Introduction
In contrast to target-based Western medicine,
Traditional Chinese Medicine (TCM) employs a conceptual framework
whereby the body's balance of Yin-Yang energy is a crucial
factor in controlling normal functioning or homeostasis. According
to the concepts of TCM, Yang represents warmth and promotion
of beneficial functions, whilst Yin represents suppression
of nourishment. Kidney Yang deficiency syndrome
(KDS-Yang) is one of the classical syndrome patterns in TCM
and is characterized by disorders of multiple metabolic pathways.
Modern research has indicated that functional disorders and damage
to the hypothalamic-pituitary-adrenal (HPA) axis are the major
pathological mechanisms underlying KDS-Yang (1), with the key issue being deregulated
expression of adrenocortical hormone (ACTH) (2,3).
Patients with KDS-Yang invariably present with symptoms
including pain and weakness of the waist and knees, feeling cold,
fatigue, impaired hearing and tooth loss, and this syndrome is
often observed in the later stages of several common chronic
diseases, including rheumatoid arthritis, diabetes and hypertension
(4–6).
Sini decoction (SND) is a well-known remedy
in TCM that has been used to restore Yang body energy and
treat KDS-Yang and associated diseases for ~1,800 years
(7,8). In recent years, advances in
metabonomics have enabled identification of metabolic signatures of
KDS-Yang (9,10) and therapeutic biomarkers of SND
(11,12). Despite this, the sites of action,
molecular targets and active components of SND have remained
largely elusive.
The present study analyzed the in vivo
effects of SND treatment on HPA-axis circulating and in situ
hormone levels in a rat model of Yang deficiency. Of note,
gene microarray analysis of rat adrenal gland tissues identified a
marked upregulation of genes involved in stress and metabolic
response pathways as a consequence of SND treatment. Furthermore,
the present study highlighted a role for SND in regulating the
expression of cyclooxygenase (COX-2), also known as
prostaglandin-endoperoxide synthase 2 (PTGS2), in adrenocortical
cells, reflecting the potential of SND treatment to control
prostaglandin release and contribute to balanced sodium and water
homeostasis, possibly through nuclear factor (NF)-κB or cyclic
adenosine monophosphate (cAMP)-mediated signalling. Of note, the
results also suggested that SND, either directly or indirectly
through COX-2 and prostaglandin synthesis, protects against
oxidative cellular damage caused by reactive oxygen species
(ROS).
Materials and methods
Reagents
Hydrocortisone injection solution was purchased from
Tianjin Jiaozuo Pharmaceutical Co. (Tianjin, China). ELISA kits for
detection of testosterone, ACTH and cortisol (CORT) were obtained
from Elabscience (cat. no. E-EL-0072c, E-EL-R0048c and E-EL-R0269c,
respectively). 2′-7′-Dichlorodihydrofluorescein diacetate (DCFH-DA)
was from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The
NCI-H295R (H295R) cell line was obtained from the American Type
Culture Collection (ATCC; no. CRL-2128™). Dulbecco's modified
Eagle's medium (DMEM) was from Gibco (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and fetal bovine serum (FBS) was from
Biowest (Nuaillé, France). The mitochondrial membrane potential
detection kit (cat. no. C2006) was from Beyotime Institute of
Biotechnology (Haimen, China).
Preparation of SND
According to the original composition of SND
recorded in the Chinese Pharmacopoeia 2010 edition (13), SND was prepared using the following
procedure: The crude drugs of Acontium carmichaeli (90 g),
Zingiber officinale (60 g) and Glycyrrhiza uralensis
(90 g) were immersed in 2.4 l water for 1 h and then decocted to
boil for 2 h. The decoction was filtered through four layers of
gauze. Next, the drugs were boiled once again for 1 h with 1.9 l
water and the decoction was filtered again using the abovementioned
method. The successive decoctions were merged and condensed under
decompression. Finally, the extracted solution was made up to a
concentration of 1.0 g crude drug/ml.
Animal experiment
Animal experiments were performed in accordance with
the Guidelines for Animal Experimentation of Guangxi University of
Chinese Medicine (Nanning, China). The protocols were approved by
the ethics committee of the First Affiliated Hospital of Guangxi
University of Chinese Medicine (Nanning, China). A total of 30 Male
Wistar rats (age, 9 weeks; body weight, 200–250 g, Shanghai SLAC
Laboratory Animal Co., Ltd., Shanghai, China) were kept under
standard conditions with regulated temperature (17–25°C) and
relative humidity (45–60%) under a 12-h light/dark cycle. The
animals had ad libitum access to food and drinking water
throughout the study period. After one week of habituation, rats
were randomly assigned to 5 groups: Blank, Model, SND-low dose,
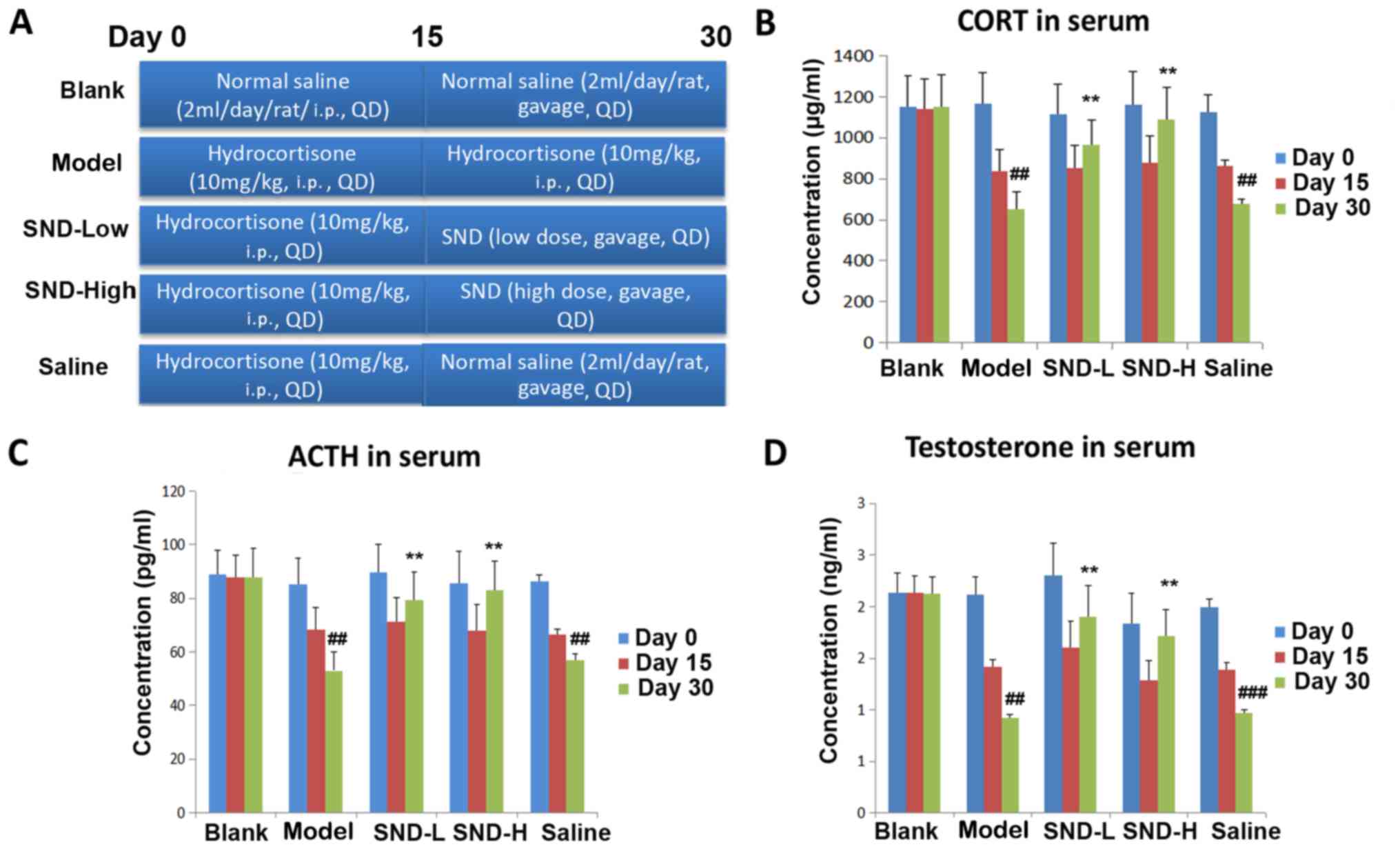
SND-high dose and Saline, as depicted in the schematic in Fig. 1A. In the SND treatment groups, rats
were intraperitoneally (i.p.) injected with hydrocortisone at a
dose of 10 mg/kg once daily for 15 days and then administered SND
extract by oral gavage twice daily for 15 days (high dose, 6.3 g
decoction/kg body weight; low dose, 1.26 g decoction/kg body
weight). In the Saline group, rats were intraperitoneally injected
with hydrocortisone at a dose of 10 mg/kg once daily for 15 days
and then orally administered normal saline twice daily for 15 days.
Rats in the Model group received 15 days +15 days of hydrocortisone
(i.p.) at a dose of 10 mg/kg once daily (9). Rats in the Blank group received 15 days
of saline (i.p.), followed by 15 days of oral saline (Fig. 1A). On Days 0, 15 and 30, 400 µl of
blood from tail vein of each rat were collected into heparinized
tubes and immediately centrifuged at 14,360 × g for 10 min. The
plasma was transferred into clean tubes and stored at −80°C until
biochemical analysis.
Immunohistochemistry
Immunohistochemistry was performed on a 4 µm thick
formalin-fixed (immersed in 10% formalin for 24 h at room
temperature) paraffin-embedded sections using the Real EnVision
Detection System and Peroxidase/diaminobenzidine (Dako; Agilent
Technologies, Inc., Santa Clara, CA, USA) according to the
manufacturer's protocol with minor modifications. Briefly, tissue
sections were deparaffinized, and rehydrated using graded alcohols.
Antigen retrieval was performed by heating the slides in a
microwave oven (500 W for 5 min 3 times in citrate buffer pH 6.0 at
96°C) (14). Endogenous peroxidase
was quenched for 10 min with peroxidase blocking reagent. The
sections were washed using PBS and incubated in blocking solution
containing 2% donkey serum (cat. no. AR009; Boster Biological
Technology, Pleasanton, CA, USA), followed by incubation overnight
at 2–4°C with primary antibodies [corticotropin-releasing hormone
(CRH) antibody, cat. no. BA1503; 1:100; ACTH antibody, cat. no.
BA0003-1; 1:100; Boster Biological Technology]. Following washing
with PBS buffer, sections were incubated with peroxidase labelled
secondary antibodies from the Envision kit for 40 min at room
temperature prior to staining with diaminobenzidine substrate
chromogen (also sourced from the kit) for 10 min at room
temperature. Images were recorded using confocal microscopy (Leica
Microsystems GmbH, Wetzlar, Germany). Cells with dark staining were
counted as positive and analysed using Image-Pro® Plus
version 5.1 (Media Cybernetics, Inc., Rockville, MD, USA)
software.
Cell culture and assays
The H295R cell line was cultured in DMEM medium
containing 10% FBS. For the 2′-7′-dichlorodihydrofluorescein
diacetate (DCFH-DA) assay, cells were cultured in 96-well plates
and labeled with DCFH-DA (10 µM) or unlabeled (none) and then
cultured for an additional 3 h. Cells were then analyzed on a
fluorescence reader using excitation/emission wavelengths of
485/535 nm. For the JC-1 assay, cells were cultured in 96-well
plates and stained with JC-1 according to the manufacturer's
instructions. Fluorescence of J-aggregates were measured on the
plate reader using excitation/emission wavelengths of 535/595 nm.
In the assay that suppressed the upregulation of COX-2, H295R cells
were treated in vitro with either vehicle control (blank
group), SND alone, H2O2 for 30 min
(H2O2 group) or SND pre-treatment for 24 h
followed by H2O2 for a further 30 min (SND +
H2O2 group). Cells were then collected for
quantitative real-time polymerase chain reaction (PCR).
Gene microarray analysis
To further investigate the potential modes of action
of SND treatment, a gene microarray analysis of mRNA derived from
rat adrenal gland tissues from the blank, control and SND treatment
groups was performed at day 30 (Fig.
1A). mRNA was isolated and analyzed using a rat-specific
Affymetrix GeneChip 1.0 ST Array (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The cluster algorithm test series of gene
expression dynamics was used to profile the gene expression series
and to identify the most probable set of clusters generating the
observed series. Gene ontology (GO) analysis was applied to analyze
the main function of differentially expressed genes according to GO
database and the AmiGO database (http://geneontology.org/; http://amigo.geneontology.org/amigo). Pathway analysis
was used to identify significant pathways of the differentially
expressed genes according to the Kyoto Encyclopedia of Genes and
Genomes database. Data were analyzed using GO analysis software.
Comparisons were made between control vs. blank, low-dose SND vs.
control and high-dose SND vs. control. Genes that were significant
in the GO as well as the pathway analyses were identified.
Differential gene expression analyses were performed using the
criteria of +2-fold or −0.5-fold differential expression and
P<0.05.
Reverse transcription (RT)
quantitative (q) PCR
RNA from H295R cells was extracted using the RNeasy
kit (Qiagen Inc., Valencia, CA, USA) according to the
manufacturer's protocol. cDNA was synthesized, and quantitative
RT-PCR analysis was performed using a Bio-Rad CFX 96 cycler
(Bio-Rad Laboratories Inc., Hercules, CA, USA) with the SuperScript
III two-step RT-qPCR kit with SYBR Green (Invitrogen; Thermo Fisher
Scientific, Inc.). The primer sequences utilized were as follows:
COX-2 forward, 5′-AGCCAGGCAGCAAATCCTT-3′ and reverse,
5′-GGGTGGGCTTCAGCAGTAAT-3′; Actin (used as the reference gene)
forward, 5′-AGAGGGAAATCGTGCGTGAC-3′ and reverse,
5′-CCATACCCAAGAAGGAAGGCT-3′. The thermocycling conditions were as
follows: Initial denaturation at 95°C for 1 min followed by 39
cycles at 95°C for 10 sec, 65°C for 30 sec, and 72°C for 30 sec.
The relative expression analysis of target genes were performed
using the 2−ΔΔCq method (15,16).
Reporter assays
H295R cells (seeded at 105 cells/well in
24-well plates) were co-transduced with NF-κB/renilla lentiviral
particles and CRE/renilla lentiviral particles. NF-κB, CRE and
renilla lentiviral particles were obtained from Novobio Scientific,
Inc. (Shanghai, China). The NF-κB reporter lenviral vector
contained an NF-κB responsive element (17), while the CRE reporter vector
contained a cAMP response-binding element (18). The Renilla lentiviral vector
expressing renilla luciferase was used as an internal control.
Following transduction at a multiplicity of infection of 50 for 8
h, blasticidin (4 µg/ml; Invitrogen; Thermo Fisher Scientific,
Inc.) were then added to screen stably transduced cells for 2
weeks. H295R cells expressing NF-κB/renilla reporters were
subjected to SND treatment (50 µg/ml) for 48 h, while CRE/renilla
expressing cells were treated with forskolin (5 µM) or forskolin (5
µM)+SND (50 µg/ml) for 48 h. Cells were collected and luciferase
assays were performed using a luminescence plate reader and
Dual-Luciferase Reporter Assay (cat. no. E1910; Promega
Corporation, Madison, WI, USA). The relative luciferase activity
for each sample was calculated as a ratio of firefly luciferase
activity divided by renilla luciferase activity.
Statistical analysis
Data were analyzed using GraphPad Prism software
version 6.0 (GraphPad Inc., La Jolla, CA, USA) and values are
expressed as the mean ± standard error of the mean. The
significance of differences was assessed using either a two-tailed,
non-paired Student's t-test or a one-way ANOVA and statistical
significance was defined using the customary threshold of P<0.05
(95% confidence interval).
Results
Treatment with SND ameliorates
KDS-Yang in a rat model
In order to assess the effects of SND treatment on
the HPA axis, a rat model of Yang deficiency was established
by treating animals daily with high-dose hydrocortisone (10 mg/kg,
i.p.) for 15 days, followed by withdrawal (8). At this point, rats were divided into
four groups (six rats in each group) and treated with either
continued hydrocortisone (10 mg/kg, i.p. Model group), low-dose
oral SND (SND-Low), high-dose oral SND (SND-High) or normal saline
(Saline) for a further 15 days. The Blank group received i.p.
saline injection for 15 days (2 ml/day), followed by oral saline
for 15 days (2 ml/day) (Fig. 1A).
Sera were collected from all rats on days 0, 15 and 30 and
subjected to ELISA to measure the levels of CORT, ACTH and
testosterone. As expected, KDS-Yang model animals exhibited
a significant decline in circulating cortisol levels, with a more
pronounced decrease observed with longer hydrocortisone treatment
(P<0.01; Fig. 1B). Of note,
animals in the low- and high-dose SND groups exhibited a marked
recovery of circulating CORT as compared with the saline group on
day 30 (P<0.01; Fig. 1B).
Similarly, KDS-Yang model group also displayed a significant
decline in circulating ACTH and testosterone levels (Fig. 1C and D), with a further decline
observed with increasing duration of hydrocortisone treatment
(P<0.01). After 15 days of hydrocortisone treatment, 15 days of
either low- or high-dose SND were sufficient for recovery of the
circulating ACTH and testosterone levels, when compared with those
in the group treated with normal saline for 15 days after
hydrocortisone withdrawal (P<0.01; Fig. 1C and D).
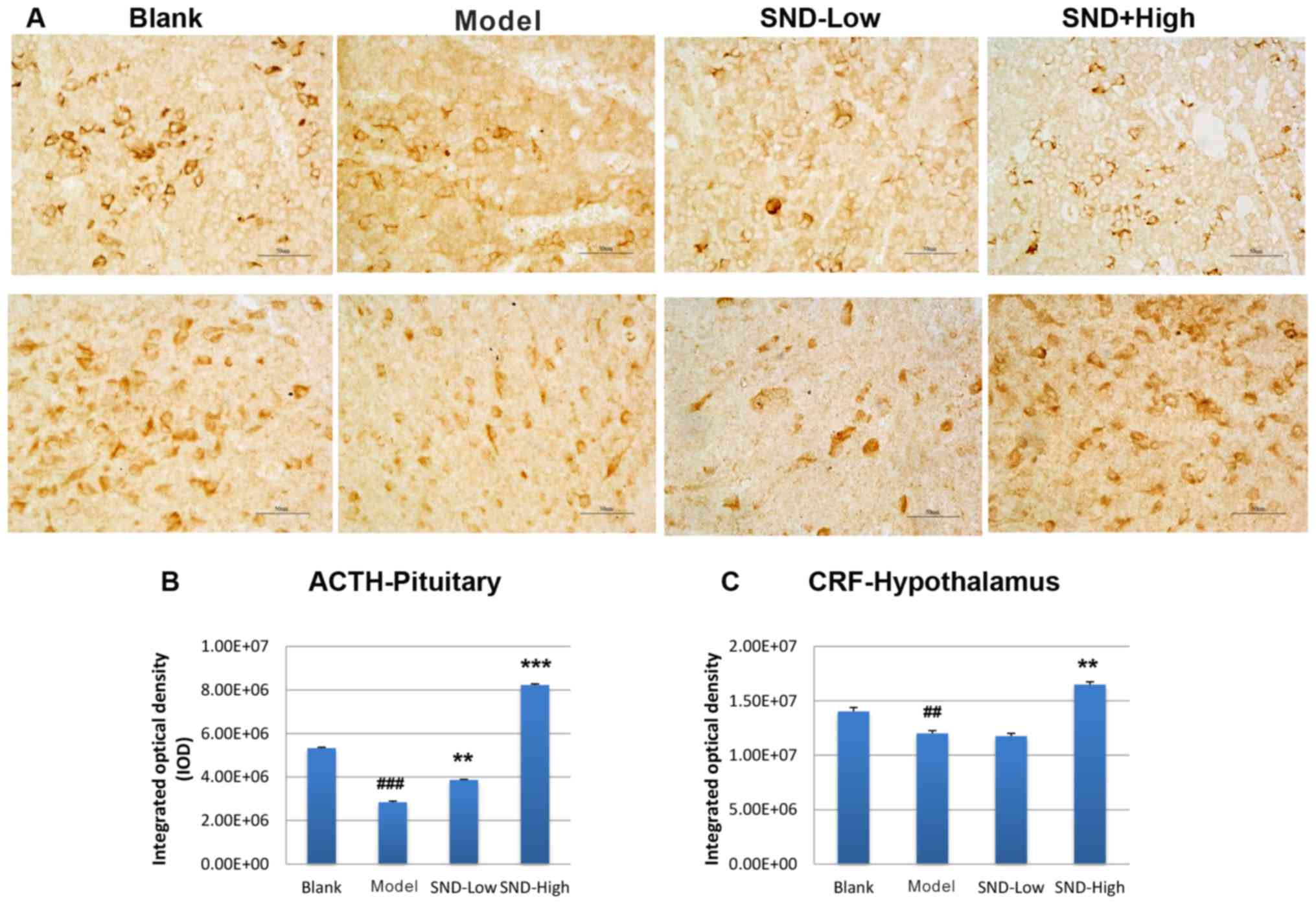
To further explore the mechanism underlying the
attenuation of HPA-axis hormone decline by SND treatment, rat
pituitary gland and hypothalamus tissues were obtained from each
treatment group at day 30. Tissue sections were subjected to
immunohistochemical analysis to detect the expression of ACTH and
CRH proteins in the pituitary gland and hypothalamus tissues,
respectively. Three slides from each rat were analysed and results
were quantified from these images. Representative images are
presented Fig. 2A. Relative to the
saline-treated control group, KDS-Yang rats exhibited a
marked decline in pituitary ACTH levels (Fig. 2A and B). Treatment with low-dose SND
produced a marginal elevation in ACTH levels and high-dose SND
resulted in a significant increase in pituitary ACTH levels to
exceed those in the saline and KDS-Yang controls (Fig. 2A and B). The levels of CRH in the
hypothalamus also exhibited a reduced trend in the KDS-Yang
rats (Fig. 2A and C). Of note, this
trend was reversed upon high-dose SND treatment, while low-dose SND
treatment had no effect (Fig.
2C).
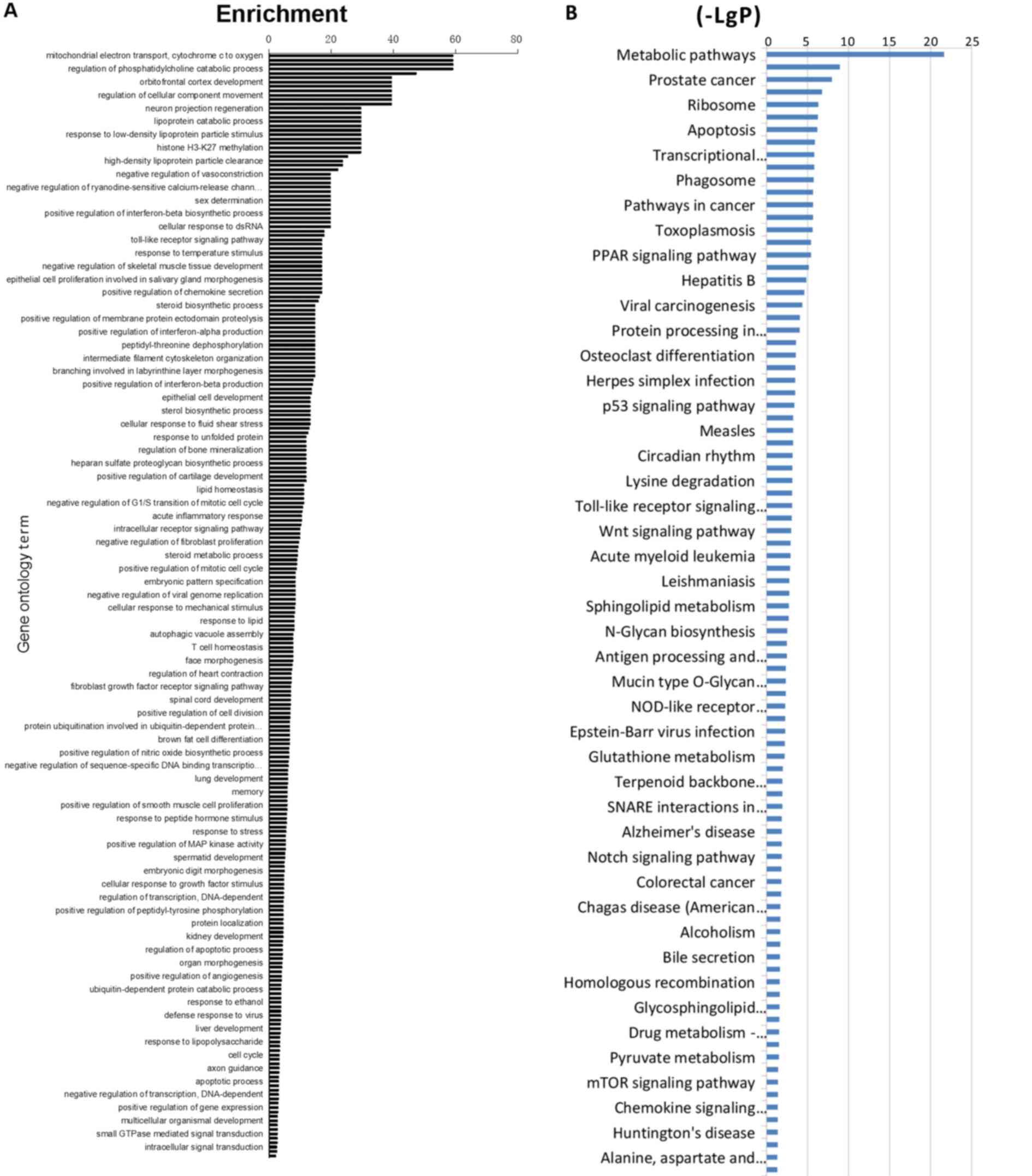
Genes involved in metabolic and stress
response pathways are upregulated in the adrenal gland in response
to SND treatment
To further investigate the potential modes of action
of SND treatment, a gene microarray analysis of mRNA derived from
adrenal gland tissues obtained from treated rats at 30 days was
performed. mRNA was isolated and analyzed using a rat-specific
Affymetrix GeneChip 1.0 ST Array. Data were analyzed using GO
analysis software and comparisons of control vs. blank, low-dose
SND vs. control and high-dose SND vs. control were made.
Differential gene expression analyses demonstrated that adrenal
gland tissues from SND-treated (high-dose) vs. control-treated rats
displayed marked differences in gene expression. Those genes with
the highest differential expression scores included COX-2, NF-κB
and genes involved in cAMP-dependent signalling. Genes were
clustered into categories based on the GO system and ordered
according to the highest enrichment (Fig. 3A). GO analysis was used to assign
these GO terms to kyto encyclopaedia of genes and genome pathways
and again, these pathways were ordered according to the highest
enrichment (Fig. 3B). According to
these analyses, differentially expressed genes in the SND-treated
(high-dose) vs. control-treated rat adrenal glands were enriched in
transcriptional regulation and metabolic signaling.
SND treatment results in partial
suppression of COX-2 upregulation in response to
H2O2 and protects adrenocortical cells from
ROS in vitro
As SND treatment resulted in significant
upregulation of genes predominantly involved in metabolic and
stress response signaling, the present study sought to further
confirm and explore the consequences of this in vitro using
the adrenocortical cell line H295R. Due to the known roles of COX-2
in regulating the response to kidney ischemia/reperfusion injury,
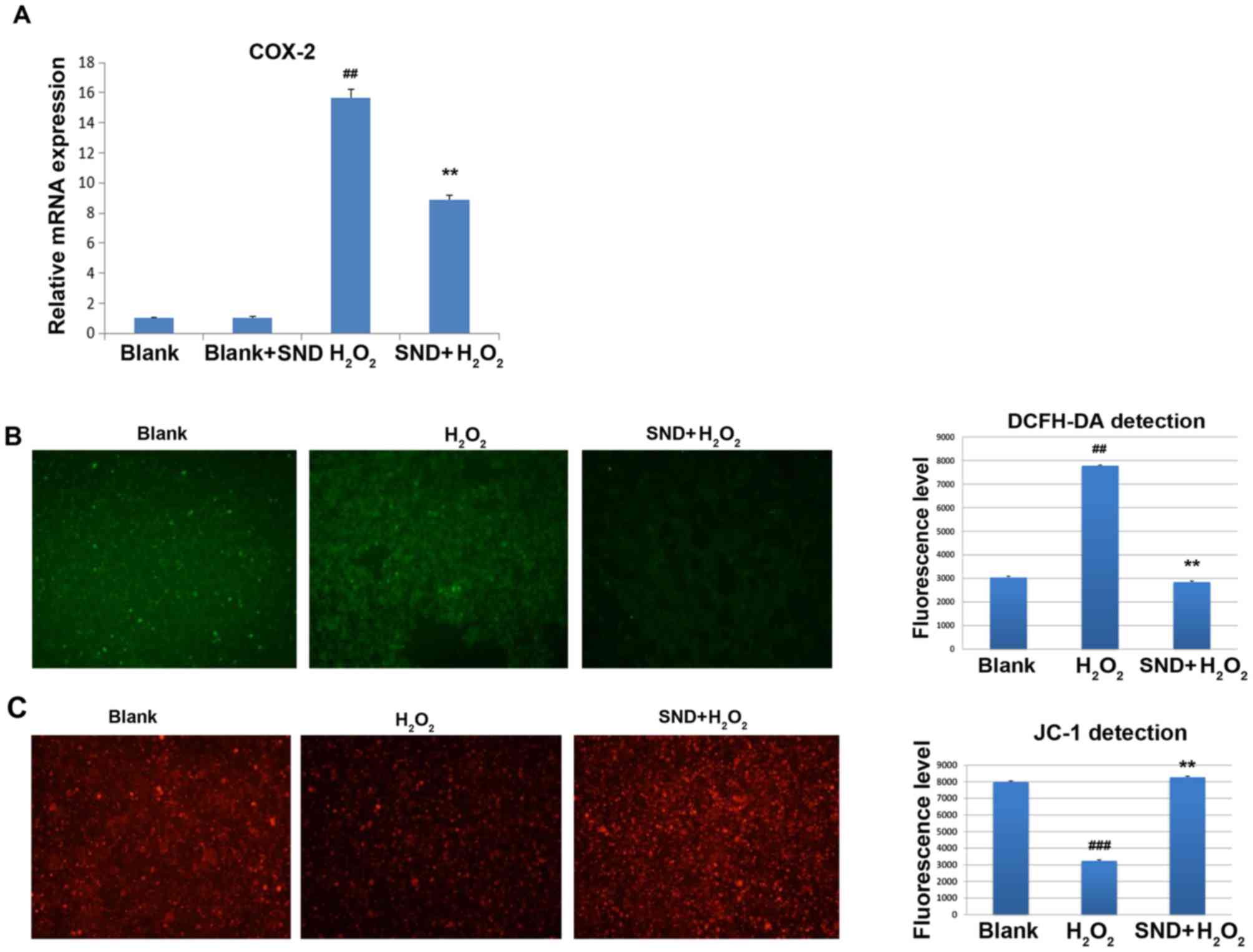
the effects of SND pre-treatment on COX-2 expression following
H2O2 treatment were assessed. COX-2 mRNA
expression was measured by reverse transcription-quantitative
polymerase chain reaction analysis. H295R cells were treated in
vitro with either vehicle control (blank group), SND alone,
H2O2 for 30 min (H2O2
group) or SND pre-treatment for 24 h followed by
H2O2 for a further 30 min (SND +
H2O2 group) (Fig.
4A). Treatment with H2O2 resulted in a
clear induction of COX-2 gene expression compared with that in the
vehicle-treated group (~15-fold induction) or SND-pre-treated blank
group (P<0.01; Fig. 4A). Of note,
pre-treatment of cells with SND for 24 h resulted in a significant
suppression of COX-2 induction compared with that in the group
treated with H2O2 only.
To further assess the functional consequences of
this SND-mediated partial suppression of COX-2 expression, the
effects of pre-treatment with SND on the
H2O2-induced generation of ROS were
determined using the cell permeable dye DCFH-DA. In the presence of
intracellular ROS, DCFH-DA is rapidly oxidized to fluorescent DCF.
As displayed in Fig. 4B, treatment
with H2O2 for 15 min resulted in a
significant elevation of ROS above the background (blank) levels.
However, pre-treatment of the cells with SND 24 h followed by
H2O2 treatment did not exhibit elevation,
indicating that ROS generation was prevented by SND.
As one of the major consequences of ROS-induced
damage is destruction of mitochondria through loss of the
mitochondrial membrane potential, ultimately leading to cellular
apoptosis, the ability of SND to protect against ROS-induced loss
of mitochondrial membrane potential was then investigated. For
this, H295R cells subjected to the above treatments were stained
with the membrane-permeant dye JC-1. JC-1 is widely used in
apoptosis studies to monitor mitochondrial health. Depending on the
mitochondrial membrane potential, the JC-1 dye accumulates in
mitochondria, which is indicated by a fluorescence emission shift
from green (~529 nm) to red (~590 nm). Consequently, mitochondrial
depolarization is indicated by a decrease in the red/green
fluorescence intensity ratio. The potential-sensitive color shift
is due to the concentration-dependent formation of red fluorescent
J-aggregates. Vehicle-treated (blank) H295R cells displayed the
expected accumulation of red JC-1 dye within the mitochondria,
whilst H2O2-treated cells exhibited a
significant decrease in the level of red JC-1 fluorescence due to
ROS-induced damage and loss of mitochondrial membrane potential
(Fig. 4C). However, pre-treatment of
H295R cells with SND prior by H2O2 treatment
increased the level of JC-1-associated red staining to a level
similar to that in the vehicle (control) group, indicating that SND
maintained the mitochondrial membrane potential and presumably
prevented mitochondrial membrane damage.
SND treatment of H295R cells results
in transcriptional activation through NF-κB and cAMP
response-binding element (CRE)-binding proteins in vitro
Based on known roles of NF-κB and CRE transcription
factor activity in regulating COX-2 gene expression and previous
studies on Yang deficiency (19–22), the
present study sought to explore whether in vitro SND
treatment resulted in activation of NF-κB or CRE activity in H295R
cells. To assess this, stably transfected H295R cells were
generated expressing lentiviral NF-κB or CRE-responsive luciferase
reporter constructs and subjected to SND treatment for 24 h.
Reporter activity was assessed using a luminescence plate reader.
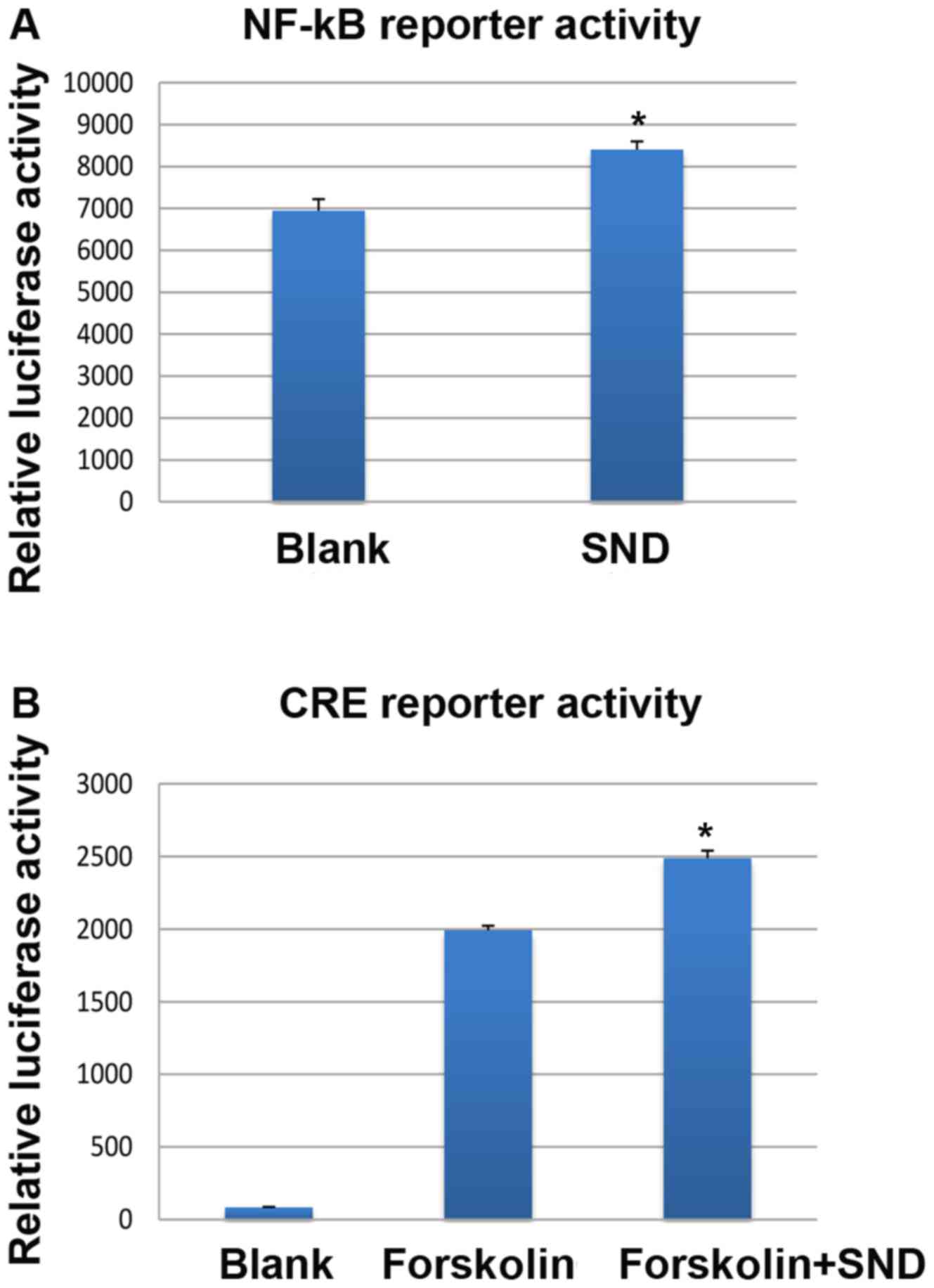
Treatment of H295R cells with SND resulted in a significant
increase in NF-κB-driven luciferase activity compared with that in
the vehicle-treated (blank) controls (~8,300 vs. ~6,900 units;
Fig. 5A). With regard to CRE-driven
luciferase activity, when the adenylyl cyclase activator forskolin
was used to artificially elevate intracellular cAMP levels, SND
treatment enhanced the effect compared with that of forskolin alone
(~2,500 vs. ~2,000 units; Fig.
5B).
Discussion
Recovery of body Yang energy using SND is
well known in TCM, but the underlying molecular mechanism has
remained to be elucidated. Using an in vivo rat model of
KDS-Yang, the present study demonstrated multiple effects of
SND treatment on the HPA-axis, including restoration of circulating
serum levels of CRH, ACTH and CORT, and importantly, an elevation
of in situ expression of ACTH. Based on prior literature
describing the ability of SND to improve adrenal gland function
(23), the present study focused on
investigating the effects of SND treatment at the molecular level
using gene microarray analysis of rat adrenal glands. To the best
of our knowledge, the present study was the first to pursue this
approach of studying the mechanisms of the biological effects of
SND, and a variety of results were obtained, several of which
indicated the restoration of HPA-axis functionality. Of note, a
marked upregulation of genes implicated in metabolic pathway
activation and stress responses were observed, including COX-2 and
NF-κB. In the context of the KDS-Yang model, in which the
HPA-axis activity is repressed as a consequence of prolonged
high-dose glucocorticoid treatment, induction of COX-2 expression
by SND is expected to reactivate HPA-axis activity due to the
ability of COX-2 to drive proliferative, pro-inflammatory and
injury responses (24). Within the
kidney, COX-2 has known roles in regulating renal physiology
through prostaglandin synthesis, control of fluid excretion and
intrarenal hemodynamics (25), thus
underscoring the importance of COX-2 expression in controlling
renal homeostasis.
The present study postulated that, in the context of
human KDS-Yang, where circulating ACTH and CORT levels are
low and patients present with a dampened immune system, low energy
metabolism and altered circadian cycles, activation of these
COX-2-driven proliferative and pro-inflammatory effects may be
beneficial in restoring the ability of the HPA-axis to respond to
physiological stresses. Similarly, despite being a pleiotropic
transcription factor, upregulation of NF-κB expression in the
adrenal gland in response to SND treatment is also consistent with
a stress response role. NF-κB is a rapidly-acting primary
transcription factor and has a key role in regulating the immune
response to infection (26).
Accordingly, SND-mediated activation of NF-κB expression and
activity is likely to restore a level of balance of the HPA-axis
and equilibrate the system.
As the present microarray data also indicated a
marked anti-oxidant response attributed to SND treatment, the
adrenocortical cell line H295R was used for further investigation
in vitro. It is worth mentioning that the TCM concept of
Yang is closely associated with energy metabolism, and
hence, aspects of mitochondrial function were also investigated. Of
note, in the context of an external insult
(H2O2 incubation), SND treatment exerted a
pronounced protective effect by repressing the induction of COX-2
expression. This is in contrast to the microarray data generated
using rat adrenal tissue from the KDS-Yang model and
suggests that SND treatment has pleiotropic effects, which may be
dependent on the cellular context and conditions (i.e. under
Yang deficiency, SND appears to provide an activating
stimulatory effect through upregulation of COX-2, but when
Yang is sufficient but cell-damaging conditions prevail, SND
may be protective by downregulating COX-2) (10). This protective effect was further
confirmed by prevention of ROS generation and maintenance of the
mitochondrial membrane potential in the presence of SND and
H2O2. In the ROS assay, although cells had
some fluorescence background as demonstrated in the blank group
(Fig. 4B), cells pretreated with SND
exhibited effects on the generation of ROS. Indeed, protective
effects of SND have also been observed in other disease states,
particularly cardiomyocytes in response to oxidative and
adriamycin-induced damage (27,28).
The present study further performed a reporter gene
induction study using NF-κB and CRE-responsive constructs for
several reasons. First, the microarray data highlighted marked
elevations in the expression of these two genes upon SND treatment
in the rat adrenal glands. Furthermore, according to DNA sequence
analysis, the COX-2 gene promoter contains binding sites for the
CRE transcription factor and finally, NF-κB and cAMP/cGMP have been
associated with the concept of Yin and Yang in TCM
(9). Indeed, Shen (29) has previously reported downregulation
of NF-κB expression in lymphocytes from a rat model of Yang
deficiency, whilst Goldberg et al (30), proposed a control mechanism for
opposing cAMP/cGMP in cellular regulation in the 1970's. The
present results indicated that SND treatment was able to stimulate
NF-κB and CRE-reporter activity in vitro, thus suggesting
that the expression of COX-2 is driven, at least in part, through
activation of these transcriptional pathways. However, it is likely
that the variety of pharmacologically active components within SND
acts via multiple pathways contributing to the gene expression
changes observed. Additional studies are currently underway to
further elucidate these mechanisms. However, the utilization of
animals was a limitation of the present study as individual
differences in hormone levels may result in variations in data
(Fig. 1D). Methods to prevent these
differences should be implemented in future studies.
Taken together, the results of the present study
confirmed that SND alleviates the hallmarks of KDS-Yang in a
rat model, through restoration of circulating and in situ
expression of HPA-axis hormones. Importantly, the characterization
of SND-induced gene expression changes in the rat adrenal glands
indicated the upregulation of metabolic and stress
response-associated signaling pathways, including those involving
COX-2 and NF-κB. Further in vitro analyses indicated the
protective effect of SND treatment within mitochondria and
suggested that the pleiotropic effects of SND are mediated, at
least in part, through activation of NF-κB and CRE signaling. These
conclusions offer a molecular insight into the well-established
effects of SND in alleviating KDS-Yang.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81573938 and
81460720).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NT and DM designed and directed the experiments. HQ
performed the experiments and wrote the manuscript. LL collected
the data. WS analyzed the data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The protocols were approved by the ethics committee
of Guangxi University of Chinese Medicine (Nanning, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shen Z: The location of deficiency
syndrome of kidney Yang. Chin Med J (Engl). 112:973–975.
1999.PubMed/NCBI
|
|
2
|
Liu X, Du J, Cai J, Liu X, Xu G, Lin A and
Teng Q: Clinical systematic observation of kangxin capsule curing
vascular dementia of senile kidney deficiency and blood stagnation
type. J Ethnopharmacol. 112:350–355. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xiong PH, Zhang L, Tian SF, Gu MH and Chen
AP: Effect of equiguard in treating patients with shen-yang
deficiency syndrome. Chin J Integr Med. 14:225–227. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lang JM, Li MZ and Wei AS: Study on
objective parameters of syndrome differentiation of diabetic
nephropathy. Zhongguo Zhong Xi Yi Jie He Za Zhi. 15:84–86. 1995.(In
Chinese). PubMed/NCBI
|
|
5
|
Deng ZZ, He YT and Yu YM: Comparison
between two diagnostic methods of computer's mathematic model and
clinical diagnosis on TCM syndromes of rheumatoid arthritis.
Zhongguo Zhong Xi Yi Jie He Za Zhi. 16:727–729. 1996.(In Chinese).
PubMed/NCBI
|
|
6
|
Gu WL, Shi ZX, Yu YX, Wu YW, Lu BW and Hui
KK: Distribution characteristics of syndrome types in essential
hypertension. Zhong Xi Yi Jie He Xue Bao. 8:842–847. 2010.(In
Chinese). View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu Y, Wu WK, Chen C and Duan XF: Effect
of delayed preconditioning induced by Sini decoction on myocardial
cell apoptosis and its mitochondrial mechanism. Zhongguo Zhong Yao
Za Zhi. 31:1269–1272. 2006.(In Chinese). PubMed/NCBI
|
|
8
|
Zhao MQ, Wu WK, Zhao DY, Duan XF and Liu
Y: Protective effects of sini decoction on adriamycin-induced heart
failure and its mechanism. Zhong Yao Cai. 32:1860–1863. 2009.(In
Chinese). PubMed/NCBI
|
|
9
|
Zhao L, Wu H, Qiu M, Sun W, Wei R, Zheng
X, Yang Y, Xin X, Zou H, Chen T, et al: Metabolic signatures of
kidney yang deficiency syndrome and protective effects of two
herbal extracts in rats using GC/TOF MS. Evid Based Complement
Alternat Med. 2013:5409572013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang D, Yang J, Lu X, Deng Y, Xiong Z and
Li F: An integrated plasma and urinary metabonomic study using
UHPLC-MS: Intervention effects of Epimedium koreanum on
‘Kidney-Yang Deficiency syndrome’ rats. J Pharm Biomed Anal.
76:200–206. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tan Y, Liu X, Lu C, He X, Li J, Xiao C,
Jiang M, Yang J, Zhou K, Zhang Z, et al: Metabolic profiling
reveals therapeutic biomarkers of processed aconitum carmichaeli
debx in treating hydrocortisone induced kidney-yang deficiency
syndrome rats. J Ethnopharmacol. 152:585–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tan G, Liao W, Dong X, Yang G, Zhu Z, Li
W, Chai Y and Lou Z: Metabonomic profiles delineate the effect of
traditional Chinese medicine sini decoction on myocardial
infarction in rats. PLoS One. 7:e341572012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
National Pharmacopeia Committee of China.
Chinese Pharmacopeia 2010 Edition: 650–660. 2010.
|
|
14
|
Munakata S and Hendricks JB: Effect of
fixation time and microwave oven heating time on retrieval of the
ki-67 antigen from paraffin-embedded tissue. J Histochem Cytochem.
41:1241–1246. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Verweij CL, Geerts M and Aarden LA:
Activation of interleukin-2 gene transcription via the T-cell
surface molecule CD28 is mediated through an NF-kB-like response
element. J Biol Chem. 266:14179–14189. 1991.PubMed/NCBI
|
|
18
|
Grewal SS, Fass DM, Yao H, Ellig CL,
Goodman RH and Stork PJ: Calcium and cAMP signals differentially
regulate cAMP-responsive element-binding protein function via a
rap1-extracellular signal-regulated kinase pathway. J Biol Chem.
275:34433–34441. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chun KS and Surh YJ: Signal transduction
pathways regulating cyclooxygenase-2 expression: Potential
molecular targerts for chemoprevention. Biochem Pharmacol.
68:1089–1100. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tanabe T and Tohnai N: Cyclooxygenase
isozymes and their gene structures and expression. Prostaglandins
Other Lipid Mediat. 68–69:95–114. 2002. View Article : Google Scholar
|
|
21
|
Sun W, Yin XJ, Tu Y, Wan YG, Liu H and Hu
H: Effects and mechanisms of Qifu decoction ameliorating renal
tubulointerstitial fibrosis through inhibiting ERK1/2 signaling
pathway in unilateral ureteral obstruction rats with Yang
deficiency. Zhongguo Zhong Yao Za Zhi. 39:4082–4089. 2014.(In
Chinese). PubMed/NCBI
|
|
22
|
Zhang JH, Xin J, Fan LX and Yin H:
Intervention effects of Zuoguiwan containing serum on osteoblast
through ERK1/2 and Wnt/β-catenin signaling pathway in models with
kidney-Yang-deficiency, kidney-Yin-deficiency osteoporosis
syndromes. Zhongguo Zhong Yao Za Zhi. 42:3983–3989. 2017.(In
Chinese). PubMed/NCBI
|
|
23
|
Huang R, Zhang Z, Xu M, Chang X, Qiao Q,
Wang L and Meng X: Effect of Sini decoction on function of
hypothalamic-pituitary-adrenal axis in patients with sepsis.
Zhonghua Wei Zhong Bing Ji Jiu Yi Xue. 26:184–187. 2014.(In
Chinese). PubMed/NCBI
|
|
24
|
Hla T and Neilson K: Human
cyclooxygenase-2 cDNA. Proc Natl Acad Sci USA. 89:7384–7388. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jia Z, Zhang Y, Ding G, Heiney KM, Huang S
and Zhang A: Role of COX-2/mPGES-1/prostaglandin E2 cascade in
kidney injury. Mediators Inflamm. 2015:1478942015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sha WC: Regulation of immune responses by
NF-kappa B/Rel transcription factor. J Exp Med. 187:143–146. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao M, Wu W, Duan X, Liu Y, Zhao D, Liang
T and Luo H: The protective effects of sini decoction on
mitochondrial function in adriamycin-induced heart failure rats.
Zhong Yao Cai. 28:486–489. 2005.(In Chinese). PubMed/NCBI
|
|
28
|
Nie Y, Wu W, Liu Y, Duan X, Zhao M and
Zhao D: Protective effects of sini decoction on cardiomyocytes in
oxidative stress damage induced by peroxide. Zhong Yao Cai.
28:395–399. 2005.(In Chinese). PubMed/NCBI
|
|
29
|
Shen ZY: Rule of tonifying the kidney in
regulating T lymphocyte apoptosis in syndrome of kidney-yang
deficiency-plasticity of gene balance. Zhong Xi Yi Jie He Xue Bao.
2:321–322. 2004.(In Chinese). View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Goldberg ND, Haddox MK, Nicol SE, Glass
DB, Sanford CH, Kuehl FA Jr and Estensen R: Biologic regulation
through opposing influences of cyclic GMP and cyclic AMP: The Yin
Yang hypothesis. Adv Cyclic Nucleotide Res. 5:307–330.
1975.PubMed/NCBI
|