oxidative
stress by up-regulating antioxidant components in human dermal
fibroblasts. Korean J Physiol Pharmacol. 20:269–277. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park SW, Kwon MJ, Yoo JY, Choi HJ and Ahn
YJ: Antiviral activity and possible mode of action of ellagic acid
identified in Lagerstroemia speciosa leaves toward human
rhinoviruses. BMC Complement Altern Med. 14:1712014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao M, Tang S-N, Marsh JL, Shankar S and
Srivastava RK: Ellagic acid inhibits human pancreatic cancer growth
in Balb c nude mice. Cancer Lett. 337:210–217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ho CC, Huang AC, Yu CS, Lien JC, Wu SH,
Huang YP, Huang HY, Kuo JH, Liao WY, Yang JS, et al: Ellagic acid
induces apoptosis in TSGH8301 human bladder cancer cells through
the endoplasmic reticulum stress- and mitochondria-dependent
signaling pathways. Environ Toxicol. 29:1262–1274. 2014.PubMed/NCBI
|
|
12
|
Hagiwara Y, Kasukabe T, Kaneko Y, Niitsu N
and Okabe-Kado J: Ellagic acid, a natural polyphenolic compound,
induces apoptosis and potentiates retinoic acid-induced
differentiation of human leukemia HL-60 cells. Int J Hematol.
92:136–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang N, Wang ZY, Mo SL, Loo TY, Wang DM,
Luo HB, Yang DP, Chen YL, Shen JG and Chen JP: Ellagic acid, a
phenolic compound, exerts anti-angiogenesis effects via VEGFR-2
signaling pathway in breast cancer. Breast Cancer Res Treat.
134:943–955. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Narayanan BA, Geoffroy O, Willingham MC,
Re GG and Nixon DW: p53/p21(WAF1/CIP1) expression and its possible
role in G1 arrest and apoptosis in ellagic acid treated cancer
cells. Cancer Lett. 136:215–221. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eskandari E, Heidarian E, Amini SA and
Saffari-Chaleshtori J: Evaluating the effects of ellagic acid on
pSTAT3, pAKT, and pERK1/2 signaling pathways in prostate cancer PC3
cells. J Cancer Res Ther. 2016.
|
|
16
|
Siveen KS, Sikka S, Surana R, Dai X, Zhang
J, Kumar AP, Tan BK, Sethi G and Bishayee A: Targeting the STAT3
signaling pathway in cancer: Role of synthetic and natural
inhibitors. Biochim Biophys Acta. 1845:136–154. 2014.PubMed/NCBI
|
|
17
|
Yu H, Lee H, Herrmann A, Buettner R and
Jove R: Revisiting STAT3 signalling in cancer: new and unexpected
biological functions. Nat Rev Cancer. 14:736–746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shukla S, Shishodia G, Mahata S, Hedau S,
Pandey A, Bhambhani S, Batra S, Basir SF, Das BC and Bharti AC:
Aberrant expression and constitutive activation of STAT3 in
cervical carcinogenesis: Implications in high-risk human
papillomavirus infection. Mol Cancer. 9:2822010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen CL, Hsieh FC, Lieblein J, Brown J,
Chan C, Wallace J, Cheng G, Hall B and Lin J: Stat3 activation in
human endometrial and cervical cancers. Brit J Cancer. 96:591–599.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ren C, Cheng X, Lu B and Yang G:
Activation of interleukin-6/signal transducer and activator of
transcription 3 by human papillomavirus early proteins 6 induces
fibroblast senescence to promote cervical tumourigenesis through
autocrine and paracrine pathways in tumour microenvironment. Eur J
Cancer. 49:3889–3899. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sobti R, Singh N, Hussain S, Suri V,
Bharti A and Das B: Overexpression of STAT3 in HPV-mediated
cervical cancer in a north Indian population. Mol Cell Biochem.
330:193–199. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zellner A, Meixensberger J, Roggendorf W,
Janka M, Hoehn H and Roosen K: DNA ploidy and cell-cycle analysis
in intracranial meningiomas and hemangiopericytomas: a study with
high-resolution DNA flow cytometry. Int J Cancer. 79:116–20. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Engelke LH, Hamacher A, Proksch P and
Kassack MU: Ellagic acid and resveratrol prevent the development of
cisplatin resistance in the epithelial ovarian cancer cell line
A2780. J Cancer. 7:353–363. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Edderkaoui M, Odinokova I, Ohno I,
Gukovsky I, Go VL, Pandol SJ and Gukovskaya AS: Ellagic acid
induces apoptosis through inhibition of nuclear factor kappa B in
pancreatic cancer cells. World J Gastroenterol. 14:3672–3680. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Salimi A, Roudkenar MH, Sadeghi L, Mohseni
A, Seydi E, Pirahmadi N and Pourahmad J: Ellagic acid, a
polyphenolic compound, selectively induces ROS-mediated apoptosis
in cancerous B-lymphocytes of CLL patients by directly targeting
mitochondria. Redox Biol. 6:461–471. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kumar D, Basu S, Parija L, Rout D, Manna
S, Dandapat J and Debata PR: Curcumin and Ellagic acid
synergistically induce ROS generation, DNA damage, p53 accumulation
and apoptosis in HeLa cervical carcinoma cells. Biomed
Pharmacother. 81:31–37. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen HS, Bai MH, Zhang T, Li GD and Liu M:
Ellagic acid induces cell cycle arrest and apoptosis through
TGF-β/Smad3 signaling pathway in human breast cancer MCF-7 cells.
Int J Oncol. 46:1730–1738. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chung YC, Lu LC, Tsai MH, Chen YJ, Chen
YY, Yao SP and Hsu CP: The inhibitory effect of ellagic Acid on
cell growth of ovarian carcinoma cells. Evid Based Complement
Alternat Med. 2013:3067052013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu FT, Jia L, Wang P, Wang H, Farren TW
and Agrawal SG: STAT3 and NF-kappaB cooperatively control in vitro
spontaneous apoptosis and poor chemo-responsiveness in patients
with chronic lymphocytic leukemia. Oncotarget. 7:32031–32045.
2016.PubMed/NCBI
|
|
30
|
Li S, Priceman SJ, Xin H, Zhang W, Deng J,
Liu Y, Huang J, Zhu W, Chen M, Hu W, et al: Icaritin inhibits
JAK/STAT3 signaling and growth of renal cell carcinoma. PLoS One.
8:e816572013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang K and Kumar R: Interferon-alpha
inhibits cyclin E- and cyclin D1-dependent CDK-2 kinase activity
associated with RB protein and E2F in Daudi cells. Biochem Biophys
Res Commun. 200:522–528. 1994. View Article : Google Scholar : PubMed/NCBI
|