Introduction
Osteosarcoma (OS) is one of the most prevalent and
aggressive bone tumors and leads to high mortality rates in
children and adolescents (1). OS
accounts for approximately 66% cases of bone cancers worldwide
(2). Although great effort has been
made in the past decades, advance achievement on OS treatment
remains very limited, and the outcomes of patients with OS are
quite poor (3). Nowadays, surgical
resection combined with chemotherapy and radiotherapy is the main
approach for OS treatment. However, this method has a poor effect
on advanced or metastatic OS (4).
Therefore, searching novel biomarkers for early diagnosis of
patients with OS and effective targets for OS therapy is urgently
required.
MicroRNAs (miRNAs) are a class of short noncoding
RNAs of approximately 22–25 nucleotides in length and has been
demonstrated to regulate gene expression by binding to the three
prime untranslated region (3′-UTR) of target mRNAs for degradation
(5). In the past decades, large
numbers of studies showed that miRNAs exert very important
functions in almost all kinds of biological processes, such as cell
survival, proliferation, migration, and invasion (6–8).
Especially in cancer, the miRNA functions have been widely
explored. For example, Du et al (9), reported that microRNA-509-3p represses
glioma cell proliferation and invasion. Wang et al (10), showed that microRNA-124-3p suppresses
the growth and metastasis of cervical cancer. In OS, several miRNAs
have also been reported to exert essential functions, such as
microRNA-643 (11), microRNA-138
(12), and microRNA-27a (13).
Two recent reports indicate that miR-466 inhibits
tumor growth and metastasis in prostate cancer and colorectal
cancer (14,15). Nevertheless, the function of miR-466
in OS remains elusive. In the present study, we found that miR-466
was significantly downregulated in OS tissues. The expression level
of miR-466 was negatively associated with OS severity. Through
functional experiments, we demonstrated that miR-466 suppressed OS
cell proliferation and induced cellular apoptosis. Mechanistically,
we found that CCND1 was a target of miR-466 in OS cells. CCND1
restoration abrogated the effects of miR-466 transfection in OS
cells. In a word, our study demonstrated that miR-466 served as a
tumor suppressor through targeting CCND1 and implied that miR-466
might be a promising prognostic biomarker and therapeutic
target.
Materials and methods
Cell lines and clinical specimens
The OS cell lines used in the present study,
including 143B, U2OS, KHOS-240S, Saos-2 and MG-63, and the normal
cell line (hFOB1.19) were purchased from The Shanghai Institute of
Cell Biology (Shanghai, China). The cells were maintained in
RPMI-1640 medium supplemented by 10% fetal bovine serum (FBS),
streptomycin (100 µg/ml) and penicillin (200 U/ml; all from
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in 5% CO2
at 37°C. A total of 87 surgically resected OS specimens and
adjacent normal tissues were acquired from patients at Beijing
Rehabilitation Hospital of Capital Medical University between June
2013 and August 2016. All patients signed formal consent forms. The
experiments involving human specimens were reviewed and approved by
the Ethics Committee of Beijing Rehabilitation Hospital of Capital
Medical University.
Cell transfection
For CCND1 overexpression, the coding sequence of
CCND1 was constructed into pCDNA3-myc vector (pCDNA3-myc-CCND1).
The miR-466 mimics, miR-466 inhibitor and negative controls were
synthesized and purchased from Sigma-Aldrich; Merck KGaA.
Corresponding plasmids were transfected into 143B and U2OS cells
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. After incubation for 48 h, cells were used
for further analysis.
Cell proliferation assay
For cell proliferation assays, the viable cells were
tested by Cell Counting Kit-8 (CCK-8) assay kit according to the
manufacturer's instructions. In brief, cells were grown in 96-well
plate with 1×104 per well and incubated in 37°C with 5%
CO2 until cell confluent rate reached 70%. After
transfected with plasmid for 48 h, cells were still incubated for
24, 48, 72 and 96 h. 10 µl CCK8 solution was seed into each well.
The absorbance at 490 nm was measured with SUNRISE Microplate
Reader (Tecan Group, Ltd., Mannedorf, Switzerland).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissues and cells using
the RNeasy Plus mini kit (Qiagen GmbH, Hilden, Germany), according
to the manufacturer's instructions. RNA was then reversely
transcribed into complementary DNA using the PrimeScript RT-PCR kit
(Takara Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's instructions. For quantification, PCR was performed
using a miRNA Q-PCR Detection kit (GeneCopoeia, Inc., Rockville,
MD, USA) on an ABI 7500 thermocycler (Thermo Fisher Scientific,
Inc.). U6 or GAPDH was used as an internal reference. The relative
expression of mRNA was determined using the 2−∆∆Cq
method (16). The primer sequences
were as follows: miR-466 (5′-AACGAGACGACGACAGAC-3′ and
5′-ATACACATACACGCAACACACAT-3′), U6 (5′-AACGAGACGACGACAGAC-3′ and
5′-GCAAATTCGTGAAGCGTTCCATA-3′), CCND1 (5′-TGAGGGACGCTTTGTCTGTC-3′
and 5′-GCCTTTGGCCTCTCGATACA-3′) and GAPDH
(5′-ATGTTGCAACCGGGAAGGAA-3′ and 5′-AGGAAAAGCATCACCCGGAG-3′).
Western blot analysis
Total proteins were isolated by
radioimmunoprecipitation (RIPA) assay buffer (Wlaterson, Barcelona,
Spain). Protein concentration was determined using a bicinchoninic
acid (BCA) protein assay kit (Wlaterson). A total of 50 µg protein
was separated by 12% sodium dodecyl sulfate polyacrylamide gels
(SDS-PAGE), and transferred to polyvinylidene fluoride (PVDF)
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
blocked with 10% skim milk (w/v) at room temperature for 2 h.
Target proteins were probed with specific antibodies against Cyclin
D1 (1:2,000; cat. no. 2978; Cell Signaling Technology, Inc.,
Danvers, MA, USA) and GAPDH (1:5,000; cat. no. 5174; Cell Signaling
Technology, Inc.), followed by incubation with horseradish
peroxidase conjugated goat-anti-rabbit second antibody (1:5,000;
cat. no. 7074; Cell Signaling Technology Inc.). The blots were
detected with the Enhanced Chemiluminescence western blot detection
kit (Pierce; Thermo Fisher Scientific, Inc.).
Statistical analysis
Each experiment was repeated at least three times.
All data are expressed in terms of means ± SD. The Kaplan-Meier
method was used to calculate the survival curve, and log-rank test
to determine statistical significance. The differences between
groups were analyzed using Two-tail Student's t-test and ANOVA
followed by Tukey's post hoc test. Pearson correlation coefficient
analysis was used to determine the expression correlation between
CCND1 and miR-466. Chi-square test was used for analysis of
correlation between miR-466 expression and clinicopathological
characteristics. P<0.05 was considered to indicate a
statistically significant difference.
Results
MiR-466 is downregulated in OS
tissues
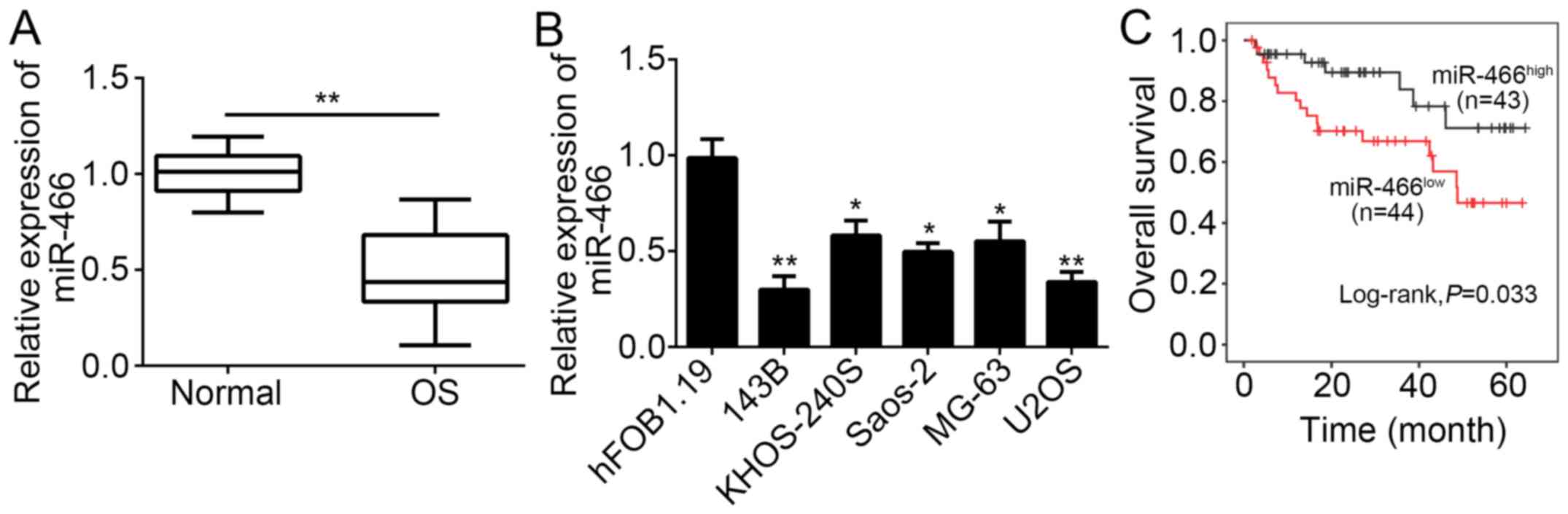
To explore the function of miR-466, we determined
its expression in OS tissues and adjacent normal tissues by
RT-qPCR. The results indicated that miR-466 was significantly
downregulated in OS tissues (n=87) compared with adjacent normal
tissues (n=87; Fig. 1A).
Consistently, miR-466 expression was also downregulated in OS cell
lines, including 143B, U2OS, KHOS-240S, UMR-106, Saos-2, and MG-63
cells compared with hFOB1.19 cells (Fig.
1B). Then, we divided these OS samples into two groups based on
miR-466 expression level and analyzed the correlation between
miR-466 expression and clinicopathological characteristics. We
found that miR-466 expression in OS tissues was negatively
correlated with differentiation, tumor metastasis, and tumor, node,
and metastasis (TNM) stage, whereas no significant correlation was
observed with age and gender (Table
I). Furthermore, we performed Kaplan-Meier curve to determine
the relationship between miR-466 expression and prognosis of
patients with OS. The results showed that high expression of
miR-466 in patients with OS was linked to high survival rate
(Fig. 1C). Overall, our data
indicated that miR-466 was downregulated in OS tissues and
correlated with patients' severity and prognosis, implying that
miR-466 may exert an important role in OS progression.
 | Table I.Correlation between miR-466 expression
and clinicopathological characteristics in OS tissues. |
Table I.
Correlation between miR-466 expression
and clinicopathological characteristics in OS tissues.
| Variables | n=87 | Low (n=44) | High (n=43) | P-value |
|---|
| Age, years |
|
|
| 0.605 |
|
<60 | 68 | 33 | 35 |
|
| ≥60 | 19 | 11 | 8 |
|
| Sex |
|
|
| 0.670 |
| Male | 45 | 24 | 21 |
|
|
Female | 42 | 20 | 22 |
|
| Differentiation |
|
|
| 0.001 |
|
Well/moderate | 45 | 15 | 30 |
|
| Poor | 42 | 29 | 13 |
|
| Metastasis |
|
|
| 0.003 |
|
Absent | 42 | 14 | 28 |
|
|
Present | 45 | 30 | 15 |
|
| TNM stage |
|
|
| 0.018 |
| I–II | 45 | 17 | 28 |
|
|
III–IV | 42 | 27 | 15 |
|
MiR-466 suppresses OS cell
proliferation and induces apoptosis
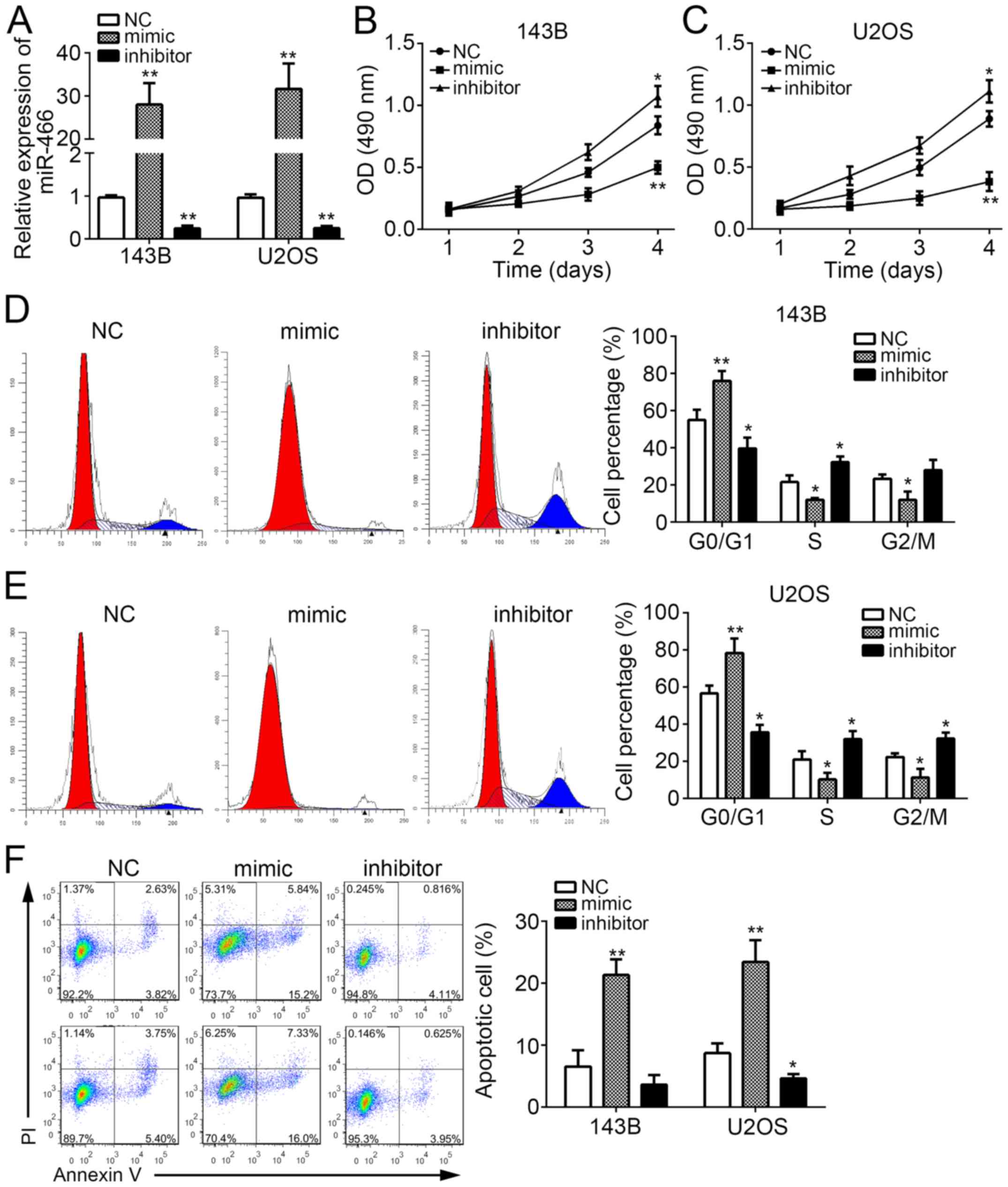
To investigate the physiological functions of
miR-466, we selected 143B and U2OS cells for the following
experiments. We effectively knocked down or overexpressed miR-466
in 143B and U2OS cells by transfection with miR-466 mimic or
inhibitor (Fig. 2A). Then, we
performed CCK8 assays to measure cell proliferation. We found that
miR-466 overexpression significantly inhibited the proliferation of
143B and U2OS cells, and vice versa (Fig. 2B and C). To explore whether the
reduced cell proliferation is induced by aberrant cell cycle, we
stained 143B and U2OS cells with PI and performed FACS analysis.
The results indicated that miR-466 overexpression significantly
increased the percent of cells in G0/G1 phase and decreased cells
in S phase, and vice versa (Fig. 2D and
E). Furthermore, we found that miR-466 overexpression
remarkably promoted the percentages of apoptotic 143B and U2OS
cells (Fig. 2F). Collectively, these
results indicated that miR-466 suppressed OS cell proliferation and
induced apoptosis.
CCND1 is a target of miR-466
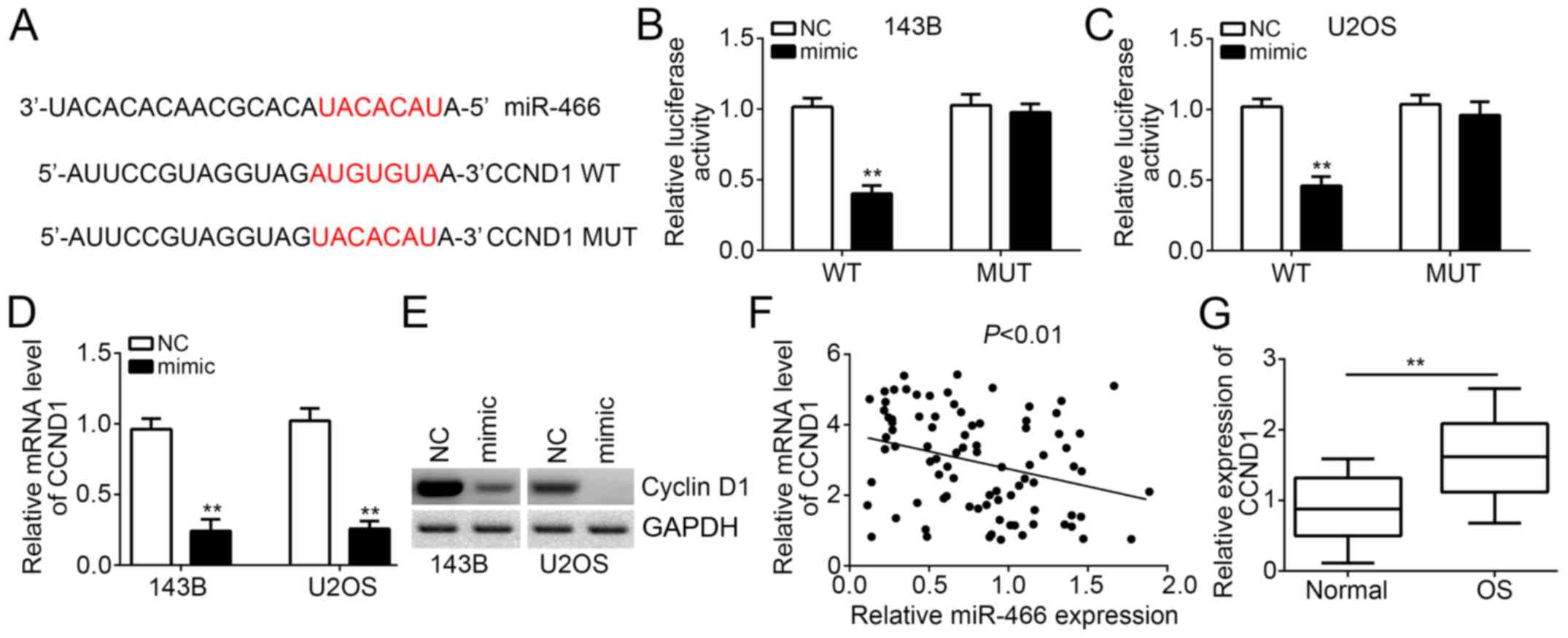
Then, we investigated the molecular mechanism of
miR-466. Through bioinformatics analysis, we found that CCND1
encoding Cyclin D1 was a potential target of miR-466. A potential
binding site of miR-466 in the 3′-UTR region of CCND1 mRNA was
observed (Fig. 3A). Then, we
constructed CCND1-3′-UTR-WT or mutant (MUT) luciferase reporter
plasmid for luciferase reporter assays. The results demonstrated
that miR-466 overexpression significantly inhibited the luciferase
activity in 143B and U2OS cells transfected with CCND1-3′-UTR-WT
(Fig. 3B and C). However, mutation
of the potential binding site in the 3′-UTR region of CCND1 mRNA
abrogated the effect of miR-466 transfection (Fig. 3B and C). Moreover, RT-qPCR and
Western blot results showed that miR-466 overexpression inhibited
the mRNA and protein levels of CCND1 in 143B and U2OS cells
(Fig. 3D and E). We also found that
the miR-466 expression was reversely correlated with that of CCND1
in OS tissues (Fig. 3F). Moreover,
we found that CCND1 was also highly expressed in OS tissues
compared with adjacent normal tissues (Fig. 3G). Overall, above results
demonstrated that CCND1 is a direct target of miR-466 in OS
cells.
Overexpression of CCND1 counteracts
the effects of miR-466 transfection in OS cells
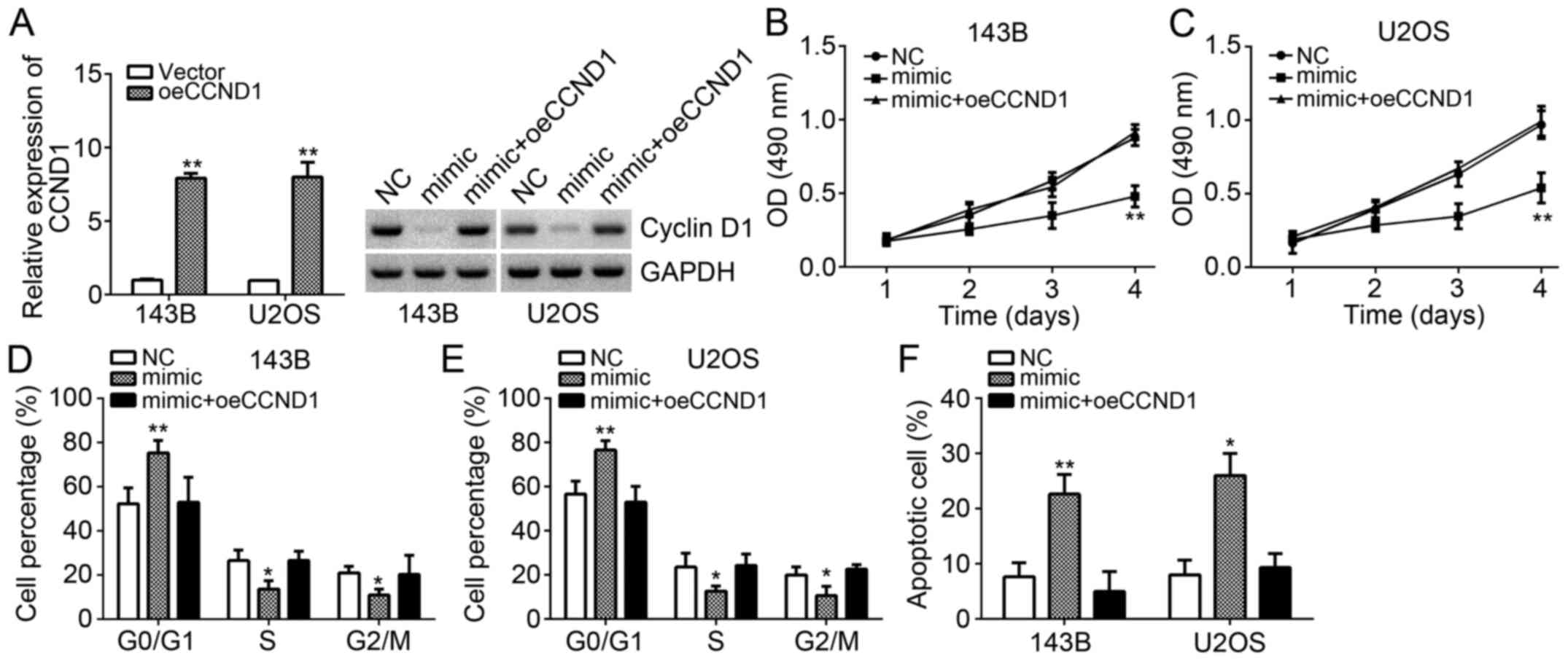
To further validate that CCND1 is important for the
function of miR-466 in OS cells, we performed rescue experiments.
We restored the protein levels of CCND1 in 143B and U2OS cells
transfected with miR-466 mimics. We successfully overexpressed
CCND1 in 143B and U2OS cells (Fig.
4A). And western blot results also indicated that CCND1 was
significantly upregulated in 143B and U2OS cells (Fig. 4A). Obviously, miR-466 overexpression
inhibited the proliferation and cell cycle, whereas CCND1
restoration enhanced cell proliferation and cell cycle in 143B and
U2OS cells (Fig. 4B-E).
Consistently, miR-466 overexpression promoted cell apoptosis,
whereas CCND1 overexpression reduced apoptotic cells (Fig. 4F). These results further confirmed
that miR-466 exerted tumor-suppressive function through targeting
CCND1.
Discussion
OS is the most prevalent and malignant bona fide
tumor in adolescents and children and has become a serious threat
for human life. No effective therapeutic method for OS complete
healing is available to date. The molecular pathogenesis of OS
still remains elusive. MiRNAs are a type of short noncoding RNAs
and have been reported to regulate gene expression
post-transcriptionally. In the past decades, accumulating studies
demonstrate the essential roles of miRNAs in human cancers through
regulating all aspects of bioactivities, such as development,
proliferation, metabolism, and invasion (17,18). In
addition, several pieces of evidence have demonstrated a close
relationship between miRNA and OS progression (19,20).
Therefore, understanding the underlying mechanism of miRNA in OS
development and progression may provide a novel orientation for
diagnosis, prognosis, and therapy of patients with OS. In the
present study, we identified miR-446 as a tumor suppressor in
OS.
Previous reports showed that miR-446 acts as a tumor
suppressor in several cancers, such as prostate cancer and
colorectal cancer (14,15). Besides, Sun et al (21), reported that the expression levels of
miR-466 were higher in cervical cancer tissues than that in
adjacent normal controls. However, whether miR-466 plays a function
in OS needs to be investigated. In this study, we found that
miR-466 was also significantly downregulated in OS tissues and cell
lines compared with adjacent normal control. Furthermore, we found
that miR-466 expression levels were negatively correlated with
tumor TNM stage and metastasis. Additionally, we showed that
miR-466 downregulation predicted a poor prognosis in patients with
OS. Through functional experiments, we found that miR-466
overexpression significantly inhibited the proliferation and
induced cellular apoptosis in OS cells. These data suggested that
miR-466 also served as a tumor suppressor in OS. However, although
we found a correlation between miR-466 expression and metastasis,
whether miR-466 regulates OS cell metastasis requires to be further
investigated by functional experiments.
CCND1 encoding Cyclin D1 is a member of cyclin
family and plays a key role in regulating cell cycle progression
(22). Increasing evidence indicated
that CCND1 exerts an essential role in promoting the development
and progression of human cancers, such as gastric cancer (23), glioma (24), renal cell cancer (25), lung adenocarcinoma (26) and (27). Except regulation of cell cycle,
recent reports also indicated that CCND1 has a pivot role in
promoting migration and tumor metastasis (28). Another study showed that Cyclin D1
recruits HDACs or histone methyltransferases to regulate gene
transcription (29). To explore the
mechanism of CCND1 expression is important because of the extensive
roles of CCND1 in cancer. In our study, we identified CCND1 as a
direct target of miR-446. We showed that miR-446 overexpression
significantly inhibited the CCND1 expression in 143B and U2OS
cells. Moreover, a reverse correlation was observed between their
expression levels in OS tissues. Through functional experiments, we
demonstrated that CCND1 restoration markedly rescued the effects of
miR-446 overexpression on OS cell proliferation and apoptosis.
In conclusion, our results showed that miR-446 is
significantly downregulated and inversely correlated with CCND1
expression in OS tissues. Besides, these data demonstrated that
miR-446 serves as a tumor suppressor and might be a promising
biomarker for prognosis of patients with OS. This study also
implied that miR-446 might serve as a potential therapeutic target
for OS treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
WC and TL initiated and designed the present study,
analyzed and interpreted the results, and wrote this manuscript.
LF, ST and HC performed certain experiments. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
For the use of human samples, the protocol for this
study was approved by the Institutional Ethics Committee of
Hospital of Capital Medical University and all enrolled patients
signed a written informed consent document.
Patient consent for publication
All patients within the present study provide
consent for the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Heare T, Hensley MA and Dell'Orfano S:
Bone tumors: Osteosarcoma and Ewing's sarcoma. Curr Opin Pediatr.
21:365–372. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shackleton M, Quintana E, Fearon ER and
Morrison SJ: Heterogeneity in cancer: Cancer stem cells versus
clonal evolution. Cell. 138:822–829. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Levings PP, McGarry SV, Currie TP,
Nickerson DM, McClellan S, Ghivizzani SC, Steindler DA and Gibbs
CP: Expression of an exogenous human Oct-4 promoter identifies
tumor-initiating cells in osteosarcoma. Cancer Res. 69:5648–5655.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fagioli F, Aglietta M, Tienghi A, Ferrari
S, Brach del Prever A, Vassallo E, Palmero A, Biasin E, Bacci G,
Picci P and Madon E: High-dose chemotherapy in the treatment of
relapsed osteosarcoma: An Italian sarcoma group study. J Clin
Oncol. 20:2150–2156. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang Y, Jiang Z, Ma N, Wang B, Liu J,
Zhang L and Gu L: MicroRNA-223 targeting STIM1 inhibits the
biological behavior of breast cancer. Cell Physiol Biochem.
45:856–866. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zavala-Yoe R, Ramirez-Mendoza RA and
Cordero LM: Entropy measures to study and model long term
simultaneous evolution of children in Doose and Lennox-Gastaut
syndromes. J Integr Neurosci. 15:205–221. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cui Y, Chen LG, Yao HB, Zhang J and Ding
KF: Upregulation of microRNA-383 inhibits the proliferation,
migration and invasion of colon cancer cells. Oncol Lett.
15:1184–1190. 2018.PubMed/NCBI
|
|
9
|
Du P, Luan X, Liao Y, Mu Y, Yuan Y, Xu J
and Zhang J: MicroRNA-509-3p inhibits cell proliferation and
invasion via downregulation of X-linked inhibitor of apoptosis in
glioma. Oncol Lett. 15:1307–1312. 2018.PubMed/NCBI
|
|
10
|
Wang P, Zhang L, Zhang J and Xu G:
MicroRNA-124-3p inhibits cell growth and metastasis in cervical
cancer by targeting IGF2BP1. Exp Ther Med. 15:1385–1393.
2018.PubMed/NCBI
|
|
11
|
Wang H, Xing D, Ren D, Feng W, Chen Y,
Zhao Z, Xiao Z and Peng Z: MicroRNA-643 regulates the expression of
ZEB1 and inhibits tumorigenesis in osteosarcoma. Mol Med Rep.
16:5157–5164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Z, Li Z, Shen Y and Chen T:
MicroRNA-138 directly targets TNFAIP8 and acts as a tumor
suppressor in osteosarcoma. Exp Ther Med. 14:3665–3673. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin T, Ma Q, Zhang Y, Zhang H, Yan J and
Gao C: MicroRNA-27a functions as an oncogene in human osteosarcoma
by targeting CCNG1. Oncol Lett. 15:1067–1071. 2018.PubMed/NCBI
|
|
14
|
Colden M, Dar AA, Saini S, Dahiya PV,
Shahryari V, Yamamura S, Tanaka Y, Stein G, Dahiya R and Majid S:
MicroRNA-466 inhibits tumor growth and bone metastasis in prostate
cancer by direct regulation of osteogenic transcription factor
RUNX2. Cell Death Dis. 8:e25722017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tong F, Ying Y, Pan H, Zhao W, Li H and
Zhan X: MicroRNA-466 (miR-466) functions as a tumor suppressor and
prognostic factor in colorectal cancer (CRC). Bosn J Basic Med Sci.
Jan 17–2018.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang Y, Shen XJ, Zou Q, Wang SP, Tang SM
and Zhang GZ: Biological functions of microRNAs: A review. J
Physiol Biochem. 67:129–139. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang ZM, Wan XH, Sang GY, Zhao JD, Zhu QY
and Wang DM: miR-15a-5p suppresses endometrial cancer cell growth
via Wnt/β-catenin signaling pathway by inhibiting WNT3A. Eur Rev
Med Pharmacol Sci. 21:4810–4818. 2017.PubMed/NCBI
|
|
19
|
Nugent M: microRNA and bone cancer. Adv
Exp Med Biol. 889:201–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao X, Han D and Fan W: Down-regulation of
RBP-J mediated by microRNA-133a suppresses dendritic cells and
functions as a potential tumor suppressor in osteosarcoma. Exp Cell
Res. 349:264–272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun P, Shen Y, Gong JM, Zhou LL, Sheng JH
and Duan FJ: A new MicroRNA expression signature for cervical
cancer. Int J Gynecol Cancer. 27:339–343. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao M, Xu P, Liu Z, Zhen Y, Chen Y, Liu
Y, Fu Q, Deng X, Liang Z, Li Y, et al: Dual roles of miR-374a by
modulated c-Jun respectively targets CCND1-inducing PI3K/AKT signal
and PTEN-suppressing Wnt/β-catenin signaling in non-small-cell lung
cancer. Cell Death Dis. 9:782018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang H, Han Y, Yang X, Li M, Zhu R, Hu J,
Zhang X, Wei R, Li K and Gao R: HNRNPK inhibits gastric cancer cell
proliferation through p53/p21/CCND1 pathway. Oncotarget.
8:103364–103374. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen DG, Zhu B, Lv SQ, Zhu H, Tang J,
Huang C, Li Q, Zhou P, Wang DL and Li GH: Inhibition of EGR1
inhibits glioma proliferation by targeting CCND1 promoter. J Exp
Clin Cancer Res. 36:1862017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xue J, Qin Z, Li X, Zhang J, Zheng Y, Xu
W, Cao Q and Wang Z: Genetic polymorphisms in cyclin D1 are
associated with risk of renal cell cancer in the Chinese
population. Oncotarget. 8:80889–80899. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yao Y, Luo J, Sun Q, Xu T, Sun S, Chen M,
Lin X, Qian Q, Zhang Y, Cao L, et al: HOXC13 promotes proliferation
of lung adenocarcinoma via modulation of CCND1 and CCNE1. Am J
Cancer Res. 7:1820–1834. 2017.PubMed/NCBI
|
|
27
|
Wu J, Cui LL, Yuan J, Wang Y and Song S:
Clinical significance of the phosphorylation of MAPK and protein
expression of cyclin D1 in human osteosarcoma tissues. Mol Med Rep.
15:2303–2307. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Z, Wang C, Prendergast GC and Pestell
RG: Cyclin D1 functions in cell migration. Cell Cycle. 5:2440–2442.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fu M, Rao M, Bouras T, Wang C, Wu K, Zhang
X, Li Z, Yao TP and Pestell RG: Cyclin D1 inhibits peroxisome
proliferator-activated receptor gamma-mediated adipogenesis through
histone deacetylase recruitment. J Biol Chem. 280:16934–16941.
2005. View Article : Google Scholar : PubMed/NCBI
|