Introduction
The endophytic is a fungi of the genus
Pestalotiopsis which exists widely in many plants of
different families, have received wide attention because of their
ability in producing a variety of secondary metabolites, such as
caryophyllene sesquiterpenoids (1,2),
disseminins and spiciferone analogues (3), prenyldepside and coumarins (4), diphenyl ether derivatives (5), benzannulated spiroketals (6), chromones (7), 1,3-dihydro isobenzofuran (8), epoxyquinols (9), and polyketide derivatives (10). It is reported that the well-known
anticancer drug taxol has been found from several cultivated
species of the genus Pestalotiopsis, which provides another
alternative way to produce such valuable anti-cancer drugs
(11,12). The active secondary metabolites
produced by the endophytic fungi can stimulate the growth and
development of the organisms, or improve the ability of the host to
resist the external environment (13). Pharmacological researches have
revealed that the secondary metabolites from species of
Pestalotiopsis possess diverse pharmacological properties,
including antimicrobial activity (14–16),
antiviral (17), antitumor (18), anti-diabetic activity (19), immunomodulatory activity (20), anticestodal activity (21), antioxidant and antihypertensive
properties (22), and 20S proteasome
inhibitory activity (23).
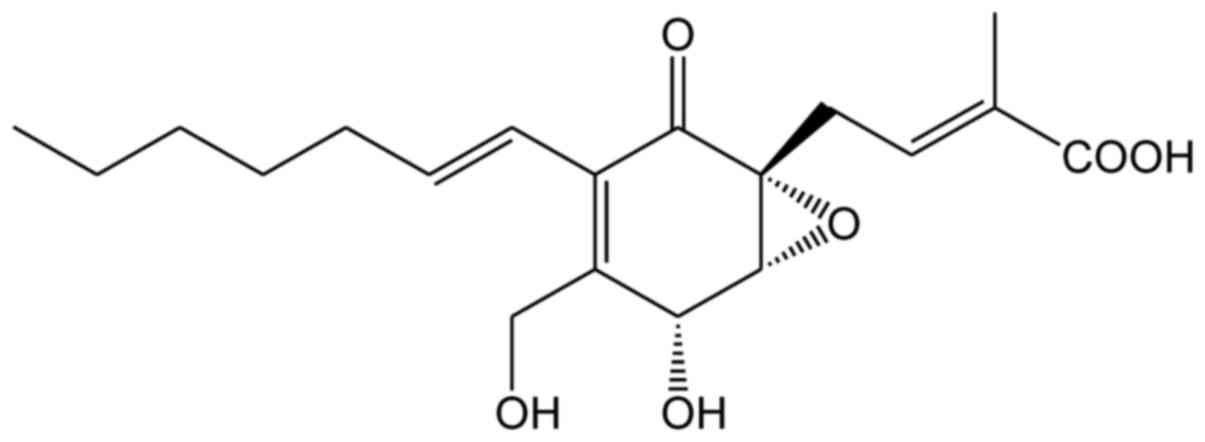
Ambuic acid (chemical structure shown in Fig. 1) was firstly reported as a novel
functionalized cyclohexenone initially isolated from
Pestalotiopsis spp. and Monochaetia sp. in 2001
(24), and its relative
stereochemistry was predicted by using a novel solid-state nuclear
magnetic resonance (NMR) approach in 2003 (25). Ambuic acid showed a certain
inhibitory effect on a variety of plant pathogenic fungi
Fusarium solani, Fusarium cubense, Helminthosporium sativum,
Diplodia natelensis, Cephalosporium gramineum, Pythium ultimum
(24), and it has a certain
antimicrobial activity against the Gram-positive bacterium
Staphyococcus aureus (26).
Ambuic acid can also effect the biosynthesis of cyclic peptide
quormones in Gram-positive bacteria and Enterococcus
faecalis (27). In our recent
studies, a plant pathogenic fungus Pestalotiopsis neglecta
(FJ-2) has been separated from the twig of Camellia sinensis
growing in Fujian Province of China. Futhermore, four new ambuic
acid derivatives together with ambuic acid and three other known
ramification, have been isolated and identified from the solid
culture of Pestalotiopsis neglecta (28). Inhibitory activities of these
compounds against LPS-induced overproduction of nitric oxide (NO)
in macrophages have been evaluated. Ambuic acid exhibited strong
inhibitory activity with IC50 value of 20.80±1.41 µM,
which was even much potent than the positive control drug.
Macrophages play an important role in providing
immediate defense against foreign stimuli. Lipopolysaccharide
(LPS)-mediated overproduction of NO and subsequent inflammatory
reaction by macrophages is proved to be closely associated with
many pathogenic diseases, including rheumatoid arthritis (29), cancer (30), atherosclerosis (31), and hepatitis (32). By reducing NO formation or removing
NO molecule, NO inhibitors may be used as powerful therapeutic
anti-inflammatory agents (33). In
the present paper, the anti-inflammatory effect of ambuic acid as a
NO inhibitor was firstly investigated and the molecular mechanism
of the anti-inflammatory action of ambuic acid was absolutely
elucidated.
Materials and methods
Fungal material
The pathogenic strain (FJ-2) used in this work was
separated from the twig of Camellia sinensis growing in
Fujian Province of China. The identification of fungus was based on
the (deoxyribonucleic acid) DNA sequences of the ITS1-5.8S-ITS2,
and the ITS regions of their ribosomal ribonucleic acid (RNA) gene.
We have submitted the sequence data derived from the fungal strain
and stored them at GenBank with accession number KJ719299. The
result of BLAST search showed that the similarity between the
sequence and the sequence of Pestalotiopsis neglecta (Thüm.)
was 99%. It was stored in GenBank with the accession number of
JX854541.
The fungal strain was placed on the slants of potato
dextrose agar and cultured at 25°C for 10 days. We inoculated the
agar plugs in 500 ml Erlenmeyer flask, it contains 120 ml of media
(0.4% glucose, 1% malt extract, and 0.4% yeast extract, before
sterilization the final pH of the medium was adjusted to 6.5), and
it was incubated at 25°C on a rotary shaker at 180 rpm for 4 days.
Large scale fermentation taked place in 40 bottles of 500 ml
Fernbach flasks, which containing 80 g of rice and 100 ml of
distilled water. Each flask was inoculated with 10.0 ml of culture
medium and cultured at 25°C for 40 days.
Isolation and identification of ambuic
acid
The ethyl acetate was used to extract the fermented
rice substrate, and the organic solvent was decompressed and
concentrated to obtain the crude extract, silica gel column
chromatography (CC) with a gradient of n-hexane-ethyl acetate,
dichloromethane-methanol were used to divided them into five groups
(A-E).
Fraction C was treated with dichloromethane-methanol
and subjected to ODS CC eluting with a gradient of methanol in
water (10%~100%) to obtain twenty-one subfractions (C1-C21).
Subfraction C13 was separated by RP-HPLC using 63% methanol in
water to afford ambuic acid. Its chemical structure was validated
by extensive NMR experiments and validated by comparison with the
data previously incorpratated into the databases of SciFinder
Scholar and PubChem. The purity was determined using
high-performance liquid chromatography (HPLC; Waters 600; Waters
Corp., Milford, MA, USA) and a ultraviolet detector (Waters 490;
Waters Corp.); peak area was normalized and the purity of ambuic
acid was 98.7%.
Chemicals and reagents
Fetal bovine serum (FBS) and RPMI 1640 culture
medium were purchased from Invitrogen (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). LPS, DMSO, MTT, PMSF and all other
chemicals were obtained from Sigma-Aldrich; Merck KGaA (Darmstadt,
Germany). Hydrocortisone succinate (080–05581, Lot CTE6574) was
product of Wako Pure Chemical Industries, Ltd. (Osaka, Japan). NO
concentration determination kit, mouse prostaglandin E2 (PGE2)
ELISA kit, mouse tumor necrosis factor-α (TNF-α) ELISA kit, mouse
interleukin-6 (IL-6) ELISA kit, and BCA protein concentration assay
kit were purchased from Yantai Science and Biotechnology Co., Ltd.
(Shandong, China). NO synthase (NOS) assay kit (fluorimetric
method) was product of Beyotime Institute of Biotechnology (Haimen,
China). COX colorimetric inhibitor screening assay kit (701050) was
product of Cayman Chemical Company (Ann Arbor, MI, USA). Mouse
anti-rabbit inducible NOS (iNOS) polyclonal antibody (160862) and
mouse anti-rabbit cyclooxygenase-2 (COX-2) polyclonal antibody
(160106) were purchased from Cayman Chemical Company. Goat
anti-rabbit phosphorylated c-Jun N-terminal kinase (p-JNK)
polyclonal antibody (AF3318), goat anti-rabbit phosphorylated
extracellular signal-regulated kinase 1/2 (p-ERK 1/2) polyclonal
antibody (AF1015), goat anti-rabbit phosphorylated p38 (p-p38)
polyclonal antibody (AF3455), and horseradish peroxidase
(HRP)-conjugated goat anti-rabbit IgG (H+L) (S0001) were products
of Affinity Biosciences. Goat anti-rabbit IκB-α polyclonal antibody
(sc-371), mouse monoclonal nuclear transcription factor-κB (NF-κB)
p65 antibody (sc-8008), and goat anti-rabbit β-actin polyclonal
antibody (sc-1616) were purchased from Santa Cruz Biotechnology,
Inc. A solution for dissolving ambuic acid with 100% cell culture
grade dimethyl sulfoxide (DMSO) into 50 mM and stored as small
aliquots at −20°C and then diluted it to desired concentrations
prior to use.
Cell culture
Mouse monocyte-macrophage RAW264.7 cells (ATCC
TIB-71; American Type Culture Collection, Manassas, VA, USA) were
cultured in RPMI-1640 medium supplemented with 10% heat inactivated
FBS in a humidified incubator with 95% air and 5% CO2 at
37°C. Every two days, the medium was routinely changed. When the
cells attained about 80% confluence, they were passaged.
MTT assay for cytotoxicity
Using the measured of mitochondrial-dependent
reduction of MTT to formazan to measure the cytotoxicity (34). RAW264.7 cells were seeded in 96-well
plates at the density of 1×106 cells/ml. After 1 h of
incubation, the cells were treated with serially diluted ambuic
acid (the concentrations were from 0.78125 to 100 µM) and incubated
for 24 h. After treatment, adding MTT solution with the
concentration of 200 µg/ml, and set the cells to the incubator for
another 4 h at 37°C. Removing the medium and adding 150 µl of DMSO
to dissolve the formazan. Using the microplate reader at 570 nm to
measure the absorbance of each group and using a reference
wavelength at 630 nm (Biotek Synergy HT; BioTek Instruments, Inc.,
Winooski, VT, USA). The control group, consisting of untreated
cells, was considered to be 100% viable. Final results are
expressed as percentage of viable cells of the experimental group
when compared with those of the control group.
NO analysis
NO was determined through measuring the nitrite
concentration in the cell culture supernatant by using Griess
reagent (mixture of equal amount of reagent A and reagent B; A: 1%
sulphanilamide in 5% H3PO4, B: 0.1%
naphthylethylene diamine dihydrochloride) (35). RAW264.7 cells were seeded in 96-well
plates at the density of 1×106 cells/ml. After 1 h
incubation, the cells were treated by LPS (1 µg/ml) with or without
ambuic acid (3.125, 6.25, 12.5, 25, 50, 100 µM), or hydrocortisone
succinate (100 µM) and then incubated for 24 h. 100 µl of the cell
culture supernatant was mixed with 100 µl of Griess reagent and
shaked for 10 min at room temperature. The absorbance was measured
at 540 nm, and using a standard calibration curve to calculate the
nitrite concentrations which was prepared from a range of different
concentrations of sodium nitrite.
PGE2 concentration
determination
PGE2, a pro-inflammatory mediator, is
produced by COX-2. RAW264.7 cells were treated by LPS (1 µg/ml)
with or without ambuic acid (3.12, 6.25, 12.5, 25, 50, 100 µM), or
hydrocortisone succinate (100 µM) for 24 h. The cell culture
supernatant of 100 µl was removed to determine the level of
PGE2 by using a commercial mouse PGE2 ELISA
kit according to the manufacturer's recommendations. The ELISA data
representing mean values ± SD (standard deviation) were obtained in
duplicate from three independent experiments (36).
Measurement of cytokines TNF-α and
IL-6
RAW264.7 cells were seeded in 96-well plates at the
density of 5×105 cells/ml. After 1 h incubation, the
cells were treated by LPS (1 µg/ml) with or without ambuic acid
(3.12, 6.25, 12.5, 25, 50, 100 µM), or hydrocortisone succinate
(100 µM) for 6 h. 100 µl of the culture supernatants was collected
to determine the levels of TNF-α or IL-6 by using respective ELISA
kit according to the manufacturer's recommendations (37). The ELISA data representing mean
values ± SD were obtained in duplicate from three independent
experiments.
Assay of iNOS enzymatic activity
Methods for the determination of iNOS enzymatic
activity as previously reported (38). Briefly, after the treatments,
removing the culture supernatant, and adding 100 µl of NOS assay
buffer (1X). Then adding 100 µl of NOS assay reaction solution and
incubated for 2 h at 37°C. Using fluorescence microplate reader
measured the fluorescence at excitation wavelength of 485 nm and
emission wavelength of 528 nm.
Assay of COX-2 enzymatic activity
The enzymatic activity of COX-2 was determined in a
cell-free system by using a COX colorimetric inhibitor screening
assay kit according to the manufacturer's instructions (38). Briefly, 160 µl of assay buffer, 10 µl
of heme, and 10 µl of DMSO were added to the background wells. 150
µl of assay buffer, 10 µl of COX-2 enzyme, 10 µl of heme, and 10 µl
of DMSO were added to the 100% initial activity wells. 150 µl of
assay buffer, 10 µl of COX-2 enzyme, 10 µl of heme, and 10 µl of
ambuic acid or hydrocortisone succinate were added to the sample
wells. The plate was gently shaken for a few sec and then incubated
for five min at 25°C. Adding 20 µl of the colorimetric substrate
solution and then adding 20 µl of arachidonic acid to all the
wells. Shaking the plate carefully for a few sec and incubating it
for five min at 25°C. The absorbance was measured at 590 nm by
using a microplate reader, and the enzymatic activity of COX-2 was
calculated when compared with the 100% initial activity wells
according to the manufacturer's instructions.
Total protein extraction
To determination the expression of iNOS and COX-2
proteins, adding the LPS (1 µg/ml) to the RAW264.7 cells with or
without indicated concentrations of ambuic acid for 24 h. Then
washing it with ice-cold phosphate-buffered saline (PBS),
harvesting the cells and adding 40 µl of cold lysis buffer [10%
NP-40, 150 mM NaCl, 10 mM Tris, 2 mM PMSF, 5 µM leupeptin, pH 7.6]
to lyse the cell. For determination of phospho-JNK, phospho-ERK
1/2, and phospho-p38 proteins, added the LPS (1 µg/ml) to the
RAW264.7 cells with or without indicated concentrations of ambuic
acid for 30 min. Then washing it with ice-cold PBS, harvesting the
cells and adding 40 µl of the same cold lysis buffer mentioned
above to lyse the cell. After incubating at 4°C for 15 min,
centrifuging it at 15,000 × g for 10 min at 4°C, the total proteins
were separated and used for western blot analysis.
Cytoplasmic and nuclear protein
extraction
After proper treatments, removing the media, and
washing the cells with ice-cold PBS, then harvesting the cells and
adding 40 µl of buffer A [10 mM HEPES, 1.5 mM MgCl2, 10
mM KCl, 500 µM DTT, 0.1% (v/v) NP-40] to lyse the cell. After
incubating at 4°C for 15 min, centrifuging it at 12,000 × g for 5
min at 4°C, the cytoplasmic proteins were separated and used for
western blot analysis of IκB-α. The ectraction of nuclei pellet is
to suspend precipitation in 30 µl of buffer B [20 mM HEPES, 1.5 mM
MgCl2, 400 mM NaCl, 100 mM DTT, 20 µM PMSF] and
incubated for 20 min at 4°C. By centrifuging at 16,000 × g for 10
min at 4°C, the supernatant was separated as nuclear protein and
used for western blot analysis of NF-κB p65 subunit.
Western blotting
Using the BCA protein concentration assay kit to
determine the protein concentration of each aliquot, and the
suspensions were boiled with SDS-PAGE loading buffer. The 30 µg of
protein in each sample were transferred to the nitrocellulose
membranes by SDS-PAGE and electrophoretically. The membranes were
blocked with 5% non-fat dried milk in Tris-buffered saline and
Tween-20 (TBST, 20 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20) for 4
h at room temperature. After washed with TBST, incubating the
membranes in respective primary antibody solution (anti-iNOS,
anti-COX-2, anti-phospho-JNK, anti-phospho-ERK, anti-phospho-p38,
anti-IκB-α, anti-p65, and anti-β-actin antibody) overnight at 4°C.
Washing the membranes with TBST and then incubating them with
HRP-conjugated secondary antibody solution for 1 h at room
temperature. The membranes were washed three times with TBST and
the blots were detected by using enhanced chemiluminescence reagent
(ECL) and exposed to photographic films (Kodak, Rochester, NY,
USA). Images were collected and the bands corresponding to iNOS,
COX-2, phospho-ERK, phospho-JNK, phospho-p38, IκB-α, p65 and
β-actin protein were quantitated by densitometric analysis using
the DigDoc100 program (Alpha Ease FC software). Data of iNOS,
COX-2, phospho-ERK, phospho-JNK, phospho-p38, IκB-α and p65 were
normalized on the basis of β-actin levels.
Statistical analysis
All data were shown as mean ± SD. Significant
differences among different treatment groups were analyzed by
two-way analysis of variance (ANOVA) followed by the post
hoc Bonferroni test. P<0.05 was considered to indicate a
statistically significant difference.
Results
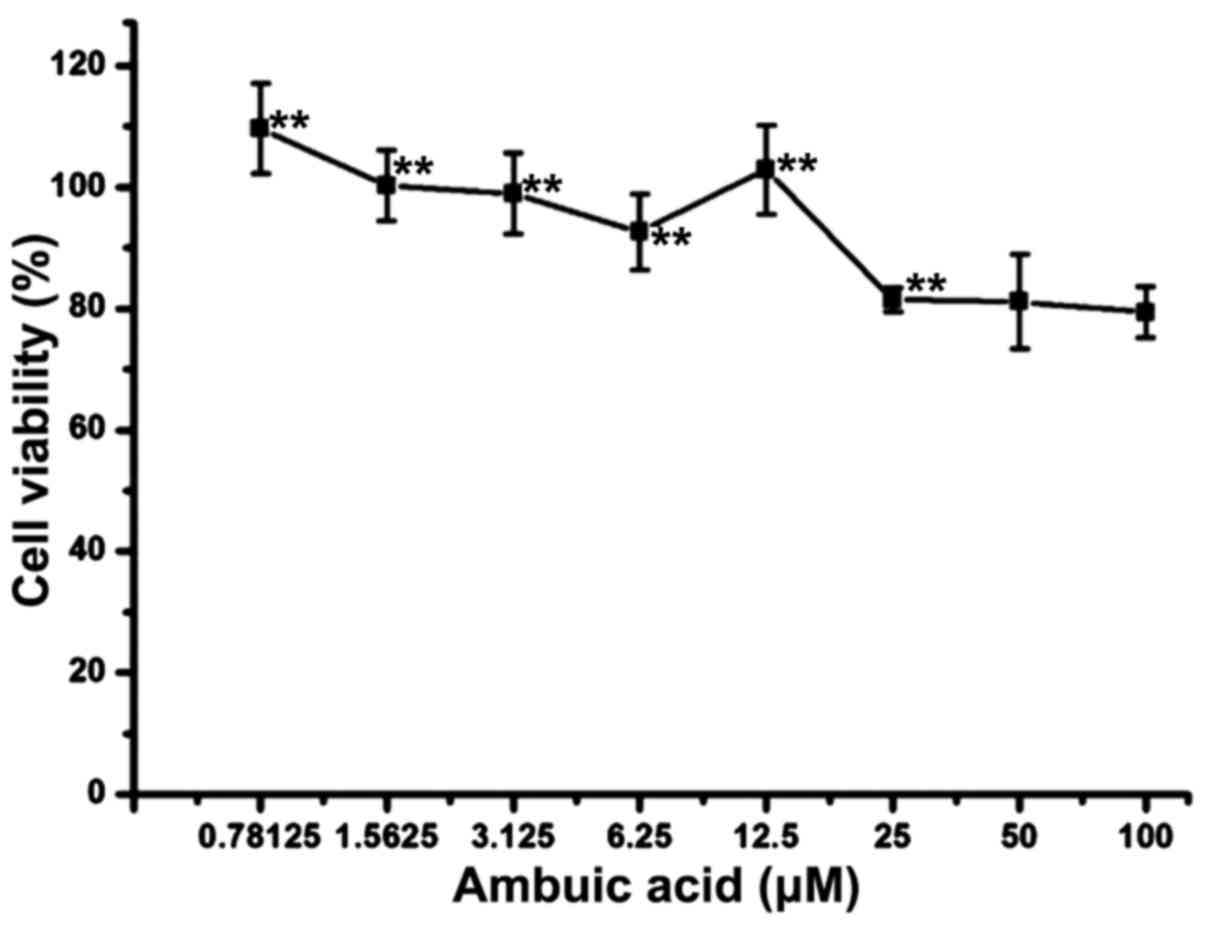
Ambuic acid did not exhibit
cytotoxicity against RAW264.7 macrophages at the concentrations of
0.78–25 µM
RAW264.7 cells were treated by 0.78–100 µM of ambuic
acid for 24 h. MTT assay described in the section of Materials and
methods was used to test the cell viability. As shown in Fig. 2, treatment with 100 µM and 50 µM of
ambuic acid showed weak cytotoxicity against RAW264.7 cells, but no
cytotoxicity was observed at the dose range of 0.78–25 µM.
Effect of ambuic acid on the NO and
PGE2 production
RAW264.7 cells were treated by 1 µg/ml of LPS with
or without ambuic acid (3.125, 6.25, 12.5, 25, 50, 100 µM), or
hydrocortisone succinate (100 µM) for 24 h. The concentration of
nitrite (NO2−) was monitored by Griess assay
as indicator of NO production, and the level of PGE2 was
determined by ELISA. Hydrocortisone succinate was used as a
positive control drug. As shown in Fig.
3A, ambuic acid significantly inhibited the overproduction of
NO induced by LPS in a dose-dependent manner. However, the
production of PGE2 induced by LPS was only weakly
inhibited by ambuic acid (Fig.
3B).
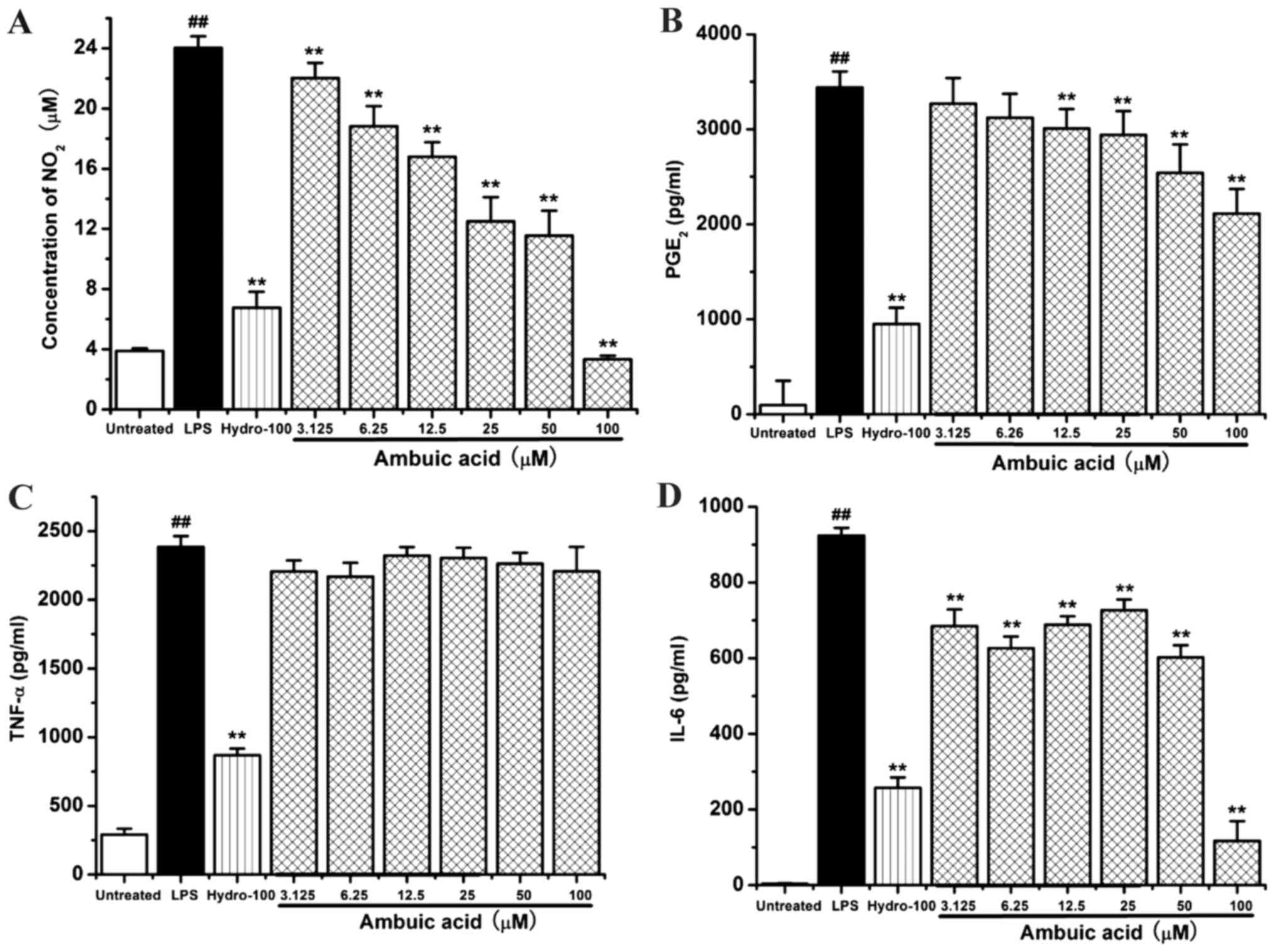 | Figure 3.Effect of ambuic acid on the
overproduction of (A) NO2, (B) PGE2 and the
release of (C) TNF-α, (D) IL-6 in LPS-activated macrophages.
RAW264.7 cells were stimulated by 1 µg/ml LPS with or without
ambuic acid (3.125, 6.25, 12.5, 25, 50 and 100 µM) or
hydrocortisone succinate (100 µM) for 24 h. (A) The nitrite
concentrations in the supernatant were calculated in triplicate by
Griess assay. (B) The levels of PGE2 in the supernatant
were measured in triplicate by ELISA. RAW264.7 cells were
stimulated by 1 µg/ml LPS with or without ambuic acid (3.125, 6.25,
12.5, 25, 50 and 100 µM) or hydrocortisone succinate (100 µM) for 6
h. The levels of (C) TNF-α and (D) IL-6 in the supernatant were
measured in triplicate by respective ELISA kit. Data are presented
as mean ± standard deviation from three separate experiments.
##P<0.01 vs. untreated group, **P<0.01 vs. LPS
treatment group. NO, nitric oxide; PGE2, prostaglandin
E2; TNF-α, tumor necrosis factor-α; IL-6, interleukin-6; LPS,
lipopolysaccharide. |
Effect of ambuic acid on the TNF-α and
IL-6 release
RAW264.7 cells were treated by 1 µg/ml of LPS with
or without ambuic acid (3.125, 6.25, 12.5, 25, 50 and 100 µM), or
hydrocortisone succinate (100 µM) for 6 h. The levels of
pro-inflammatory cytokine TNF-α and IL-6 were measured by using
respective ELISA kit. As shown in Fig.
3C, the positive control group hydrocortisone succinate
potently can effectively inhibit the release of inflammatory factor
TNF-α induced by LPS. However, ambuic acid did not show any
significant inhibitory activity on the release of inflammatory
factor TNF-α. As shown in Fig. 3D,
ambuic acid significantly inhibited the release of IL-6 induced by
LPS in a dose-dependent manner.
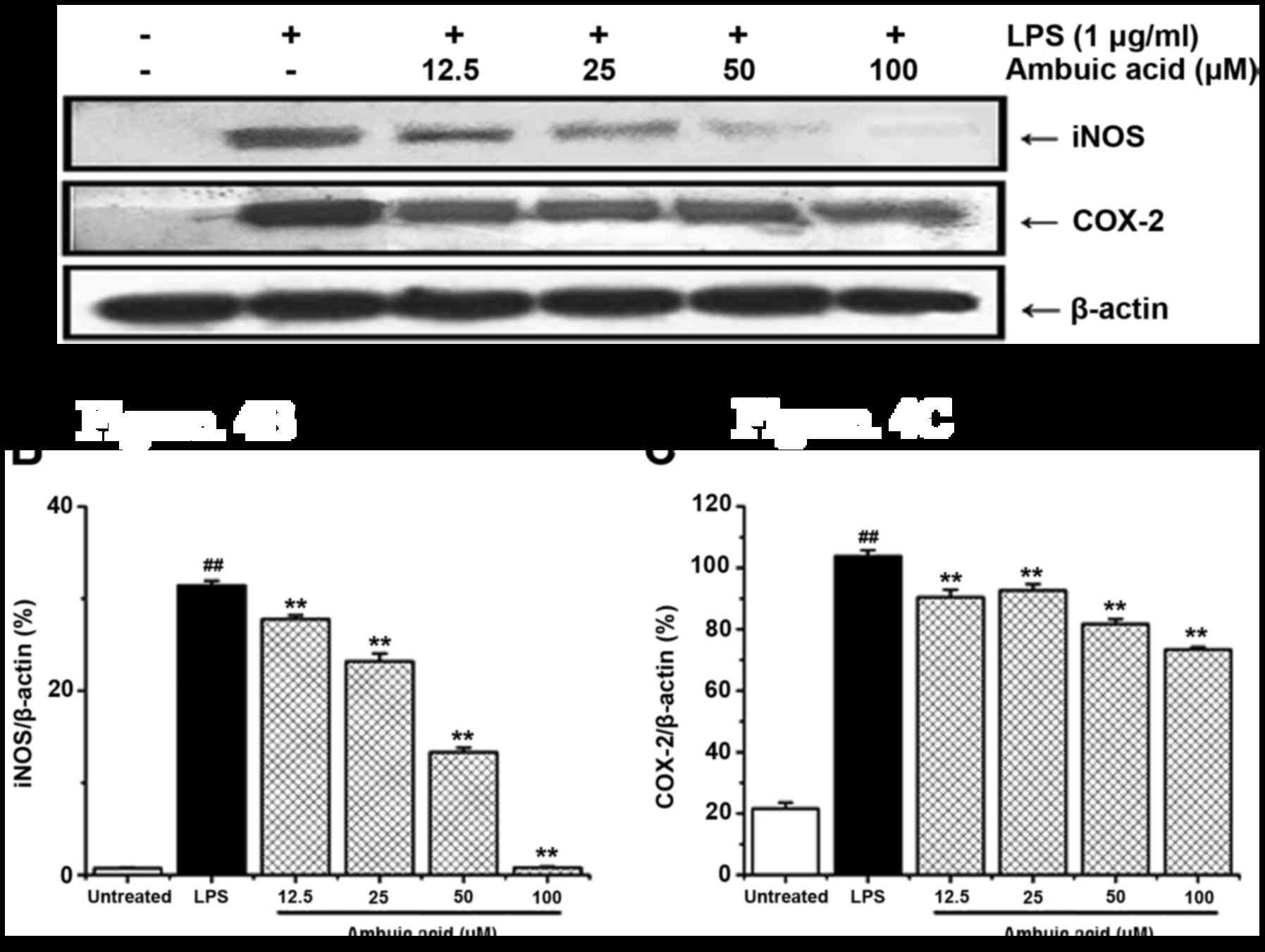
Effect of ambuic acid on the
expression of iNOS and COX-2 proteins
As the overproduction of NO and PGE2 is
always correlated with the high expression of iNOS and COX-2
proteins, and we currently using Western blot analysis to
investigate the expression of iNOS together with COX-2 protein. As
shown in Fig. 4A, the expression of
iNOS and COX-2 proteins was potently increased after stimulation of
LPS for 24 h. Ambuic acid can markedly down regulated the
expression of iNOS protein in a dose-dependent manner, which
strongly suggested that the decrease of NO production was due to
the inhibitory effect of ambuic acid on the suppression of iNOS
expression. Furthermore, ambuic acid also inhibited the expression
of COX-2 protein. The density of bands corresponding to the iNOS
and COX-2 proteins were standardized on the basis of β-actin and
shown in Fig. 4B and C,
respectively.
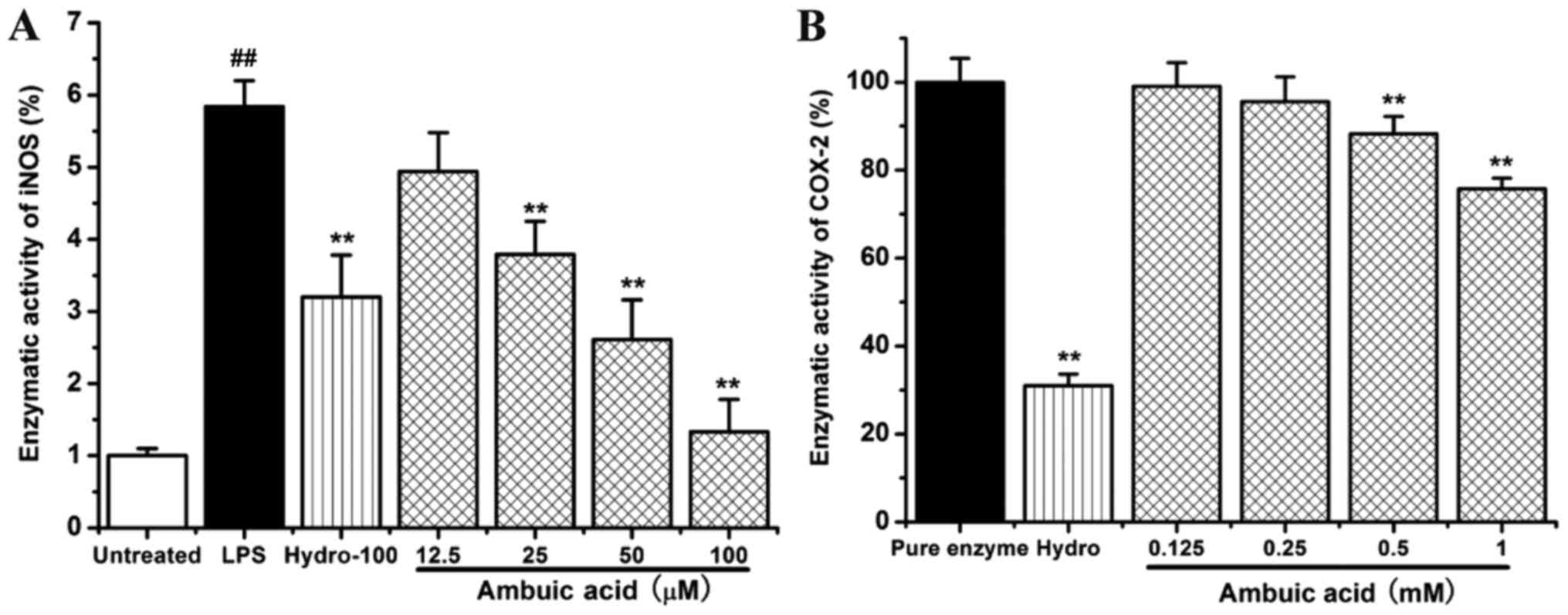
Effect of ambuic acid on the iNOS and
COX-2 enzymatic activity
RAW264.7 cells were treated by 1 µg/ml of LPS with
or without indicated concentrations of ambuic acid (12.5, 25, 50
and 100 µM) or hydrocortisone succinate (100 µM). The inhibitory
effect of ambuic acid at different concentration on the activity of
iNOS enzymatic activity was detected by fluorimetric method. As
shown in Fig. 5A, LPS treatment
caused about a six-fold increase of iNOS enzymatic activity within
120 min. Ambuic acid could significantly inhibit the activity of
iNOS enzymatic induced by LPS in RAW264.7 cells and showed a good
dose-dependency.
The inhibitory effect of ambuic acid on the COX-2
enzymatic activity was examined by the cell-free colorimetric
method. As shown in Fig. 5B, ambuic
acid also inhibited the COX-2 enzymatic activity.
Effect of ambuic acid on the
activation of mitogen-activated protein kinase (MAPK) signaling
pathway
Previous studies have already demonstrated that the
MAPK signaling pathways are associated with the upregulation of
LPS-induced inflammatory responses. LPS activates MAPKs including
the protein kinases ERK 1/2, JNK and p38 kinase. In order to
investigate whether the three MAPK protein pathways are involved in
the anti-inflammatory responses by ambuic acid, LPS-induced
phosphorylation of ERK1/2, JNK and p38 in RAW264.7 cells were
examined by Western blot analysis. As shown in Fig. 6, LPS treatment significantly induced
the phosphorylation of ERK 1/2, JNK and p38. Ambuic acid could
remarkably reduced LPS-induced phosphorylation of ERK1/2 and JNK in
a dose-dependent manner, whereas phosphorylation of p38 was not
affected. The density of bands corresponding to the phospho-ERK,
phospho-JNK and phospho-p38 protein was standardized on the basis
of β-actin and shown in Fig. 6.
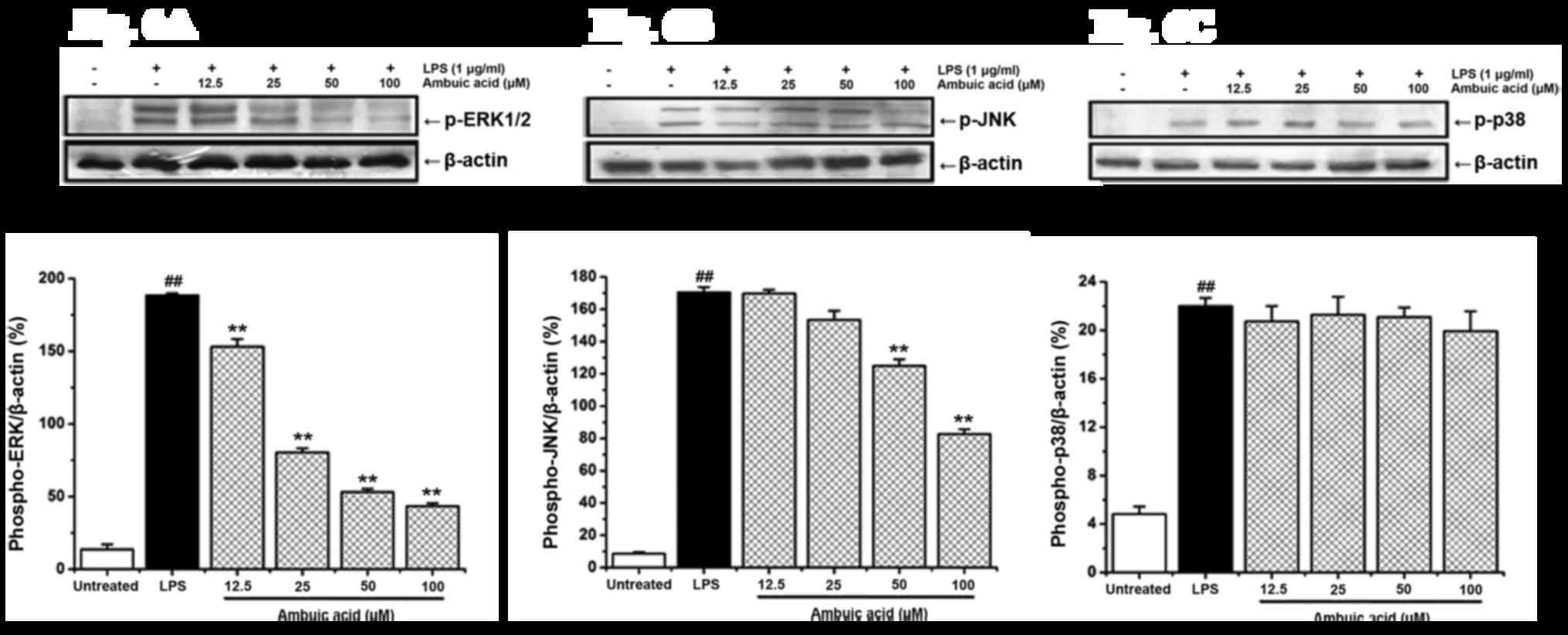 | Figure 6.Effect of ambuic acid on the
phosphorylation of ERK 1/2, JNK and p38 proteins induced by LPS.
RAW264.7 cells were stimulated by LPS (1 µg/ml) with or without
ambuic acid (12.5, 25, 50 and 100 µM) for 30 min, and western blot
analysis was used to investigate the expression of phospho-ERK 1/2,
phospho-JNK and phospho-p38 proteins. To confirm the equal loading
of proteins, the detection of β-actin was carried out.
Densitometric analysis of (A) phospho-ERK 1/2 protein, (B)
phospho-JNK protein and (C) phospho-p38 protein is represented by
mean ± standard deviation of three separate experiments. Data were
standardized on the basis of β-actin levels. ##P<0.01
vs. untreated group; **P<0.01 vs. LPS treatment group. ERK,
extracellular signal-regulated kinase; JNK, c-Jun N-terminal
kinase; LPS, lipopolysaccharide. |
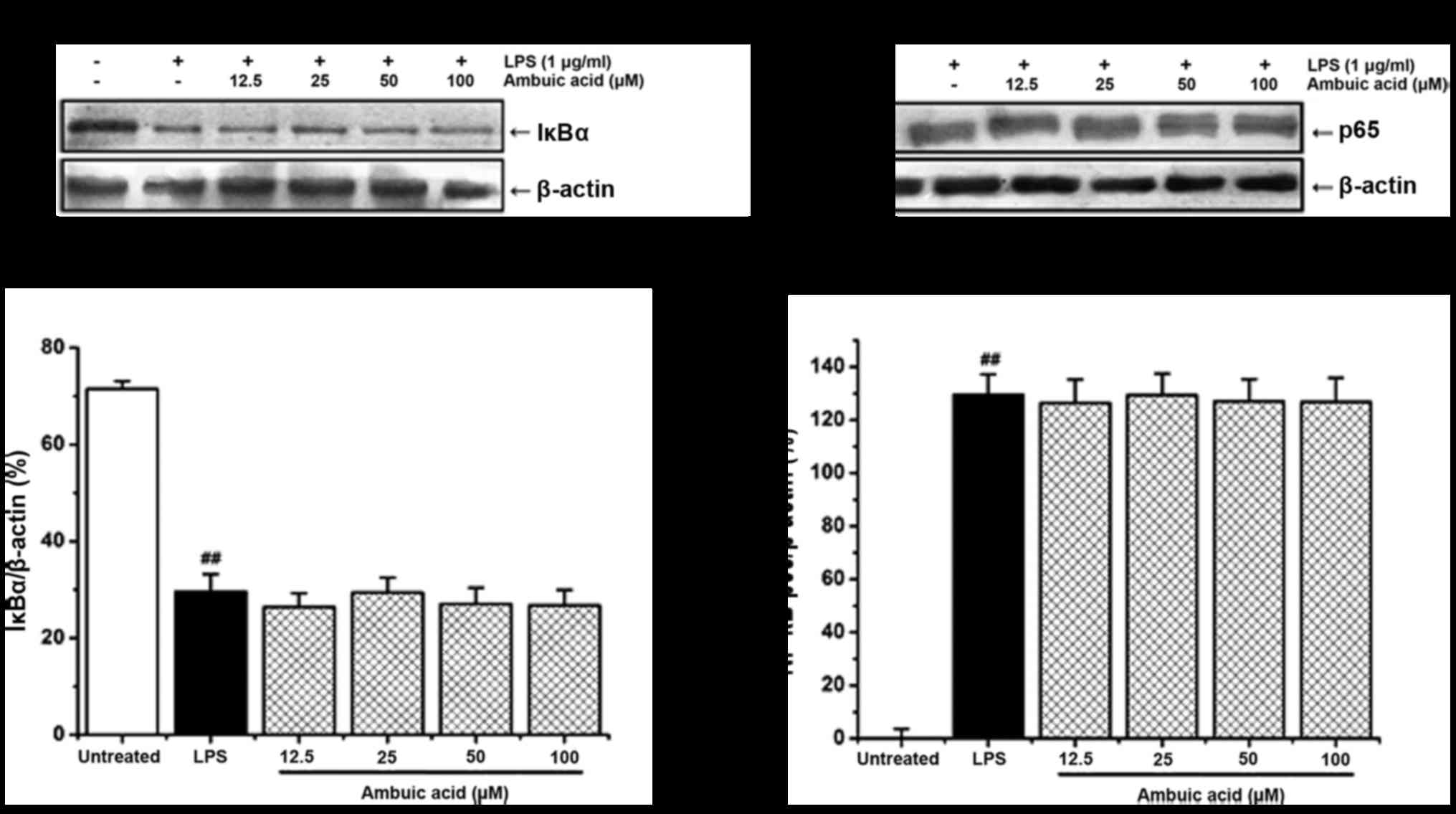
Effect of ambuic acid on the
activation of NF-κB signaling pathway
NF-κB translocated to the nucleus always due to the
phosphorylation and degradation of IκB-α protein. The effect of
ambuic acid on the degradation of IκB-α and the translocation of
NF-κB p65 to the nucleus was investigated by Western blot analysis.
As shown in Fig. 7A and B, LPS
treatment induced the degradation of IκB-α protein and the
translocation of NF-κB p65 to the nucleus. However, ambuic acid
showed no obvious suppression on both the degradation of IkB-α
protein and the translocation of NF-κB p65 to the nucleus.
Discussion
Ambuic acid has been reported to show a certain
inhibitory effect on a variety of plant pathogenic fungi
Fusarium solani, Fusarium cubense, Helminthosporium sativum,
Diplodia natelensis, Cephalosporium gramineum, Pythium ultimum
(24), and it has a certain
antimicrobial activity against the Gram-positive bacterium
Staphyococcus aureus (26).
Ambuic acid can also effect the biosynthesis of cyclic peptide
quormones in Gram-positive bacteria Enterococcus faecalis
(27). Our research group firstly
found that ambuic acid significantly inhibited LPS-induced
overproduction of NO, an important pro-inflammatory mediator that
could promote the expression of some enzymes implicated in diverse
human inflammatory diseases. These results imply the potential
anti-inflammatory effect of ambuic acid and its related
derivatives. The present study was designed to elucidate the
anti-inflammatory effects of ambuic acid, as well as to elucidate
its molecular mechanism.
As shown in the results, ambuic acid strongly
inhibited various LPS-induced responses of macrophages including
the NO overproduction, the PGE2 production, and the
release of pro-inflammatory cytokine IL-6. This is the first direct
evidence to show that the ambuic acid has an anti-inflammatory
effect. To further investigate the possible anti-inflammatory
molecular mechanism of ambuic acid, the effect on the expression of
iNOS and COX-2 proteins, the effect on the enzymatic activity of
iNOS and COX-2 enzymes, the effect on the activation of MAPK
pathway and NF-κB pathway were further investigated.
The Toll like receptor is a superfamily of receptors
that are mainly distributed in inflammatory cells to identify
pathogen molecules, among them, TLR4 mainly identified the LPS that
is the component of gram negative bacteria cell wall. Activation of
myeloid activating factor 88 dependent and non dependent two signal
pathway pathways in combination with LPS and TLR4. The former
activates MAPK and NF-κB signaling pathways, the latter activates
the NF-κB and interferon regulatory factor-3 (IRF3) signaling
pathways. Through these signaling pathways, TLR4 induces
inflammatory cells to release inflammatory factors that mediate
inflammatory responses. RAW264.7 cells have been shown to express
TLR4 in the literature, and LPS is able to recognize TLR4 receptors
on macrophages and activate macrophages to produce inflammatory
factors (39).
MAPK cascades have been shown to be important in the
transduction of extracellular signals to cellular immune responses
(40). In mammalian cells, three
MAPK families have been clearly characterized: classical MAPK, also
known as ERK, JNK/MAPK (JNK/SAPK) and p38 kinase. The MAPK pathways
relay, amplify and integrate signals from a diverse range of
stimuli, and elicit appropriate physiological responses, including
inflammatory responses, cellular proliferation, differentiation and
apoptosis in mammalian cells. LPS can activate the ERK, JNK and p38
MAPK signaling pathways, which can promote the production of
pro-inflammatory cytokines and the expression of iNOS and COX-2
proteins (41,42). Thus, the MAPK signaling pathways are
vital approaches to regulate inflammatory responses, and are also
considered suitable targets for anti-inflammatory therapy (43).
NF-κB plays an important role in many cellular
responses to environmental changes so it is the most valuable
transcription factor. NF-κB complex containing the p50 and the p65
subunits exists in the cytoplasm in a latent form, which is
stabilized by an inhibitory subunit called IκB-α (44,45).
After stimulation of cells with LPS, IκB-α becomes phosphorylated
followed by a rapidly protein degradation. As a result, NF-κB is
activated and translocates into the nucleus promptly.
Our results demonstrated that ambuic acid strongly
inhibited both the expression of iNOS protein and the enzymatic
activity of iNOS enzyme in a dose-dependent manner, which strongly
suggested that the decrease of NO production was due to the
suppression of iNOS expression and enzymatic activity. Ambuic acid
also inhibited the expression of COX-2 protein and the enzymatic
activity of COX-2 enzyme, which indicated that the decrease of
PGE2 production was due to the suppression of COX-2
expression and enzymatic activity. Further investigations showed
that ambuic acid suppressed the phosphorylation of ERK and JNK
proteins, but did not suppress the phosphorylation of p38 protein.
Ambuic acid showed no obvious inhibition on the degradation of
IκB-α protein and the translocation of NF-κB to the nucleus. In
conclusion, ambuic acid exerts anti-inflammatory effect through
inhibiting LPS-induced production of much pro-inflammatory
mediators such as NO, PGE2 and IL-6, and its molecular
mechanism was proved to be via blocking the ERK/JNK MAPK signaling
pathway but has no concern with the inactivation of NF-κB
pathway.
The mechanism of inflammation is that LPS combines
with TLR4 receptors on macrophages to stimulate the release of
inflammatory factors such as NO, TNF-α, IL-6 and PGE2,
and can activate the high expression of upstream protein MAPK
signaling pathway and downstream proteins iNOS and COX-2. In the
experiment, the cells were cultured, and then add the ambuic acid
after a period of incubation, the medium was changed, and the cells
were washed to remove the medium of ambuic acid, then add LPS, it
combines with the TLR4 cell receptor, which can prevent the
reaction of LPS and ambuic acid, and appear the false positive
results.
From the experimental results we can see that ambuic
acid may have mediated changes in the production of inflammatory
cytokines by RAW264.7 cells by influencing the ability of LPS to
bind to TLR4. In the future studies will investigate the role of
TLR4 in the ambuic acid-mediated reduction of pro-inflammatory
cytokines in RAW264.7 cells and investigate the mechanisms
underlying the ambuic acid-mediated reduction in pro-inflammatory
cytokines using specific inhibitors of ERK and JNK.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Ministry of
Science and Technology of China (grant nos. 2014CB138304 and
2012ZX09301002-003), The National Nature Science Foundation (grant
no. 21472233), The Youth Innovation Promotion Association of
Chinese Academy of Sciences (grant no. 2014074) and The Shandong
Provincial Natural Science Foundation (grant no. ZR2016HL54).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HuL cultured the cells and detected the cytotoxicity
of ambuic acid by MTT assay. YL and PL determined the concentration
of nitric oxide and PGE2. LW and DL determined the
concentration of TNF-α and IL-6. MW and QZo analyzed the enzymatic
activity of iNOS and COX-2. Protein extraction was carried out by
KM. The first authors QZh and RL detected the extracted protein by
western blotting, and performed statistical analysis of the data;
they were also major contributors in writing the manuscript. HoL
purified and identified ambuic acid. HoL and FZ conducted the
experimental design and manuscript writing. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liu Y, Yang MH, Wang XB, Li TX and Kong
LY: Caryophyllene sesquiterpenoids from the endophytic fungus,
Pestalotiopsis sp. Fitoterapia. 109:119–124. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Deyrup ST, Swenson DC, Gloer JB and
Wicklow DT: Caryophyllene sesquiterpenoids from a fungicolous
isolate of Pestalotiopsis disseminata. J Nat Prod.
69:608–611. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hwang IH, Swenson DC, Gloer JB and Wicklow
DT: Disseminins and spiciferone analogues: Polyketide-derived
metabolites from a fungicolous isolate of Pestalotiopsis
disseminate. J Nat Prod. 79:523–530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang XL, Awakawa T, Wakimoto T and Abe I:
Induced production of novel prenyldepside and coumarins in
endophytic fungi Pestalotiopsis acaciae. Tetrahedron Lett.
54:5814–5817. 2013. View Article : Google Scholar
|
|
5
|
Klaiklay S, Rukachaisirikul V, Tadpetch K,
Sukpondma Y, Phongpaichit S, Buatong J and Sakayaroj J: Chlorinated
chromone and diphenyl ether derivatives from the mangrove-derived
fungus Pestalotiopsis sp. PSU-MA69. Tetrahedron.
68:2299–2305. 2012. View Article : Google Scholar
|
|
6
|
Li J, Li L, Si Y, Jiang X, Guo L and Che
Y: Virgatolides A-C, benzannulated spiroketals from the plant
endophytic funfus Pestalotiopsis virgatula. Org Lett.
13:2670–2673. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu J, Kjer J, Sendker J, Wray V, Guan H,
Edrada R, Lin W, Wu J and Proksch P: Chromones from the endophytic
fungus Pestalotiopsis sp. isolated from the chinese mangrove
plant Rhizophora mucronata. J Nat Prod. 72:662–665. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Harper JK, Arif AM, Ford EJ, Strobel GA,
Porco JA Jr, Tomer DP, Oneill KL, Heider WM and Grant DM: Pestacin:
A 1,3-dihydro isobenzofuran from Pestaliopsis microspora possessing
antioxidant and antimycotic activities. Tetrahedron. 59:2471–2474.
2003. View Article : Google Scholar
|
|
9
|
Akone SH, Amrani ME, Lin W, Lai D and
Proksch P: Cytosporins F-K, new epoxyquinols from the endophytic
fungus Pestalotiopsis theae. Tetrahedron Lett. 54:6751–6754.
2013. View Article : Google Scholar
|
|
10
|
Xu J, Aly AH, Wray V and Proksch P:
Polyketide derivatives of endophytic fungus Pestalotiopsis
sp. isolated from the Chinese mangrove plant Rhizophora
mucronata. Tetrahedron Lett. 52:21–25. 2010. View Article : Google Scholar
|
|
11
|
Strobel G, Yang X, Sears J, Kramer R,
Sidhu RS and Hess WM: Taxol from Pestalotiopsis microspora,
an endophytic fungus of Taxus wallachiana. Microbiology.
142:435–440. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kumaran RS, Kim HJ and Hur BK:
Taxol-producing [corrected] fungal endophyte, Pestalotiopsis
species isolated from Taxus cuspidata. J Biosci Bioeng.
110:541–546. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tan RX and Zou WX: Endophytes: A rich
source of functional metabolites. Nat Prod Rep. 18:448–459. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding G, Liu S, Guo L, Zhou Y and Che Y:
Antifungal metabolites from the plant endophytic fungus
Pestalotiopsis foedan. J Nat Prod. 71:615–618. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei MY, Li D, Shao CL, Deng DS and Wang
CY: (±)-Pestalachloride D, an antibacterial racemate of chlorinated
benzophenone derivative from a soft coral-derived fungus
Pestalotiopsis sp. Mar Drugs. 11:1050–1060. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu D, Zhang BY and Yang XL: Antifungal
monoterpene derivatives from the plant endophytic fungus
Pestalotiopsis foedan. Chem Biodivers. 13:1422–1425. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia YL, Wei MY, Chen HY, Guan FF, Wang CY
and Shao CL: (+)- and (−)-pestaloxazine A, a pair of antiviral
enantiomeric alkaloid dimers with a symmetric
spiro[oxazinane-piperazinedione] skeleton from Pestalotiopsis
sp. Org Lett. 17:4216–4219. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen L, Zhang QY, Jia M, Ming QL, Yue W,
Rahman K, Qin LP and Han T: Endophytic fungi with antitumor
activities: Their occurrence and anticancer compounds. Crit Rev
Microbiol. 42:454–473. 2016.PubMed/NCBI
|
|
19
|
Kiho T, Itahashi S, Sakushima M, Matsunaga
T, Usui S, Ukai S, Mori H, Sakamoto H and Ishiguro Y:
Polysaccharides in fungi. XXXVIII. Anti-diabetic activity and
structural feature of a galactomannan elaborated by
Pestalotiopsis species. Biol Pharm Bull. 20:118–121. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kumar DS, Lau CS, Wan JM, Yang D and Hyde
KD: Immunomodulatory compounds from Pestalotiopsis
leucothës, an endophytic fungus from Tripterygium wilfordii.
Life Sci. 78:147–156. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Verma VC, Gangwar M, Yashpal M and Nath G:
Anticestodal activity of endophytic Pestalotiopsis sp. on
protoscoleces of hydatid cyst Echinococcus granulosus. Biomed Res
Int. 2013:3085152013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tejesvi MV, Kini KR, Prakash HS, Subbiah V
and Shetty HS: Antioxidant, antihypertensive, and antibacterial
properties of endophytic Pestalotiopsis species from
medicinal plants. Can J Microbiol. 54:769–780. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xia X, Kim S, Liu C and Shim SH: Secondary
metabolites produced by an endophytic fungus Pestalotiopsis
sydowiana and their 20S proteasome inhibitory activities.
Molecules. 21:pii: E944. 2016. View Article : Google Scholar
|
|
24
|
Li JY, Harper JK, Grant DM, Tombe BO,
Bashyal B, Hess WM and Strobel GA: Ambuic acid, a highly
functionalized cyclohexenone with antifungal activity from
Pestalotiopsis spp. and Monochaetia sp.
Phytochemistry. 56:463–468. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harper JK, Barich DH, Hu JZ, Strobel GA
and Grant DM: Stereochemical analysis by solid-state NMR:
Structural predictions in ambuic acid. J Org Chem. 68:4609–4614.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ding G, Li Y, Fu S, Liu S, Wei J and Che
Y: Ambuic acid and torreyanin acid derivatives from the
endolichenic fungus Pestalotiopsis sp. J Nat Prod.
72:182–186. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakayama J, Uemura Y, Nishiguchi K,
Yoshimura N, Igarashi Y and Sonomoto K: Ambuic acid inhibits the
biosynthesis of cyclic peptide quormones in gram-positive bacteria.
Antimicrob Agents Chemother. 53:580–586. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qi QY, Li EW, Han JJ, Pei YF, Ma K, Bao L,
Huang Y, Zhao F and Liu HW: New ambuic acid derivatives from the
solid culture of Pestalotiopsis neglecta and their nitric
oxide inhibitory activity. Sci Rep. 5:99582015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ali AM, Habeeb RA, El-Azizi NO, Khattab
DA, Abo-Shady RA and Elkabarity RH: Higher nitric oxide levels are
associated with disease activity in Egyptian rheumatoid arthritis
patients. Rev Bras Reumatol. 54:446–451. 2014.(In Portuguese).
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Coulter JA, McCarthy HO, Xiang J, Roedl W,
Wagner E, Robson T and Hirst DG: Nitric oxide-a novel therapeutic
for cancer. Nitric Oxide. 19:192–198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Husain K, Hernandez W, Ansari RA and
Ferder L: Inflammation, oxidative stress and rennin angiotensin
system in atherosclerosis. World J Biol Chem. 6:209–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Beyazit Y, Efe C, Tanoglu A, Purnak T,
Sayilir A, Taskıran I, Kekilli M, Turhan T, Ozaslan E and Wahlin S:
Nitric oxide is a potential mediator of hepatic inflammation and
fibrogenesis in autoimmune hepatitis. Scand J Gastroenterol.
50:204–210. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Muscará MN and Wallace JL: Nitric Oxide.
V. therapeutic potential of nitric oxide donors and inhibitors. Am
J Physiol. 276:G1313–G1316. 1999.PubMed/NCBI
|
|
34
|
Denizot F and Lang R: Rapid colorimetric
assay for cell growth and survival. Modifications to the
tetrazolium dye procedure giving improved sensitivity and
reliability. J Immunol Methods. 89:271–277. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao F, Chen L, Zhang M, Bi C, Li L, Zhang
Q, Shi C, Li M, Zhou S and Kong L: Inhibition of
lipopolysaccharide-induced iNOS and COX-2 expression by indole
alkaloid,
3-(hydroxy-methyl)-6,7-dihydroindolo[2,3-a]quinolizin-(12H)-one,
via NF-κB inactivation in RAW 264.7 macrophages. Planta Med.
79:782–787. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wohlmuth H, Deseo MA, Brushett DJ,
Thompson DR, Macfarlane G, Stevenson LM and Leach DN:
Diarylheptanoid from Pleuranthodium racemigerum with in vitro
prostaglandin E(2) inhibitory and cytotoxic activity. J Nat Prod.
73:743–746. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao F, Xu H, He EQ, Jiang YT and Liu K:
Inhibitory effects of sesquiterpenes from Saussurea lappa on the
overproduction of nitric oxide and TNF-alpha release in
LPS-activated macrophages. J Asian Nat Prod Res. 10:1045–1053.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao F, Wang L and Liu K: In vitro
anti-inflammatory effects of arctigenin, a lignan from Arctium
lappa L., through inhibition on iNOS pathway. J Ethnopharmacol.
122:457–462. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Murshid A, Gong J, Prince T, Borges TJ and
Calderwood SK: Scavenger receptor SREC-I mediated entry of TLR4
into Lipid microdomains and triggered inflammatory cytokine release
in RAW 264.7 Cells upon LPS activation. PLoS One. 10:e01225292015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu Y, Shepherd EG and Nelin LD: MAPK
phosphatases-regulating the immune response. Nat Rev Immunol.
7:202–212. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Arthur JS and Ley SC: Mitogen-activated
protein kinases in innate immunity. Nat Rev Immunol. 13:679–692.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rao KM: MAP kinase activation in
macrophages. J Leukoc Biol. 69:3–10. 2001.PubMed/NCBI
|
|
43
|
Kaminska B: MAPK signalling pathways as
molecular targets for anti-inflammatory therapy-from molecular
mechanisms to therapeutic benefits. Biochim Biophys Acta.
1754:253–262. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Baeuerle PA and Baltimore D: I kappa B: A
specific inhibitor of the NF-kappa B transcription factor. Science.
242:540–546. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Beg AA and Baldwin AS Jr: The I kappa B
proteins: Multifunctional regulators of Rel/NF-kappa B
transcription factors. Genes Dev. 7:2064–2070. 1993. View Article : Google Scholar : PubMed/NCBI
|