Introduction
Gastric cancer (GC) is common in developing
countries (1). In 2012, there were
700,000 mortalities, making GC the third leading cause of
cancer-associated mortality after lung and liver cancer (2). Worldwide, ~42% of male and ~19% of
female patients with GC are from China (3). Risk factors for GC include stress, a
diet of charred or salty foods, alcohol consumption, smoking,
obesity and physical inactivity (4).
Current treatments for GC include surgery (5), chemotherapy and/or radiation therapy
(6). The outcomes of patients with
locally advanced GC are sub-optimal due to the likelihood of
relapse (hazard ratio=1.52) (7) and
chemotherapy with external beam radiotherapy is the standard
adjuvant treatment (8).
Radio-resistance makes the successful treatment of GC challenging
(9). Target therapy is a novel
strategy currently being studied in clinical trials (10). Thus, the identification of novel
molecular targets is required to fulfill unmet clinical needs.
B-cell lymphoma 2 (Bcl-2) is an anti-apoptotic
member of the Bcl-2 family, which regulates cell apoptosis
(11). Deregulation of apoptosis is
strongly associated with various diseases, such as schizophrenia,
which is thought to result from the decreased expression of Bcl-2
and increased expression of caspase-3 (12).
MicroRNA (miRNA or miR) are a class of non-coding
RNA that are ~22 nucleotides long, which act as endogenous
suppressors of gene expression by binding to the 3′-untranslated
region (3′-UTR) of target mRNA (13,14).
miRNA serve a role in cell proliferation and apoptosis, which are
associated with tumorigenesis (15).
The regulation of Bcl-2 by miRNA and its role in GC progression
remain unknown and further investigation is required to identify
the pathogenic mechanism of GC.
The present study aimed to investigate the function
of miR-744 in GC and the possible molecular mechanisms. RT-qPCR
analyses were performed on tumor and adjacent tissues obtained from
30 patients to explore the differencential expression of miR-744.
SGC-7901 cells were randomly divided into control, miR-negative
control (NC) and miR-744 mimic groups. Cell Counting Kit-8 (CCK-8)
assay and flow cytometry were performed to determine the cell
proliferation and cell apoptosis of SGC-7901 cells, respectively.
Western blotting was performed to evaluate the expression of Bcl-2,
B cell lymphoma-2-associated X protein (Bax) and caspase-3. A
dual-luciferase assay was used for verification of the interaction
between miR-744 and Bcl-2. It was found that miR-744 serves a tumor
suppressive role in GC by targeting Bcl-2.
Materials and methods
Patients
GC tumor tissues and adjacent normal tissues from 30
patients (14 males and 16 females; mean age, 47.30±9.82 years) who
underwent surgery on the stomach or esophageal junction at the
Dongying People's Hospital (Dongying, China) were collected between
November 2014 and November 2016 for the current study. The adjacent
tissues were removed at a distance of ≥5 cm away from the tumor
tissues from the antrum and corpus mucosa, which were stripped
along the side of the lesser curvature. Prior informed and written
consent was obtained from each patient. Patients who underwent
chemotherapy or radiotherapy prior to surgery were excluded from
the current study. The present study was approved by the Ethics
Committee of Dongying People's Hospital.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
miRNA from patient tissues were extracted and lysed
using an miRNeasy Mini kit (Qiagen China Co., Ltd., Shanghai,
China). RT for hsa-miR-744 was performed using a Transcriptor First
Strand cDNA Synthesis kit (Roche Diagnostics, Basel, Switzerland)
at 65°C for 1 h. Subsequently, cDNA was used for qPCR, which was
performed using a SYBR-Green Master mix (ROX; Roche Diagnostics) in
a PCR Thermal Cycler Dice (Takara Bio, Inc., Otsu, Japan) at 95°C
for 10 min, followed by 50 cycles of 95°C for 10 sec, 55°C for 10
sec, 72°C for 5 sec, 99°C for 1 sec, 59°C for 15 sec, 95°C for 1
sec and final cooling to 40°C. Relative expression of miR-744 was
normalized against U6 with the 2−ΔΔCq method (16). Primer sequences were as follows:
miR-744 forward, 5′-AATGCGGGGCTAGGGCTA-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; and U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and
reverse, 5′-AACGCTTCACGAATTTGCGT-3′ (17).
Cell culture
Human GC cell line SGC-7901 (American Type Culture
Collection, Manassas, VA, USA) was cultured in RPMI-1640/F12 medium
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% (v/v) fetal bovine serum (FBS; Invitrogen;
Thermo Fisher Scientific, Inc.), streptomycin (100 mg/ml) and
penicillin (100 U/ml) at 37°C in a 95% humidified chamber
containing 5% CO2.
Cell transfection
SGC-7901 cells (1×105) were first seeded
onto 24-well plates and cultured for 24 h at 37°C. Cells were
divided into the following groups: The control group consisting of
untreated SGC-7901 cells; the miR-NC mimics group consisting of
SGC-7901 cells transfected with miR-NC mimics, and the miR-744
mimic group consisting of SGC-7901 cells transfected with miR-744
mimics. The miR-744 mimics and miR-NC mimics were purchased from
GeneCopoeia, Inc. (Rockville, MD, USA). Cell transfection of 30 µM
miR-744 mimics or miR-NC mimics was performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) and cells were incubated for 48 h at 37°C in
accordance with the manufacturer's protocols prior to the
performance of subsequent experimentation. The sequences used were
as follows: miR-744 mimics forward, 5′-UGCGGGGCUAGGGCUAACAGCA-3′
and reverse, 5′-UGCUGUUAGCCCUAGCCCCGCA-3′; and miR-NC mimics
forward, 5′-AAUUCUCCGAACGUGUCACTT-3′ and reverse,
5′-GUGACACGUUCGGAGAAUUTT-3′, according to a previous study
(18).
Western blot analysis
SGC-7901 cells were first washed with PBS three
times, 5 min each time, and lysed using lysis buffer containing
protease inhibitors (Beyotime Institute of Biotechnology, Haimen,
China). The concentration of each protein sample was normalized
using the bicinchoninic acid method. Equal amounts of proteins from
all the obtained samples (15 µg/lane) were loaded and then
separated by 8–10% SDS-PAGE. Samples were then transferred on to
nitrocellulose membranes. Membranes were blocked using 5% (v/v)
skimmed milk for 1 h at room temperature and then incubated and
probed with the following primary antibodies: Bax (cat. no. 2772),
Bcl-2 (cat. no. 3498), caspase-3 (cat. no. 9662) and GAPDH (cat.
no. 5174; all 1:1,000; Cell Signaling Technology, Inc., Danvers,
MA, USA) at 4°C overnight. Membranes were then incubated with
bovine anti-rabbit horseradish peroxidase-conjugated secondary
antibodies (cat. no. 2370; 1:1,000; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) at room temperature for 2 h. Bands were developed
using an enhanced chemiluminescent kit (Beyotime Institute of
Biotechnology, Shanghai, China) with GAPDH acting as an endogenous
loading control.
Cell proliferation assay
SGC-7901 cell proliferation rate was assessed using
CCK-8 (Dojindo Molecular Technologies, Inc., Kumamoto, Japan)
according to the manufacturer's protocol. Briefly, SGC-7901 cells
were seeded in each well of a 96-well plate at a concentration of
5×103 cells/well and cultured at 37°C for 1–5 days. A
total of 10 µl CCK-8 solution was then added into each well and
incubated for 1 h at 37°C. The optical density value from each
sample was read at a wavelength of 450 nm using a microplate
reader.
Cell apoptosis analysis
SGC-7901 cell apoptosis was evaluated using an
Annexin V-fluorescein isothiocyanate/propidium iodide (FITC/PI)
double-staining apoptosis detection kit (Beyotime Institute of
Biotechnology) in accordance with the manufacturer's protocols.
FITC specifically binds to the phosphatidyl serine residues on the
cell membrane, while PI binds to DNA when the cell membrane is
permeable (19). SGC-7901 cells were
collected at 48 h following transfection and resuspended in 400 µl
of 90% RPMI-1640 medium with 10% FBS to the final concentration of
1×106 cells/ml. A total of 5 µl Annexin V-FITC was then
added to the suspension and SGC-7901 cells were incubated at room
temperature in the dark for 15 min prior to the addition of 5 µl
PI. After 5 min, SGC-7901 cells were analyzed using a flow
cytometer and quantified using FlowJo software version 7.6.2
(FlowJo LLC, Ashland, OR, USA).
Luciferase reporter assay
Wild-type (WT) or mutant Bcl-2 3′-UTR was cloned
into a pmirGLO luciferase reporter vector (Promega Corporation,
Madison, WI, USA). Following the incubation of SGC-7901 cells at
37°C for 24 h, SGC-7901 cells were co-transfected with miR-744
mimics and pmir-Bcl-2WT-3′-UTR or pmir-Bcl-2mut-3′-UTR using
Lipofectamine® 2000, according to the manufacturer's
protocol. SGC-7901 cells were then seeded in 24-well plates at a
concentration of 5×104 cells/well and cultured at 37°C
for 18 h. Subsequently, SGC-7901 cells were transfected with
firefly luciferase vector (500 ng/µl) and mixed with Vigofect
transfection reagent (Vigorous Biotechnology, Beijing, China),
according to the manufacturer's protocol. Following continuous
exposure for 48 h, the luciferase activities from firefly and
Renilla were measured with a Dual-luciferase reporter assay
system (Promega Corporation), with Renilla activity acting
as a normalized parameter.
Statistical analysis
All data are expressed as the mean ± standard
deviation and all experiments were repeated three times. GraphPad
Prism 5 software (GraphPad Software, Inc., La Jolla, CA, USA) and a
Student's two-sample unpaired t-test were used for the analyses of
data, while data among three groups were compared by one-way
analysis of variance followed by a Tukey's post hoc test. A
Pearson's correlation test was used to perform correlation analysis
between miR-744 expression and GC progression in Table I. P<0.05 was considered to
indicate a statistically significant difference.
 | Table I.Clinicopathological characteristics
and expression of miR-774 in tissues from patients with gastric
cancer. |
Table I.
Clinicopathological characteristics
and expression of miR-774 in tissues from patients with gastric
cancer.
|
| Characteristics |
|---|
|
|
|
|---|
| Patient number | Sex | Age, years | TNM stage | Relative miR-774
level |
|---|
| 1 | F | 57 | IV | 0.40 |
| 2 | F | 30 | I A | 1.21 |
| 3 | M | 38 | III A | 0.93 |
| 4 | F | 46 | II | 1.01 |
| 5 | M | 39 | I A | 1.20 |
| 6 | F | 54 | II | 1.10 |
| 7 | M | 62 | III A | 0.97 |
| 8 | M | 37 | IV | 0.52 |
| 9 | F | 60 | III B | 0.88 |
| 10 | M | 58 | III B | 0.64 |
| 11 | M | 39 | III B | 0.78 |
| 12 | M | 34 | III B | 0.84 |
| 13 | F | 45 | IV | 0.45 |
| 14 | F | 28 | I A | 1.23 |
| 15 | M | 32 | I A | 1.32 |
| 16 | M | 45 | III A | 0.91 |
| 17 | F | 51 | III B | 0.74 |
| 18 | M | 36 | IV | 0.62 |
| 19 | F | 42 | IV | 0.60 |
| 20 | F | 52 | IV | 0.47 |
| 21 | F | 54 | III A | 0.98 |
| 22 | F | 46 | I B | 1.21 |
| 23 | M | 49 | III B | 0.78 |
| 24 | F | 52 | III B | 0.75 |
| 25 | M | 57 | IV | 0.56 |
| 26 | M | 55 | III B | 0.67 |
| 27 | F | 53 | III B | 0.85 |
| 28 | M | 47 | III A | 0.95 |
| 29 | F | 59 | IV | 0.48 |
| 30 | F | 62 | II | 1.04 |
Results
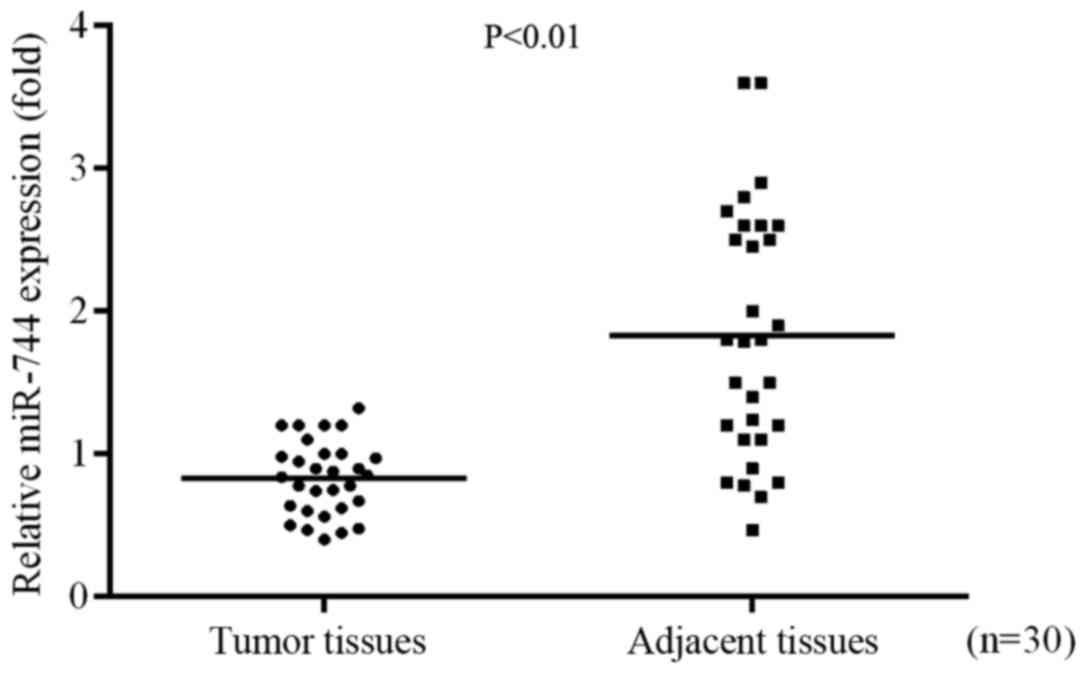
miR-744 expression levels are
decreased in GC tissues
RT-qPCR was used to identify the role of miR-744 in
GC progression by determining the differential expression of
miR-744 in GC tissues and adjacent tissues. The results
demonstrated that the expression of miR-744 was significantly
decreased in tumor tissues compared with the level in adjacent
normal tissues (Fig. 1).
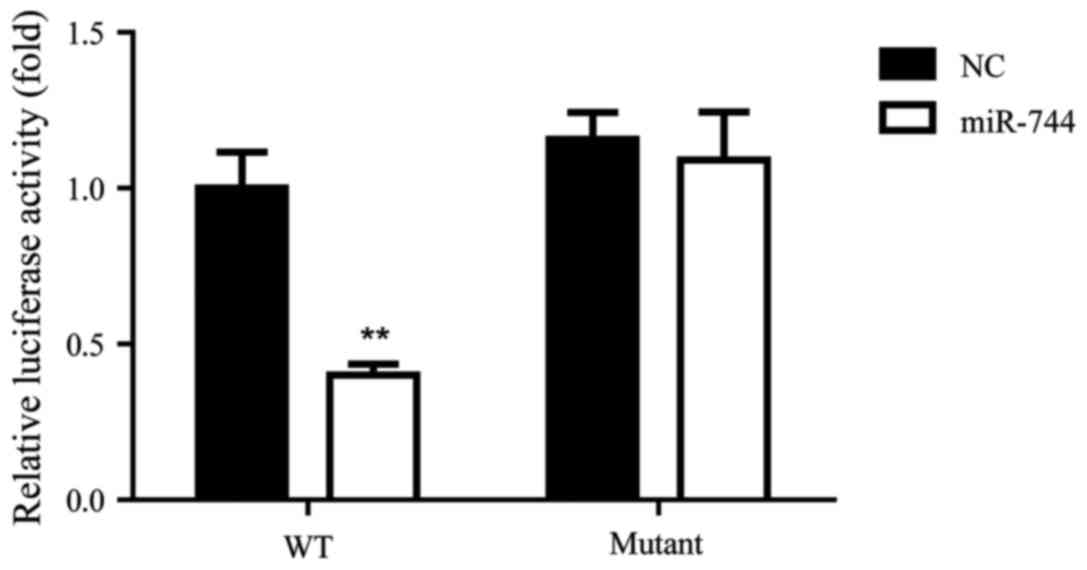
Bcl-2 is a direct target of miR-744 in
GC
A previous study indicated that miR-744 was involved
in cervical cancer through targeting Bcl-2 (16). Therefore, the current study aimed to
determine whether miR-744 regulated Bcl-2 in GC using a luciferase
target assay for the detection of an association between miR-744
and Bcl-2 in SGC-7901 cells. The results indicated that miR-744
significantly decreased the relative luciferase activity of the WT
Bcl-2 3′-UTR compared with that observed in the NC group, while the
luciferase activity of the mutant Bcl-2 3′-UTR was not
significantly different between the NC and miR-744 groups (Fig. 2). These results suggest that Bcl-2
was a direct target of miR-744 in SGC-7901 cells.
Expression profile of miR-744 in
tissues from patients with GC
There were 14 male and 16 female patients included
in the current study. The mean age of patients was 47.30±9.82 years
and 16 patients were aged <50 years old, while 14 patients were
aged ≥50 years old. The tumor, node and metastasis (TNM) staging
system according to seventh edition of the American Joint Committee
on Cancer TNM Classification was used for this study (20). There were 5 patients at histological
stage I (4 IA and 1 IB), 3 patients (the lowest rate) at
histological stage II, 14 patients (the highest rate) at
histological stage III (5 IIIA and 9 IIIB) and 8 patients (the
highest rate) at histological stage IV. The expression profile of
miR-744 was also determined, and it was demonstrated that the more
serious the TNM stage of patients the lower the miR-744 mRNA level,
suggesting that miR-744 was negatively associated with the
progression of GC (Table I). This
association requires further study and verification in a larger
sample size.
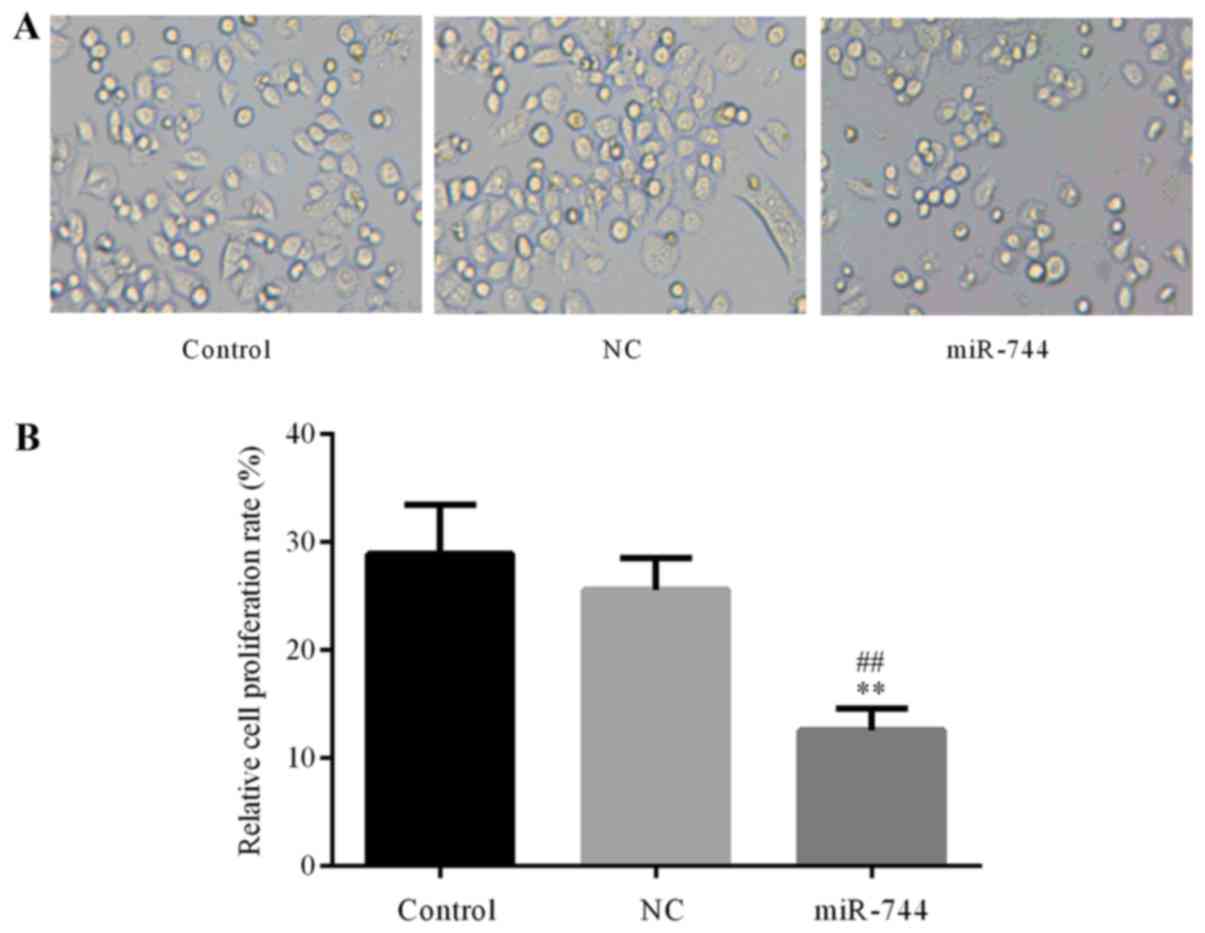
miR-744 inhibits SGC-7901 cell
proliferation
The role of miR-744 in GC cell proliferation was
further confirmed using a CCK-8 assay to determine SGC-7901 cell
proliferation in the different groups. The results demonstrated
that the proliferation of SGC-7901 cells was significantly
decreased in the miR-744 mimics group compared with that observed
in the control and miR-NC mimics groups (Fig. 3). These results suggest that miR-744
inhibited cell proliferation in GC cells.
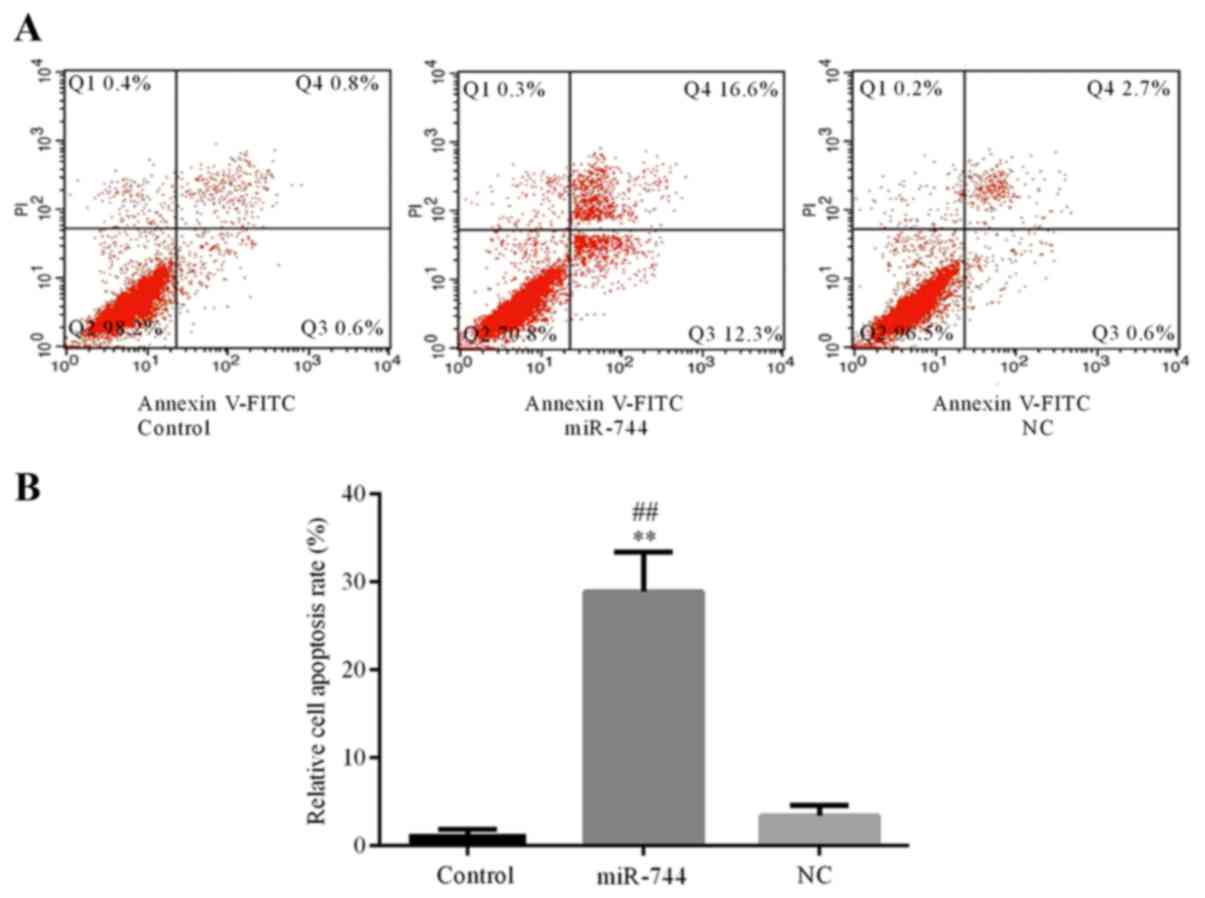
miR-744 induces SGC-7901 cell
apoptosis
Flow cytometry was used to determine the apoptosis
rate of SGC-7901 cells in the different groups. The results
demonstrated that cell apoptosis was significantly increased in the
miR-744 mimics group compared with that observed in the control and
miR-NC mimics groups (Fig. 4). These
results suggest that miR-744 promotes cell apoptosis in GC
cells.
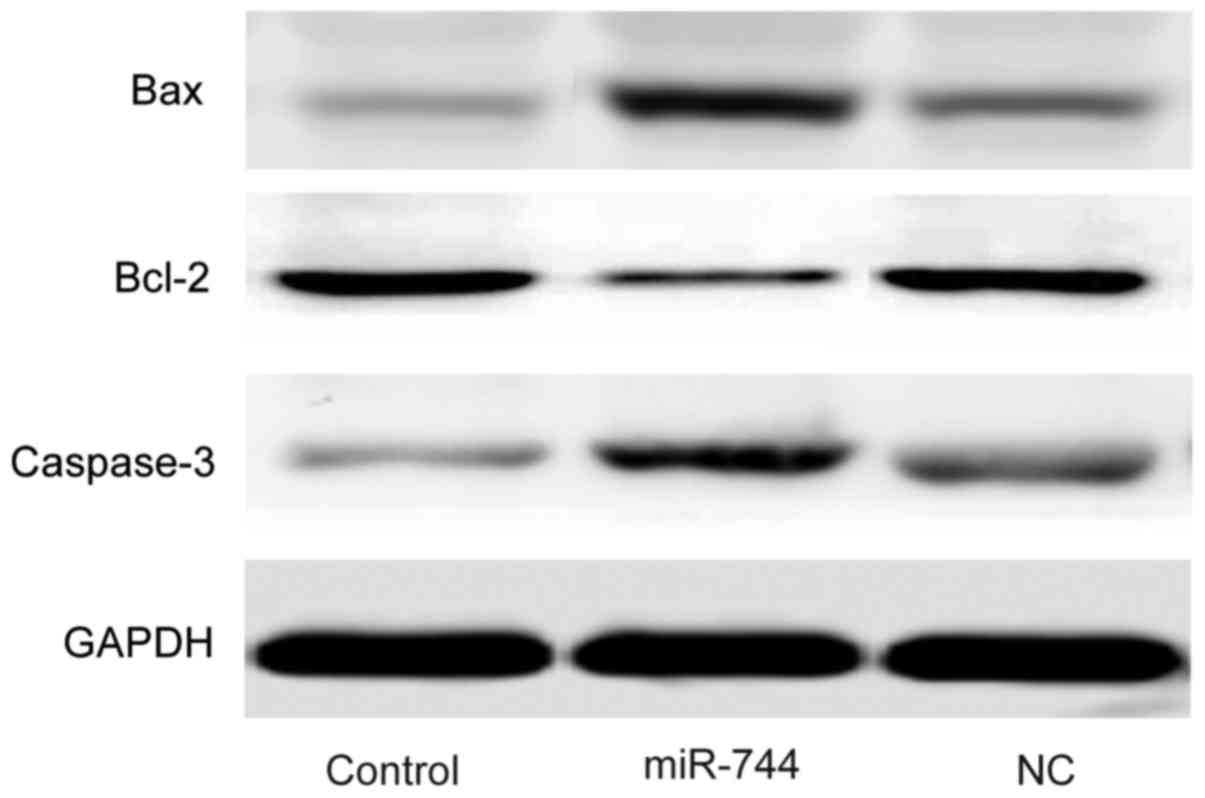
miR-744 decreases the expression of
anti-apoptosis protein Bcl-2 and increases the expression of
pro-apoptosis proteins Bax and caspase-3 in SGC-7901 cells
Western blot analysis was used to determine the
expression levels of Bcl-2, Bax and caspase-3 in SGC-7901 cells. It
was demonstrated that the expression levels of Bax and caspase-3
were markedly increased while the expression of Bcl-2 was decreased
in the miR-744 mimics group compared with the levels observed in
the control and miR-NC mimics groups (Fig. 5).
Discussion
GC is common in developing countries (1). Targeted therapy is a novel treatment
strategy for GC, which is being studied in clinical trials
(10). However, due to the
heterogeneity of GC, the identification of potential targets is
required to fulfill the unmet clinical need.
miRNA are endogenous suppressors of gene expression,
which bind to mRNA 3′-UTRs (13,14) and
are associated with cell proliferation and apoptosis (15). Apoptosis has been demonstrated to
modulate schizophrenia, which may be caused by decreased expression
of Bcl-2 and increased expression of caspase-3 (12). miRNA that may regulate Bcl-2 were
investigated in the present study. Bcl-2 is an integral protein of
the external mitochondrial membrane, which reduces cell apoptosis
(21). Upregulation of the
expression of Bcl-2 by secretory leukocyte protease inhibitor has
been demonstrated to promote the proliferation of GC cells
(22).
A previous study indicated that miR-744 targets
Bcl-2 (23), thus it was chosen as
the focus of the current study. The differential expression of
miR-744 between tumor tissues and adjacent normal tissues was
investigated in the present study. RT-qPCR demonstrated that the
expression level of miR-744 was significantly decreased in tumor
tissues compared with the level in adjacent normal tissues. These
results suggest that miR-744 may be a tumor suppressor gene, which
is consistent with a previous study that indicated the decreased
expression of miR-744 in human hepatocellular carcinoma (17). Notably, miR-744 has been suggested to
inhibit various types of cancer, including colon cancer, breast
cancer and GC (24). Furthermore,
the relationship between Bcl-2 and miR-744 was demonstrated using a
dual-luciferase assay in SGC-7901 cells in the current study, which
indicated that miR-744 significantly reduced the relative
luciferase activity of the WT Bcl-2 3′-UTR, while the luciferase
activity of the mutant Bcl-2 3′-UTR was not significantly
changed.
The expression of miR-744 in patients with GC at
different TNM stages was also examined. The results demonstrated
that the more serious the TNM stage of patients the lower the
miR-744 mRNA level, the expression of miR-744 was negatively
associated with the progression of GC. This association requires
further study and verification in a larger sample size. Thus, the
results of the current study indicated that miR-744 serves a role
in inhibiting the progression of GC in patients, with Bcl-2 as the
target gene in GC cells.
The impact of miR-744 on GC cell behavior was
further investigated by determining the cell proliferation and
apoptosis rates of SGC-7901 cells using a CCK-8 assay and flow
cytometry, respectively. The results of the CCK-8 assay
demonstrated that the proliferation of SGC-7901 cells was
significantly decreased in the miR-744 mimics group compared with
the control and miR-NC mimics groups. In addition, flow cytometry
analysis indicated that apoptosis of SGC-7901 cells was
significantly increased in the miR-744 mimics group compared with
the control and miR-NC mimics groups.
However, the molecules regulated by miR-744 remained
unclear. Therefore, western blot analysis was performed and the
results indicated that the expression levels of Bax and caspase-3
were increased, while the expression of Bcl-2 was decreased in the
miR-744 mimics group compared with the levels observed in the
control and miR-NC mimics groups in SGC-7901 cells. The
differential expression of Bcl-2, Bax and caspase-3 were consistent
with the results that were indicated for SGC-7901 cell apoptosis
and proliferation. Caspase-3 is an essential indicator of apoptosis
(25) and Bax activates cell
apoptosis by activating the activity of caspase-3 (26).
In conclusion, the results of the current study
demonstrated that miR-744 was downregulated in GC tissues compared
with normal gastric tissues; miR-744 decreased the expression of
Bcl-2 and SGC-7901 cell proliferation, while it increased the
expression of caspase-3, Bax and the SGC-7901 cell apoptosis rate.
Thus, miR-744 may serve as a tumor suppressor via the inhibition of
cell proliferation and promotion of apoptosis by targeting Bcl-2 in
GC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
Availability of data and materials: All data
generated or analyzed during this study are included in this
published article.
Authors' contributions
JL designed and conducted the experiments, and
analyzed the data. YW and SL conducted the experiments and analyzed
the data. YL and HL conducted the experiments. JL analyzed the
data. XZ designed the experiments, analyzed the data and wrote the
manuscript.
Ethics approval and consent to
participate
Prior informed and written consent was obtained from
each patient. The present study was approved by the Ethics
Committee of Dongying People's Hospital.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lozano R, Naghavi M, Foreman K, Lim S,
Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, et
al: Global and regional mortality from 235 causes of death for 20
age groups in 1990 and 2010: A systematic analysis for the global
burden of disease study 2010. Lancet. 380:2095–2128. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
An JY, Kim KM, Kim YM, Cheong JH, Hyung WJ
and Noh SH: Surgical complications in gastric cancer patients
preoperatively treated with chemotherapy: Their risk factors and
clinical relevance. Ann Surg Oncol. 19:2452–2458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oh DY, Choi KS, Shin HR and Bang YJ:
Public awareness of gastric cancer risk factors and disease
screening in a high risk region: A population-based study. Cancer
Res Treat. 41:59–66. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen K, Xu XW, Zhang RC, Pan Y, Wu D and
Mou YP: Systematic review and meta-analysis of laparoscopy-assisted
and open total gastrectomy for gastric cancer. World J
Gastroenterol. 19:5365–5376. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pretz JL, Wo JY, Mamon HJ, Kachnic LA and
Hong TS: Chemoradiation therapy: Localized esophageal, gastric, and
pancreatic cancer. Surg Oncol Clin N Am. 22:511–524. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Macdonald JS, Smalley SR, Benedetti J,
Hundahl SA, Estes NC, Stemmermann GN, Haller DG, Ajani JA,
Gunderson LL, Jessup JM and Martenson JA: Chemoradiotherapy after
surgery compared with surgery alone for adenocarcinoma of the
stomach or gastroesophageal junction. N Eng J Med. 345:725–730.
2001. View Article : Google Scholar
|
|
8
|
Wang J, Tian Y, Tang Y, Wang X, Li N, Ren
H, Fang H, Feng Y, Wang S, Song Y, et al: A Phase II prospective
nonrandomized trial of magnetic resonance imaging-guided
hematopoietic bone marrow-sparing radiotherapy for gastric
cancerpatients with concurrent chemotherapy. Onco Targets Ther.
9:2701–2707. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kang M, Xiao J, Wang J, Zhou P, Wei T,
Zhao T and Wang R: MiR-24 enhances radiosensitivity in
nasopharyngeal carcinoma by targeting SP1. Cancer Med. 5:1163–1173.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meza-Junco J, Au HJ and Sawyer MB:
Critical appraisal of trastuzumab in treatment of advanced stomach
cancer. Cancer Manag Res. 3:57–64. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Azimian H, Bahreyni-Toossi MT, Rezaei AR,
Rafatpanah H, Hamzehloei T and Fardid R: Up-regulation of Bcl-2
expression in cultured human lymphocytes after exposure to low
doses of gamma radiation. J Med Phys. 40:38–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Glantz LA, Gilmore JH, Lieberman JA and
Jarskog LF: Apoptotic mechanisms and the synaptic pathology of
schizophrenia. Schizophr Res. 81:47–63. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Van Rooij E: The art of microRNA research.
Circ Res. 108:219–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Munker R and Calin GA: MicroRNA profiling
in cancer. Clin Sci (Lond). 121:141–158. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong
Q, Qin L, Wu X, Zheng Y, Yang Y, et al: Identification of miRNomes
in human liver and hepatocellular carcinoma reveals miR-199a/b-3p
as therapeutic target for hepatocellular carcinoma. Cancer Cell.
19:232–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin F, Ding R, Zheng S, Xing D, Hong W,
Zhou Z and Shen J: Decrease expression of microRNA-744 promotes
cell proliferation by targeting c-Myc in human hepatocellular
carcinoma. Cancer Cell Int. 14:582014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ding J, Yuan L and Chen G: Aspirin
enhances the cytotoxic activity of bortezomib against myeloma cells
via suppression of Bcl-2, survivin and phosphorylation of AKT.
Oncol Lett. 13:647–654. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roy M, Chakraborty S, Siddiqi M and
Bhattacharya RK: Induction of apoptosis in tumor cells by natural
phenolic compounds. Asian Pac J Cancer Prev. 3:1–67.
2002.PubMed/NCBI
|
|
22
|
Du XY, Liu X, Wang ZJ and Wang YY: SLPI
promotes the gastric cancer growth and metastasis by regulating the
expression of P53, Bcl-2 and caspase-8. Eur Rev Med Pharmacol Sci.
21:1495–1501. 2017.PubMed/NCBI
|
|
23
|
Chen XF and Liu Y: MicroRNA-744 inhibited
cervical cancer growth and progression through apoptosis induction
by regulating Bcl-2. Biomed Pharmacother. 81:379–387. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song MY, Pan KF, Su HJ, Zhang L, Ma JL, Li
JY, Yuasa Y, Kang D, Kim YS and You WC: Identification of serum
microRNAs as novel non-invasive biomarkers for early detection of
gastric cancer. PLoS One. 7:e336082012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Basu S, Rajakaruna S, De Arcangelis A,
Zhang L, Georges-Labouesse E and Menko AS: α6 integrin
transactivates insulin-like growth factor receptor-1 (IGF-1R) to
regulate caspase-3-mediated lens epithelial cell differentiation
initiation. J Biol Chem. 289:3842–3855. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Z and Wang Z: Effects of rapamycin on
expression of Bcl-2 and Bax in human lens epithelial cells and cell
cycle in rats. J Huazhong Univ Sci Technolog Med Sci. 31:555–559.
2011. View Article : Google Scholar : PubMed/NCBI
|