Introduction
Cervical cancer is one of the most common cancer
among females in the world. It is estimated that approximately over
500,000 new cases are reported each year (1). Previous studies have indicated that
human papilloma virus (HPV) infection is the major cause of
cervical cancer (2,3). Undoubtedly, long term exposure to
oxidative stress by cervical epithelial tissues will lead to the
persistent, chronic viral infections, thereby causing genetic
rearrangements and genomic instability due to the integration of
the viral genome (4,5).
In the normal physiological status, an antioxidant
system in cervical epithelial tissue can maintain the homeostasis
through nuclear factor erythroid 2-related factor 2 (NRF2)
(6,7). NRF2 mainly binds to the antioxidant
response element (ARE) in target gene promoters thereby maintaining
redox balance (8). Increasing
evidence has shown that upregulation of NRF2 tightly correlated
with multiple tumor development and progression (9,10). It is
showed that NRF2 protects normal cells from transformation and
enhances cancer cell proliferation and survival (11). Therefore, it is important to maintain
NRF2 at a normal status for the prevention and treatment of
cervical cancer patients.
Allicin (diallyl thiosulfinate) is the major
component present in freshly crushed garlic, and it is also a key
active compounds of garlic (12,13). In
the past years, allicin has been extensively applied in the clinic
based on the characteristics of anti-inflammatory, anti-microbial,
cardiovascular protection, and immunity functions (14,15).
Emerging evidence has demonstrated the antitumor activity of
allicin in gastric carcinoma, breast cancer, and glioblastoma
mainly by inhibiting cell proliferation and inducing cell apoptosis
(16,17). However, the functional role of
allicin in cervical cancer cells and the potential molecular
mechanism is still unknown.
In the present study, we found that allicin
significantly suppressed cervical cancer proliferation and
migration mainly by inhibiting the expression of NRF2, thereby
maintaining the intracellular oxidative homeostasis.
Materials and methods
Cell culture and transfections
SiHa cells (ATCC; Manassas, VA, USA), a human
cervical squamous cell carcinoma cell line, were cultured in
RPMI-1640 plus 10% calf serum and 1% penicillin/streptomycin in a
5% CO2 humidified incubator at 37°C.
Transient transfection
NRF2 in the eukaryotic expression vector pcDNA3.1,
and NRF2 siRNA, and the scrambled sequence
(CCAACCAGUUGACAGUGAACUCAUU/CAAACUGACAGAAGUUGACAAUUAU) were
constructed by GenePharma (Shanghai, China).
In brief, SiHa cells were seeded in 6-well plates
and grown to 60–80% confluence overnight. Transfection complexes
were formed with Lipofectamine RNAiMAX (Invitrogen, CA, USA) in
Opti-MEMI (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) according to manufacturer guidelines. All transfections were
performed in triplicate. Cell proliferation was determined by
counting cells 24, 48, and 72 h after transfection.
RNA isolation and qRT-PCR
Total RNA was isolated from SiHa cells using Trizol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. RNA was reverse transcribed into
cDNA using the Prime-Script one-step RT-PCR kit (C28025-032,
Invitrogen; Thermo Fisher Scientific, Inc.). Detailed RT-PCR
procedure was described as follows: 95°C for 10 min followed by 50
cycles of 95°C for 10 sec, 55°C for 10 sec, 72°C for 5 sec; 99°C
for 1 sec; 59°C for 15 sec; 95°C for 1 sec; then cooling to 40°C.
The relative expression levels were calculated with the
2−∆∆Cq method and experiments were repeated in
triplicate. The primers used were listed as follows: NRF2-forward
primer: 5′-TCAGCGACGGAAAGAGTATGA-3′ and reverse primer:
5′-CCACTGGTTTCTGACTGGATGT-3′.
Protein isolation and western
blotting
Firstly, protease inhibitors (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) were added to cell lysates and maintained
on ice for 15 min. Then, cell lysates were centrifuged at 12,000 ×
g for 10 min at 4°C. And the supernatant was collected and were
boiled for 5 min in sample buffer. All the samples were separated
on 12% gels by SDS-PAGE and transferred to polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA). Then, the membranes
was blocked in 10% skim milk for 40 min at 37°C. A primary antibody
against NRF2 (Abcam, Cambridge, MA, USA) or β-Actin (Sangon,
Shanghai, China) was added overnight to blots at 4°C. Blots were
washed in PBS Tween three times, after which the secondary antibody
(horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin
G; Zhongshan Gold Bridge, Beijing, China) was added at room
temperature for 2 h. Chemiluminescent substrate (Thermo Fisher
Scientific, Inc.) was added to visualize bands. Quantity One
software was used to quantify the intensity of each band and was
normalized to the intensity of the internal control β-actin.
Detection of reactive oxygen
species
Cells were seeded at 1×104 cells per well
into 96-well plates and treated with 40 nM allicin. The cells were
incubated for 48 h at 37°C and 5% CO2. At the end of the
incubation period, cells were washed twice in PBS and incubated in
200 µM ROS Fluorescent Probe-DHE (Vigorous, Beijing, China) for 15
min (Sigma-Aldrich; Merck KGaA).
Analyses of cell cycle and apoptotic
changes by flow cytometry
SiHa cells were seeded in 6-well culture plates at a
density of 5×104 cells/well in RPMI 1640 supplemented
with 10% calf serum and 1% penicillin/streptomycin. After allicin
treatment for 48 h, cell cycle distributions were examined by
measuring PI-fluorescence with a BD FACS Calibur flow cytometer (BD
Biosciences, San Jose, CA, USA) through an FL-2 filter (585
nm).
Annexin V staining was performed to evaluate
apoptosis. Control and treated SiHa cells were added at
5×105 cells/ml in binding buffer (10 mM HEPES
[(4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid] [Ph 7.4], 140
mM NaCl, 2.5 mM CaCl2). FITC-Annexin V (10 µl) in 190 µl of cell
suspension was incubated for 10 min at room temperature. Cell
mixtures were centrifuged and resuspended in 190 µl binding buffer,
and 10 µl PI (1 mg/ml) solution was added. Then, the cells were
washed with cold PBS and resuspended at a final concentration of
1×106 cells/ml. FITC-Annexin V (5 µl) and propidium
iodide were gently mixed and incubated with the cells for 15 min at
a room temperature. After incubation, the samples were analyzed by
flow cytometry within 1 h. The Annexin V− and
PI+ represented necrotic cells, the Annexin
V+ and PI+ represented late apoptotic cell,
the Annexin V+ and PI− represented early
apoptotic cell, and the Annexin V− and PI−
represented normal cells.
Transwell migration and invasion
assays
Migration and invasion assays were performed as
previously described. Migration was evaluated in Transwell cell
culture chambers with 6.5-mm-diameter polycarbonate membrane
filters containing 8-µm pores (Corning Incorporated, Corning, NY,
USA). Cells were added in 100 ml serum-free media to the upper
chamber. The lower chamber contained 600 ml culture media with 10%
calf serum. After 10 h at 37°C, cells were removed from the upper
surface of the membrane with a cotton swab. Filters were fixed in
methanol for 20 min and stained with Giemsa solution for 30 min.
The number of cells that had migrated were counted. Five random
fields (Nikon ECLIPSE TS100; Nikon Corporation, Tokyo, Japan) were
counted per well, and the mean was calculated. The membrane of the
upper chamber of the transwell was pre-coated with 100 ml of a 1
mg/ml solution of Matrigel (BD Biosciences, Franklin Lakes, NJ,
USA).
TUNEL assay
Cells were stained by terminal deoxy-nucleotidyl
transferase-mediated dUTP nick-end labeling (TUNEL; In situ
Cell Death Detection kit; Roche Diagnostics, Basel, Switzerland).
In brief, cells were fixed in 4% paraformaldehyde, added
permeabilisation solution, and incubated with TUNEL reaction
mixture. TUNEL positive cells and total cells were counted and
percent apoptotic cells calculated.
Statistical analysis
The data are represented as the mean ± standard
error of the mean (SEM). The two-tailed unpaired Student's t-tests
were used for comparisons of two groups. The ANOVA multiple
comparison test (SPSS 13.0; SPSS, Inc., Chicago, IL, USA) followed
by Turkey post hoc test were used for comparisons of two more
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
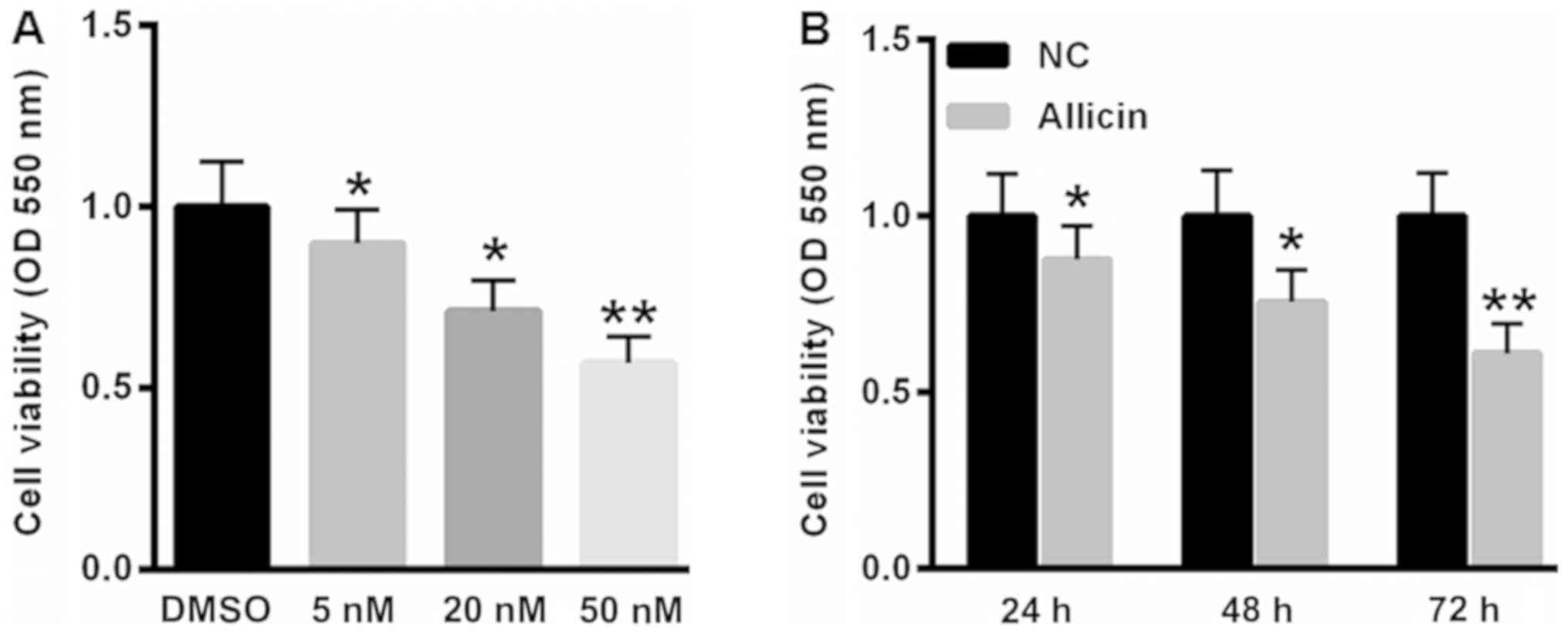
Allicin suppressed SiHa cell viability
in time- and dose-dependent manner
Firstly, we analyzed the effects of allicin on SiHa
cell viability. As shown in Fig. 1A,
treatment with 5, 20 and 50 nM allicin significantly decreased cell
viability by 10.2, 27.8 and 43.1%, respectively. Furthermore,
incubation of 20 nM allicin reduced SiHa cell viability by 12.3,
24.3 and 38.9 at 24, 48 and 72 h (Fig.
1B).
Allicin-induced accumulation of
reactive oxygen species and SiHa cell apoptosis
Next, we evaluated the role of allicin in the
accumulation of ROS. Compared with blank control,
H2O2 treatment markedly increased the
fluorescence density of DHE, but allicin treatment could reverse
H2O2-induced ROS accumulation (Fig. 2A). We also quantified the contents of
glutathione (GSH), superoxide dismutase (SOD) and malondialdehyde
(MDA) contents. Our data showed that H2O2
treatment decreased GSH and SOD contents, but increased MDA content
(Fig. 2B-D). In contrast, GSH and
SOD contents were increased, but MDA was decreased after allicin
incubation for 48 h (Fig. 2B-D).
Futhermore, TUNEL staining showed H2O2
treatment induced cell apoptosis, but allicin treatment could
decrease cell apoptosis (Fig.
2E).
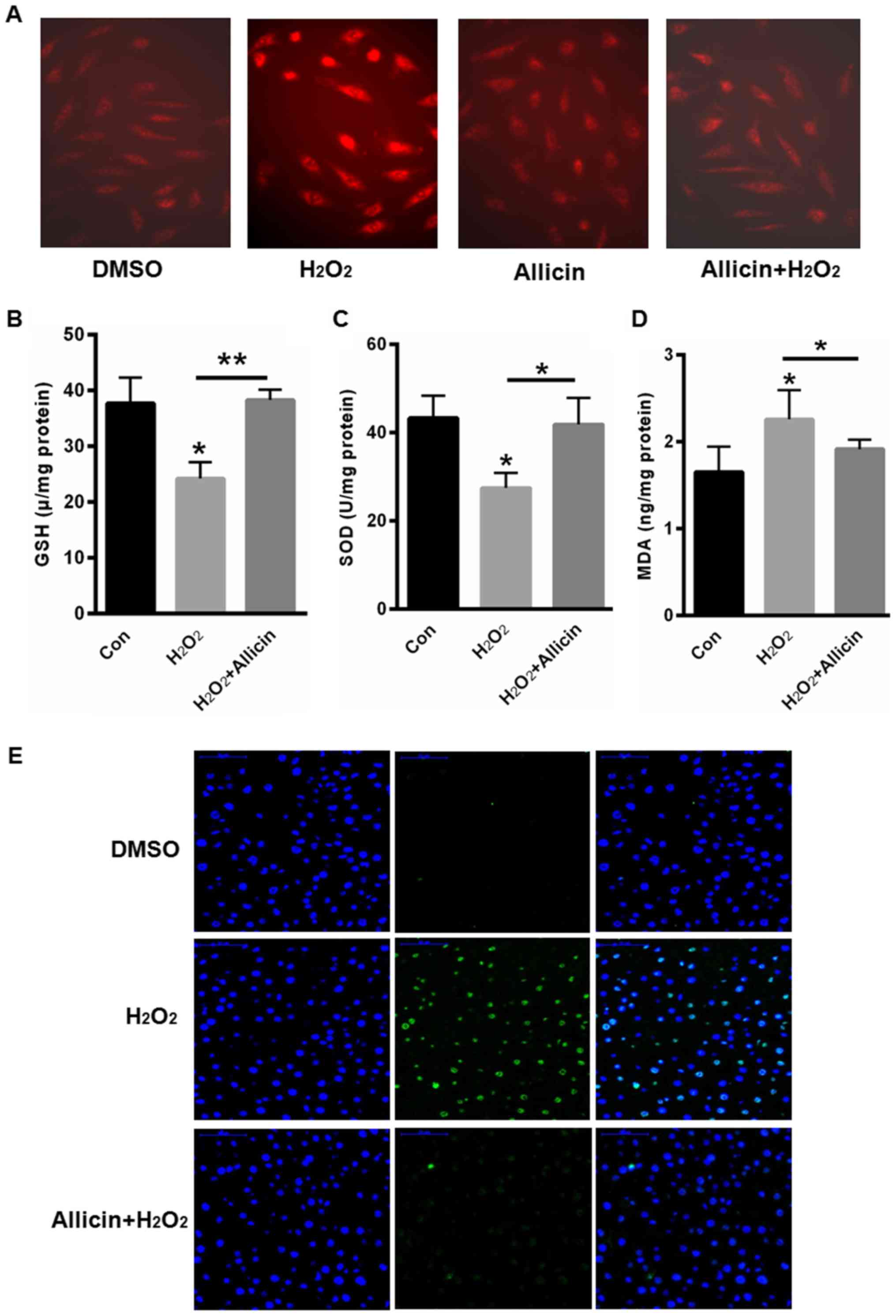 | Figure 2.Allicin-induced accumulation of
reactive oxygen species and SiHa Cell apoptosis. (A) DHE staining.
Quantification of (B) GSH, (C) SOD and (D) MDA contents. (E) TUNEL
staining. *P<0.05, **P<0.01, vs. control. DHE,
dihydroethidium; GSH, glutathione; SOD, superoxide dismutase; MDA,
malondialdehyde; TUNEL, terminal deoxy-nucleotidyl
transferase-mediated dUTP nick-end labeling; DMSO, dimethyl
sulfoxide; con, control. |
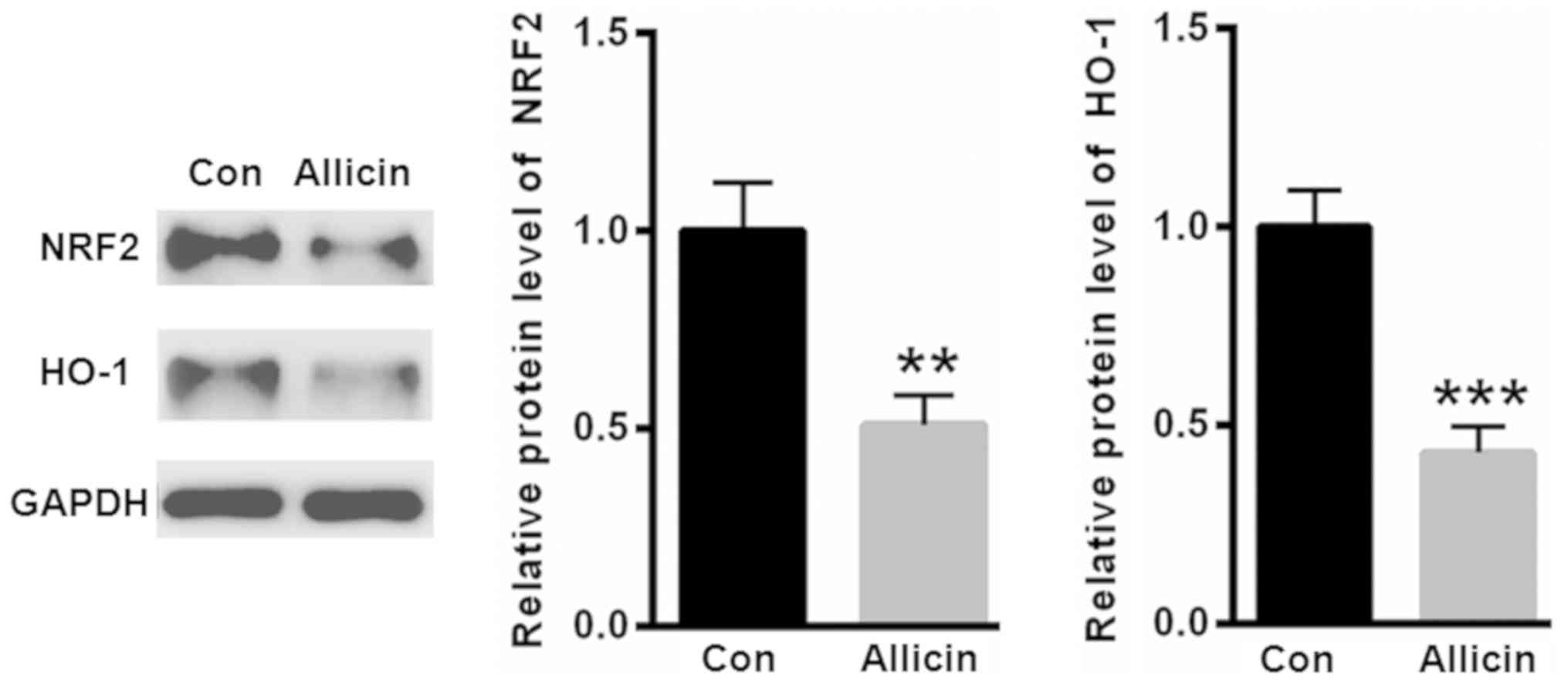
Allicin inhibited the expression of
NRF2 in SiHa cells
NRF2 is a key transcription factor that is widely
involved in the regulation of antioxidant genes. Thus, we evaluated
the expression of NRF2 after allicin treatment. As shown in
Fig. 3, treatment with allicin
significantly suppressed the level of NRF2. Furthermore, heme
oxygenase 1 (HO-1), an antioxidant enzyme regulated by NRF2, was
decreased by allicin incubation (Fig.
3).
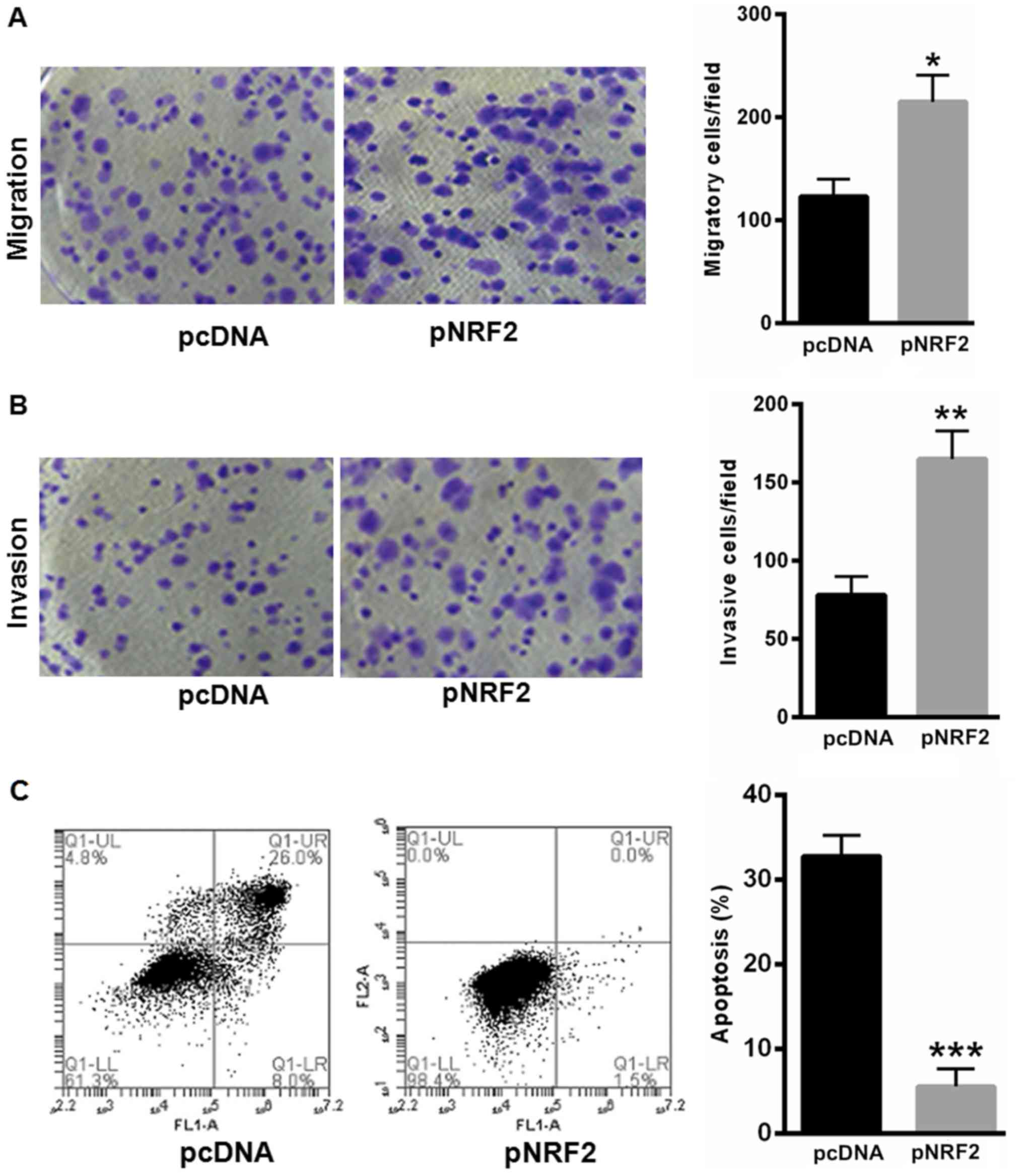
NRF2 prompted SiHa cell proliferation
and reduced SiHa cell apoptosis
Furthermore, we explored the effects of NRF2 in SiHa
cell proliferation and apoptosis. Our data showed that
overexpressing NRF2 significantly enhanced SiHa cell migration and
invasion capacity (Fig. 4A and B).
Furthermore, overexpressing NRF2 could largely reverse
H2O2-induced cell apoptosis (Fig. 4C).
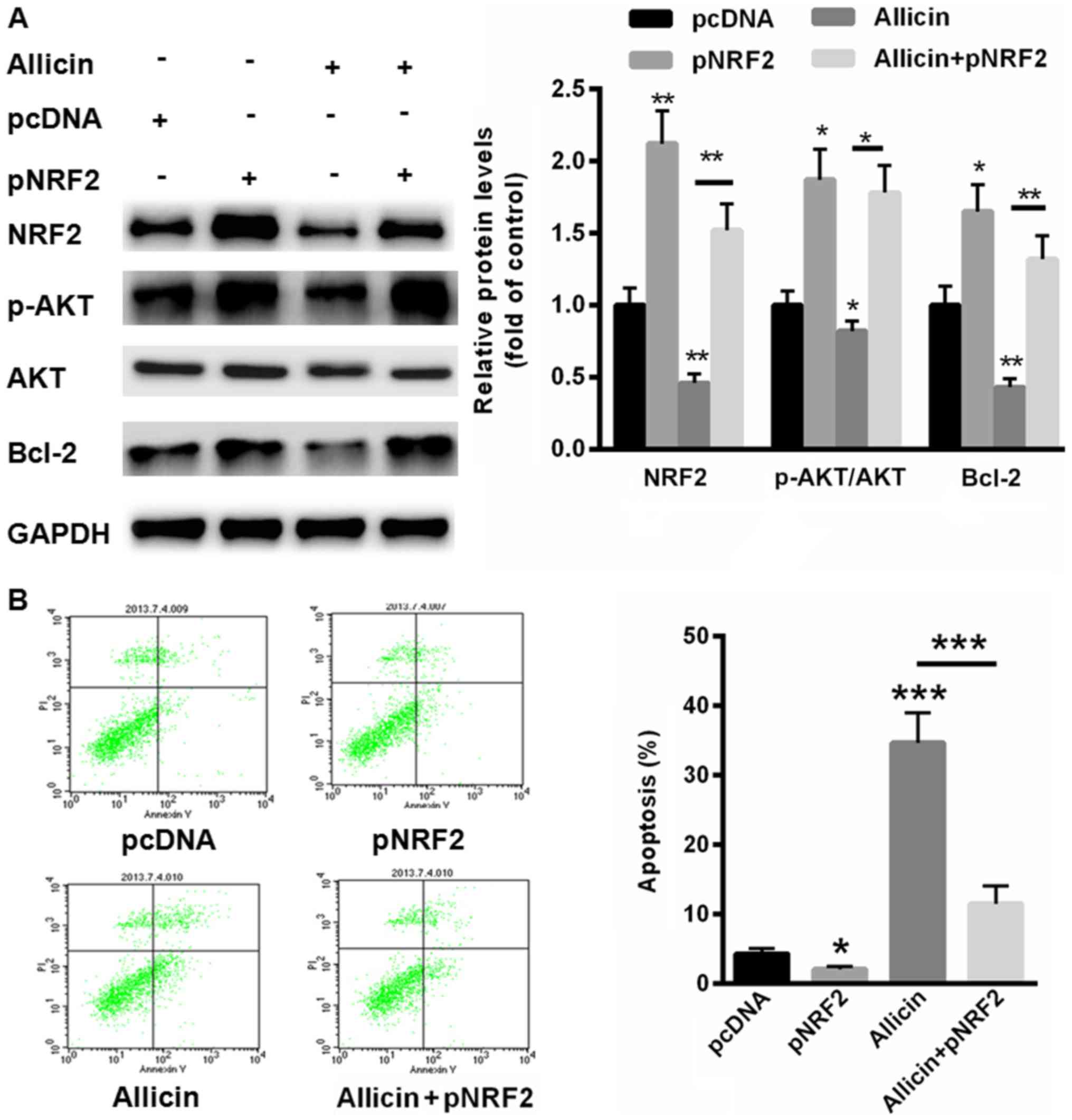
Allicin suppressed the malignant
phenotype of SiHa cells by inhibiting NRF2
We further evaluated whether allicin inhibits SiHa
cell proliferation through suppressing NRF2. Thus, a full-length
human NRF2 was successfully transfected into SiHa cells in the
presence or absence of allicin. Compared with normal control,
transfection of NRF2 significantly enhanced the activation of
PI3K/AKT signaling (Fig. 5A).
However, pre-incubation of allicin could decrease the
phosphorylation levels of PI3K/AKT and the protein level of Bcl-2
(Fig. 5A). In contrast,
allicin-inactivated PI3K/AKT signaling and Bcl-2 expression could
partially reversed by overexpressing of NRF2 (Fig. 5A). We also evaluated cell apoptosis
in SiHa cells transfected with pNRF2. Our data showed that
overexpressing of NRF2 decreased cell apoptosis rate by 3.1±0.54%
(Fig. 5B). More importantly,
allicin-induced cell apoptosis (43.5±3.8%) could largely be
abolished by upregulation of NRF2 (12.3±2.08%). These data
indicated that allicin induced SiHa cell apoptosis mainly by
suppressing the expression of NRF2.
Discussion
Cervical cancer is ranked as the second most common
cancer in women worldwide (18,19). A
most important feature of cervical cancer is the high mortality
rate, which is mainly attributed to the lack of effective therapies
for women with high-grade cervical cancer (20). It is reported that high-risk human
papillomaviruses (HPVs) is a key causal factor for cervical cancer
(21). And HPV infection is also
suggested to correlate with other anogenital cancers as well as a
small fraction of head & neck cancer (22). Therefore, it is important to identify
effective prevention methods of human cervical cancers.
Allicin is characterized by antitumor effect in
multiple cancers through suppressing cancer cell growth and
increasing cell apoptosis (23,24). For
example, allicin is reported to sensitize hepatocellular carcinoma
(HCC) cells to 5-FU induced apoptosis mainly by modulating
ROS-mediated mitochondrial pathway, showing the application of
allicin as a novel chemotherapy regimen in HCC (25). In addition, allicin is reported to
enhance MGC-803 human gastric carcinoma cell apoptosis by
activating the p38 mitogen-activated protein kinase/caspase-3
signaling pathway (26). In the
current study, we mainly evaluated the role of allicin in the
malignant proliferation of cervical cancer cells. Our data showed
that allicin suppressed cervical cancer viability in a time- and
dose-dependent manner. Further study revealed that allicin
inhibited cervical cancer cell proliferation and migration. These
data showed an antitumor role of allicin in cervical cancer
cells.
These above findings prompts us to further explore
the underlying mechanism in which allicin modulates the progression
of cervical cancer. Here, we mainly focused on NRF2, an
anti-oxidant enzyme. It is reported that abnormal activation of
NRF2 enhances the expression of enzymes for the detoxification of
chemical carcinogens, thereby leading to the protection against
carcinogenicity, mutagenicity and various toxicity (27,28).
Increasing evidence has showed the protective role of NRF2 in
intracellular oxidative stress, chemotherapeutic agents and
radiotherapy (29,30). However, disruption of NRF2 also
enables the cells towards carcinogens, which resulting in the
development of inflammation and cancer formation (31,32).
Therefore, it is important to maintain the expression of NRF2 in
normal status, or else the excessive NRF2 expression confers to the
abberant survival of cancer cells.
Here, we found that treatment with allicin could
significantly suppress the expression of NRF2 and the downstream
enzyme, HO-1. Meanwhile, we also evaluated the functional role of
NRF2 on cervical cancer cell proliferation. We found that
overexpressing of NRF2 enhanced cervical cancer cell invasion and
migration, indicating an oncogenic role of NRF2 in cervical cancer
cells. More importantly, allicin-induced cell apoptosis could
largely be abolished by overexpressing of NRF2, indicating the
antitumor role of allicin in cervical cancer cells mainly by
suppressing NRF2.
In summary, our data showed allicin was effective to
suppress the malignant phenotype of cervical cancer cells mainly by
inhibiting the expression of NRF2, showing the potential clinical
benefits of allicin in cervical cancer patients.
References
|
1
|
Ma JQ, Tuersun H, Jiao SJ, Zheng JH, Xiao
JB and Hasim A: Functional role of NRF2 in cervical carcinogenesis.
PLoS One. 10:e01338762015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Feng Y, Wang Y, Jiang C, Fang Z, Zhang Z,
Lin X, Sun L and Jiang W: Nicotinamide induces
mitochondrial-mediated apoptosis through oxidative stress in human
cervical cancer HeLa cells. Life Sci. 181:62–69. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alvarez-Olmedo DG, Biaggio VS, Koumbadinga
GA, Gómez NN, Shi C, Ciocca DR, Batulan Z, Fanelli MA and O'Brien
ER: Recombinant heat shock protein 27 (HSP27/HSPB1) protects
against cadmium-induced oxidative stress and toxicity in human
cervical cancer cells. Cell Stress Chaperones. 22:357–369. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen L, Wang L, Shen H, Lin H and Li D:
Anthelminthic drug niclosamide sensitizes the responsiveness of
cervical cancer cells to paclitaxel via oxidative stress-mediated
mTOR inhibition. Biochem Biophys Res Commun. 484:416–421. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Souza RP, Bonfim-Mendonca PS, Gimenes F,
Ratti BA, Kaplum V, Bruschi ML, Nakamura CV, Silva SO, Maria-Engler
SS and Consolaro ME: Oxidative stress triggered by apigenin induces
apoptosis in a comprehensive panel of human cervical cancer-derived
cell lines. Oxid Med Cell Longev. 2017:15127452017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shah P, Trinh E, Qiang L, Xie L, Hu WY,
Prins GS, Pi J and He YY: Arsenic induces p62 expression to form a
positive feedback loop with nrf2 in human epidermal keratinocytes:
Implications for preventing arsenic-induced skin cancer. Molecules.
22:E1942017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bao L, Wu J, Dodson M, Rojo de la Vega EM,
Ning Y, Zhang Z, Yao M, Zhang DD, Xu C and Yi X: ABCF2, an Nrf2
target gene, contributes to cisplatin resistance in ovarian cancer
cells. Mol Carcinog. 56:1543–1553. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cho HY, Kim K, Kim YB, Kim H and No JH:
Expression patterns of Nrf2 and keap1 in ovarian cancer cells and
their prognostic role in disease recurrence and patient survival.
Int J Gynecol Cancer. 27:412–419. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duong HQ, You KS, Oh S, Kwak SJ and Seong
YS: Silencing of NRF2 reduces the expression of aldh1a1 and aldh3a1
and sensitizes to 5-fu in pancreatic cancer cells. Antioxidants
(Basel). 6:E522017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fabrizio FP, Costantini M, Copetti M, la
Torre A, Sparaneo A, Fontana A, Poeta L, Gallucci M, Sentinelli S,
Graziano P, et al: Keap1/Nrf2 pathway in kidney cancer: Frequent
methylation of KEAP1 gene promoter in clear renal cell carcinoma.
Oncotarget. 8:11187–11198. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gonzalez-Donquiles C, Alonso-Molero J,
Fernandez-Villa T, Vilorio-Marqués L, Molina AJ and Martín V: The
NRF2 transcription factor plays a dual role in colorectal cancer: A
systematic review. PLoS One. 12:e01775492017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang H, Zheng F, Dong X, Wu F, Wu T and
Li H: Allicin inhibits tubular epithelial-myofibroblast
transdifferentiation under high glucose conditions in vitro. Exp
Ther Med. 13:254–262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gruhlke MC, Nicco C, Batteux F and
Slusarenko AJ: The effects of allicin, a reactive sulfur species
from garlic, on a selection of mammalian cell lines. Antioxidants
(Basel). 6:E12016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen X, Pang S, Lin J, Xia J and Wang Y:
Allicin prevents oxidized low-density lipoprotein-induced
endothelial cell injury by inhibiting apoptosis and oxidative
stress pathway. BMC Complement Altern Med. 16:1332016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding G, Zhao J and Jiang D: Allicin
inhibits oxidative stress-induced mitochondrial dysfunction and
apoptosis by promoting PI3K/AKT and CREB/ERK signaling in
osteoblast cells. Exp Ther Med. 11:2553–2560. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li S, Chen S, Yang W, Liao L, Li S, Li J,
Zheng Y and Zhu D: Allicin relaxes isolated mesenteric arteries
through activation of PKA-KATP channel in rat. J Recept Signal
Transduct Res. 37:17–24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang D, Lv Z, Zhang H, Liu B, Jiang H, Tan
X, Lu J, Baiyun R and Zhang Z: Activation of the Nrf2 signaling
pathway involving KLF9 plays a critical role in allicin resisting
against arsenic trioxide-induced hepatotoxicity in rats. Biol Trace
Elem Res. 176:192–200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mahmoodi P, Motamedi H, Seyfi Abad
Shapouri MR, Bahrami Shehni M and Kargar M: Molecular detection and
typing of human papillomaviruses in paraffin-embedded cervical
cancer and pre-cancer tissue specimens. Iran J Cancer Prev.
9:e37522016.PubMed/NCBI
|
|
19
|
Kim M, Kim YS, Kim H, Kang MY, Park J, Lee
DH, Roh GS, Kim HJ, Kang SS, Cho GJ, et al: O-linked
N-acetylglucosamine transferase promotes cervical cancer
tumorigenesis through human papillomaviruses E6 and E7 oncogenes.
Oncotarget. 7:44596–44607. 2016.PubMed/NCBI
|
|
20
|
Piroozmand A, Mostafavi Zadeh SM, Madani
A, Soleimani R, Nedaeinia R, Niakan M, Avan A, Manian M, Moradi M
and Eftekhar Z: The association of high risk human papillomaviruses
in patients with cervical cancer: an evidence based study on
patients with squamous cell dysplasia or carcinoma for evaluation
of 23 human papilloma virus genotypes. Jundishapur J Microbiol.
9:e327282016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gu Y, Ma C, Zou J, Zhu Y, Yang R, Xu Y and
Zhang Y: Prevalence characteristics of high-risk human
papillomaviruses in women living in Shanghai with cervical
precancerous lesions and cancer. Oncotarget. 7:24656–24663. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zandnia F, Doosti A, Mokhtari-Farsani A,
Kardi MT and Movafagh A: Application of multiplex PCR for rapid and
sensitive detection of human papillomaviruses in cervical cancer.
Pak J Med Sci. 32:444–447. 2016.PubMed/NCBI
|
|
23
|
Müller A, Eller J, Albrecht F, Prochnow P,
Kuhlmann K, Bandow JE, Slusarenko AJ and Leichert LI: Allicin
induces thiol stress in bacteria through s-allylmercapto
modification of protein cysteines. J Biol Chem. 291:11477–11490.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tu G, Zhang YF, Wei W, Li L, Zhang Y, Yang
J and Xing Y: Allicin attenuates H2O2-induced
cytotoxicity in retinal pigmented epithelial cells by regulating
the levels of reactive oxygen species. Mol Med Rep. 13:2320–2326.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zou X, Liang J, Sun J, Hu X, Lei L, Wu D
and Liu L: Allicin sensitizes hepatocellular cancer cells to
anti-tumor activity of 5-fluorouracil through ROS-mediated
mitochondrial pathway. J Pharmacol Sci. 131:233–240. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Zhu Y, Duan W, Feng C and He X:
Allicin induces apoptosis of the MGC-803 human gastric carcinoma
cell line through the p38 mitogen-activated protein
kinase/caspase-3 signaling pathway. Mol Med Rep. 11:2755–2760.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang XY, Zhu XS, Xu HY, Zhao ZX, Li SY,
Li SZ, Cai JH and Cao JM: Diallyl trisulfide suppresses tumor
growth through the attenuation of Nrf2/Akt and activation of
p38/JNK and potentiates cisplatin efficacy in gastric cancer
treatment. Acta Pharmacol Sin. 38:1048–1058. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kontostathi G, Zoidakis J, Makridakis M,
Lygirou V, Mermelekas G, Papadopoulos T, Vougas K, Vlamis-Gardikas
A, Drakakis P, Loutradis D, et al: Cervical cancer cell line
secretome highlights the roles of transforming growth
factor-beta-induced protein ig-h3, peroxiredoxin-2 and nrf2 on
cervical carcinogenesis. Biomed Res Int. 2017:41807032017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Krajka-Kuźniak V, Paluszczak J and
Baer-Dubowska W: The Nrf2-ARE signaling pathway: An update on its
regulation and possible role in cancer prevention and treatment.
Pharmacol Rep. 69:393–402. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lisek K, Walerych D and Del Sal G: Mutant
p53-Nrf2 axis regulates the proteasome machinery in cancer. Mol
Cell Oncol. 4:e12179672016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu K, Alcivar AL, Ma J, Foo TK, Zywea S,
Mahdi A, Huo Y, Kensler TW, Gatza ML and Xia B: NRF2 induction
supporting breast cancer cell survival is enabled by oxidative
stress-induced DPP3-KEAP1 interaction. Cancer Res. 77:2881–2892.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mandal A, Bhatia D and Bishayee A:
Anti-Inflammatory mechanism involved in pomegranate-mediated
prevention of breast cancer: the role of NF-κB and Nrf2 Signaling
pathways. Nutrients. 9:E4362017. View Article : Google Scholar : PubMed/NCBI
|