Introduction
Lung cancer is a respiratory disease that is
responsible for the highest rates of cancer-associated mortality
and air contamination worldwide (1).
Small cell lung cancer (SCLC) and non-small cell lung cancer
(NSCLC) are two main types of human lung cancer that are
characterized by tumor morphology (2), and they account for ~95% of all lung
cancer (3). NSCLC which has the
highest incidence among all kinds of cancer originates from
non-small cells in the lungs (4).
NSCLC, which is also the most frequent type of lung cancer, can be
divided into squamous cell carcinoma, large cell carcinoma and
adenocarcinoma determined by tumor cell genetics. According to the
clinical statistics investigations of lung cancer cases >80% of
newly diagnosed NSCLC patients were in middle or severe stage
(5–7).
Though increasing research has endeavored to improve
the efficacy of treatment for patients with NSCLC, the survival
rate remained poor, with <15% survival observed in the 5 years
after clinical treatment (6,8,9). In
addition, the majority of newly diagnosed patients with NSCLC are
in the advanced stage. Previous research has reported that
migration and invasion in NSCLC are predominantly responsible for
the poor survival rate during treatment and recurrence for patients
with NSCLC (10,11). As a result, the exploration for
effective agents for the inhibition of migration and invasion has
become critically important for the treatment of cancer patients
(12,13). The present study investigated H358
NSCLC cell migration and invasion. Furthermore, the inhibitory
effects of anti-cysteine-rich angiogenic inducer-61 (CYR-61) on
H358 NSCLC cell migration and invasion were analyzed in
vitro and in vivo.
CYR-61 is a member of the CYR-61/connective tissue
growth factor/nephroblastoma overexpressed (CCN) protein family
(14). Previous studies have
demonstrated that CYR-61 promotes human lung cancer cell migration
and metastasis and it is closely related to patient prognosis in
NSCLC (15,16). In addition, CYR-61 is involved in
tumor cell mitogenesis, cellular adhesion, migration,
differentiation, wound healing, angiogenesis and survival (17). Previous reports have demonstrated the
important roles of CYR-61 in cancer development and metastasis,
indicating that CYR-61 may be an important target for gene therapy
and tumor suppression (16, 17). In addition, CYR61 has been
demonstrated to induce angiogenesis by supplying oxygen for tumor
cells during proliferation (18).
Many studies have focused on the functions of the
CCN protein family in cancer biology (17,19). A
study by Barnett et al (7)
reported that CYR-61 demonstrated potential as an oncogene or a
tumor suppressor, depending on tumor cell type. Clinically,
expression of CYR-61 has been associated with the prognosis of
breast cancer and prostate cancer (20). However, few studies have investigated
the function of CYR-61 in NSCLC. Therefore, the present study
investigated the expression of CYR-61 in NSCLC cells and tumors.
Results indicated that CYR-61 was expressed at higher levels in
NSCLC cells, when compared with normal lung cells of MRC-5.
Furthermore, an antibody against CYR-61 (anti-CYR-61) was
constructed and its therapeutic effects in mice with NSCLC were
investigated.
Recently, numerous studies have indicated that
mechanistic target of rapamycin (mTOR) may regulate tumor cell
growth, migration and cancer metastasis (21,22).
Epithelial-mesenchymal transition (EMT) has an essential role in
tumor growth, migration and cancer metastasis. In addition, the EMT
process reduces tumor cell adhesion and results in tumor cells
gaining migratory and invasive properties through cell-cell
connections (23). Previous research
has indicated that CYR-61 is associated with NSCLC migration and
cancer metastasis (17). However,
little is known about the signaling mechanisms regulating mTOR,
CYR-61 and EMT in NSCLC. Therefore, the present study examined the
association between CYR-61 and EMT in NSCLC cells. EMT biomarker
expression levels of vimentin, fibronectin, α-smooth muscle actin
(SMA) and N-cadherin were analyzed. Mitogen-activated protein
kinase (MAPK) and phosphoinositide 3-kinase (PI3K)/protein kinase B
(AKT)/mTOR signaling pathways in EMT were also investigated in
vitro and in vivo in NSCLC cells and tissues,
respectively.
The aim of the present study was to determine the
effects of anti-CYR-61 on CYR-61-associated invasion and metastasis
in NSCLC through MAPK/EMT signaling pathways. It was concluded that
CYR-61 may be considered as a potential prognostic biomarker for
NSCLC, and anti-CYR-61 may provide a potential minimally invasive
therapy for NSCLC.
Materials and methods
Ethics statement
The present study was carried out in strict
accordance with the approval and recommendations from the Ethics
Committee of the Care and Use of Laboratory Animals of Qilu
Hospital of Shandong University (Jinan, China). All surgery and
euthanasia were performed under sodium pentobarbital anesthesia,
and all efforts were made to minimize suffering.
Cell culture
The H358 NSCLC cell line and MRC-5 normal lung cell
line were purchased from American Type Culture Collection
(Manassas, VA, USA). The cell lines were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) at 37°C, 5% CO2 and 100%
humidity.
Construction of full-length
anti-CYR-61 antibody
A mouse anti-human CYR-61 monoclonal antibody was
constructed using a conventional approach and screened by
fluorescence-activated cell sorting (FACS). The full length of the
anti-CYR-61 antibody was constructed, as previously described
(24). The single chain variable
fragments of the mouse anti-human CYR-61 monoclonal antibody (Sino
Biological, Beijing, China) were cloned and inserted into a Pklight
vector (termed Pklight-anti-CYR-61 vector; Biovector NTCC, Inc.,
Beijing, China). The constant domain heavy chain Fc and light chain
fragments of mouse anti-human CYR-61 monoclonal antibody were
subcloned into the Pklight-anti-CYR-61 vector. Pklight-anti-human
CYR-61 monoclonal antibody and IREX-enhanced green fluorescent
protein (EGFP) were subcloned into the Peedual12.4 vector
(BioVector NTCC, Inc.), which contained the glutamine synthetase
gene. The CHO-K1SV cell line (American Type Culture Collection) was
cultured in Iscove's modified Dulbecco's medium (Sigma-Aldrich;
Merck KGaA) supplemented with 10% fetal bovine serum and 2 mM
L-glutamine. Peedual12.4-anti-human CYR-61 monoclonal antibody was
transfected into the fluorescein isothiocyanate (Shanghai Xinyu
Biotechnology Pharmaceutical Co., Ltd.)-labeled CHO-K1SV cells
(1×105 cells/ml) using Lipofectamine 2000 (Tiangen
Biotech Co., Ltd., Beijing, China). CHO-K1SV cells were washed and
resuspended in 0.01 mol/l pH=7.4 PBS twice. The cells were
identified and sorted in a flow cytometer at 488 nm. Thus,
anti-human CYR-61 monoclonal antibody and EGFP were stably
expressed in the CHO-K1SV cells. Stable and high expression of
anti-CYR-61 antibodies in bacterial cells was screened using FACS
(BD Biosciences, Franklin Lakes, NJ, USA). Data analysis and
statistics were performed using BD Accuri™ C6 Plus (BD
Biosciences) and GraphPad Prism 5 (GraphPad Software, Inc., La
Jolla, CA, USA).
MTT assay
A total of 3,000 of H358 cells were cultured into
each well of a 96-well plate and cells were used to investigate the
inhibitory effects of anti-CYR-61 on cell viability when ~90% cell
confluence was reached. Anti-CYR-61 (1:1,000) or 30 µl PBS
(control) were added into each well of the 96-well plate and
incubated at 37°C for 12 h. Subsequently, 10 µl MTT (5 mg/ml;
Sigma-Aldrich; Merck KGaA) was added to the cells and incubated at
37°C for 4 h. Following this, dimethylsulfoxide (Amresco, LLC,
Solon, OH, USA) was added for incubation for 30 min to dissolve the
precipitate after the supernatant had been removed. Results were
determined using a spectrophotometer (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) at 540 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from H358 cells and a normal
lung cell line MRC-5 cells with or without treatment of anti-CYR-61
using an RNeasy mini kit (Qiagen Sciences, Inc., Gaithersburg, MD,
USA), according to the manufacturer's protocol. RNA (1.0 µg) was
reverse transcribed into cDNA using QuantiTect Reverse
Transcription kit (Qiagen Sciences, Inc.), according to the
manufacturer's protocol. The primers (Table I) were designed using Primer Express
software (version 2.0; Thermo Fisher Scientific, Inc.) qPCR
analysis was performed using the SYBR®
Premix™ Ex Taq™ (Perfect Real
Time; Takara Biotechnology Co., Ltd., Dalian, China) in a total
volume of 20 µl using a 7300 Real-Time PCR System (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
thermocycling conditions were as follows: 95°C for 30 sec, and 40
cycles of 95°C for 5 sec and 60°C for 30 sec. All relative mRNA
expression levels were calculated using the by 2−ΔΔCq
method (25). Results were expressed
as the n-fold relative to the housekeeping gene, β-actin.
 | Table I.Primer sequences utilized in
RT-qPCR. |
Table I.
Primer sequences utilized in
RT-qPCR.
| Gene | Primer | Sequence |
|---|
| CYR-61 | Forward |
5′-CAAGGAGCTGGGATTCGATG-3′ |
|
| Reverse |
5′-AAAGGGTTGTATAGGATGCGAG-3′ |
| Bak | Forward |
5′-CACCTTACCTCTGCAACCTAG-3′ |
|
| Reverse |
5′-TGCAACATGGTCTGGAACTC-3′ |
| Bax | Forward |
5′-AGTAACATGGAGCTGCAGAG-3′ |
|
| Reverse |
5′-AGTAGAAAAGGGCGACAACC-3′ |
| Caspase-9 | Forward |
5′-GTTTGAGGACCTTCGACCAG-3′ |
|
| Reverse |
5′-GCATTAGCGACCCTAAGCAG-3′ |
| Apaf-1 | Forward |
5′-CCTCTCATTTGCTGATGTCG-3′ |
|
| Reverse |
5′-TCACTGCAGATTTTCACCAGA-3′ |
| Caspase-10 | Forward |
5′-AATCTGACATGCCTGGAG-3′ |
|
| Reverse |
5′-ACTCGGCTTCCTTGTCTAC-3′ |
| Caspase-8 | Forward |
5′-ATGCAAACTGGATGATGACA-3′ |
|
| Reverse |
5′-TTCATATCTTCAGCAGGTCT-3′ |
| FasL | Forward |
5′-AACCAAGTGGACCTTGAGACCACA-3′ |
|
| Reverse |
5′-TTCACATGGCAGCCCAGAGTTCTA-3′ |
| FADD | Forward |
5′-CCTGGTACAAGAGGTTCAGC-3′ |
|
| Reverse |
5′-CTGTGTAGATGCCTGTGGTC-3′ |
| β-actin | Forward |
5′-ACCTTCTACAATGAGCTGCG-3′ |
|
| Reverse |
5′-CCTGGATAGCAACGTACATGG-3′ |
ELISA
The affinity of anti-CYR-61 was examined for its
target antigens, CYR-61, using ELISA in H358 cells. CYR-61 (0.2–1.4
mg/ml) was added into an enzyme-linked-immuno microplate and
incubated at 4°C for 12 h. Anti-CYR-61 (4 µg/ml) was added to the
wells and incubated for 60 min at 37°C, and bovine serum albumin
(BSA; Atlanta Biologicals, Inc., Flowery Branch, GA, USA) was used
as a control. Subsequently, 100 µl human horseradish
peroxidase-conjugated CYR-61 antibodies (1:1,000; eBioscience,
Inc.; Thermo Fisher Scientific, Inc.) was added and incubated at
37°C for 60 min. The
3,3′-diaminobenzidine/H2O2 system was used
for the detection of anti-CYR-61 affinity. Results were analyzed at
450 nm using an ELISA plate reader (Bio-Rad Laboratories,
Inc.).
Cell invasion and migration
assays
H358 cells were treated with anti-CYR-61 and
non-treated H358 cells were used as control. H358 cells were
adjusted to a density of 1×106 cells in 500 µl
serum-free RPMI-1640 medium for the invasion assay. H358 cells were
treated with anti-CYR-61 (1:1,000) for 12 h at room temperature and
then added to the tops of BD BioCoat Matrigel Invasion Chambers (BD
Biosciences), according to the manufacturer's protocol. Transwell
chambers (Costar; Corning Incorporated, Corning, NY, USA) with 8 µm
diameter pores were utilized. Matrigel (100 µl; BD Biosciences, San
Jose, CA, USA) was added to the Transwell apparatus. A total of 106
H358 cells in 100 µl serum-free culture medium (Gibco; Thermo
Fisher Scientific, Inc.) was placed in the upper chamber and 500 µl
complete culture medium containing 20% fetal bovine serum
(Sigma-Aldrich; Merck KGaA) was added to the lower chamber as a
chemoattractant. Following incubation at 37°C for 24 h, Transwell
chambers were stained with 0.4% crystal violet for 5 min at room
temperature and washed with PBS three times. Non-migrating and
non-invading cells were carefully wiped from the upper chambers
with cotton wool. Results were examined using a CX21 Olympus light
microscope (Olympus Corporation, Tokyo, Japan; magnification,
×100).
For the migration assay, H358 (1×104)
cells were inoculated with anti-CYR-61 (1:1,000) for 12 h at room
temperature and Control inserts (BD Biosciences) were used instead
of a Matrigel Invasion Chamber. Tumor cell invasion and migration
were observed in at least three stained fields in every membrane
using a light microscope at a magnification of ×100. The invasion
and migration assays used 24 well dishes.
Apoptosis assay
Apoptosis was determined by staining H358 cells with
Annexin V-PE/ and 7-aminoactinomycin (BD Biosciences) for 15 min at
25°C in the dark, and flow cytometry analysis was performed. The
Annexin V-positive cells were counted as early apoptotic cells.
7-amino actinomycin-positive cells were counted as necrotic cells.
Double positive cells were counted as late apoptotic cells. Double
negative cells were counted as live cells.
Animal study
A total of 50 female specific pathogen-free C57BL/6
mice (aged 6–8 weeks; weighing 20±2 g) were purchased from Shanghai
Slack Experimental Animals Co., Ltd., (Shanghai, China). The mice
were given free accessible to food and water, and were housed at
20°C with 60% humidity and 12 h light/dark cycle. C57BL/6 mice were
subcutaneously implanted with H358 tumor cells and were divided
into two groups (20 per group). Treatments were initiated on day 5
after tumor implantation when the tumor diameter had reached 6–8
mm. H358-bearing mice were intravenously injected with anti-CYR-61
or 10 µl PBS as a control. The treatment was administered once
daily for 14 days. Tumor volumes were calculated according to
previous study (26). The survival
rate for animals treated with anti-CYR-61 (1:1,000) or 10 µl PBS
was detected during a 120-day observation period.
Western blotting
Total protein was extracted using
CytoBuster™ Protein Extraction Reagent (EMD Millipore,
Billerica, MA, USA) and protein concentration was measured by the
DC™ Protein Assay (Bio-Rad Laboratories, Inc.)
using the Bradford method (27). A
total of 40 µg of protein from each sample were separated by 10%
SDS-PAGE and transferred to an equilibrated polyvinylidene
difluoride membrane (GE Healthcare, Chicago, IL, USA). The membrane
was blocked by 5% milk in 0.1% Tween 20 in Tris-buffer solution
(TBS) at room temperature for 1 h. After incubation with specific
primary antibody at 4°C overnight and the membranes were washed
four times with TBS, 10 min each time. The membranes were then
incubated with horseradish peroxidase-conjugated secondary
antibodies (1:800; cat. no. ab7090; Abcam, Cambridge, MA, USA) at
37°C for 30 min and the proteins were detected by enhanced
chemiluminescence (GE Healthcare) and quantified using an image
analyzer Quantity One System (Bio-Rad Laboratories, Inc.). The
primary antibodies directed against CYR-61 (cat. no. ab24448),
extracellular signal-regulated kinase (cat. no. ab54230), AKT (cat.
no. ab8805), phosphorylated (p)ERK (cat. no. ab79483), pAKT (cat.
no. ab8933), fibronectin (cat. no. ab23750), SMA (cat. no. ab21027)
and N-cadherin (cat. no. ab98952) were purchased from Abcam
(Cambridge, UK) all at a dilution of 1:500 and were incubated at
4°C overnight. Primary antibodies directed against Caspase 8
(1;800; cat. no, sc81656), 9 (1;800; cat. no. sc56076) and 10
(1;800; cat. no. sc134299), Fas ligand (FasL; 1;800; cat. no.
sc33716), Fas-associated protein with Death Domain (FADD; 1:800;
cat. no. sc5559), apoptotic protease activating factor 1 (Apaf-1;
1:800; cat. no. sc135836), B cell lymphoma-2 antagonist/killer
(Bak; 1:800; cat. no. sc517390), B cell lymphoma-2-associated X
protein (Bax; 1:800; cat. no. sc20067), vimentin (1:800; cat. no.
sc80975) were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). The housekeeping proteins β-actin (1:500; cat.
no. 3700) and GAPDH (1:500; cat. no. 97166) were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA). Density
analysis was performed using Image J v.1.49 (National Institutes of
Health, Bethesda, MD, USA).
Histological immunostaining
For immunostaining, H358 cells or tumors from
xenograft mice with NSCLC were fixed using 10% formaldehyde at room
temperature for 24 h and subsequently embedded in paraffin.
Following this, tumor samples were sliced into 4-µm-thick sections
and antigen retrieval was also performed with 0.1 M citrate buffer
for 15 min at 100°C, rehydrated in a descending series of ethanol
(100, 95, 85, 80 and 75%) watched in xylene in tumor sections.
After blocking by 10% BSA at room temperature for 1 h, H358 cells
and tumor sections were incubated with pERK (cat. no. ab192591),
pAKT (cat. no. ab38449) and CYR-61 (cat. no. ab24448; all 1:600;
Abcam) primary antibodies at 37°C for 2 h. Following washing three
times with TBST, HRP-conjugated goat anti-rabbit Immunoglobulin G
secondary antibodies (1:800; cat. no. ab97051; Abcam) were
incubated 30 min at 37°C. Specimens were then visualized using a
binocular light microscope (Eclipse E100-LED; Nikon Corporation,
Tokyo, Japan) at a magnification of ×200. A Ventana Benchmark
automated staining system (Ventana Medical Systems, Inc., Tucson,
AZ, USA) was used for observation of CYR-61.
Statistical analysis
All data were presented as the mean ± standard
deviation of triplicates. Unpaired data were compared using
Student's t-tests and comparisons of data between multiple groups
were made using one-way analysis of variance with a post-hoc
Tukey's test using SPSS 17.0 software (SPSS, Inc., Chicago, IL,
USA). Kaplan-Meier tests were used to estimate the survival rate
during 120-day long-term treatment. P<0.05 was considered to
indicate a statistically significant difference.
Results
CYR-61 expression and anti-CYR-61
characteristics
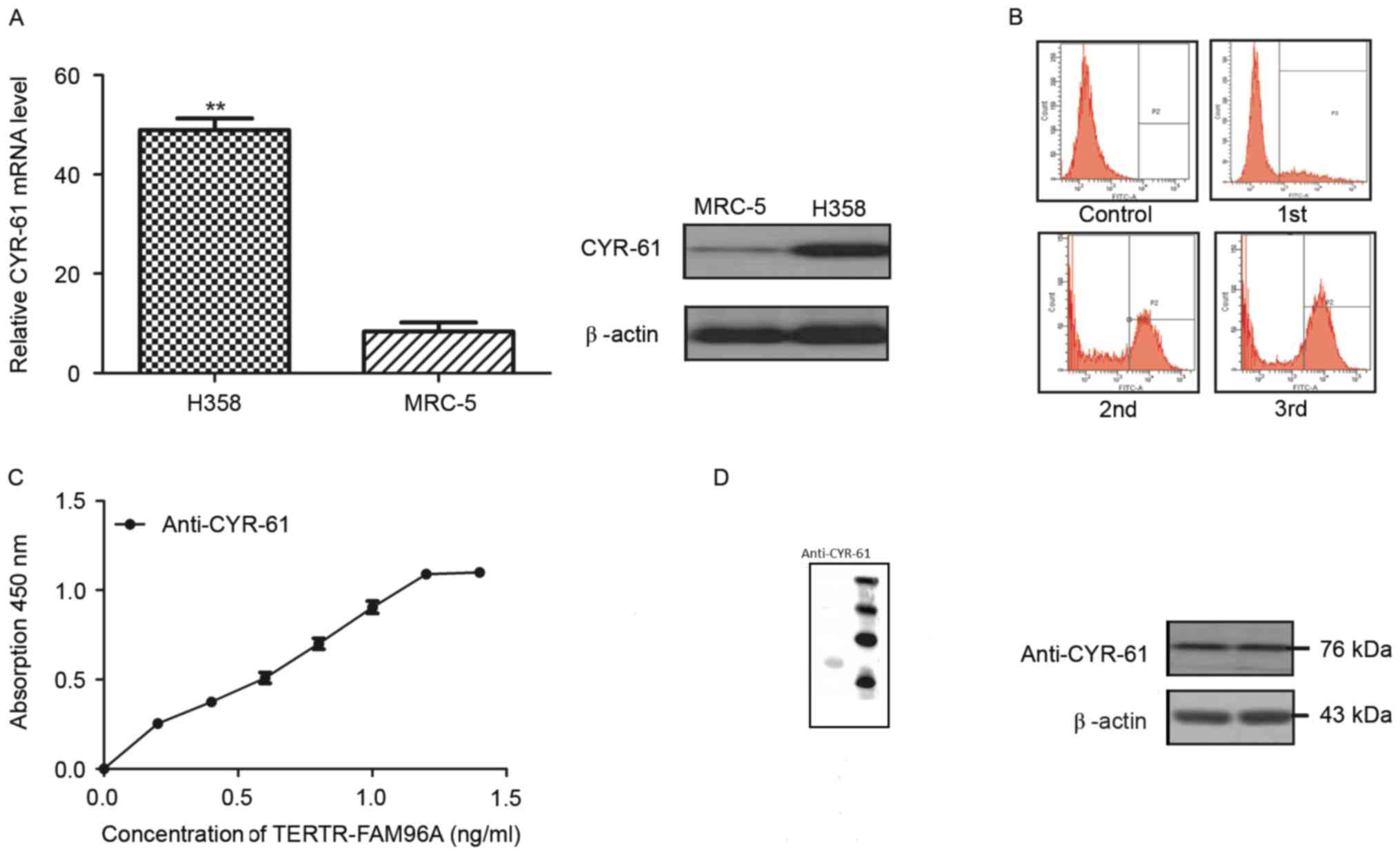
In order to analyze the role of CYR-61 in NSCLC,
mRNA and protein expression levels of CYR-61 were detected in the
NSCLC cell line H358 and a normal lung cell line MRC-5 by RT-qPCR
analysis and western blotting. The results demonstrated that CYR-61
mRNA expression levels were significantly higher (P<0.01) and
protein expression levels were markedly higher in the H358 cell
line compared with the levels in the MRC-5 cell line (Fig. 1A). These findings suggested that
CYR-61 was a potential molecular target for NSCLC. Therefore, a
full-length antibody was constructed to target CYR-61, termed
anti-CYR-61, which was screened by flow cytometry. The population
of CYR-61 positive cells began to reduce 1 h after anti-CYR-61
treatment, and reduced to its lowest level 3 h after the treatment
(Fig. 1B). An ELISA assay of
anti-CYR-61 demonstrated a high affinity with CYR-61 (Fig. 1C). Western blotting revealed that
anti-CYR-61 was 76 kDa under non-reducing conditions (Fig. 1D). Western blotting also demonstrated
that anti-CYR-61 was able to specifically bind with CYR-61. These
results indicated that CYR-61 was upregulated in NSCLC cell lines
and that the anti-CYR-61 antibodies we constructed exhibited high
affinity for CYR-61.
Effect of anti-CYR-61 on
CYR-61-induced NSCLC viability, migration and invasion
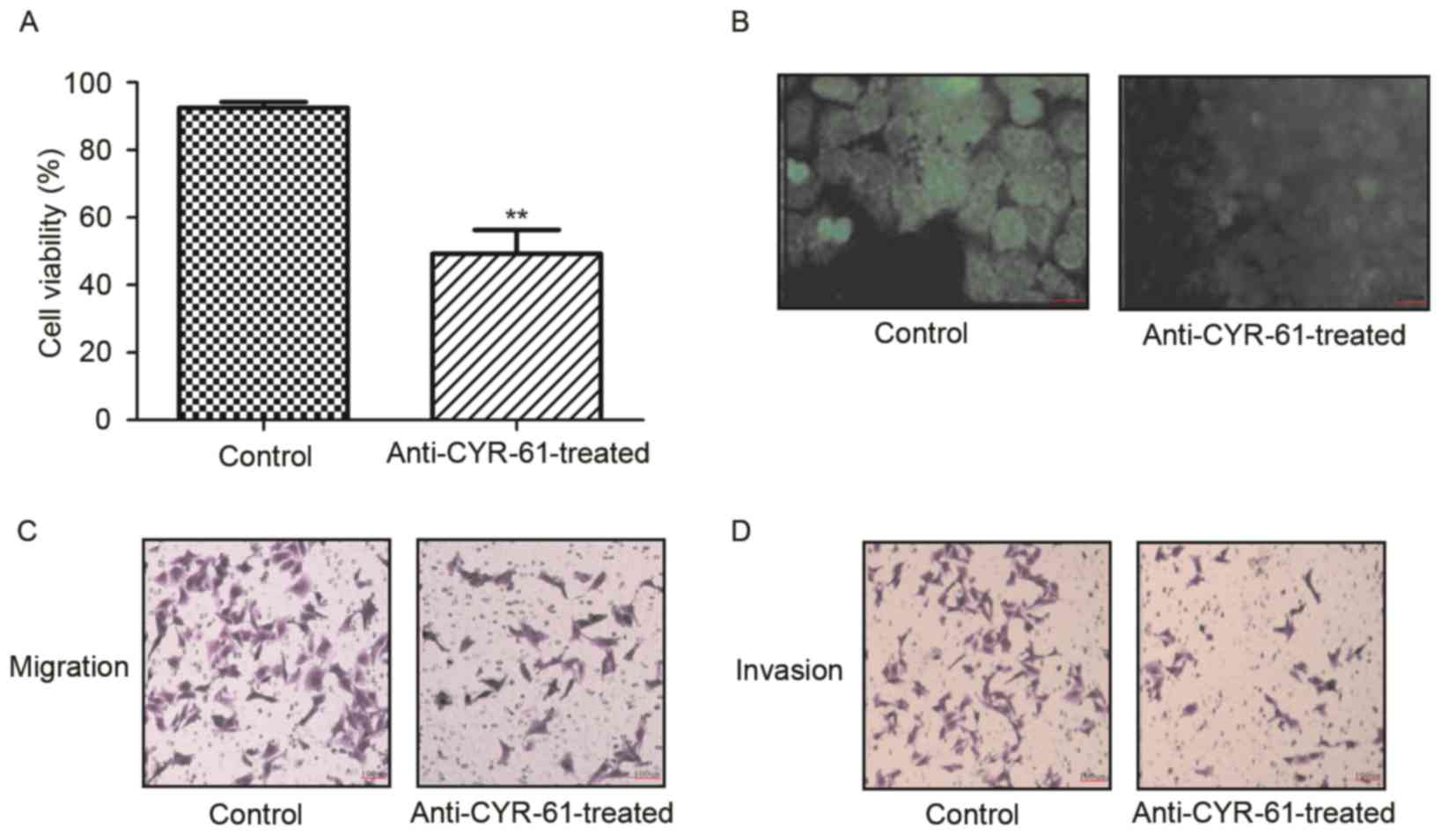
To identify the function of CYR-61 in NSCLC, we
investigated the inhibitory effects of anti-CYR-61 on cell
viability, migration and invasion of H358 cells. As demonstrated in
Fig. 2A and B, H358 cell viability
was significantly inhibited following treatment with anti-CYR-61,
which decreased CYR-61 expression (P<0.01). In migration and
invasion assays, anti-CYR-61 treatment inhibited the migration and
invasion of H358 cells (Fig. 2C and
D). These results indicated that anti-CYR-61 may have potential
therapeutic effects by inhibiting NSCLC cell viability, migration
and invasion by targeting CYR-61.
Effect of anti-CYR-61 on NSCLC cell
migration and the AKT and ERK signaling pathways
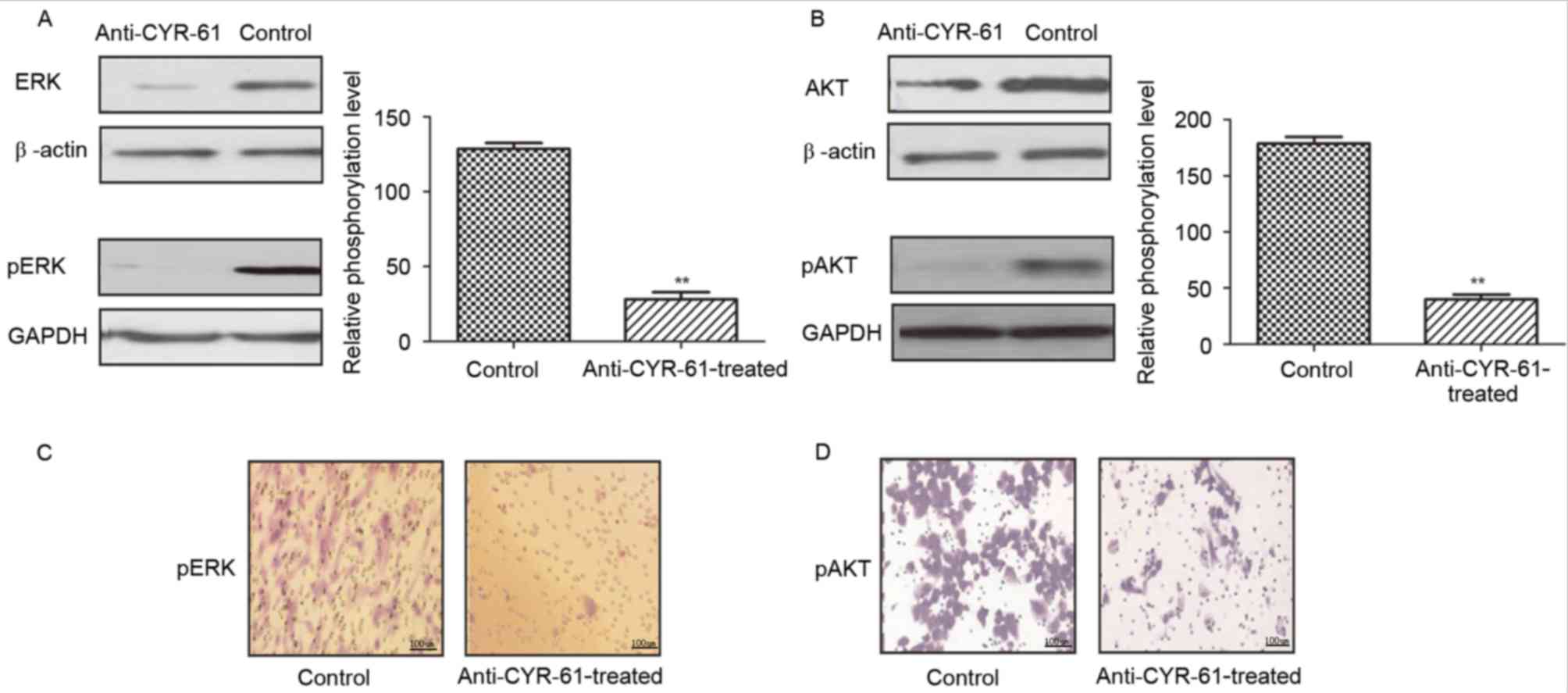
To further investigate the effects and mechanism of
anti-CYR-61 on cell migration, we analyzed ERK and AKT expression
and phosphorylation levels in H358 cells prior to and following
treatment with anti-CYR-61. As demonstrated in Fig. 3A and B, anti-CYR-61 treatment
significantly suppressed AKT and ERK protein expression and
phosphorylation compared with untreated controls (P<0.01),
indicating a reduction in AKT and ERK activity in NSCLC cells
treated with anti-CYR-61. Immunofluorescence also indicated that
expression and phosphorylation levels of AKT and ERK were inhibited
in H358 cells treated with anti-CYR-61 (Fig. 3C and D). These results suggested that
the inhibitory effect of anti-CYR-61 on migration may involve AKT
and ERK phosphorylation and the ERK and AKT signaling pathways.
Role of anti-CYR-61 on cell
apoptosis
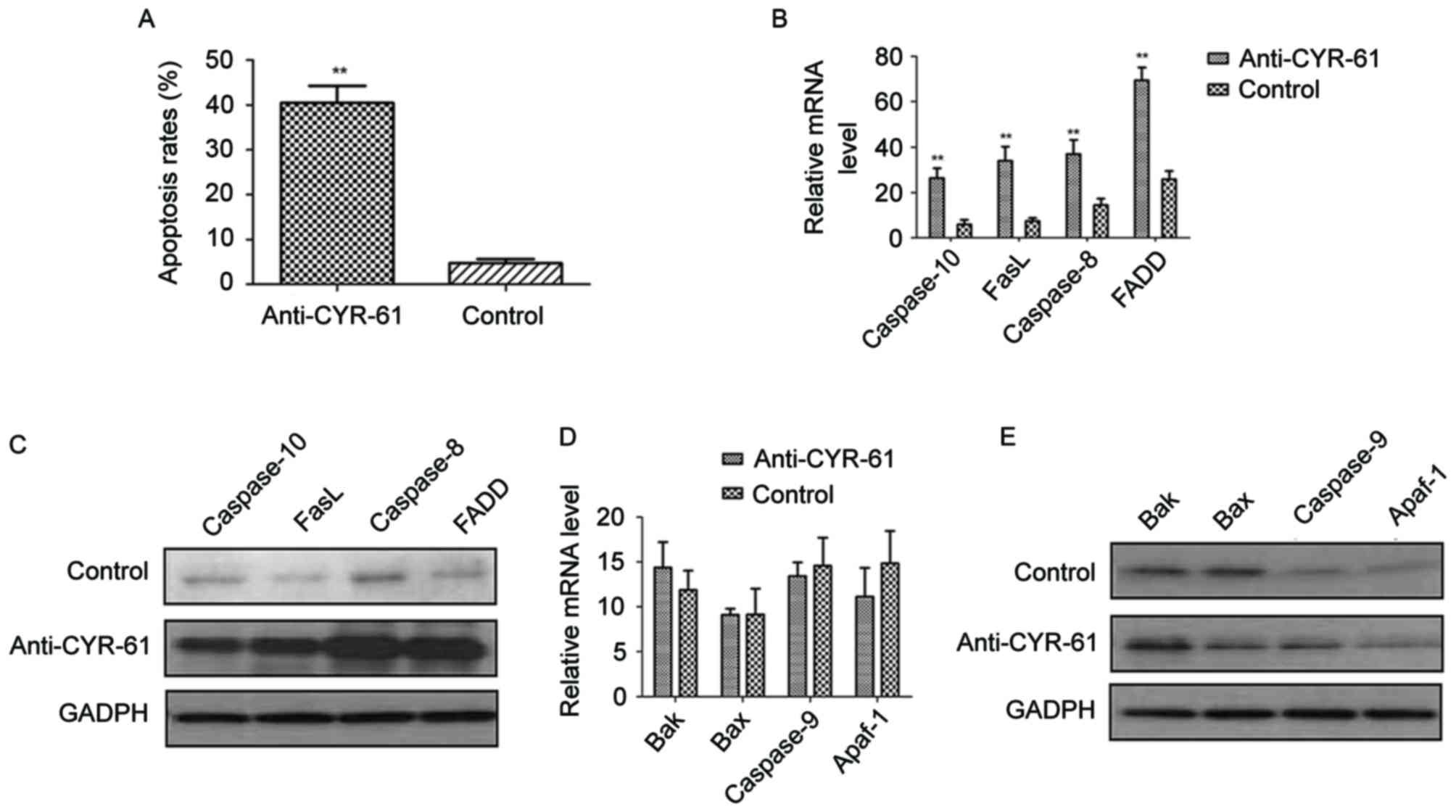
As the results demonstrated that anti-CYR-61
regulated NSCLC cell migration through phosphorylation of ERK and
AKT process, the apoptotic effects of anti-CYR-61 on H358 cells by
blocking the CYR-61 were analyzed. NSCLC H358 cells were treated
with or without anti-CYR-61. Results demonstrated that anti-CYR-61
significantly promoted H358 cell apoptosis compared with the
control cells (P<0.01; Fig. 4A).
In addition, the relationship between anti-CYR-61 and apoptosis
signaling pathways in H358 cells was investigated. Results
indicated that FasL, FADD, caspase-8 and −10 mRNA expression levels
were significantly elevated by anti-CYR-61 treatment compared with
the control cells (P<0.01; Fig.
4B). Western blot analysis also revealed that FasL, FADD,
caspase-8 and −10 protein expression levels were markedly
upregulated following treatment with anti-CYR-61 compared with
control cells (Fig. 4C). However, no
significant difference in the expression levels of Bak, Bax, Apaf-1
and caspase-9 were observed between anti-CYR-61-treated cells and
the control cells (Fig. 4D and E).
These results suggested that anti-CYR-61 has an important role in
regulating exogenous cell apoptosis signaling pathways for NSCLC
cells.
Anti-tumor effects of anti-CYR-61 in
mice with NSCLC
Following observation of the inhibitory effects of
anti-CYR-61 on NSCLC cell viability in vitro, the anti-tumor
efficacy of anti-CYR-61 in H358-bearing mice was investigated in
vivo. As illustrated in Fig. 5A,
tumor growth was significantly inhibited 21 days after anti-CYR-61
treatment, as determined via tumor volume, compared with
PBS-treated mice (P<0.01). In addition, immunohistology
demonstrated that CYR-61 expression was downregulated in
anti-CYR-61-treated tumors compared with PBS-treated tumors
(Fig. 5B). In agreement with the
in vitro results, the findings demonstrated that AKT and ERK
expression were decreased in tumors from experimental mice treated
with anti-CYR-61 on day 25 compared with control mice treated with
PBS (Fig. 5C). To analyze the
relationship between anti-CYR-61 and the EMT process in
vivo, EMT biomarker expression levels of vimentin, fibronectin,
SMA and N-cadherin were analyzed. The results in Fig. 5D demonstrated that the protein
expression levels of EMT markers were decreased in
anti-CYR-61-treated tumors compared with PBS-treated tumors.
Furthermore, it was observed that the survival rate of Anti-CYR-61
treatment is significantly higher than that of PBS treatment after
day 40 during a 120-day observation period (P<0.01; Fig. 5E). Anti-CYR-61 treatment
significantly inhibited tumor metastasis compared with PBS-treated
mice (P<0.01; Fig. 5E and F). In
conclusion, our findings suggested that anti-CYR-61 presented
potential anti-cancer efficacy in NSCLC treatment in murine
model.
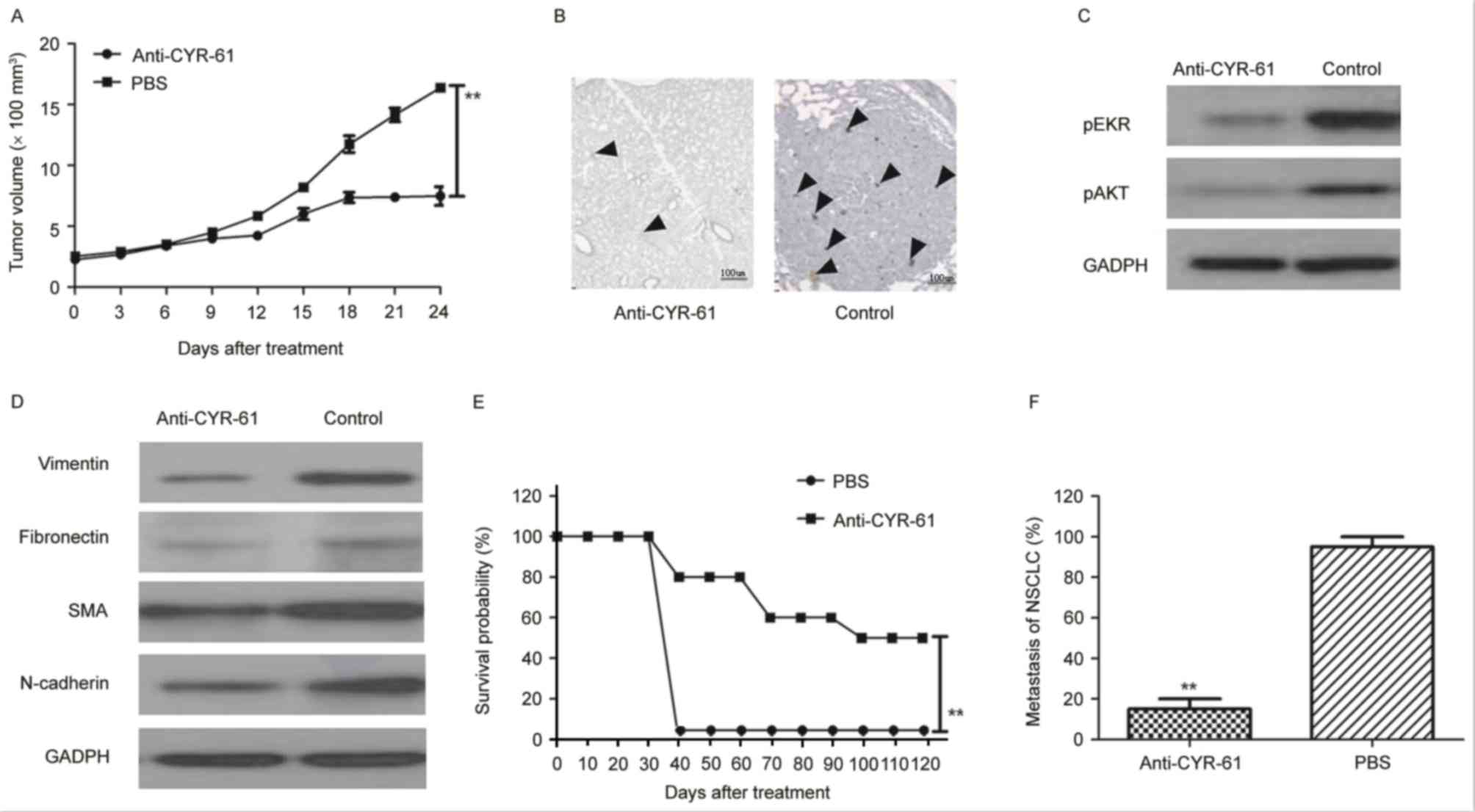 | Figure 5.Therapeutic and metastasis effects of
anti-CYR-61 in H358-bearing mice. (A) Tumor growth was analyzed
during a 24-day short-term observation period. Data are presented
as the mean ± SD. **P<0.01 vs. PBS on day 21. (B) CYR-61
expression was analyzed after anti-CYR-61 treatment and DAB
staining. Black arrows, in situ CYR-61 (C) Phosphorylation
levels of ERK and AKT were analyzed by western blotting in tumors
following treatment with anti-CYR-61. (D) EMT biomarker expression
levels were analyzed by western blotting in tumors following
treatment with anti-CYR-61. (E) Long-term survival probability was
compared over a 120-day observation between the anti-CYR-61 and PBS
treatment groups. It was demonstrated that the survival rate of
Anti-CYR-61 treatment was significantly higher than that of PBS
treatment after day 40 during the 120 day observation period. (F)
NSCLC metastasis was analyzed in experimental mice. Data are
expressed as the mean ± SD. **P<0.01 vs. PBS. CYR-61,
cysteine-rich angiogenic inducer-61; ERK, extracellular
signal-regulated kinase; AKT, protein kinase B; EMT,
epithelial-mesenchymal transition; PBS, phosphate-buffered saline;
NSCLC, non-small cell lung cancer; p, phosphorylated; SMA, α-smooth
muscle actin; SD, standard deviation. |
Discussion
The incidence and mortality rate of human lung
cancer has been growing rapidly in recent years and is one of the
most threatening types of malignant tumors to health and survival
(28). NSCLC represents more than
85% of lung cancer cases according to statistical clinical data
(29). Therefore, NSCLC has
attracted much attention to identify anti-cancer agents, ranging
from molecular markers to immunotherapy, in order to improve the
treatment and prognosis of patients with NSCLC. Notably, the
majority of newly diagnosed NSCLC cases are often in the moderate
or severe stage, decreasing the recovery probability and survival
period (30). Therapeutic protocols
for advanced NSCLC have expanded to include a number of targeted
interventions, including chemotherapy, radiotherapy, small molecule
target therapy, personalized treatment and immunotherapy (31). Various types of treatment for NSCLC
have been introduced with different mechanisms of action; they have
been shown to change tumor architecture and the tumor
microenvironment, however the outcomes were not promising as an
early reduction in tumor mass was not achieved (32). The reasons of treatment failure were
predominantly that these treatments failed to control NSCLC
migration and inhibit invasion in patients during treatment
periods.
CYR-61 is a secreted protein of the CCN family that
is associated with the extracellular matrix signaling pathway.
Previous research has indicated that CYR-61 is potent in regulating
tumor cell activities, such as tumor cell growth, apoptosis,
proliferation, migration, adhesion, differentiation and the EMT
process, in the majority of human cancer cells (17). A study by Sabile et al
(33) reported that CYR-61 signaling
was regulated by phosphorylation of AKT and ERK in osteosarcoma
tumor and lung cancer cells. In addition, a study by Chen et
al (34) described that
phosphorylation of AKT and ERK was significantly associated with
the migration and invasion of prostate carcinoma PC-3 cells. In the
present study, a full-length antibody target for CYR-61 was
constructed and its anti-tumor efficacy in a murine model of lung
cancer was investigated. In agreement with results from a previous
report (35), the present study
demonstrated that anti-CYR-61 treatment inhibited phosphorylation
of the AKT and ERK signaling pathway in human NSCLC cells.
Furthermore, it was demonstrated that anti-CYR-61 inhibited tumor
growth and downregulated EMT biomarker expression of vimentin,
fibronectin, SMA and N-cadherin.
In a previous report, CYR-61 expression was
demonstrated to be downregulated via inhibition of the ERK and AKT
pathways, resulting in suppression of colon cancer cell migration
(19). A study by Lee et al
(36) suggested that activities of
CYR-61 protein were modulated through extracellular acidification
and the PI3K/AKT signaling pathway in prostate carcinoma cells.
Furthermore, Chen et al (34)
indicated that transforming growth factor-β induced CYR-61
production to enhance tumor cell migration and invasion in
vitro and in vivo. These observations were supported in
the present study in NSCLC cells and the results of the present
study also suggested that inhibition of CYR-61 production is
beneficial for inhibiting NSCLC cell viability, migration and
invasion via the AKT and ERK signaling pathways.
In the present study, CYR-61 production in human
NSCLC cells and normal lung cells was investigated. The critical
role of the CYR-61 signaling pathway for the EMT process was
confirmed in NSCLC H358 cells. Previous research has reported that
activation of ERK and the PI3K/AKT signal pathway has an important
role in regulation of tumor cell migration (37). In the present study, antibody
targeting of CYR-61 not only inhibited viability of NSCLC H358
cells, it also suppressed migration of NSCLC H358 cells in
vitro and in vivo.
In conclusion, the aim of the present study was to
investigate the association between CYR-61 and the prognosis of
NSCLC in a murine model. Anti-CYR-61was demonstrated to be a
potential anti-tumor agent for NSCLC by targeting CYR-61, which led
to inhibition of migration by decreasing the production of CYR-61
via suppressing phosphorylation of AKT and ERK. Taken together, the
therapeutic efficacy of anti-CYR-61 was examined and the results
suggested that this targeted strategy may represent an attractive
method of inhibiting tumor cell migration for the treatment of
NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and YQ constructed and performed the cell
invasion and migration assays, and were major contributors in
writing the manuscript. NY analyzed and interpreted the RT-qPCR,
western blotting and histological immunostaining data. LL performed
and analyzed the animal study and ELISA assay. LY performed and
analyzed the MTT and apoptosis assays.
Ethics approval and consent to
participate
The present study was carried out in strict
accordance with the approval and recommendations from the Ethics
Committee of the Care and Use of Laboratory Animals of Qilu
Hospital of Shandong University (Jinan, China). All surgery and
euthanasia were performed under sodium pentobarbital anesthesia,
and all efforts were made to minimize suffering.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fenton-Ambrose L and Kazerooni EA:
Preventative care: Lung-cancer screens now worth the cost. Nature.
514:352014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Awad R and Nott L: Radiation recall
pneumonitis induced by erlotinib after palliative thoracic
radiotherapy for lung cancer: Case report and literature review.
Asia Pac J Clin Oncol. 12:91–95. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang SY, Zhao J, Wang MZ, Huo Z, Zhang J,
Zhong W and Xu Y: Small-cell lung cancer transformation in patients
with pulmonary adenocarcinoma: A case report and review of
literature. Medicine (Baltimore). 95:e27522016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kong R, Feng J, Ma Y, Zhou B, Li S, Zhang
W, Jiang J, Zhang J, Qiao Z, Zhang T, et al: Silencing NACK by
siRNA inhibits tumorigenesis in non-small cell lung cancer via
targeting Notch1 signaling pathway. Oncol Rep. 35:2306–2314. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brody H: Lung cancer. Nature. 513:S12014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moro-Sibilot D, Smit E, de Castro Carpeño
J, Lesniewski-Kmak K, Aerts JG, Villatoro R, Kraaij K, Nacerddine
K, Dyachkova Y, Smith KT, et al: Non-small cell lung cancer
patients with brain metastases treated with first-line
platinum-doublet chemotherapy: Analysis from the European FRAME
study. Lung Cancer. 90:427–432. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Barnett SA, Downey RJ, Zheng J, Plourde G,
Shen R, Chaft J, Akhurst T, Park BJ and Rusch VW: Utility of
routine PET imaging to predict response and survival after
induction therapy for non-small cell lung cancer. Ann Thorac Surg.
101:1052–1059. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xie FJ, Lu HY, Zheng QQ, Qin J, Gao Y,
Zhang YP, Hu X and Mao W: The clinical pathological characteristics
and prognosis of FGFR1 gene amplification in non-small-cell lung
cancer: A meta-analysis. Onco Targets Ther. 9:171–181. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lim SH, Sun JM, Lee SH, Ahn JS, Park K and
Ahn MJ: Pembrolizumab for the treatment of non-small cell lung
cancer. Expert Opin Biol Ther. 16:397–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Müller B, Bovet M, Yin Y, Stichel D, Malz
M, González-Vallinas M, Middleton A, Ehemann V, Schmitt J, Muley T,
et al: Concomitant expression of far upstream element (FUSE)
binding protein (FBP) interacting repressor (FIR) and its splice
variants induce migration and invasion of non-small cell lung
cancer (NSCLC) cells. J Pathol. 237:390–401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao Q, Yue J, Zhang C, Gu X, Chen H and
Xu L: Inactivation of M2 AChR/NF-κB signaling axis reverses
epithelial-mesenchymal transition (EMT) and suppresses migration
and invasion in non-small cell lung cancer (NSCLC). Oncotarget.
6:29335–29461. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang H, Zhu X, Li N, Li D, Sha Z, Zheng X
and Wang H: miR-125a-3p targets MTA1 to suppress NSCLC cell
proliferation, migration, and invasion. Acta Biochim Biophys Sin
(Shanghai). 47:496–503. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Roth MT, Ivey JL, Esserman DA, Crisp G,
Kurz J and Weinberger M: Individualized medication assessment and
planning: Optimizing medication use in older adults in the primary
care setting. Pharmacotherapy. 33:787–797. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin Y, Xu T, Tian G and Cui M:
Cysteine-rich, angiogenic inducer, 61 expression in patients with
ovarian epithelial carcinoma. J Int Med Res. 42:300–306. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu ST, Ding X, Ni QF and Jin SJ: Targeting
MACC1 by RNA interference inhibits proliferation and invasion of
bladder urothelial carcinoma in T24 cells. Int J Clin Exp Pathol.
8:7937–7944. 2015.PubMed/NCBI
|
|
16
|
Han S, Bui NT, Ho MT, Kim YM, Cho M and
Shin DB: Dexamethasone inhibits TGF-β1-induced cell migration by
regulating the ERK and AKT pathways in human colon cancer cells via
CYR61. Cancer Res Treat. 48:1141–1153. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Osaki M, Inaba A, Nishikawa K, Sugimoto Y,
Shomori K, Inoue T, Oshimura M and Ito H: Cysteine-rich protein 61
suppresses cell invasion via down-regulation of matrix
metalloproteinase-7 expression in the human gastric carcinoma cell
line MKN-45. Mol Med Rep. 3:711–715. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hviid CV, Erdem JS, Kunke D, Ahmed SM,
Kjeldsen SF, Wang YY, Attramadal H and Aasen AO: The matri-cellular
proteins ‘cysteine-rich, angiogenic-inducer, 61’ and ‘connective
tissue growth factor’ are regulated in experimentally-induced
sepsis with multiple organ dysfunction. Innate Immun. 18:717–726.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ito T, Hiraoka S, Kuroda Y, Ishii S, Umino
A, Kashiwa A, Yamamoto N, Kurumaji A and Nishikawa T: Effects of
schizophrenomimetics on the expression of the CCN1 (CYR 61) gene
encoding a matricellular protein in the infant and adult neocortex
of the mouse and rat. Int J Neuropsychopharmacol. 10:717–725. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sabile AA, Arlt MJ, Muff R, Bode B,
Langsam B, Bertz J, Jentzsch T, Puskas GJ, Born W and Fuchs B:
Cyr61 expression in osteosarcoma indicates poor prognosis and
promotes intratibial growth and lung metastasis in mice. J Bone
Miner Res. 27:58–67. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aoki M and Fujishita T: Oncogenic roles of
the PI3K/AKT/mTOR axis. Curr Top Microbiol Immunol. 407:153–189.
2017.PubMed/NCBI
|
|
22
|
Chen X, Cheng H, Pan T, Liu Y, Su Y, Ren
C, Huang D, Zha X and Liang C: mTOR regulate EMT through RhoA and
Rac1 pathway in prostate cancer. Mol Carcinog. 54:1086–1095. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Risolino M, Mandia N, Iavarone F, Dardaei
L, Longobardi E, Fernandez S, Talotta F, Bianchi F, Pisati F,
Spaggiari L, et al: Transcription factor PREP1 induces EMT and
metastasis by controlling the TGF-β-SMAD3 pathway in non-small cell
lung adenocarcinoma. Proc Natl Acad Sci USA. 111:E3775–E3784. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bai F, Tian H, Niu Z, Liu M, Ren G, Yu Y,
Sun T, Li S and Li D: Chimeric anti-IL-17 full-length monoclonal
antibody is a novel potential candidate for the treatment of
rheumatoid arthritis. Int J Mol Med. 33:711–721. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhuang T, Djemil T, Qi P, Magnelli A,
Stephans K, Videtic G and Xia P: Dose calculation differences
between Monte Carlo and pencil beam depend on the tumor locations
and volumes for lung stereotactic body radiation therapy. J Appl
Clin Med Phys. 14:40112013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu TS, Yiao SY, Lim K, Jensen RV and Hsiao
LL: Interpretation of biological and mechanical variations between
the Lowry versus Bradford method for protein quantification. N Am J
Med Sci. 2:325–328. 2010.PubMed/NCBI
|
|
28
|
Jang BI and Hwang MJ: Do esophageal
squamous cell carcinoma patients have an increased risk of
coexisting colorectal neoplasms? Gut Liver. 10:6–7. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kulkarni S, Vella ET, Coakley N, Cheng S,
Gregg R, Ung YC and Ellis PM: The use of systemic treatment in the
maintenance of patients with non-small cell lung cancer: A
systematic review. J Thorac Oncol. 11:989–1002. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yoshiba S, Jansen M, Matsushima N, Chen S
and Mendell J: Population pharmacokinetic analysis of patritumab, a
HER3 inhibitor, in subjects with advanced non-small cell lung
cancer (NSCLC) or solid tumors. Cancer Chemother Pharmacol.
77:987–996. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Weller A, O'Brien MER, Ahmed M, Popat S,
Bhosle J, McDonald F, Yap TA, Du Y, Vlahos I and deSouza NM:
Mechanism and non-mechanism based imaging biomarkers for assessing
biological response to treatment in non-small cell lung cancer. Eur
J Cancer. 59:65–78. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rossi A and Di Maio M: Platinum-based
chemotherapy in advanced non-small-cell lung cancer: Optimal number
of treatment cycles. Expert Rev Anticancer Ther. 16:653–660. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sabile AA, Arlt MJ, Muff R, Husmann K,
Hess D, Bertz J, Langsam B, Aemisegger C, Ziegler U, Born W and
Fuchs B: Caprin-1, a novel Cyr61-interacting protein, promotes
osteosarcoma tumor growth and lung metastasis in mice. Biochim
Biophys Acta. 1832:1173–1182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen J, Song Y, Yang J, Gong L, Zhao P,
Zhang Y and Su H: The up-regulation of cysteine-rich protein 61
induced by transforming growth factor beta enhances osteosarcoma
cell migration. Mol Cell Biochem. 384:269–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wong WR, Chen YY, Yang SM, Chen YL and
Horng JT: Phosphorylation of PI3K/Akt and MAPK/ERK in an early
entry step of enterovirus 71. Life Sci. 78:82–90. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee YJ, Lee DM and Lee SH: Production of
Cyr61 protein is modulated by extracellular acidification and
PI3K/Akt signaling in prostate carcinoma PC-3 cells. Food Chem
Toxicol. 58:169–176. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shelton JG, Steelman LS, Lee JT, Knapp SL,
Blalock WL, Moye PW, Franklin RA, Pohnert SC, Mirza AM, McMahon M
and McCubrey JA: Effects of the RAF/MEK/ERK and PI3K/AKT signal
transduction pathways on the abrogation of cytokine-dependence and
prevention of apoptosis in hematopoietic cells. Oncogene.
22:2478–2492. 2003. View Article : Google Scholar : PubMed/NCBI
|