Introduction
Podocytes, also known as visceral glomerular
epithelial cells, serve a critical role in maintaining the normal
structure and function of the glomerular filtration barrier
(1). A wide variety of glomerular
diseases can lead to many abnormal structural deformations of
podocytes, including podocyte foot process fusion and detachment
from the glomerular basement membrane (GBM), GBM thickening and
ultimately a reduction in podocytes (2). Manifold stimuli can lead to podocyte
abnormality and apoptosis, including immune-mediated and oxidative
stress, hemodynamic changes, hyperglycemia and hyperlipidemia
(3). To date, it has been
established that glycation end-products and reactive oxygen species
(ROS) induced by hyperglycemia can result in ultrastructural
alterations of the glomerular filtration barrier in diabetic kidney
diseases (2). Tight control of blood
glucose can delay the development of diabetic kidney disease (DKD),
but this alone is not capable of preventing progression of diabetic
nephropathy to end-stage renal damage (ESRD).
Oxidative stress is a particularly important cause
of kidney injury and podocyte apoptosis, activating a series of
phosphorylated kinases. These, in turn, induce expression of
downstream transcription factors and contribute to increased
apoptosis and detachment of podocytes that line the GBM (4). As highly specialized and terminally
differentiated cells, podocytes lack regenerative capabilities
(4,5). ROS include free radicals, such as
superoxide, as well as non-radical species (i.e.,
H2O2). A certain concentration (200 µmol/l)
of H2O2 results in cell apoptosis (6). It is well established that signaling
pathways associated with podocyte apoptosis mainly include the
caspase, cyclin kinase and other such pathways. Caspase-9 is the
initiator caspase in the intrinsic or mitochrondrial caspase
pathway that functions to activate downstream caspase-3 in response
to apoptotic signals (7).
Honokiol (HNK) is a natural bi-phenolic compound
isolated from the Magnoliae officinalis Cortex, which is
commonly used in Traditional Chinese Medicine. Magnolia extracts
have been confirmed to exhibit anti-oxidative, anti-microbial,
anti-inflammatory, and antitumor pharmacological effects (8). It has also been reported that Magnolia
extracts may protect contrast-induced nephropathy in rats through
antioxidation and antiapoptosis in the kidney (9). Previous studies revealed that HNK
protects against renal or myocardial ischemia/reperfusion injury
via the suppression of oxidative stress, inducible nitric oxide
synthase and inflammation in rats (10–12).
Therefore, the aim of the present study was to explore the
protective effects exerted by HNK on cultured mouse podocytes and
its effect on apoptosis induced by H2O2. The
potential associated molecular mechanisms/signaling pathways were
also investigated.
Materials and methods
Reagents
HNK (purity, 98.7%) was obtained from the National
Institute for the Control of Pharmaceutical and Biological Products
of China (Beijing, China). H2O2 was obtained
from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). A CellTiter
96® AQueous Non-Radioactive Cell Proliferation Assay
(MTS) was procured from Promega Corporation (Madison, WI, USA).
Annexin V-FITC Apoptosis Detection kit was purchased from BD
Biosciences (Franklin Lakes, NJ, USA). Polyclonal antibodies
against cleaved caspase-3 (cat. no. 9664), cleaved-caspase-9 (cat.
no. 7237), protein kinase B (Akt; cat. no. 4685), extracellular
signal-regulated kinase (Erk) 1/2 (cat. no. 4695), phosphorylated
(p-)Akt (cat. no. 4060), p-Erk ½ (cat. no. 4376) and β-actin (cat.
no. 4970S) were all obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Cell culture
Conditionally immortalized mouse podocytes were
purchased from Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China). Cells were cultured in RPMI 1640
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% heat-inactivated fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.), 25 mM glucose and antibiotics
(penicillin and streptomycin) at 37°C in humidified air with 5%
CO2. The morphology of podocytes treated with
H2O2 in the absence or presence of different
concentrations (1.25, 5 and 20 µM) of HNK was observed using images
obtained from an inverted microscope (Olympus IX81; Olympus
Corporation, Tokyo, Japan; magnification, ×100).
Viability evaluation
Cultured mouse podocytes (1×104
cells/well in 96-well plate) were pre-treated with HNK (0, 1.25, 5
and 20 µM) for 2 h at 37°C and further incubated in the presence of
100 µM H2O2 for 24 h at 37°C The groups
(excluding group 1; 0 µM H2O2+0 µM HNK) were
pretreated with HNK 2 h prior to the addition of
H2O2 and then sustained with the same
concentration of HNK for 24 h to assess the effects of HNK.
Additionally, group 6 was treated with 20 µM HNK without
H2O2 in order to assess if a high
concentration of HNK affected the viability of cells. Cell
viability was evaluated using an MTS assay. Following incubation in
the appropriate medium, 20 µl phenazine methosulfate (an electron
coupling reagent) was added to each well for 1 h at 37°C in 5%
CO2 and absorbance was measured at 490 nm.
Flow cytometry analysis
At 24 h following H2O2
treatment, the apoptosis of cells treated with or without HNK were
monitored. Annexin V binding and propidium iodide (PI) staining
were determined by flow cytometry. Cells were washed with PBS
twice, and double stained at 37°C with the fluorescein
isothiocyanate (FITC)-conjugated Annexin V protein and PI for 20
min. Flow cytometry was performed using a 488 nm laser coupled to a
flow cytometer (FACSCalibur; BD Biosciences, San Jose, CA, USA) to
detect intact cells (FITC−/PI−), apoptotic
cells (FITC+/PI−) and necrotic cells
(FITC−/PI+). The data was analyzed using BD
FACSDiva 6.0 software (BD Biosciences).
Western blotting
Cultured mouse podocytes (5×106/10-cm
dish) were pre-treated with different concentrations (0, 1.25, 5
and 20 µM) of HNK for 2 h and followed by 100 µM
H2O2 for 24 h at 37°C. Cells were collected
and lysed with lysis buffer (20 mM Tris-HCl (pH 7.5), 150 mM NaCl,
1 mM EDTA, 1% Triton, 1% NP-40, 2.5 mM sodium pyrophosphate, 1 mM
β-glycerophosphate, 1 mM leupeptin, 1 mM phenylmethylsulfonyl
fluoride) for 30 min at 4°C. Extracted protein in each cell lysate
was determined using a bicinchoninic acid protein assay kit
(Pierce; Thermo Fisher Scientific, Inc.). Proteins were transferred
to a polyvinylidene difluoride membrane and blocked with 5% non-fat
dry milk in PBS with 0.02% v/v Tween-20 (PBS-Tween) for 2 h at room
temperature. The membrane was incubated for 16 h at 4°C with the
aforementioned primary antibodies at a dilution of 1:1,000.
However, anti-β-actin was used at a dilution of 1:2,000. The
membrane was washed and subsequently incubated for 1 h at room
temperature with a peroxidase-labeled Rabbit anti-Goat IgG antibody
(cat. no. SA00001-4; 1:5,000; ProteinTech Group, Inc., Chicago, IL,
USA). Following further washing, the membrane was analyzed using an
enhanced chemiluminescence kit (Pierce; Thermo Fisher Scientific,
Inc.).
Extraction of total RNA and reverse
transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted from cell lines using RNAiso
Plus (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's instructions. Samples were stored at −80°C prior to
further use. First chain cDNA synthesis was performed using the
PrimeScript™ RT Reagent kit (Takara Bio, Inc.) according to the
manufacturer's instructions and PCR was subsequently performed
using the SYBR Premix Ex Taq kit (Takara Bio, Inc.). Primer
sequences used were as follows: Caspase-3 forward,
5′-CGTGGTTCATCCAGTCCCTTT-3′ and reverse, 5′-ATTCCGTTGCCACCTTCCT-3′;
caspase-9 forward, 5′-ATCGACCCTCCGCCAGA-3′ and reverse,
5′-CAAAGGAAGCAGAACCCAT-3′; and β-actin forward,
5′-CTCTTCCAGCCTTCCTTCCT-3′ and reverse, 5′-CACCTTCACCGTTCCAGTTT-3′.
The thermocycling conditions were as follows: 95°C for 5 min,
followed by 40 cycles at 95°C for 15 sec, 60°C for 20 sec and 72°C
for 20 sec, and a final extension at 72°C for 10 min.
Statistical analysis
All experiments were performed in triplicate and
data are presented as means ± standard deviation. Statistical
significance was determined using one-way analysis of variance
followed by a Fisher's least significant difference test.
Statistical analyses were performed using SPSS 16.0 software (SPSS,
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of HNK on viability of mouse
podocytes treated with H2O2
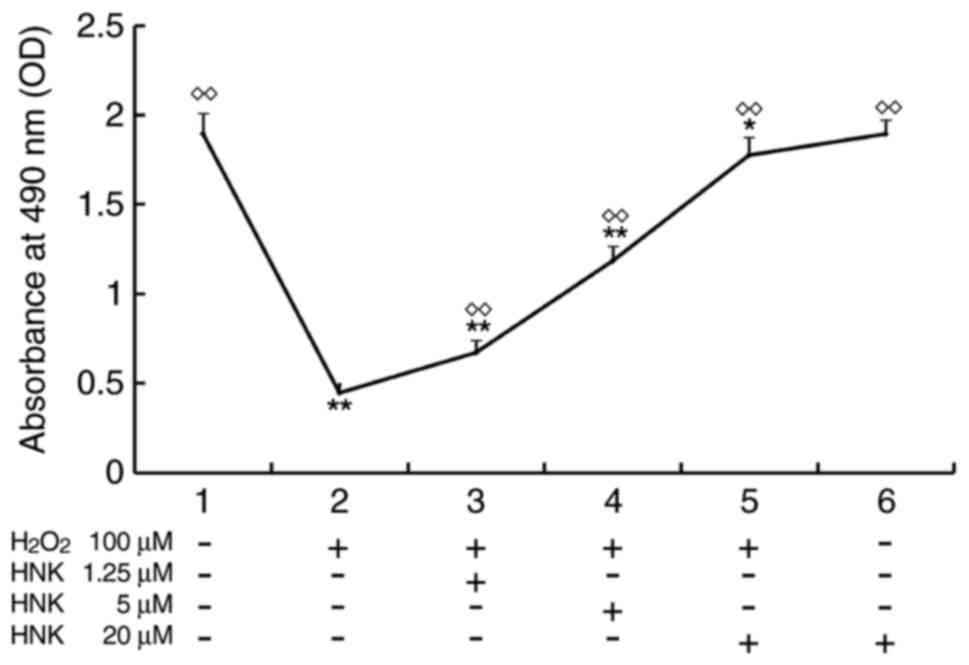
Cultured mouse podocytes were pre-treated with HNK
(0, 1.25, 5, 20 µM) for 2 h and further incubated with 100 µM
H2O2 (excluding groups 1 and 6) for 24 h.
Cell viability was evaluated via MTS assay. Results revealed that
mouse podocytes retained almost the same levels of viability
following exposure to incubation conditions with HNK concentrations
of 20 µM (group 6) compared with the normal control (group 1),
suggesting that HNK did not affect podocyte viability at certain
concentrations. However, as 100 µM H2O2
significantly reduced cell viability (group 2), the OD value was
only ~25% of that in group 1 (P<0.01). Following pre-treatment
with different concentrations of HNK, the rate of cell viability
increased in a concentration-dependent manner. OD values of groups
treated with HNK at low, medium and high concentrations were 1.39
(P<0.01), 2.5 (P<0.01) and 3.73 times (P<0.01) higher,
respectively, when compared with the group treated with
H2O2 alone (Fig.
1).
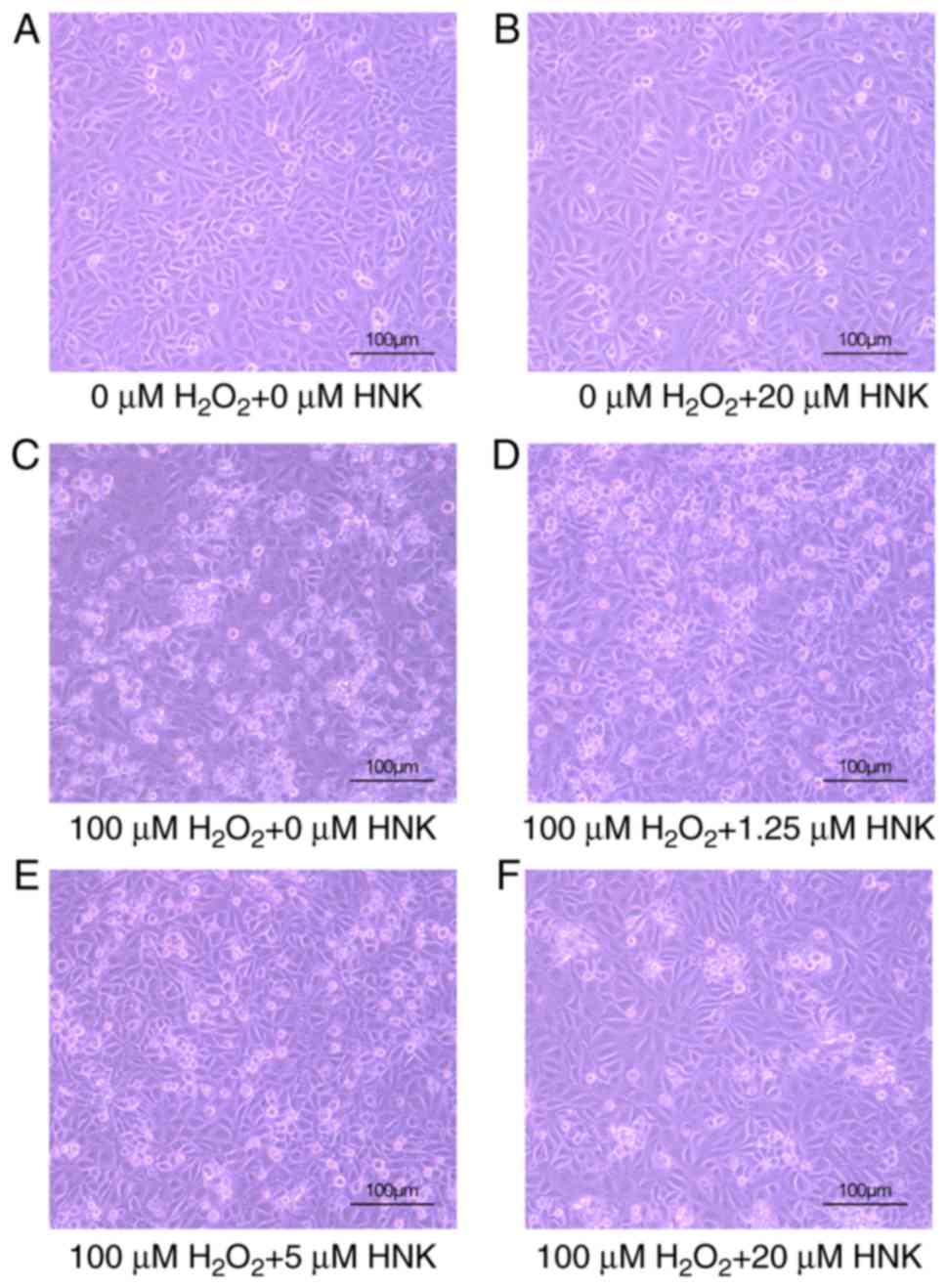
Effect of HNK on morphologic changes
in podocytes treated with H2O2
Following pre-treatment with different
concentrations of HNK for 2 h and 100 µM H2O2
for 24 h, podocytes were observed and photographed using an
inverted microscope. Microscopic observation revealed that 100 µM
H2O2 markedly affected cell physiology.
Numerous dead podocytes were noted floating in the supernatant,
whereas weakly adhered cells were observed as opaque, black-spotted
masses. Podocytes treated with HNK alone exhibit a morphology
similar to the control group with only a few floating cells in the
supernatant. Following treatment with H2O2
and different concentrations of HNK for 24 h, the morphology of
podocytes improved in a dose-dependent manner and cells gradually
became more transparent (Fig.
2).
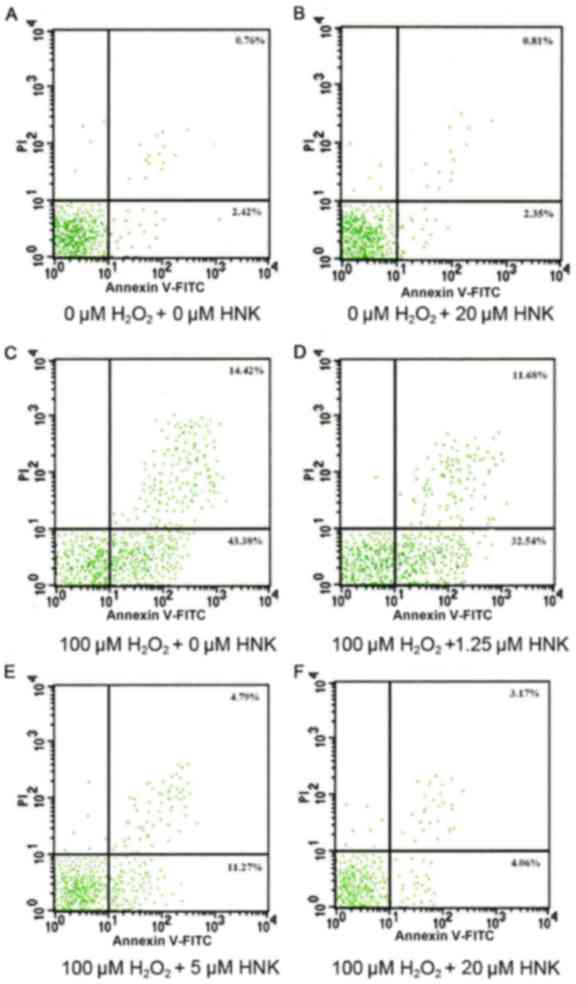
Effects of HNK on podocyte apoptosis
induced by H2O2
Results of flow cytometry revealed that the ratio of
apoptotic podocytes did not markedly differ between control and 20
µM HNK treatment groups following 24 h. Although
H2O2 typically results in an increased
apoptotic ratio, treatment with different concentrations of HNK
resulted in markedly decreases in the ratio of apoptosis in
podocytes. Compared with the H2O2-treated
group, the ratio of apoptosis in groups treated with low, medium
and high concentrations of HNK were decreased gradually. However,
the apoptosis ratio of groups treated with high concentrations of
HNK remained slightly higher than that of the control group
(Fig. 3).
Effects of HNK on the expression of
caspase-3 and caspase-9
Caspases (proteases) serve an important role in
extrinsic and intrinsic apoptotic pathways. The RT-PCR results
indicated that mRNA expression of caspase-3 and caspase-9 both
increased in podocytes treated with H2O2
compared with control cells. Additionally, although mRNA levels of
caspase 9 were marginally higher in cells treated with 100 µM
H2O2 +1.25 µM HNK than in those treated with
100 µM H2O2 alone, HNK gradually
downregulated expression of cleaved caspase-3 and caspase-9 mRNA in
a concentration from 5 to 20 µM (Fig.
4). Furthermore, western blotting analysis of cleaved caspase-3
and caspase-9 protein levels produced similar findings as those of
their mRNA levels, although the protein level of cleaved caspase-3
was margnally higher in cells treated with 100 µM
H2O2 +1.25 µM HNK than in those treated with
100 µM H2O2 alone (Fig. 5). These results suggested an obvious
inhibition of cleaved caspase-3, −9 protein and caspase-3, −9 mRNA
expression by HNK.
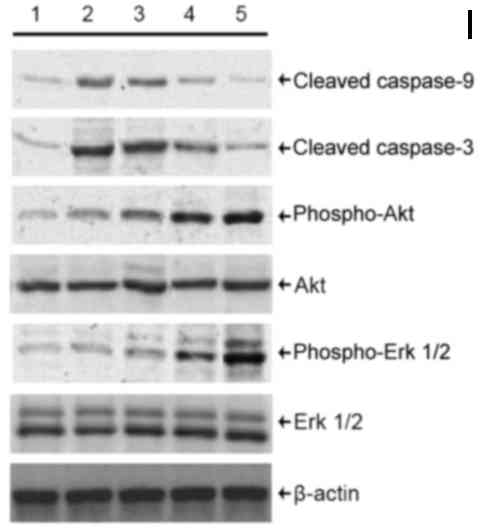 | Figure 5.Detection of cleaved caspase-3 and −9
protein, p-Akt and p-Erk 1/2 protein levels in podocytes treated
with HNK and H2O2. Levels of cleaved caspase-3 and −9 protein, Akt,
Erk 1/2, p-Akt and p-Erk 1/2 protein were detected by western
blotting. Cells were divided into the following treatment groups:
1, 0 µM H2O2 + 0 µM HNK (normal control); 2, 100 µM H2O2 + 0 µM
HNK; 3, 100 µM H2O2 +1.25 µM HNK; 4, 100 µM H2O2 + 5 µM HNK; and 5,
100 µM H2O2 + 20 µM HNK. HNK, honokiol. p, phosphorylated; Akt,
protein kinase B; Erk, extracellular signal-regulated kinase; HNK,
honokiol. |
Mechanisms associated with
HNK-inhibition of H2O2-induced oxidative
stress in mouse podocytes
Recent findings have demonstrated that Akt and Erk
signaling pathways is associated with the regulation of apoptosis
in various of cell types, tissues and organs in many diseases
(13–15). In the present study, protein levels
of total Akt and Erk 1/2 exhibited no marked changes, whereas
levels of p-Akt and p-Erk 1/2 appeared to increase following HNK
treatment, peaking at the highest concentration of HNK (20 µM).
However, levels of p-Akt and p-Erk 1/2 did not exhibit any marked
declines following H2O2 treatment alone. To
some extent, p-Akt and p-Erk 1/2 levels increased in cells treated
with H2O2 compared with the control group,
suggesting an inconsistent tendency associated with the expression
of caspase in cells pre-treated with HNK (Fig. 5).
Discussion
An increased amount of urinary protein, termed
albuminuria (typically 67 kDa), is among the earliest signs of DKD
and strongly correlates with progression towards ESRD (2). The glomerular filtration barrier
consists of three layers: Capillary endothelium, GBM and the
glomerular epithelial cell (or podocyte) layer. Numerous studies
(16–18) concerning DKD emphasized damage to
mesangial cells and the glomerular basement membrane. Glomerular
hypertrophy, mesangial matrix expansion and GBM thickening are
classic signs of diabetic glomerular diseases (19). Previous evidence (20–22) have
demonstrated that the onset of proteinuria is closely associated
with pathological changes in podocytes, such as hypertrophy,
detachment, apoptosis and epithelial-to-mesenchymal transition.
ROS promotes renal injury, exacerbating the
progression of kidney disease (2).
Previous studies (23,24) have demonstrated that ameliorating
oxidative stress through treatment with antioxidants may be an
effective strategy for reducing diabetic complications. Several
clinical (25,26) trials have confirmed the effects of
certain antioxidants on the prevention of diabetic complications.
Under normal physiological conditions, the levels of cellular ROS
remain stable in a dynamic equilibrium. The destruction of this
balance promotes ROS accumulation, which causes molecular, cellular
and clinical abnormalities (6). In
the present study, data from the MTS assay revealed that 100 µM
H2O2 significantly reduced cell viability,
with the OD value being ~25% of the normal control group with
identical flow cytometry outcomes. In addition, observation under
inverted microscopy indicated that 100 µM
H2O2 significantly affected cellular
physiology.
HNK serves an anti-oxidative role by inhibiting
NADPH oxidase, myeloperoxidase and cyclooxygenase while increasing
glutathione peroxidase activity in neutrophils to promote
metabolism of H2O2 (27). The present study demonstrated that
H2O2 reduces the ratio of viable podocytes as
well as increases the ratio of cell apoptosis. Additionally,
cultured mouse podocytes were pre-treated with HNK (0, 1.25, 5 and
20 µM) for 2 h and further incubated in 100 µM
H2O2 for 24 h. The effect of HNK on
peroxide-induced podocyte apoptosis was subsequently investigated.
Podocytes were also treated with high concentrations (20 µM) of HNK
alone to observe the influence of HNK on cellular viability and
apoptosis as well. Following 24 h treatment with different
concentrations of HNK and 100 µM H2O2, the
morphology of adherent podocytes improved in a dose-dependent
manner compared with H2O2 treatment alone. In
addition, HNK treatment alone did not markedly alter podocyte
physiology in comparison with the control group. These results
demonstrated that HNK, within certain safe ranges of concentration,
can protect podocytes from damage induced by oxidative stress. MTS
assay and flow cytometry further confirmed that HNK treatment lead
to significantly lower apoptotic ratios and greater viability rates
in podocytes. Nonetheless, the ratio of apoptosis in cells treated
with HNK at high concentrations remained slightly higher as
compared with the control group.
Casepase-9 is generally considered to be an initial
mediator of apoptosis induced by H2O2
(28). The eventual release of
cytochrome C promotes activation of caspase-3 (which can also be
activated to strengthen caspase-3), thus resulting in activation of
caspase-associated DNase. Activated DNase degrades DNA to mere
fragments. This is one important marker of cellular apoptosis
(28). In the present study, RT-PCR
and western blotting results confirmed that the expression of both
mRNA and protein of cleaved caspase-3 and −9 markedly increased in
podocytes treated with H2O2. Pretreatment
with HNK markedly downregulated expression of both protein and mRNA
of caspase-3 and −9 in a concentration-dependent manner.
Previous studies also reported the anti-apoptotic
effects of hepatocyte growth factor (HGF) on podocytes in
vitro. HGF stimulation resulted in the phosphorylation of Akt
and Erk, and induction of an X-linked inhibitor of apoptosis
protein (XIAP) in podocytes (29).
Furthermore, phosphorylation of Akt and Erk 1/2 was attenuated,
whereas the expression of cleaved caspase-3 and the number of TUNEL
positive cells was enhanced in vascular endothelial cells exposed
to H2O2 (30).
In another study, cultured mouse neural progenitor cells were
treated with H2O2, apoptotic signaling
pathways were activated and the phosphorylation of Akt and Erk
decreased. Astaxanthin pretreatment, however, significantly
inhibited H2O2-mediated caspase activation
(31). The activation of
phosphorylation of Erk 1/2, which blocked the release of cytochrome
C from mitochondria, resulted in inhibition of caspase-9 and −3
activation. The ratio of apoptosis also decreased (31). The phosphoinositide 3-kinase
(PI3K)/Akt/glycogen synthase kinase (GSK)-3 signal transduction
pathway is a vital modality of intracellular membrane receptor
signal transduction and serves an important role in the regulation
of apoptosis in a variety of organs (32). Akt is a serine/threonine protein
kinase characterized by multiple sites of phosphorylation. PI3K
inhibits the downstream apoptosis-related protein GSK-3β by
enhancing the activation of Akt, thus having an anti-apoptotic
effect. GSK-3 serves a critical role in regulation of apoptosis,
which can inhibit transcription factors, such as heat shock
factor-1 (a cyclic adenosine monophosphate binding protein) and
activate members of the caspase family, leading to apoptosis
(32,33).
The present data indicated that as HNK concentration
increased, total Akt and Erk 1/2 protein levels did not markedly
change. However, levels of p-Akt and Erk 1/2 gradually increased,
peaking at 20 µM HNK in a concentration-dependent manner. As the
expression of cleaved caspase-3 and −9 gradually decreased, levels
of p-Akt and -Erk 1/2 increased with HNK treatment. Notably, in the
present study, levels of p-Akt and -Erk 1/2 in the group treated
with H2O2 alone did not significantly
decrease as compared with the normal control group. This phenomenon
seemed inconsistent with the tendency of the expression of caspase
in HNK-treated cells and contradicted the results of MTS and flow
cytometry. It was speculated that the potency of
H2O2 inevitably weakens as time passes, and
cells exhibit a certain degree of self-repair capacity. Levels of
p-Akt and Erk 1/2 therefore increased slightly under
H2O2 treatment alone following 24-h
incubation. Nevertheless, the effects observed in the present study
are not necessarily indicative of a cause-and-effect relationship.
Furthermore, it is necessary to interfere with Akt and Erk at the
genetic level to confirm the targets of HNK in
H2O2-treated podocytes.
In conclusion, the present study confirmed that HNK
serves a vital role in protecting against apoptosis in podocytes
treated with H2O2 by means of inhibiting
caspase-3 and −9 activation as well as enhancing phosphorylation of
Akt and Erk 1/2. HNK may have potential as a treatment for kidney
diseases exacerbated by oxidative stress damage.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Zhejiang
Provincial Natural Science Foundation of China (LY14H070002 and
LQ16H070001), the Zhejiang Provincial Medical Science and
Technology Program (Backbone Project of Platform Program;
2015RCA013) and the Zhejiang Provincial Administration of
traditional Chinese Medicine Project (2015ZA058).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FW and HY conceived and designed the study, wrote
the protocol, performed the experiments, analyzed the data,
performed the literature search and approved the final manuscript.
FZ, JZ and HL supervised the research and contributed to the flow
cytometric analysis and cell viability evaluation. XL, LL and ST
were involved in data acquisition and helped perform the cell
biology experiments. FW wrote the first draft of the manuscript, HL
helped revise the manuscript. All co-authors approved the final
version of the manuscript. FW and HY had full access to all the
data in this study and take responsibility for the integrity of the
data.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Thorner PS, Ho M, Eremina V, Sado Y and
Quaggin S: Podocytes contribute to the formation of glomerular
crescents. J Am Soc Nephrol. 19:495–502. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gnudi L, Coward RJM and Long DA: Diabetic
nephropathy: Perspective on novel molecular mechanisms. Trends
Endocrinol Metab. 27:820–830. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marshall CB and Shankland SJ: Cell cycle
and glomerular disease: A minireview. Nephron Exp Nephrol.
102:e39–e48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xia H, Bao W and Shi S: Innate immune
activity in glomerular podocytes. Front Immunol. 8:1222017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shibata S, Nagase M, Yoshida S, Kawachi H
and Fujita T: Podocyte as the target for aldosterone: Roles of
oxidative stress and Sgk1. Hypertension. 49:355–364. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Johansen JS, Harris AK, Rychly DJ and
Ergul A: Oxidative stress and the use of antioxidants in diabetes:
Linking basic science to clinical practice. Cardiovasc Diabetol.
4:52005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zou H, Yang R, Hao J, Wang J, Sun C, Fesik
SW, Wu JC, Tomaselli KJ and Armstrong RC: Regulation of the
Apaf-1/caspase-9 apoptosome by caspase-3 and XIAP. J Biol Chem.
278:8091–8098. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fried LE and Arbiser JL: Honokiol, a
multifunctional antiangiogenic and antitumor agent. Antioxid Redox
Signal. 11:1139–1148. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang F, Zhang G, Zhou Y, Gui D, Li J, Xing
T and Wang N: Magnolin protects against contrast-induced
nephropathy in rats via antioxidation and antiapoptosis. Oxid Med
Cell Longev. 2014:2034582014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Zhang ZZ, Wu Y, Zhan J, He XH and
Wang YL: Honokiol protects rat hearts against myocardial ischemia
reperfusion injury by reducing oxidative stress and inflammation.
Exp Ther Med. 5:315–319. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu F, Zhang W, Li L, Zheng F, Shao X, Zhou
J and Li H: Inhibitory effects of honokiol on
lipopolysaccharide-induced cellular responses and signaling events
in human renal mesangial cells. Eur J Pharmacol. 654:117–121. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu Y, Li M, Su N, Zhang Z, Zhao H, Yu H
and Xu Y: Honokiol protects against renal ischemia/reperfusion
injury via the suppression of oxidative stress, iNOS, inflammation
and STAT3 in rats. Mol Med Rep. 13:1353–1360. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zeng W, Tang J, Li H, Xu H, Lu H, Peng H,
Lin C, Gao R, Lin S, Lin K, et al: Caveolin-1 deficiency protects
pancreatic β cells against palmitate-induced dysfunction and
apoptosis. Cell Signal. 47:65–78. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li L, Wang X, Sharvan R, Gao J and Qu S:
Berberine could inhibit thyroid carcinoma cells by inducing
mitochondrial apoptosis, G0/G1 cell cycle arrest and suppressing
migration via PI3K-AKT and MAPK signaling pathways. Biomed
Pharmacother. 95:1225–1231. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang B, Shen Q, Chen Y, Pan R, Kuang S,
Liu G, Sun G and Sun X: myricitrin alleviates oxidative
stress-induced inflammation and apoptosis and protects mice against
diabetic cardiomyopathy. Sci Rep. 7:442392017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sargin AK, Can B and Turan B: Comparative
investigation of kidney mesangial cells from increased oxidative
stress induced diabetic rats by using different microscopy
techniques. Mol Cell Biochem. 390:41–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Thomson SE, McLennan SV, Kirwan PD,
Heffernan SJ, Hennessy A, Yue DK and Twigg SM: Renal connective
tissue growth factor correlates with glomerular basement membrane
thickness and prospective albuminuria in a non-human primate model
of diabetes: Possible predictive marker for incipient diabetic
nephropathy. J Diabetes Complications. 22:284–294. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang A, Ziyadeh FN, Lee EY, Pyagay PE,
Sung SH, Sheardown SA, Laping NJ and Chen S: Interference with
TGF-beta signaling by Smad3-knockout in mice limits diabetic
glomerulosclerosis without affecting albuminuria. Am J Physiol
Renal Physiol. 293:F1657–F1665. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Osterbt R and Gundersen HJ: Glomerular
size and structure in diabetes mellitus. I. Early abnormalities.
Diabetologia. 11:225–229. 1975.
|
|
20
|
Jefferson JA, Shankland SJ and Pichler RH:
Proteinuria in diabetic kidney disease: A mechanistic viewpoint.
Kidney Int. 74:22–36. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
White KE, Bilous RW, Marshall SM, El Nahas
M, Remuzzi G, Piras G, De Cosmo S and Viberti G: Podocyte number in
normotensive type 1 diabetic patients with albuminuria. Diabetes.
51:3083–3089. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
DallaVestra M, Masiero A, Roiter AM,
Saller A, Crepaldi G and Fioretto P: Is podocyte injury relevant in
diabetic nephropathy? Studies in patients with type 2 diabetes.
Diabetes. 52:1031–1035. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mekinová D, Chorváthová V, Volkovová K,
Staruchová M, Grancicová E, Klvanová J and Ondreicka R: Effect of
intake of exogenous vitamins C, E and beta-carotene on the
antioxidative status in kidneys of rats with streptozotocin-induced
diabetes. Nahrung. 39:257–261. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Obrosova I, Fathallah L and Greene D:
Early changes in lipid peroxidation and antioxidative defense in
rat retina: Effect of DL-alpha-lipoic acid. Eur J Pharm.
398:139–146. 2000. View Article : Google Scholar
|
|
25
|
Skyrme-Jones RA, O'Brien RC, Berry KL and
Meredith IT: Vitamin E supplementation improves endothelial
function in type I diabetes mellitus: A randomized,
placebo-controlled study. J Am Coll Cardiol. 36:94–102. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gaede P, Poulsen HE, Parving HH and
Pedersen O: Double-blind, randomised study of the effect of
combined treatment with vitamin C and E on albuminuria in Type 2
diabetic patients. Diabet Med. 18:756–760. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liou KT, Shen YC, Chen CF, Tsao CM and
Tsai SK: The anti-inflammatory effect of honokiol on neutrophils:
Mechanisms in the inhibition of reactive oxygen species production.
Eur J Pharmacol. 475:19–27. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tuo QH, Wang C, Yan FX and Liao DF: MAPK
pathway mediates the protective effects of onychin on oxidative
stress-induced apoptosis in ECV304 endothelial cells. Life Sci.
76:487–497. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Agustian PA, Schiffer M, Gwinner W,
Schäfer I, Theophile K, Modde F, Bockmeyer CL, Traeder J, Lehmann
U, Grosshennig A, et al: Diminished met signaling in podocytes
contributes to the development of podocytopenia in transplant
glomerulopathy. Am J Pathol. 178:2007–2019. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang B, Oo TN and Rizzo V: Lipid rafts
mediate H2O2 prosurvival effects in cultured endothelial cells.
FASEB J. 20:1501–1503. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim JH, Choi W, Lee JH, Jeon SJ, Choi YH,
Kim BW, Chang HI and Nam SW: Astaxanthin inhibits H2O2-mediated
apoptotic cell death in mouse neural progenitor cells via
modulation of P38 and MEK signaling pathways. J Microbiol
Biotechnol. 19:1355–1363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim JW, Lee JE, Kim MJ, Cho EG, Cho SG and
Choi EJ: Glycogen synthase kinase 3 beta is a natural activator of
mitogen-activated protein kinase/extracellular signal-regulated
kinase kinase kinase 1 (MEKK1). J Biol Chem. 278:13995–14001. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pap M and Cooper GM: Role of glycogen
synthase kinase-3 in the phosphatidylinositol 3-Kinase/Akt cell
survival pathway. J Biol Chem. 273:19929–19932. 1998. View Article : Google Scholar : PubMed/NCBI
|