Introduction
In type 2 diabetes, insulin resistance triggers the
compensation response in β cells, including increased biosynthesis
and secretion of insulin as well as proliferation of β cells
(1). In eukaryotic cells, augmented
protein folding demand perturb ER homeostasis and lead to a
condition defined as endoplasmic reticulum (ER) stress (2). Likewise, increased demand for insulin
secretion brings about a state of metabolic ER stress in β cells,
and β cells are considered to be very sensitive to ER stress, which
contributes to loss of β-cells that underlies the pathogenesis of
type 2 diabetes (3).
The unfolded protein response (UPR), identified as
expanding the protein-folding capacity of the ER, plays a pivotal
role in the control of cell fate for survival under ER stress
(4). If the UPR fails to restore
homeostasis under excessive ER stress, cells will undergo
apoptosis. Three mammalian ER-resident transmembrane proteins,
inositol-requiring enzyme 1 (IRE1), PKR-like endoplasmic reticulum
kinase (PERK), and activating transcription factor 6 (ATF6),
mediate the canonical signaling branches of the UPR that
orchestrate the adaptive response to resolve ER stress (5). IRE1α is the most highly conserved ER
stress sensor which processes dual Ser/Thr kinase and
endoribonuclease (RNase) activities in its cytoplasmic portion
(6,7). In response to ER stress, IRE1α is
activated through oligomerization and autophosphorylation (8,9). In
mammals, activation of IRE1α's RNase activity leads to removal of a
26-nucleotide intron within the mRNA encoding the downstream
transcription factor X-box binding protein 1 (XBP1). This
non-conventional splicing event generates the spliced active form
of XBP1 (XBP1s), which drives a major transcriptional program of
the UPR (10–12). Proper activation of the IRE1a elicits
the cytoprotective actions of the UPR and is essential to maintain
cellular homeostasis and survival in response to ER stress
(13). Notably, the IRE1α-Xbp1
pathway has been implicated in the homeostatic regulation of
pancreatic islet β cells (14).
GLP-1 and its analogy exendin-4 (EX-4), has been
shown to promote β cell replication and prevent β cell exhaustion
under the diabetic conditions (15),
and this effect was explained partly through the inhibition of ER
stress (16). Therefore, in the
present study, we tested the hypothesis that EX-4 could regulate β
cell mass through IRE1α-Xbp1 signaling pathway. Our results
demonstrate that the induction of IRE1α phosphorylation and the
resultant splicing of Xbp1 mRNA evoked by EX-4 treatment could
promote rapid phosphorylation of protein kinase B (Akt) and nuclear
exclusion of FoxO1 in INS-1 cells, which improve cells survival
under lipotoxic-induced ER stress condition.
Materials and methods
Cell culture and treatments
INS-1 832/13 (Cell Culture Centre, CAMS, Beijing,
China) cells were maintained in RPMI-1640 containing 11.1 mM
D-glucose, 10% fetal calf serum, penicillin (100 U/ml) and
streptomycin (100 mg/ml), 10 mM Hepes, 2 mM L-glutamine, 1 mM
sodium pyruvate, and 0.05 mM 2-bmercaptoethanol. After the cells
reached 70% confluence, the medium was replaced with RPMI-1640
containing BSA-conjugated sodium palmitate (PA; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) at concentrations of 0.5 mM, with
or without EX-4 (50 nM; Sigma-Aldrich, Merck KGaA), or 0.5% BSA as
control for 24 h.
Cell transfection
To knock down IRE1α or RACK1 in INS-1 cells, cells
were transfected with small interfering RNA (siRNA; Genepharma,
Shanghai, China) targeting IRE1α (sequences:
5′-GGAATTACTGGCTTCTCATAG′) or RACK1 (target sequences:
5′-GCTAAAGACCAACCACATTGG-3′) at a concentration of 20 µM. Parallel
cell cultures were transfected with control siRNA containing
scrambled non-targeted sequence (5′-GTTCTCCGAACGTGTCACGTTT-3′) at
the equal concentrations (Genepharma). INS-1 cells were transfected
with the plasmid containing human XBP1 s (Addgene Inc., Cambridge,
MA, USA) for XBP1s overexpression. At 48 h after transfection, the
medium was replaced with regular medium containing PA (0.5 mM),
with or without EX-4 (50 nM), Forskolin (10 µM; Sigma-Aldrich;
Merck KGaA) or H89 (10 µM; Sigma-Aldrich; Merck KGaA) for 24 h,
INS-1 cells precultured for 30 min with PKA inhibitor H89 were
treated with EX-4.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from INS-1 cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). First-strand cDNA was synthesized with moloney murine
leukemia virus (M-MLV) reverse transcriptase (Invitrogen) and
random hexamer primers (Invitrogen; Thermo Fisher Scientific,
Inc.). RT-qPCR was conducted using the SYBR Green PCR system,
following the manufacturer's recommendations (Applied Biosystems;
Thermo Fisher Scientific, Inc.). GAPDH was used as an internal
control for normalization. The oligonucleotide primers used are
shown in Table I.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Primer | Forward sequence
(5′-3′) | Reverse sequence
(5′-3′) |
|---|
| Bim |
AGAGATACGGATCGCACAGG |
GTCTTCCGCCTCTCGGTAAT |
| GAPDH |
AGTTCAACGGCAGTCAAG |
TACTCAGCACCAGCATCACC |
Co-immunoprecipitation (Co-IP)
For coimmunoprecipitation analysis, INS-1 cells were
lysed with the lysis buffer [20 mM tris-HCl (pH 7.5), 100 mM KCl,
0.1% Nonidet P-40, 1 mM EDTA, and 10% glycerol containing 1 mM
phenyl-methyl-sulfonyl-fluoride (PMSF), 1% Protease Inhibitor
Cocktail (Sigma-Aldrich; Merck KGaA), and 1% Phosphatase Inhibitor
Cocktails I/II (Sigma-Aldrich; Merck KGaA) for 0.5 h at 4°C. After
incubation with the desired primary antibody for 18 h at 4°C via
gentle rocking, immune complexes were captured by mixing with a
final concentration of 2.5% protein G Sepharosebeads (Amersham; GE
Healthcare, Chicago, IL, USA) for 2 h at 4°C on a rotator.
Anti-IRE1α antibody was used in the Co-IP assays. Beads were
subsequently washed three times with the washing buffer [20 mM
tris-HCl (pH 7.5), 150 mM KCl, 0.5% Nonidet P-40, 1 mM EDTA, and
10% glycerol supplemented with 1 mM PMSF, 1% Protease Inhibitor
Cocktail, and 1% Phosphotase Inhibitor Cocktails I/II], followed by
SDS-PAGE and immunoblotting analysis after elution by boiling in 2X
SDS loading buffer.
Antibodies and immunoblotting
Antibodies against phospho-Akt (Ser473, no. 9271),
phosphor-eIF2α (Ser 51, no. 3597), phospho-FoxO1 (Thr24, no. 9464),
IRE1α (no. 3294), XBP1s (no. 12782), phosphor-CREB (Ser133, no.
9196), eIF2α (no. 9722), Akt (no. 4691), FoxO1 (no. 2880), CREB
(no. 9197), RACK1 (no. 5432), PKA (no. 4782) were purchased from
Cell Signaling Technologies, Inc. (Danvers, MA, USA). α-tubulin
antibody (T6199) from Sigma-Aldrich and antibody against
phospho-IRE1α (Ser724, ab124945) from Abcam (Cambridge, MA, USA).
Tubulin antibody was diluted 1:10,000 and all other antibodies were
diluted 1:1,000. For immune-blotting, cellular lysates were
prepared by RIPA buffer. Protein extracts were separated by
SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred
onto a polyvinylidene difluoride (PVDF) membrane filter (Merck
KGaA, Darmstadt, Germany). After incubation with desired
antibodies, the blots were developed with Thermo Scientific's
SuperSignal West Pico Chemiluminescent substrate or Millipore's
Immunobilon Western Chemiluminescent HRP substrate.
Cell viability and apoptosis
assay
INS-1 cell viability was measured using WST-8 assay
using Cell Counting Kit-8 (CCK-8; Dojindo, Gaithersburg, MD, USA)
according to manufacturer's instruction. INS-1 β cells were seeded
in 96-well culture plates at a density of 105 /ml. The
next day, the culture medium was replaced with RPMI-1640 containing
BSA-conjugated PA at concentrations of 0.5 mM, with or without
EX-4, or 0.5% BSA as control for 24 h. The CCK-8 assay reagent was
added to the culture medium for the final 3 h. A microplate reader
was used to measure the absorbance at 450 nm.
For apoptosis analysis, cells were washed twice with
1X binding buffer then labelled with Annexin V and propidium iodide
(PI) following the manufacturer's instructions. The Apoptosis
Analysis kit was ordered from Beyotime Institute of Biotechnology
(Haimen, China). Cell apoptosis was analyzed by
fluorescence-activated cell sorting (FACS) using a FACScan flow
cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Statistical analysis
All data are expressed as the mean ± standard error
mean. Student's t-test was used to compare mean values between two
groups using the Graph-Pad Prism 5 program (GraphPad Software,
Inc., La Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
EX-4 inhibits ER stress and stimulates
IRE1α-Xbp1 signaling pathway
Exposure to free fatty acids such as PA has been
shown to cause ER stress (17). In
our study, 0.5 mM PA for 24 h did cause a significant increase in
the protein level of eukaryotic initiation factor 2α (eIF2α), a ER
stress marker, as well as the decrease in the phosphorylation of
Akt (p-Akt) and its physiological target FoxO1 (p-FoxO1) (Fig. 1A), which leads to decreased cell
viability (Fig. 1B) and increased
Bim mRNA expression in INS-1 cells (Fig.
1C). The activation of Bim has been supposed to trigger
cleavage of caspases and is critical for the apoptosis in β cells
(18). In accord with this, PA
induced significant cell apoptosis in INS-1 cells (Fig. 1D). However, EX-4 treatment induced a
significant reverse in the protein level of p-Akt and p-FoxO1,
displaying a protective role on the INS-1 cells, as demonstrated by
the increased cell viability and decreased expression of Bim.
IRE1α-Xbp1 signaling pathway has been implicated in the homeostatic
regulation of pancreatic islet β cells, we then tested whether
EX-4-induced protective roles on INS-1 cells was associated with
this signaling pathway. We found that PA triggers the mild increase
in the phosphorylation at the Ser 724 activation site in IRE1α as
detected by a phosphorylation site-specific antibody, which
triggered the increase of XBP1 s protein levels (Fig. 1A). Interestingly, the protein level
of p-IRE1α was further increased in response to EX-4 treatment
(Fig. 1A), accompanied by the
enhancement in XBP1s, suggesting that EX-4 could stimulate
IRE1α-Xbp1 signaling pathway.
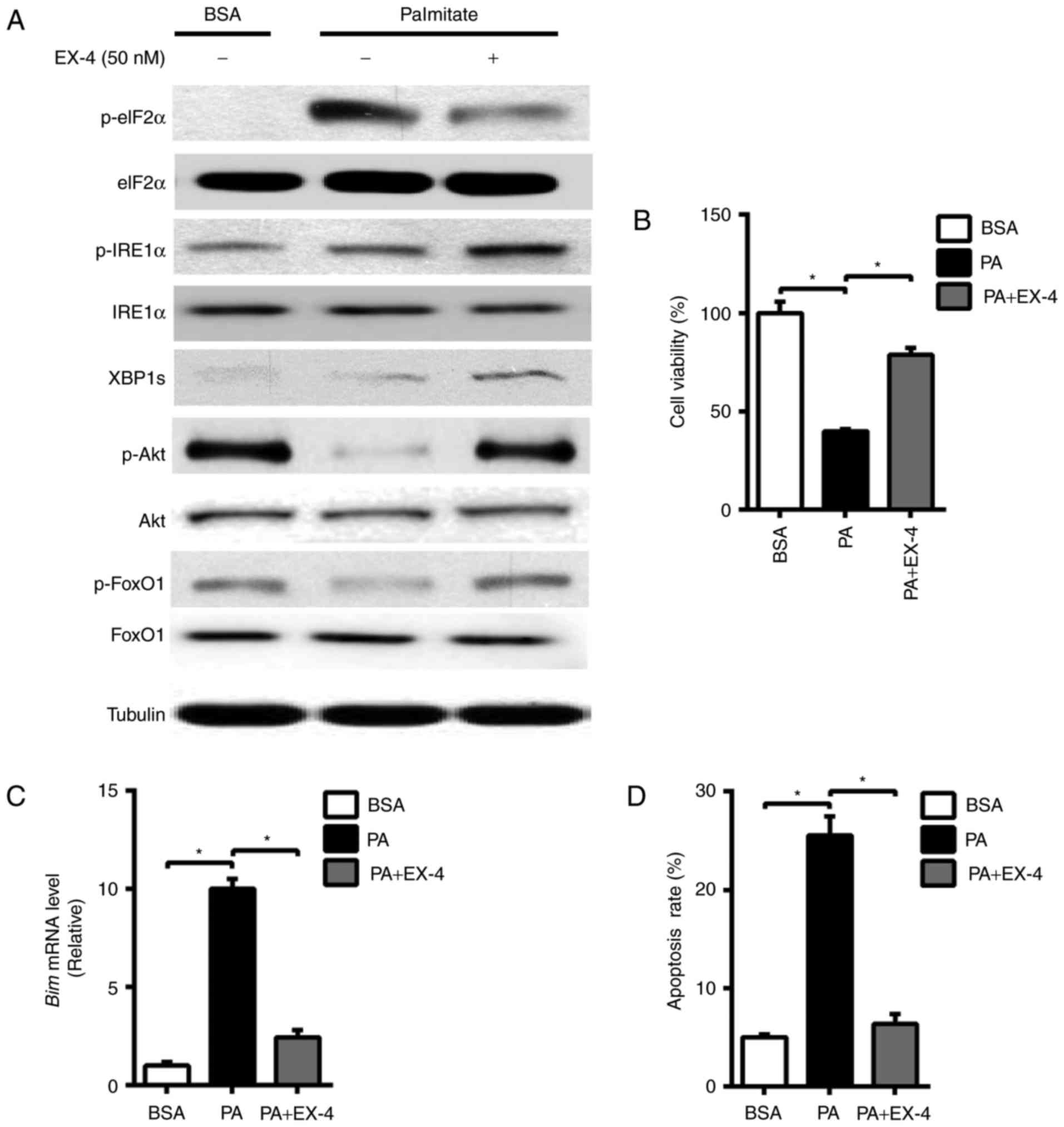 | Figure 1.EX-4 exerts a protective role on
palmitate-induced apoptosis and stimulates the IRE1α-Xbp1 signaling
pathway in INS-1 cells. INS-1 cells were treated with PA (0.5 mM)
and to which EX-4 (50 nM) was then added for 24 h. (A) Western blot
analysis of p-eIF2α, eIF2α, p-Akt, Akt, p-FoxO1, FoxO1, p-IRE1α,
IRE1α, XBP1s and Tubulin. (B) Cell viability. (C) Reverse
transcription-quantitative polymerase chain reaction for Bim mRNA
expression. (D) Cell apoptosis rate was determined by flow
cytometry using Annexin V and propidium iodide. Tubulin was used as
the internal control. Data are presented as the mean ± standard
error mean of three independent experiments. *P<0.05, as
indicated. IRE1α, inositol-requiring enzyme 1α; Xbp1, X-box binding
protein 1; EX-4, exendin-4; p-, phosphorylated-; eIF2α, eukaryotic
initiation factor 2α; Akt, protein kinase B; FoxO1, forkhead box
protein O1; Bim, B-cell lymphoma-2-like protein 11; BSA, bovine
serum albumin; PA, sodium palmitate. |
EX-4-induced inhibition of ER stress
and improvement of p-Akt is mediated by the stimulation of IRE1
α-Xbp1 signaling pathway
To determine whether IRE1α-Xbp1 signaling pathway
acts as a critical component in mediating EX-4's beneficial effects
in PA-induced INS-1 cells, we used siRNA to silence the IRE1α gene.
INS-1 cells were transfected with siRNA specific for IRE1α and the
efficiency of IRE1α knockdown was validated (Fig. 2A). In nontransfected cells treated
with EX-4, a significant increase in p-Akt and a drastic decrease
in p-eIF2α protein levels was observed (Fig. 1A), in contrast, transfection of INS-1
cells with siRNA targeting IRE1α, the expression of p-Akt was only
partially improved by EX-4 treatment under PA-treated conditions
(Fig. 2A). The decreased cell
viability and increased mRNA level of Bim and cell apoptosis were
also not ameliorated (Fig. 2B-D). As
IRE1α associates with various signaling molecules, we then tested
whether EX-4-induced upregulation of p-Akt is through the splicing
of Xbp1 mRNA by IRE1α phosphorylation. In INS-1 cells with
RNAi-mediated knockdown of IRE1α, the decreased protein levels of
p-Akt and cell viability under EX-4 treatment was rescued by XBP1s
overexpression (Fig. 2A and B),
while the mRNA level of Bim and cell apoptosis markedly decreased
(Fig. 2C and D) This data indicates
that the beneficial roles of EX-4 treatment on PA-induced INS-1
cells might be associated with the IRE1α-Xbp1 signaling
pathway.
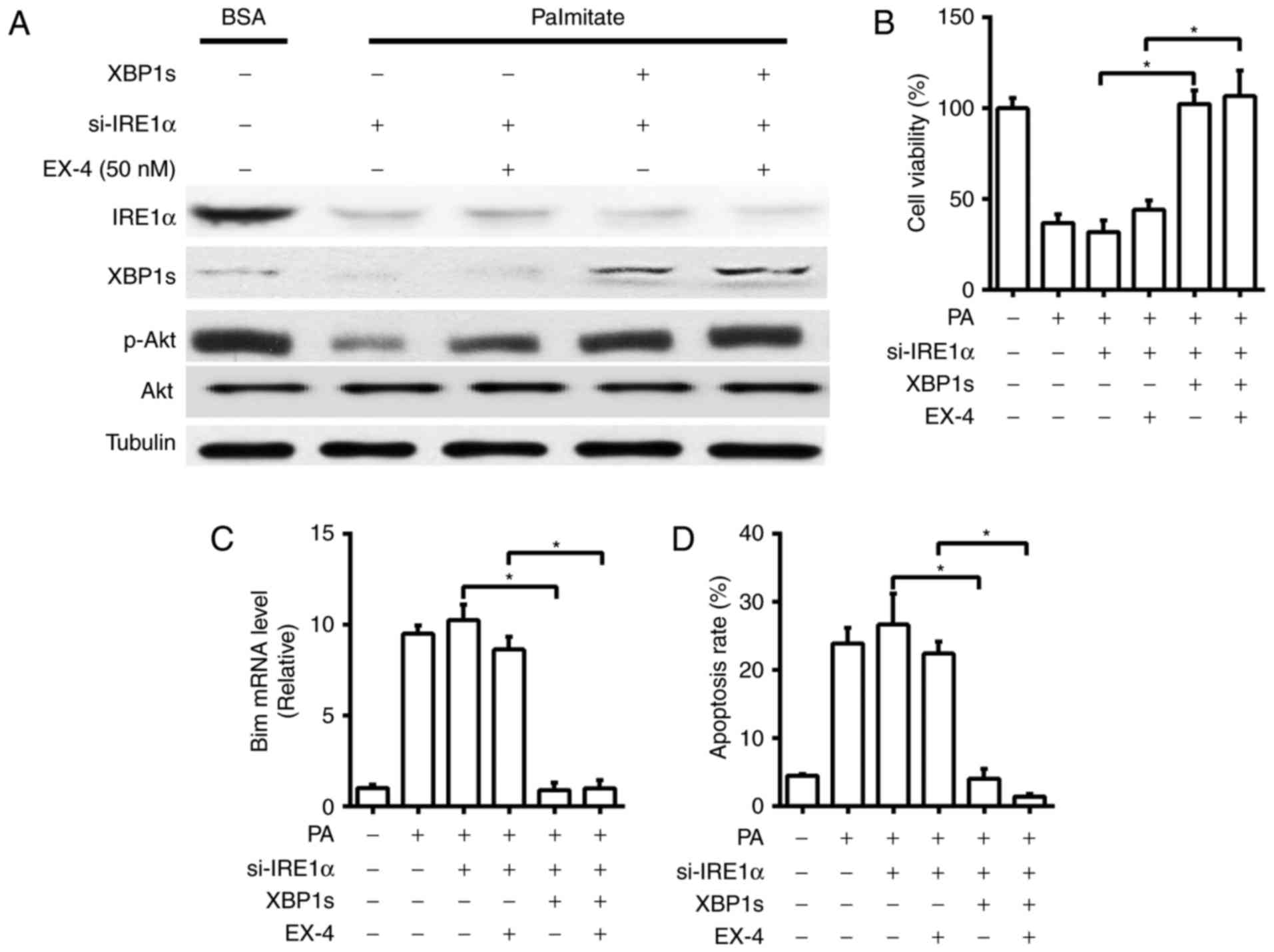 | Figure 2.IRE1α-Xbp1 signaling pathway mediates
the protective role of EX-4 on the palmitate-induced apoptosis in
INS-1 cells. INS-1 cells with RNAi-mediated knockdown of IRE1α,
coupled with or without the overexpression of XBP1s by transfection
with a plasmid containing human XBP1s, were treated with PA (0.5
mM), with or without EX-4 (50 nM), for 24 h. (A) Western blot
analysis of IRE1α, XBP1s, p-Akt, Akt and Tubulin. (B) Cell
viability. (C) Reverse transcription-quantitative polymerase chain
reaction for Bim mRNA expression. (D) Cell apoptosis rate was
determined by flow cytometry using Annexin V and propidium iodide.
Tubulin was used as the internal control. Data are presented as the
mean ± standard error mean of three independent experiments.
*P<0.05, as indicated. IRE1α, inositol-requiring enzyme 1α;
Xbp1, X-box binding protein 1; EX-4, exendin-4; p-,
phosphorylated-; si-, small interfering; Akt, protein kinase B;
Bim, B-cell lymphoma-2-like protein 11; BSA, bovine serum albumin;
PA, sodium palmitate. |
EX-4-induced phosphorylation of IRE1α
is dependent on PKA
Previous research has supported the idea that under
ER stress conditions, IRE1α is activated through dimerization and
transautophosphorylation (8,9). To test the idea that a protein kinase
other than IRE1α itself may link EX-4 with the observed stimulation
of IRE1α phosphorylation, we examined whether protein kinase A
(PKA) is involved in the stimulation of IRE1α phosphorylation by
EX-4. We found that EX-4 could promote the activation of PKA, as
evidenced by the increased expression of cAMP response
element-binding protein (CREB) phosphorylation (p-CREB), as well as
the upregulation of IRE1α phosphorylation (Fig. 3). In accordance with this, forskolin,
a chemical activator of PKA, also triggered the increase of IRE1α
phosphorylation, while the increase in the IRE1α phosphorylation by
EX-4 was suppressed by the inhibition of PKA using H89 (Fig. 3), a pharmacological PKA inhibitor,
implying that EX-4-regulated IRE1α-phosphorylation is mediated
through a PKA-dependent manner.
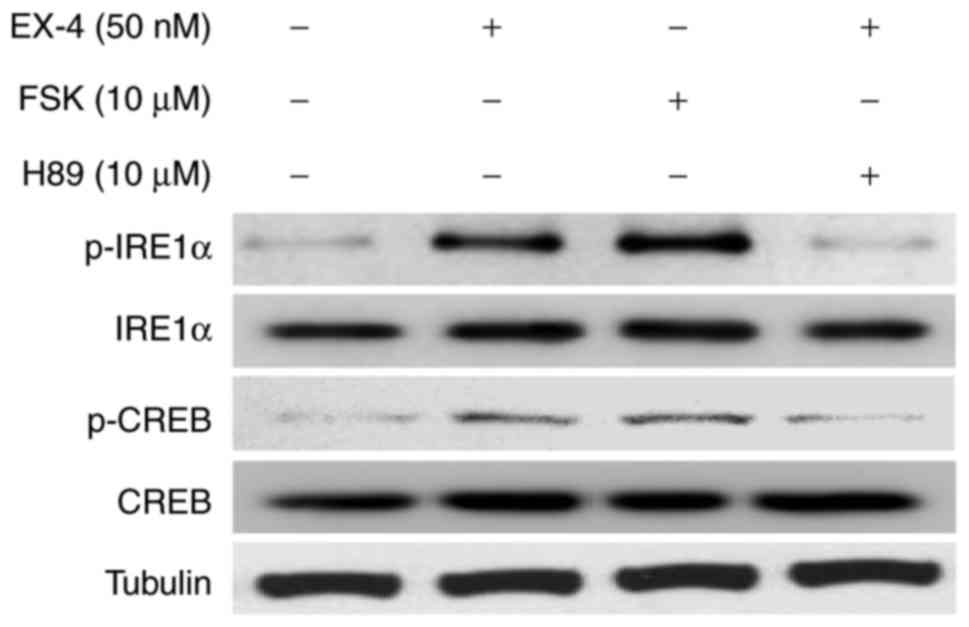 | Figure 3.Protein kinase A is required for
EX-4-induced phosphorylation of IRE1α. INS-1 cells were treated
with Sodium palmitate (0.5 mM), to which FSK (10 µM), EX-4 (50 nM)
or EX-4 with H89 (10 µM), was then added for 24 h. Western blot
analysis was then performed for p-CREB, CREB, p-IRE1α, IRE1α and
Tubulin. Tubulin was used as the internal control. Tubulin was used
as the internal control. IRE1α, inositol-requiring enzyme 1α; EX-4,
exendin-4; FSK, Forskolin; p-, phosphorylated-; CREB, cAMP response
element-binding protein. |
RACK1 is essential for PKA-dependent
IRE1α phosphorylation in response to EX-4
RACK1, binding to membrane receptors and protein
kinases, coordinates the interactions between signaling components
in multiple cellular processes (19). We found that EX-4 treatment induced
the association of IRE1α with PKA and RACK1 by Co-IP analysis
(Fig. 4A). More importantly, INS-1
cells with knockdown of RACK1, although presenting increased p-CREB
by EX-4, showed the absent increased IRE1α phosphorylation compared
to cells transfected with a scrambled negative control siRNA
(Fig. 4B). This confirms a possible
function of RACK1 in the recruitment of PKA to phosphorylated IRE1α
in response to EX-4 treatment.
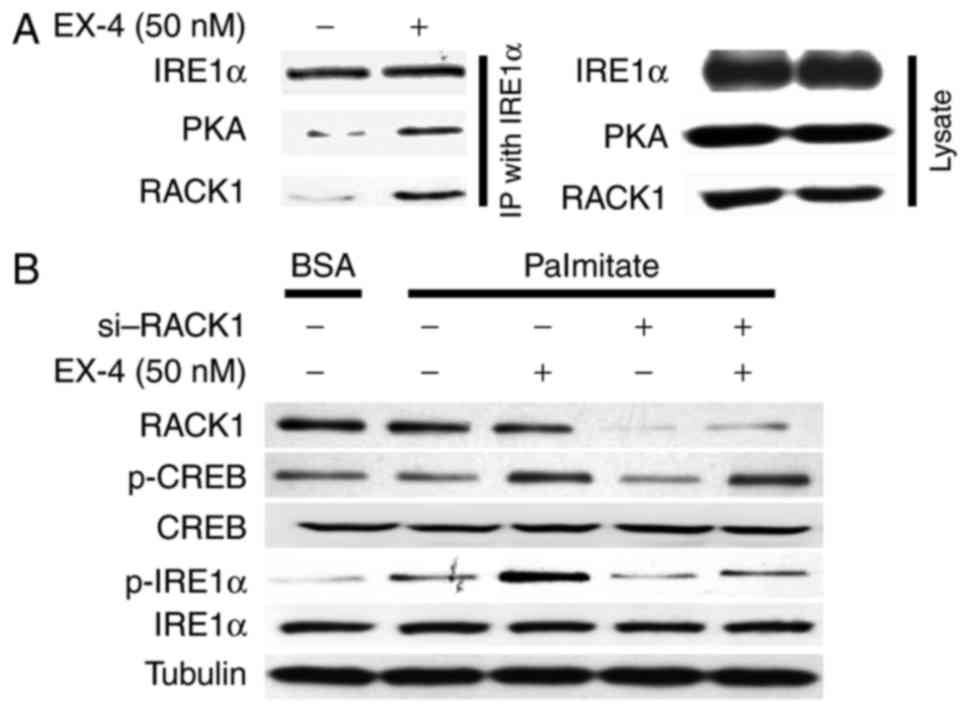 | Figure 4.RACK1 is essential for PKA-dependent
IRE1α phosphorylation in response to EX-4 treatment. (A) INS-1
cells were treated with Sodium palmitate (0.5 mM) and then with or
without EX-4 (50 nM) for 24 h. Immunoprecipitation was performed
with anti-IRE1α, followed by immunoblotting with RACK1, PKA and
IRE1α antibodies. Immunoprecipitation reactions were replicated
three times. (B) INS-1 cells with RNAi-mediated knockdown of RACK1,
were then treated with Sodium palmitate (0.5 mM), with or without
EX-4 (50 nM) for 24 h. Western blot analysis of RACK1, p-CREB,
CREB, p-IRE1α, IRE1α and Tubulin was then performed. Tubulin was
used as the internal control. RACK1, receptor for activated C
kinase 1; PKA, protein kinase A; IRE1α, inositol-requiring enzyme
1α; EX-4, exendin-4; BSA, bovine serum albumin; p-,
phosphorylated-; CREB, cAMP response element-binding protein. |
Discussion
The suppression of ER stress and the activation of
Akt in GLP-1-mediated β cell survival under lipotoxic conditions
has previously been demonstrated (16,20), but
the mechanisms are not well established. The present study provides
evidence that induction of the IRE1α-Xbp1 axis, a signaling branch
of the UPR, by EX-4 treatment contributes to the alleviation of ER
stress and the activation of Akt, which exerts anti-apoptotic roles
on PA-treated INS-1 cells. These novel insights connect the EX-4
and the UPR response in a complex chain of events that inhibits ER
stress and promotes β cell survival.
In mammals, IRE1α is recognized as a signaling
branch to mediate UPR by splicing Xbp1 mRNA to generate an active
form of this transcription factor that induces a major
transcriptional program of the UPR (10,11,14). UPR
can ameliorate ER stress by enhancing the ER's capacity to manage
the workload of protein folding (5).
In an in vitro ‘ER stress’ model produced by prolonged
exposure of PA, EX-4-induced inhibition of ER stress and the
improvement of cell viability is linked to the activation of
IRE1α-Xbp1 signaling pathway, suggesting that IRE1α-Xbp1 may be
involved in mediating the anti-apoptotic role of EX-4 on INS-1
cells by suppressing ER stress.
Activation of Akt results in the phosphorylation of
various downstream protein targets that affect proliferation, cell
cycle entry and intracellular apoptotic pathways (21,22). Akt
signaling has been shown to protect β cells from ER stress-induced
apoptosis and mediate part of the anti-apoptotic effects of GLP-1
agonists (23–25). FoxO1, a downstream target of Akt
pathway, triggers cell death in different types of cells which
involves transactivation of BH3-only molecule Bim (26–29). Bim
is a member of pro-apoptotic BH3-only protein that plays an
important role in mediating β cell apoptosis (18). Phosphorylation of FoxO1 by Akt
results in the inhibition of FoxO1-dependent transcription that
elicits the protective roles of pancreatic β cells against cell
lipoapoptosis (30,31). Our data showed that IRE1α may be
involved in the improvement of the phosphorylation of Akt, as the
deficiency of IRE1α down-regulated its phosphorylation. Our global
analysis of protein expression in IRE1α-deficient cells pointed to
the suppression of ER stress and increase in the Akt signaling as
possible mechanisms for the IRE1α-mediated β cell protection in
response to EX-4 treatment. In agreement with anti-apoptotic role
of IRE1α-Xbp1 signaling pathway, IRE1α has been previously shown to
drive compensatory β cell proliferation during metabolic ER stress
through XBP1s regulation of cell cycle machinery, as demonstrated
by decreased islet mass and hypoinsulinemia in Ire1αf/f:
Cre mice when challenged with HFD feeding (14). Previous research also demonstrated
the role of XBP1s on the protection of β cells against apoptosis
through the induction of Akt (32).
It is worth noting that our data showed that IRE1α may modulate Akt
phosphorylation through an XBP1 dependent manner as well. In fact,
the regulatory role of XBP1s on Akt activity has also been reported
in other cell types (33,34). Besides, an earlier study reported
that in islets of db/db mice, increased activation of xbp1 mRNA was
observed, presumably a compensatory mechanism to inhibit ER stress
and β cell apoptosis induced by obesity (35). Our data raise the possibility that
IRE1α-Xbp1 axis may serve as a component to regulate Akt
phosphorylation in response to EX-4 treatment. Nonetheless,
additional signaling regulators may also be involved in
coordinating the anti-apoptotic actions of the IRE1α-Xbp1 branch.
For instance, PI3K of the insulin/IGF-1 pathway can interact with
XBP1s to modulate its functional behavior, implying that XBP1s may
likely serve as a component of the insulin/IGF-1 signaling cascade
in promoting β cell proliferation (36).
Notably, it was documented that IRE1α could also
regulate insulin biosynthesis. For instance, increased
phosphorylation of IRE1α is coupled to insulin production through
mechanisms that do not involve Xbp1 splicing in response to acute
glucose stimulation, whereas prolonged activation of IRE1α leads to
suppression of insulin production after chronic exposure to high
glucose (2). Interestingly, genetic
deletion of Xbp1 specifically in pancreatic islet β cells of mice
was reported to cause defective proinsulin processing and insulin
secretion, as a result of the feedback hyperactivation of IRE1α
that in turn degrades mRNAs encoding enzymes for proinsulin
processing (37). Based on this
evidence, it's likely that the the role of IRE1α in integrating ER
stress could regulate both the cell mass and the function of β
cells.
The activation of IRE1α is considered to be
activated through autophosphorylation to initiate a key signaling
arm of the mammalian UPR pathways (8). Previous research showed the
anti-apoptotic effects of the GLP-1 analog in primary-cultured
neonatal rat β cells could be reproduced by the activator of PKA
(38), implying that PKA plays an
important role in mediating the anti-apoptotic effect of GLP-1
analog. In our study, the early induction of IRE1α phosphorylation
by PA may constitute an important component of UPR to cope with ER
stress, while the further augment of IRE1α phosphorylation in
response to EX-4 treatment was blunted by the pharmacological PKA
inhibitor H89, suggesting that PKA, other than IRE1α itself, may
link EX-4 with the observed stimulation of IRE1α phosphorylation.
This finding is supported by a recent study showed that hepatic PKA
can directly phosphorylate IRE1α at Ser 724 that modulates the
metabolic activation of IRE1α in the mice with obesity (7). Although a direct activation of Akt by
PKA cannot be completely excluded in the present study, our data
favor the idea that PKA activates Akt through IRE1α-Xbp1 signaling
pathway by EX-4. Furthermore, our study revealed a crucial role of
RACK1, a multifaceted scaffolding protein, in the regulation of
IRE1α phosphorylation by PKA in response to EX-4. RACK1 is a
scaffold protein which contains seven Trp-Asp 40 (WD40) repeats,
which binds to membrane receptors and protein kinases as well as
coordinates the interactions between signaling components in the
cellular processes (19,39,40).
However, the precise pathophysiological effect of RACK1-dependent
regulation of IRE1α by PKA in response to EX-4 remains to be
further investigated.
In conclusion, we show that IRE1α-Xbp1 signaling
pathway contributes to cope with ER stress and activate Akt
signaling, thereby preventing PA-induced β cell apoptosis under
EX-4 treatment conditions. A better understanding of the molecular
components that link the EX-4 and UPR with the anti-apoptotic
control of β cells may shed light on the new mechanisms of the EX-4
treatment for type 2 diabetes.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National 863
Hi-Tech Project (grant no. 2015AA033703) and the National Key
R&D Program of China (grant nos. 2016YFC1100300,
2016YFC1100304, 2017YFC0840100 and 2017YFC0840106).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FW and DJ conceived the initial idea of the work,
supervised the study and wrote the manuscript. DJ contributed to
the design of the study, and performed the experiments and data
analysis. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sachdeva MM and Stoffers DA: Minireview:
Meeting the demand for insulin: Molecular mechanisms of adaptive
postnatal beta-cell mass expansion. Mol Endocrinol. 23:747–758.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lipson KL, Fonseca SG, Ishigaki S, Nguyen
LX, Foss E, Bortell R, Rossini AA and Urano F: Regulation of
insulin biosynthesis in pancreatic beta cells by an endoplasmic
reticulum-resident protein kinase IRE1. Cell Metab. 4:245–254.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wajchenberg BL: Beta-cell failure in
diabetes and preservation by clinical treatment. Endocr Rev.
28:187–218. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hetz C: The unfolded protein response:
Controlling cell fate decisions under ER stress and beyond. Nat Rev
Mol Cell Biol. 13:89–102. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bell GI and Polonsky KS: Diabetes mellitus
and genetically programmed defects in beta-cell function. Nature.
414:788–791. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mao T, Shao M, Qiu Y, Huang J, Zhang Y,
Song B, Wang Q, Jiang L, Liu Y, Han JD, et al: PKA phosphorylation
couples hepatic inositol-requiring enzyme 1alpha to glucagon
signaling in glucose metabolism. Proc Natl Acad Sci USA.
108:15852–15857. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shamu CE and Walter P: Oligomerization and
phosphorylation of the Ire1p kinase during intracellular signaling
from the endoplasmic reticulum to the nucleus. EMBO J.
15:3028–3039. 1996.PubMed/NCBI
|
|
9
|
Welihinda AA and Kaufman RJ: The unfolded
protein response pathway in Saccharomyces cerevisiae.
Oligomerization and trans-phosphorylation of Ire1p (Ern1p) are
required for kinase activation. J Biol Chem. 271:18181–18187. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Calfon M, Zeng H, Urano F, Till JH,
Hubbard SR, Harding HP, Clark SG and Ron D: IRE1 couples
endoplasmic reticulum load to secretory capacity by processing the
XBP-1 mRNA. Nature. 415:92–96. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee K, Tirasophon W, Shen X, Michalak M,
Prywes R, Okada T, Yoshida H, Mori K and Kaufman RJ: IRE1-mediated
unconventional mRNA splicing and S2P-mediated ATF6 cleavage merge
to regulate XBP1 in signaling the unfolded protein response. Genes
Dev. 16:452–466. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshida H, Matsui T, Yamamoto A, Okada T
and Mori K: XBP1 mRNA is induced by ATF6 and spliced by IRE1 in
response to ER stress to produce a highly active transcription
factor. Cell. 107:881–891. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin JH, Li H, Yasumura D, Cohen HR, Zhang
C, Panning B, Shokat KM, Lavail MM and Walter P: IRE1 signaling
affects cell fate during the unfolded protein response. Science.
318:944–949. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu T, Yang L, Yan C, Wang X, Huang P, Zhao
F, Zhao L, Zhang M, Jia W, Wang X and Liu Y: The IRE1α-XBP1 pathway
regulates metabolic stress-induced compensatory proliferation of
pancreatic β-cells. Cell Res. 24:1137–1140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Drucker DJ: Glucagon-like peptide-1 and
the islet beta-cell: Augmentation of cell proliferation and
inhibition of apoptosis. Endocrinology. 144:5145–5148. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsunekawa S, Yamamoto N, Tsukamoto K, Itoh
Y, Kaneko Y, Kimura T, Ariyoshi Y, Miura Y, Oiso Y and Niki I:
Protection of pancreatic beta-cells by exendin-4 may involve the
reduction of endoplasmic reticulum stress; in vivo and in vitro
studies. J Endocrinol. 193:65–74. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Achard CS and Laybutt DR: Lipid-induced
endoplasmic reticulum stress in liver cells results in two distinct
outcomes: Adaptation with enhanced insulin signaling or insulin
resistance. Endocrinology. 153:2164–2177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ren D, Sun J, Mao L, Ye H and Polonsky KS:
BH3-only molecule Bim mediates β-cell death in IRS2 deficiency.
Diabetes. 63:3378–3387. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
McCahill A, Warwicker J, Bolger GB,
Houslay MD and Yarwood SJ: The RACK1 scaffold protein: A dynamic
cog in cell response mechanisms. Mol Pharmacol. 62:1261–1273. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Buteau J, El-Assaad W, Rhodes CJ,
Rosenberg L, Joly E and Prentki M: Glucagon-like peptide-1 prevents
beta cell glucolipotoxicity. Diabetologia. 47:806–815. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Scheid MP and Woodgett JR: PKB/AKT:
Functional insights from genetic models. Nat Rev Mol Cell Biol.
2:760–768. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hajduch E, Litherland GJ and Hundal HS:
Protein kinase B (PKB/Akt)-a key regulator of glucose transport?
FEBS Lett. 492:199–203. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wrede CE, Dickson LM, Lingohr MK, Briaud I
and Rhodes CJ: Fatty acid and phorbol ester-mediated interference
of mitogenic signaling via novel protein kinase C isoforms in
pancreatic beta-cells (INS-1). J Mol Endocrinol. 30:271–286. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Srinivasan S, Ohsugi M, Liu Z, Fatrai S,
Bernal-Mizrachi E and Permutt MA: Endoplasmic reticulum
stress-induced apoptosis is partly mediated by reduced insulin
signaling through phosphatidylinositol 3-kinase/Akt and increased
glycogen synthase kinase-3beta in mouse insulinoma cells. Diabetes.
54:968–975. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cunha DA, Ladrière L, Ortis F,
Igoillo-Esteve M, Gurzov EN, Lupi R, Marchetti P, Eizirik DL and
Cnop M: Glucagon-like peptide-1 agonists protect pancreatic
beta-cells from lipotoxic endoplasmic reticulum stress through
upregulation of BiP and JunB. Diabetes. 58:2851–2862. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gilley J, Coffer PJ and Ham J: FOXO
transcription factors directly activate bim gene expression and
promote apoptosis in sympathetic neurons. J Cell Biol. 162:613–622.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stahl M, Dijkers PF, Kops GJ, Lens SM,
Coffer PJ, Burgering BM and Medema RH: The forkhead transcription
factor FoxO regulates transcription of p27Kip1 and Bim in response
to IL-2. J Immunol. 168:5024–5031. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dijkers PF, Birkenkamp KU, Lam EW, Thomas
NS, Lammers JW, Koenderman L and Coffer PJ: FKHR-L1 can act as a
critical effector of cell death induced by cytokine withdrawal:
Protein kinase B-enhanced cell survival through maintenance of
mitochondrial integrity. J Cell Biol. 156:531–542. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dijkers PF, Medema RH, Lammers JW,
Koenderman L and Coffer PJ: Expression of the pro-apoptotic Bcl-2
family member Bim is regulated by the forkhead transcription factor
FKHR-L1. Curr Biol. 10:1201–1204. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kitamura T, Nakae J, Kitamura Y, Kido Y,
Biggs WH III, Wright CV, White MF, Arden KC and Accili D: The
forkhead transcription factor Foxo1 links insulin signaling to Pdx1
regulation of pancreatic beta cell growth. J Clin Invest.
110:1839–1847. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Martinez SC, Tanabe K, Cras-Méneur C,
Abumrad NA, Bernal-Mizrachi E and Permutt MA: Inhibition of Foxo1
protects pancreatic islet beta-cells against fatty acid and
endoplasmic reticulum stress-induced apoptosis. Diabetes.
57:846–859. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cunha DA, Gurzov EN, Naamane N, Ortis F,
Cardozo AK, Bugliani M, Marchetti P, Eizirik DL and Cnop M: JunB
protects β-cells from lipotoxicity via the XBP1-AKT pathway. Cell
Death Differ. 21:1313–1324. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu MC, Gong HY, Lin GH, Hu SY, Chen MH,
Huang SJ, Liao CF and Wu JL: XBP-1, a key regulator of unfolded
protein response, activates transcription of IGF1 and Akt
phosphorylation in zebrafish embryonic cell line. Biochem Biophys
Res Commun. 359:778–783. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Akiyama M, Liew CW, Lu S, Hu J, Martinez
R, Hambro B, Kennedy RT and Kulkarni RN: X-box binding protein 1 is
essential for insulin regulation of pancreatic α-cell function.
Diabetes. 62:2439–2449. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Laybutt DR, Preston AM, Akerfeldt MC,
Kench JG, Busch AK, Biankin AV and Biden TJ: Endoplasmic reticulum
stress contributes to beta cell apoptosis in type 2 diabetes.
Diabetologia. 50:752–763. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Winnay JN, Boucher J, Mori MA, Ueki K and
Kahn CR: A regulatory subunit of phosphoinositide 3-kinase
increases the nuclear accumulation of X-box-binding protein-1 to
modulate the unfolded protein response. Nat Med. 16:438–445. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee AH, Heidtman K, Hotamisligil GS and
Glimcher LH: Dual and opposing roles of the unfolded protein
response regulated by IRE1alpha and XBP1 in proinsulin processing
and insulin secretion. Proc Natl Acad Sci USA. 108:8885–8890. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bregenholt S, Møldrup A, Blume N, Karlsen
AE, Nissen Friedrichsen B, Tornhave D, Knudsen LB and Petersen JS:
The long-acting glucagon-like peptide-1 analogue, liraglutide,
inhibits beta-cell apoptosis in vitro. Biochem Biophys Res Commun.
330:577–584. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Arimoto K, Fukuda H, Imajoh-Ohmi S, Saito
H and Takekawa M: Formation of stress granules inhibits apoptosis
by suppressing stress-responsive MAPK pathways. Nat Cell Biol.
10:1324–1332. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
López-Bergami P, Habelhah H, Bhoumik A,
Zhang W, Wang LH and Ronai Z: RACK1 mediates activation of JNK by
protein kinase C [corrected]. Mol Cell. 19:309–320. 2005.
View Article : Google Scholar : PubMed/NCBI
|














